1. Introduction
In eukaryotic photosynthetic organisms, CO
2 assimilation (fixation) is carried out by a series of biochemical reactions (the Calvin cycle) that take place in chloroplasts [
1]. This biochemical cycle is subdivided into three stages. In stage one, a CO
2 molecule is incorporated into a five-carbon backbone of a ribulose-1,5-bisphosphate that is cleaved into two molecules of phosphoglyceric acid. In stage two, three carbon backbone moieties, created in stage one, are handled and streamlined into other biochemical pathways. In stage three, a pentacarbon substrate is regenerated. In essence, the whole act of CO
2 incorporation is carried out in stage one by the enzyme ribulose-1,5-bisphosphate carboxylase/oxygenase (RUBISCO, E.C. 4.1.1.39). This enzyme is one of the most abundant proteins in our biosphere, and it is mostly responsible for the biomass creation. It catalyzes the carboxylation reaction in which the CO
2 is added to the C2 of ribulose-1,5-bisphosphate. Subsequently, the hexacarbon intermediate is cleaved forming two 3-phosphoglycerates [
2]. These simple sugars can be utilized in several ways including glycolysis (energy production), storage (starch production) or regeneration of the initial substrate ribulose-1,5-bisphosphate. The enzyme can also incorporate O
2 and produce 3-phosphoglycerate and 2-phosphoglycolate. This second reaction is considered parasitic and diminishes the overall rate of CO
2 fixation. This particular species of algae has a high specificity ratio for the CO
2/O
2 incorporation and should provide insights into the molecular origin of specificity and slow catalytic turnover rate.
RUBISCO from higher plants, as well as from cyanobacteria, red algae and some photosynthetic bacteria, forms a hexadecameric structure, built out of eight large and eight small subunits. The molecular weight of a large subunit is around 55kDa, while a small subunit is ~ 16kDa. The three-dimensional structure of RUBISCO’s from several species of plants, as well as from more primitive organisms, have been elucidated by X-ray crystallography. The large subunit containing the active site is folded into the α/β (TIM) barrel, and the small subunit has an α + β fold. The small subunit seems to play mostly a structural role in the assembly of this large molecule, although recent reports have also suggested some catalytic role, especially in the selectivity towards the CO2. The genes for both subunits of G. sulphuraria RUBISCO are coded by the chloroplast DNA. This is in contrast to RUBISCO from plants (e.g. tobacco) where the gene for small subunit is usually coded by the nuclear DNA. Its nuclear location in most species is probably the reason for a higher sequence divergence of the small subunit, especially when compared to that from higher plants.
X-ray crystallography provides a wealth of structural information about the different stages of RUBISCO catalysis. Structures of the unactivated enzyme [
3], the activated one with metal ions and transition state analog [
4], the structures with substrate analogs [
5] as well as products are available in the PDB. A few years ago, the structure of RUBISCO from a similar species
G. partita was solved and published [
6]. However, structural details of activation were not fully described nor the catalytic cycle fully elucidated [
7].
In order for the enzyme to carry out its catalytic mission, it needs to be activated by addition of one molecule of CO
2 to a crucial Lys residue. The active site Lys must be carbamylated to provide an essential ligand for binding of the metal ion. This activation process in plants is mediated by RUBISCO activase [
8] that facilitates the release of blocking agents. It may also happen nonenzymatically, by the prolonged exposure to higher levels of CO
2. In an activated enzyme, the substrate is directly coordinated to the metal ion. It is believed that the second CO
2 molecule is transiently coordinated to the metal ion. In the proposed reaction, the metal ion provides necessary polarization to activate the CO
2 molecule, while the carbamyl moiety plays the role of a base in the reaction. The catalytic mechanism was investigated by a number of biochemical methods such as: isotope exchange, NMR and fast kinetics, as well as by computational methods. The results were summarized by several reviews; one notable that summarizes the kinetic mechanism is by Cleland
et. al.[
7].
