Cell Adhesion Regulates Expression of the Androgen Receptor and Coregulators in Different Prostate Cancer Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
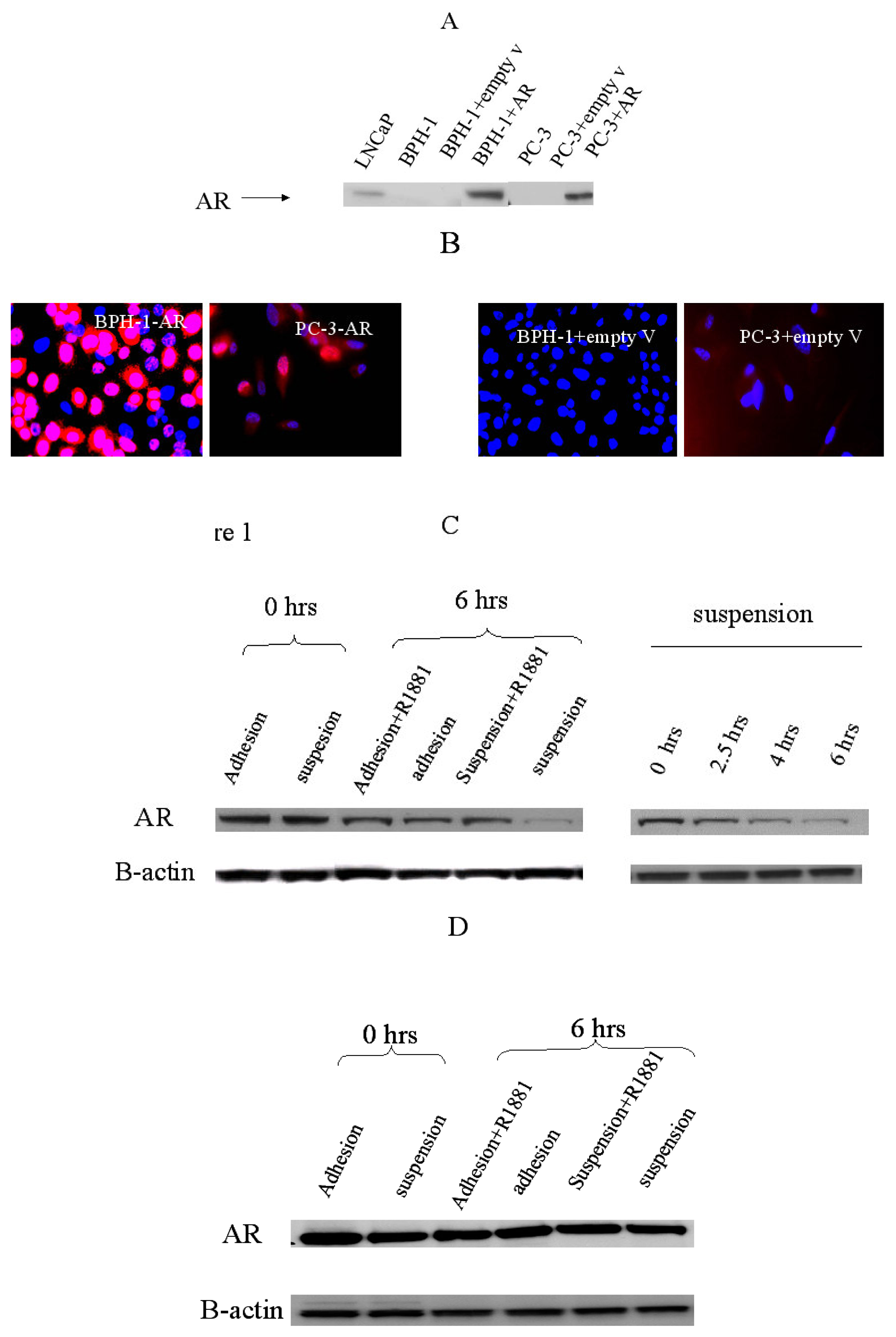
2.1. Establishment of the cell line model to study cell-matrix adhesion
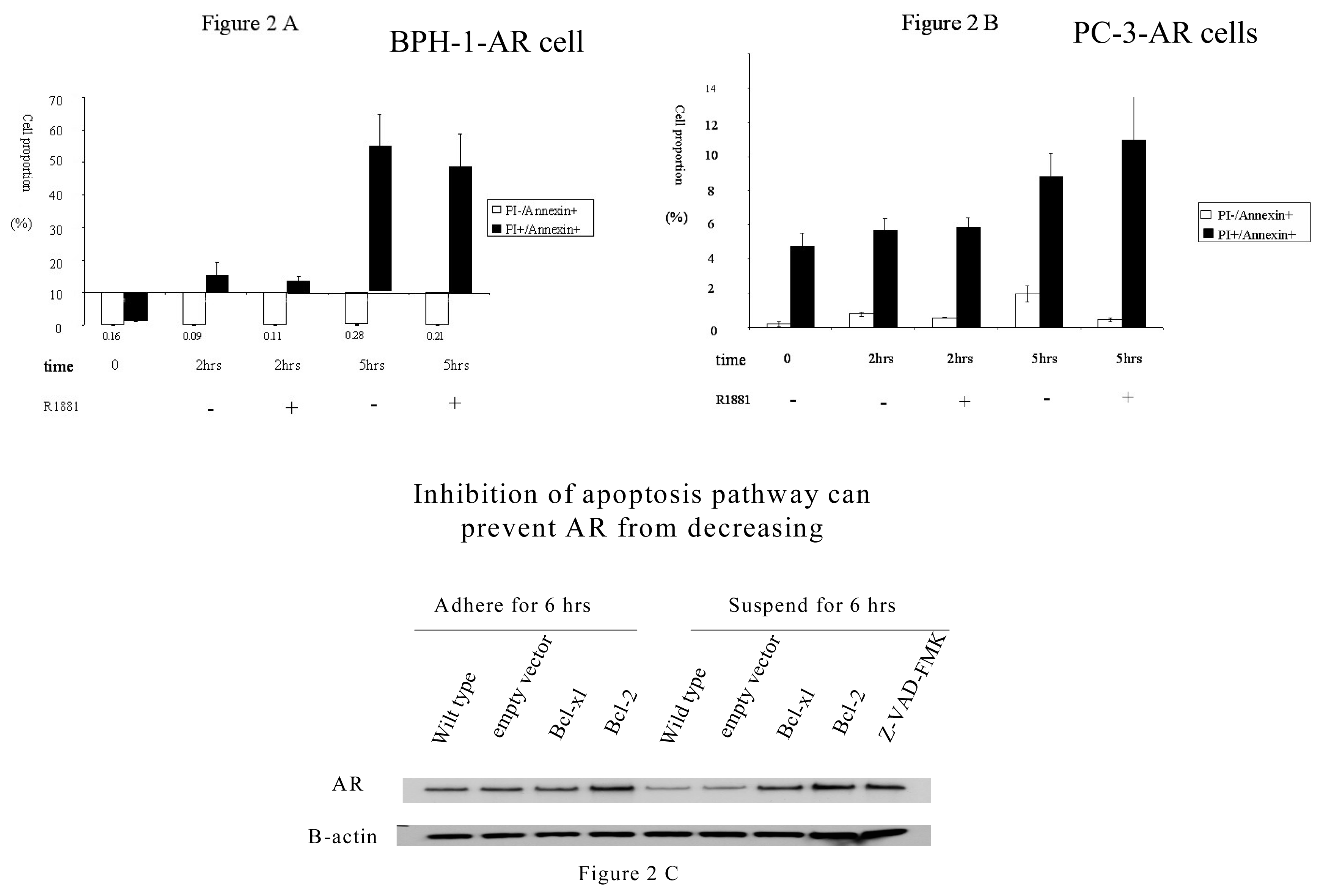
2.2 Adhesion effect on AR expression in prostate cancer cells
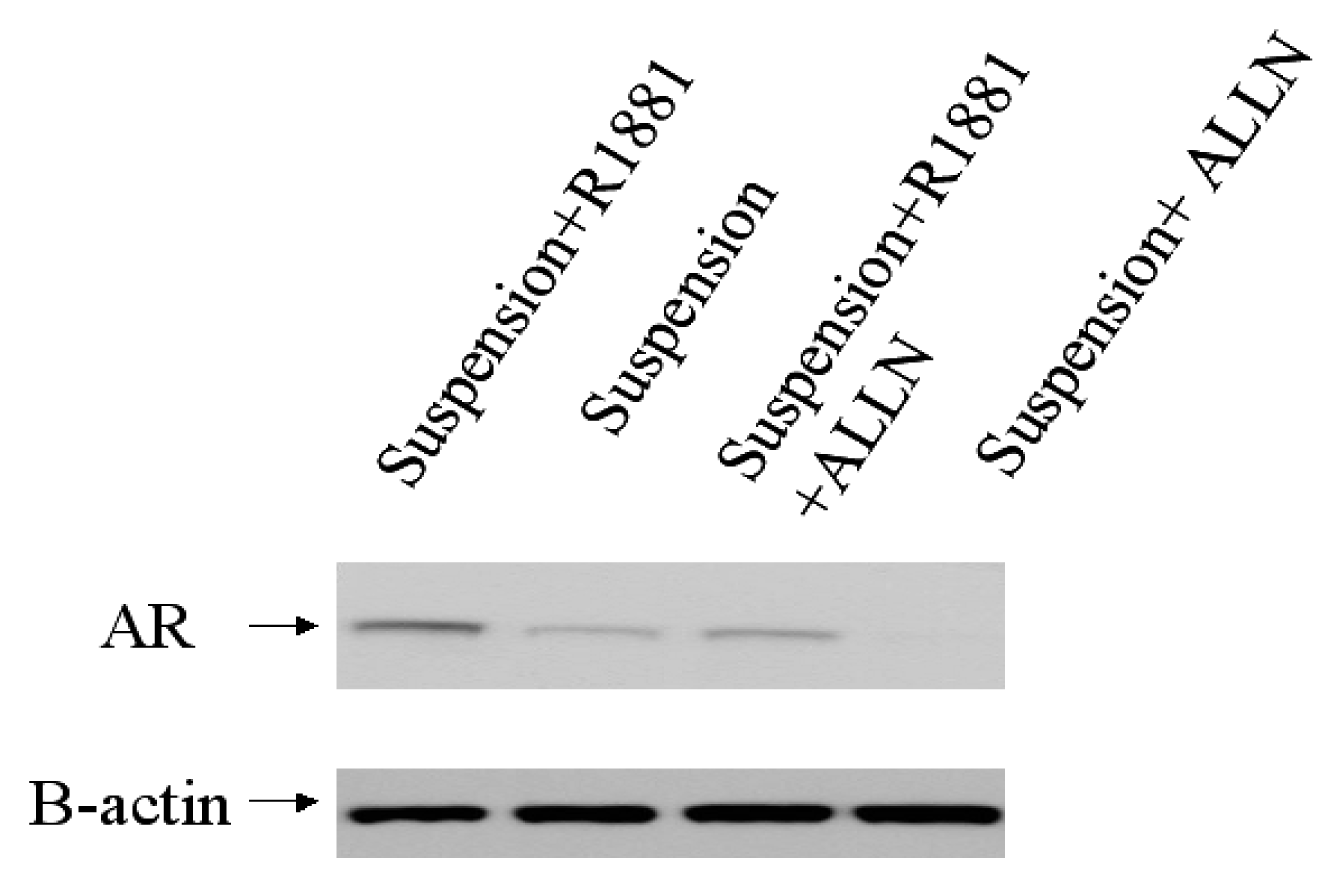
2.3. The mechanism involved in the regulation of AR by adhesion/suspension
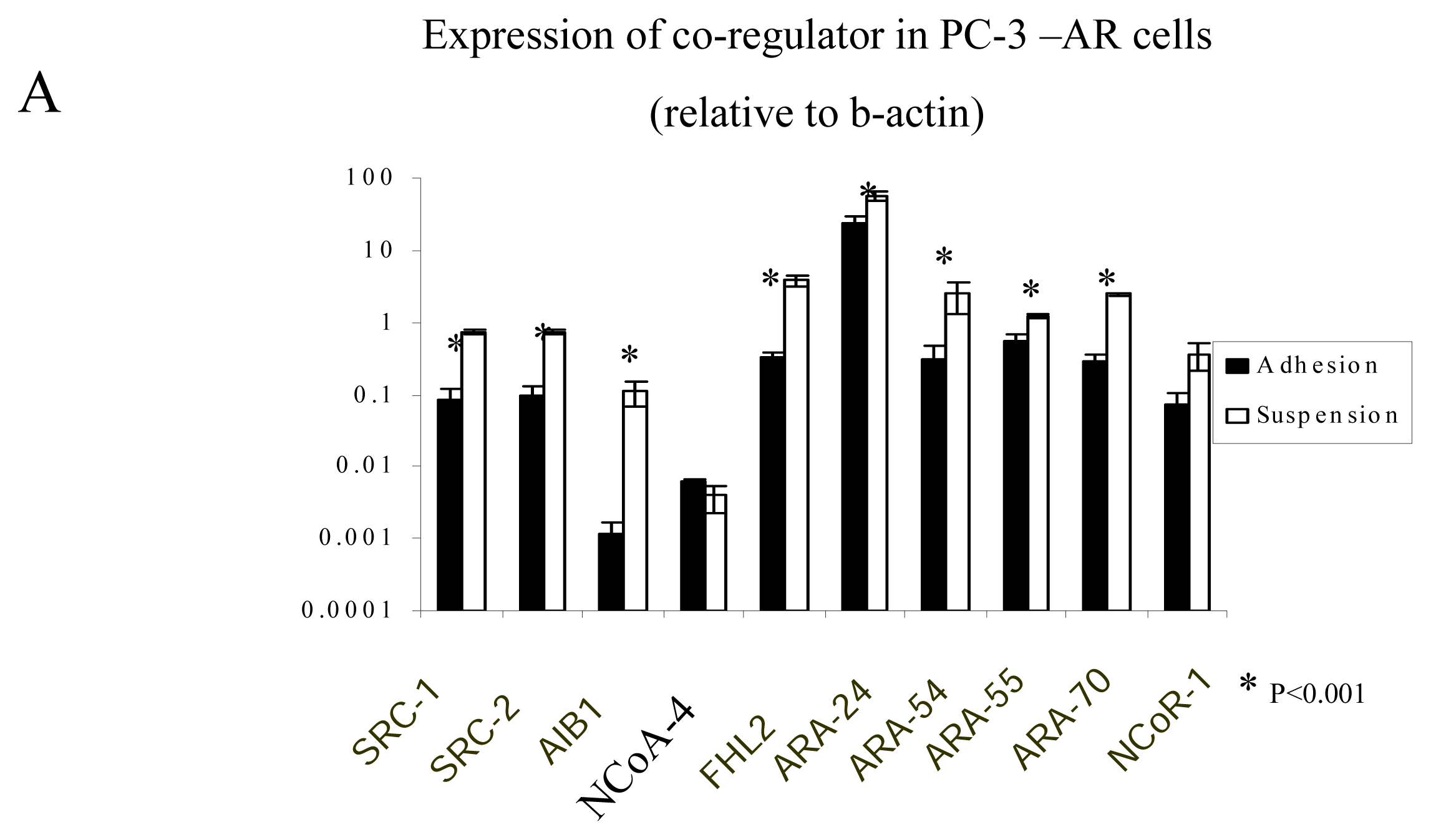
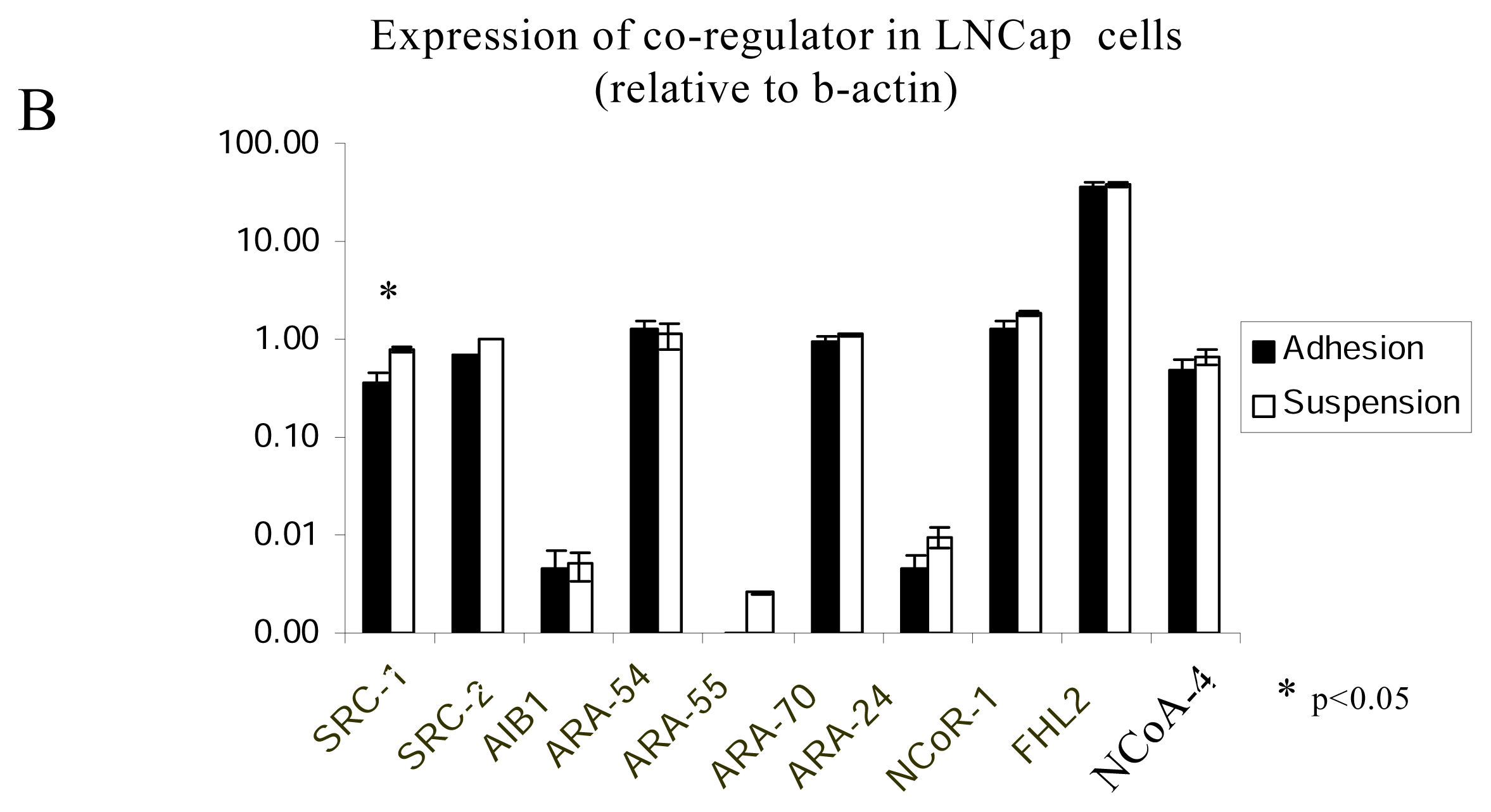
2.4. The regulation of AR-coregulators in adherent and suspended prostate cancer cell lines
3. Experimental
Cell cuture
Cell lysis and western blot
Immunofluorescent analysis
AR measurement in adherent and suspended cells
Apoptosis assay by double staining with anexin V-FITC and propidium iodide( PI)
Real-time PCR to measure AR-coregulators in adherent and suspended prostate cancer cell lines
Statistics
Acknowledgements
References