The red algae
G. sulphuraria (previously known as
Cyanidium caldarium, Allen strain) appears to be an old organism with a large capacity to adapt to different environments. It can be found in almost every hot spring around the globe and can grow under very diverse conditions that include significantly changed gaseous environments (anaerobic) and varied temperatures (20–50°C). It is an extremophile that prefers acidic conditions (pH ~2) and achieves optimum growth at 42°C, but can dwell at pH=0 and higher temperatures [
9].
The large subunit of RUBISCO from red algae
G. sulphuraria has 493 amino acids while the small subunit has 138 amino acids [
10]. The structure of RUBISCO from this species is interesting since the attempts to reengineer the specificity of RUBISCO towards a more selective enzyme that would suppress or eliminate the oxygenase activity have been unsuccessful [
11]. The red algae may hold great promise in environmental remediation, especially in the era of global warming caused by green-house gasses (e.g. CO
2), because red algae RUBISCO was proven to have the highest specificity toward CO
2[
12].
3. Experimental Section
3.1 Materials
The live cells were gift of Dr. R Troxler (Boston University Medical School). Reagent grade chemicals were purchased from SIGMA.
3.2 Cell Growth
The cells were grown in two stages: a glycolytic-dark stage (rapidly dividing) and a photosynthetic-light stage (a greening stage). They were grown in minimal media containing dextrose (10g/L) enriched with a cocktail of micronutrients (Na, K, Mg, Ca in high μM range, V, Mo, Co, Cu, B in low nM range) at pH 2. The cells after having grown in dark phase for 10 days were spun down and washed with buffer without dextrose. Then the cells were re-suspended in the same medium without the dextrose and exposed to white light for a period of at least 10 days. The cells when harvested were dark green. A small volume of cells collected after dark stage were mixed with 20% glycerol and stored frozen for later use at −80 °C.
3.3 Purification
The cells, collected after the greening stage, were centrifuged at 25,000 × g for 30 min. The cell pellet was re-suspended in a small volume of the breaking buffer (100 mM Tris, 10 mM NaCl, pH 8) and opened by sonication. The cell debris was isolated by centrifugation and the supernatant separated. Subsequently, the soluble proteins were precipitated in 50% saturated solution of ammonium sulfate. The precipitate was re-suspended and dialyzed against pure water for 24 h. The first chromatographic step was carried out on a brushite column (Ca2PO4). The protein was eluted with the gradient of the phosphate buffer (5 mM to 100 mM, pH 8.0). The selected fractions containing colored proteins were precipitated by 50% AS and dialyzed against pure water. The sample applied on the Biogel-A DEAE ion exchange column was eluted with the gradient of 30 mM to 300 mM NaCl (pH 7.5). The colored proteins separated from the main non-colored peak, eluted at approximately 100 mM NaCl. The non-colored peak was confirmed by gel electrophoresis to be RUBISCO. The collected protein fractions encompassing major peaks were combined and dialyzed against 0.01 mM phosphate buffer at pH 7.5 and stored at 4 °C.
3.4 Crystallization
Concentrated protein (~11 mg/mL) was crystallized using the hanging drop vapor diffusion method. The ammonium sulfate variable concentration screen was used. Concentrations ranging from 20% to 60% v/v saturated AS at three pH values (6, 7, 8) were tried. The sample containing 5 μL solution of ~11 mg/mL protein in 0.01 M phosphate buffer mixed with 5 μL of 50 mM phosphate buffer (pH = 7.5) containing 55% v/v AS yielded crystals of the form 1. The crystal of form 1 had a very unusual shape (resembling the optical lens,
Figure 1a) and took longer time to grow (approximately 1 month) than the form 2. The crystals of form 2 were initially obtained by screening with the HAMPTON crystal screen 1. The optimization of crystallization conditions led to the conditions in which the protein suspended in phosphate buffer was mixed with 25% v/v AS and 12% w/v PEG 4000, yielding large crystals for the crystal form 2 (
Figure 1b). The crystals usually grew within 1–2 weeks to typical dimensions of 0.2 × 0.2 × 0.3 mm, in the shape of a square pyramid. The crystals of form 2 were harvested, stabilized in the cryoprotectant containing 50% w/v AS and 20% w/v glycerol. They were suspended in a nylon loop and frozen in N
2 gas stream at 100K for data collection.
3.5 Data Collection
The initial crystal characterization and data collection on the crystals of form 1 was carried out on an old multiwire area detector MarcII (Multiwire Area Detector Systems, Poway, CA) at room temperature. The first data set was used for structure solution. The second data set was collected later at 100 K on the Rigaku RaxisIV++ area detector system mounted on the MicroMax 007 rotating anode generator operated at 40 kV and 20 mA and processed with HKL2000 [
21]. The system is located at the Chemistry Department, University of Texas at El Paso. The data clearly showed high symmetry space group I422 with the diffraction extending to ~2 Å resolution.
The images for the second crystal form showed diffraction to approximately ~2.4 Å resolution (
Table 1). The crystal system for the form 2 was monoclinic with the space group P2
1, as determined by the analysis of systematic absences. The X-ray data for both crystal phases of form 2 were collected at 100 K. The diffraction data for the form 2 were collected on the same equipment, as described for form 1. The data were processed with HKL2000.
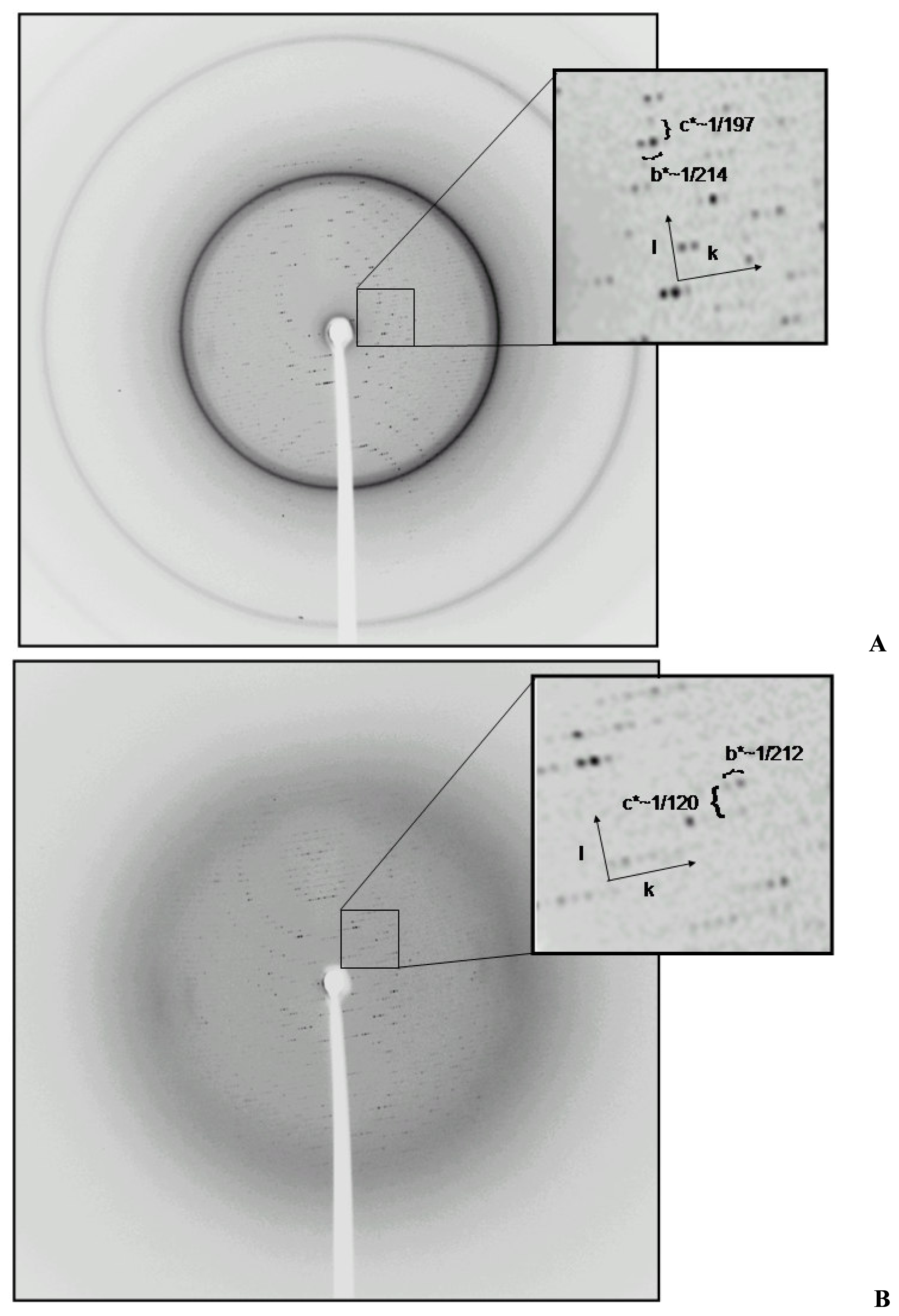
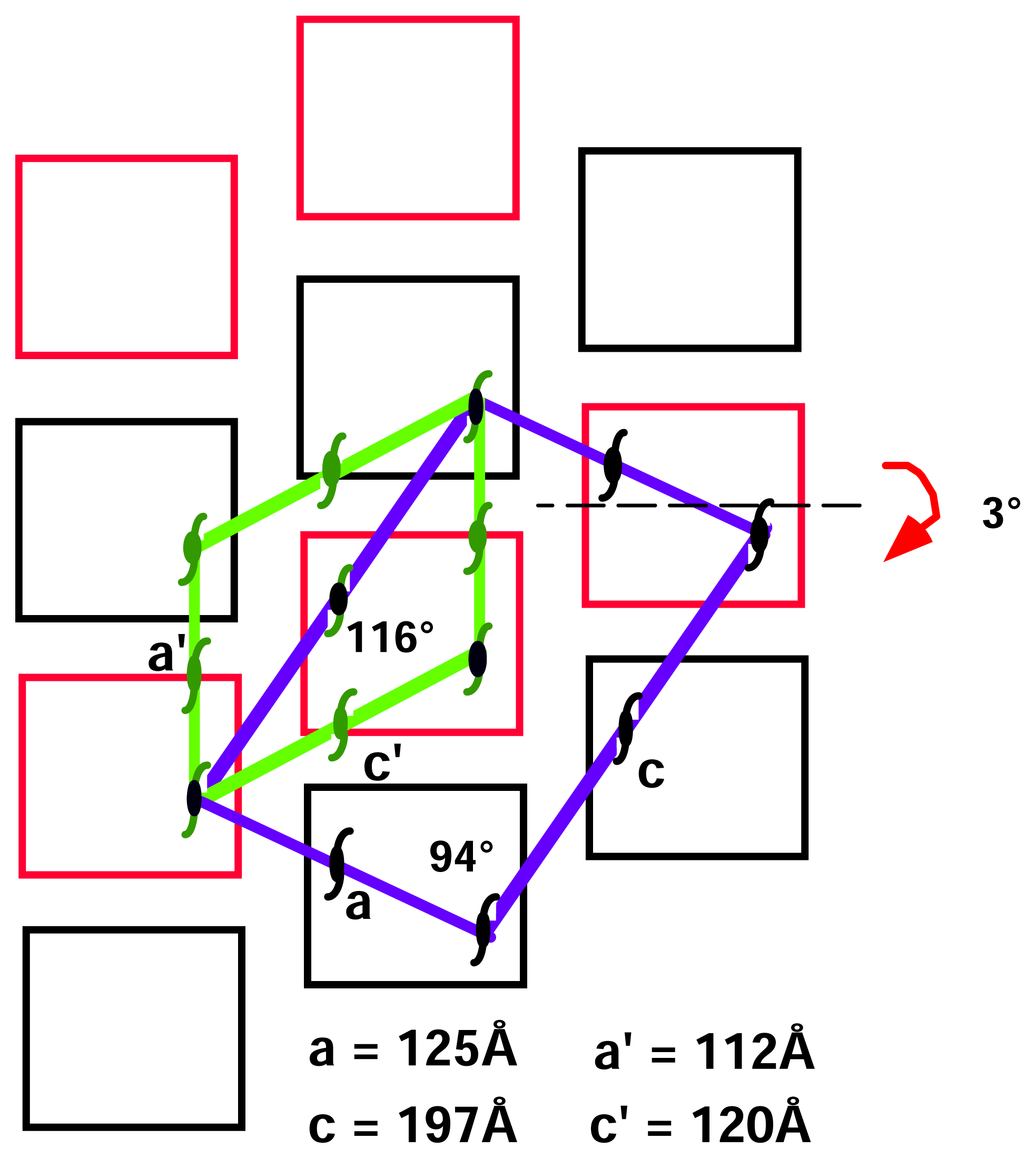
After the first successful data set collection and data reduction for the monoclinic form, we decided to eliminate the background coming from imperfect cryoprotection. We thawed the crystal, bathed it in fresh stabilizing solution and froze again in the cold stream. To our surprise, we saw a strong yet different diffraction pattern (
Figure 2). This new pattern suggested an internal change in the crystal packing. We collected a second data set using the same crystal. The data reduction showed a change in cell dimensions with preservation of monoclinic system (
Table 1).
3.6 Molecular replacement
The phase problem for both crystal forms was solved using molecular replacement. The structure for the first crystal form was solved by molecular replacement as implemented in CNS [
22] using spinach RUBISCO as a template (1AUS). The cell dimensions as well as the symmetry suggested a single large and a single small subunit in the asymmetric unit. The initial solution was unique, taking into account the high symmetry of the lattice with the signal to noise ratio for rotation function of 15.8 sigma remaining peaks around 3 sigma (R factor ~0.5). The translation solution was unique and fixed by the internal symmetry of the assembly. The initial placement was followed by a long process of establishing the correct sequence. Only in the last two years with advances in sequencing technologies the unique sequence for
Galdieria sulphuraria RUBISCO emerged. The initial fit with
Cyanidium caldarium sequence (~86% identity, large subunit and 63% identity, small subunit) showed many mismatches that turned out to be sequence changes between three closely related species of