- Mattila, E.; Pellinen, T.; Nevo, J.; Vuoriluoto, K.; Arjonen, A.; Ivaska, J. Integrins and EGFR coordinately regulate the pro-apoptotic protein Bim to prevent anoikis. Nat. Cell. Biol 2003, 5, 733–740. [Google Scholar]
- Burridge, K.; Wennerberg, K. Rho and Rac Take Center Stage. Cell 1992, 70, 389–399. [Google Scholar]
- Knudsen, B.S.; Miranti, C.K. The impact of cell adhesion changes on proliferation and survival during prostate cancer development and progression. J. Cell Biochem 2006, 99, 345–361. [Google Scholar]
- Yu, H.M.; Frank, D.E.; Zhang, J.; You, X.; Carter, W.G.; Knudsen, B.S. Basal prostate epithelial cells stimulate the migration of prostate cancer cells. Mol. Carcinog 2004, 41, 85–97. [Google Scholar]
- Heinlein, C.A.; Chang, C. Androgen receptor in prostate cancer. Endocr. Rev 2004, 25, 276–308. [Google Scholar]
- Cress, A.E.; Rabinovitz, I.; Zhu, W.; Nagle, R.B. The alpha 6 beta 1 and alpha 6 beta 4 integrins in human prostate cancer progression. Cancer Metastasis Rev 1995, 14, 219–228. [Google Scholar]
- Rahman, M.; Miyamoto, H.; Chang, C. Expression of Androgen Receptor Coregulators in Prostate Cancer. Clin. Cancer Res 2004, 10, 2208–2219. [Google Scholar]
- Nagle, RB. Role of the Extracellular Matrix in Prostate Carcinogenesis. Journal of Cellular Biochemistry 2004, 91, 36–40. [Google Scholar]
- Gelmann, E.P. Molecular Biology of the Androgen Receptor. J. Clin. Oncol 2002, 20, 3001–3015. [Google Scholar]
- Yang, L.; Xie, S.; Jamaluddin, M.S.; Altuwaijri, S.; Ni, J.; Kim, E.; Chen, Y.T.; Hu, Y.C.; Wang, L.; Chuang, KH.; Wu, C.T.; Chang, C. Induction of androgen receptor expression by phosphatidylinositol 3-kinase/Akt downstream substrate, FOXO3a, and their roles in apoptosis of LNCaP prostate cancer cells. J. Biol. Chem 2005, 280, 33558–33565. [Google Scholar]
- Krongrad, A.; Wilson, C.M.; Wilson, J.D.; Allman, D.R.; McPhaul, M.J. Androgen increases androgen receptor protein while decreasing receptor mRNA in LNCaP cells. Mol. Cell. Endocrinol 1991, 76, 79–88. [Google Scholar]
- Zhou, Z.X.; Lane, M.V.; Kemppainen, J.A.; French, F.S.; Wilson, E.M. Specificity of ligand-dependent androgen receptor stabilization: receptor domain interactions influence ligand dissociation and receptor stability. Mol. Endocrinol 1995, 9, 208–218. [Google Scholar]