Galdieria partita,
Cyanidium caldarium and
Galdieria sulphuraria. When the search was conducted with the
G. partita model, the rigid body refinement lowered the R factor to 0.39 thus confirming the solution obtained before.
The phase problem for the crystal form 2a was solved by MR with
G. partita model as a template (1IWA) using the CNS [
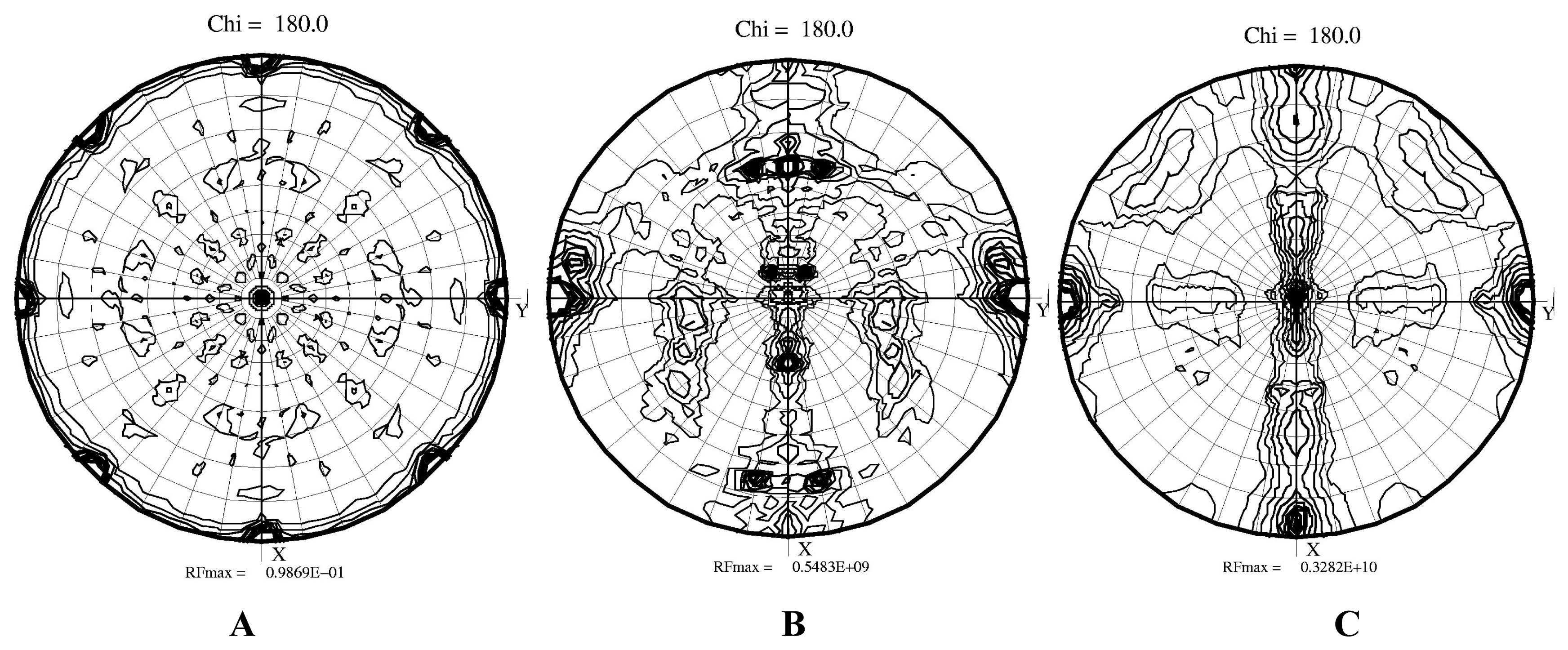
22]. The cell dimensions suggested that two hexadecameric molecules were present in the asymmetric unit. The self-rotation function suggested that the fourfold axis of the hexadecamers were close to the Y axis while both hexadecamers were rotated against each other (
Figure 3). Indeed, two hexadecamers were located in the asymmetric unit with the resulting R factor of 0.49. The initial rotation function produced eight peaks approximately 30 sigma above the noise level, while the next peaks on the list were at the level of 5 sigma. All those peaks were equivalent by non-crystallographic 422 symmetry. We are presently continuing the refinement of the initial model because the ~10,000 amino acid contents of the asymmetric unit makes it a formidable technical task. We are working on implementing effective refinement strategies.
A similar process to solve the phase problem was carried out for the second monoclinic form using the same model of G. partita RUBISCO. As suggested by the cell dimensions, only a single hexadecamer was located in the asymmetric unit. The highest peaks were again above 30 sigma and R ~0.45, indicating the correctness of the solution. The self-rotation function clearly showed that a major fourfold axis was positioned as in form 2a along the Y direction while the perpendicular two-fold symmetry axis was aligned with the X direction. The simplification of the self-rotation function combined with detected directions of two-fold axes when compared to form 2a was consistent with the presence of a single hexadecamer in the asymmetric unit.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}