- Sheflin, L.; Keegan, B.; Zhang, W.; Spaulding, S.W. Inhibiting proteasomes in human HepG2 and LNCaP cells increases endogenous androgen receptor levels. Biochem. Biophys. Res. Commun 2000, 276, 144–150. [Google Scholar]
- Lin, H.K.; Wang, L.; Hu, Y.C.; Altuwaijri, S.; Chang, C. Phosphorylation-dependent ubiquitylation and degradation of androgen receptor by Akt require Mdm2 E3 ligase. EMBO J 2002, 21, 4037–4048. [Google Scholar]
- Lin, H.K.; Altuwaijri, S.; Lin, W.J.; Kan, P.Y.; Collins, L.L.; Chang, C. Proteasome activity is required for androgen receptor transcriptional activity via regulation of androgen receptor nuclear translocation and interaction with coregulators in prostate cancer cells. J. Biol. Chem 2002, 277, 36570–36576. [Google Scholar]
- Brzozowski, A.M.; Pike, A.C.; Dauter, Z.; Hubbard, R.E.; Bonn, T.; Engelstrom, O.; Ohman, L.; Greene, G.L.; Gustafsson, J-A.; Carlquist, M. Molecular basis of agonism and antagonism in the oestrogen receptor. Nature 1997, 389, 753–758. [Google Scholar]
- Hsu, C.L.; Chen, Y.L.; Ting, H.J.; Lin, W.J.; Yang, Z.; Zhang, Y.; Wang, L.; Wu, C.T.; Chang, H.C.; Yeh, S.; Pimplikar, S.W.; Chang, C. Androgen receptor (AR) NH2- and COOH-terminal interactions result in the differential influences on the AR-mediated transactivation and cell growth. Mol. Endocrinol 2005, 19, 350–361. [Google Scholar]
- Manin, M.; Baron, S.; Goossens, K.; Beaudoin, C.; Jean, C.; Veyssiere, G.; Verhoeven, G.; Morel, L. Androgen receptor expression is regulated by the phosphoinositide 3-kinase/Akt pathway in normal and tumoral epithelial cells. Biochem. J 2002, 366, 729–736. [Google Scholar]
- Sharma, M.; Chuang, W.W.; Sun, Z. Phosphatidylinositol 3-kinase/Akt stimulates androgen pathway through GSK3beta inhibition and nuclear beta-catenin accumulation. J. Biol. Chem 2002, 30935–30941. [Google Scholar]
- Yang, L.; Xie, S.; Jamaluddin, M.S.; Altuwaijri, S.; Ni, J.; Kim, E.; Chen, Y.T.; Hu, Y.C.; Wang, L.; Chuang, K.H.; Wu, C.T. Chang CInduction of androgen receptor expression by phosphatidylinositol 3-kinase/Akt downstream substrate, FOXO3a, and their roles in apoptosis of LNCaP prostate cancer cells. J Biol Chem 2005, 280, 33558–33565. [Google Scholar]
- Ellerby, L.M.; Hackam, A.S.; Propp, S.S.; Ellerby, H.M.; Rabizadeh, S.; Cashman, N.R.; Trifiro, M.A.; Pinsky, L.; Wellington, C.L.; Salvesen, G.S.; Hayden, M.R.; Bredesen, D.E. Kennedy’s disease: caspase cleavage of the androgen receptor is a crucial event in cytotoxicity. J Neurochem 1999, 72, 185–195. [Google Scholar]
- Rokhlin, O.W.; Taghiyev, A.F.; Guseva, N.V.; Glover, RA.; Chumakov, P.M.; Kravchenko, J.E.; Cohen, M.B. Androgen regulates apoptosis induced by TNFR family ligands via multiple signaling pathways in LNCaP. Oncogene 2005, 24, 6773–6784. [Google Scholar]
- Liao, X.; Tang, S.; Thrasher, J.B.; Griebling, T.L.; Li, B. Small-interfering RNA-induced androgen receptor silencing leads to apoptotic cell death in prostate cancer. Mol Cancer Ther 2005, 4, 505–515. [Google Scholar]
- Nessler-Menardi, C; Jotova, I.; Culig, Z.; Eder, I.E.; Putz, T.; Bartsch, G.; Klocker, H. Expression of Androgen Receptor Coregulatory Proteins in Prostate Cancer and Stromal-Cell Culture Models. Prostate 2000, 45, 124–131. [Google Scholar]
- Kinoshita, M.; Nakagawa, T.; Shimizu, A.; Katsuoka, Y. Expression of Androgen Receptor Coregulators in Prostate Cancer. Clin Cancer Res 2004, 10, 1032–1040. [Google Scholar]





© 2007 by MDPI Reproduction is permitted for noncommercial purposes.
Share and Cite
Zhang, J.; Xiang, J.-Q.; Zhang, Y.-W.; Chen, Y.-Y.; Zhou, X.-Y.; Liu, Y.; Li, S. Cell Adhesion Regulates Expression of the Androgen Receptor and Coregulators in Different Prostate Cancer Cells. Int. J. Mol. Sci. 2007, 8, 156-165. https://doi.org/10.3390/i8020156
Zhang J, Xiang J-Q, Zhang Y-W, Chen Y-Y, Zhou X-Y, Liu Y, Li S. Cell Adhesion Regulates Expression of the Androgen Receptor and Coregulators in Different Prostate Cancer Cells. International Journal of Molecular Sciences. 2007; 8(2):156-165. https://doi.org/10.3390/i8020156
Chicago/Turabian StyleZhang, Jie, Jia-Qing Xiang, Ya-Wei Zhang, Yan-Yan Chen, Xiao-Yan Zhou, Yun Liu, and Sheng Li. 2007. "Cell Adhesion Regulates Expression of the Androgen Receptor and Coregulators in Different Prostate Cancer Cells" International Journal of Molecular Sciences 8, no. 2: 156-165. https://doi.org/10.3390/i8020156