1. Introduction
Csx/Nkx2.5 is an evolutionarily conserved homeodomain-containg transcription factor that is essential for early cardiac development. It was first isolated on the basis of its homology to the Drosophila gene tinman [
1]. The expression of Csx/Nkx2.5 is restricted to the heart and heart progenitor cells from the very early developmental stage when the two heart primordial are symmetrically situated in the anterior lateral mesoderm [
2]. Targeted disruption of murine Csx/Nkx2.5 resulted in embryo ionic lethality due to the abnormal looping morphogenesis of the primary heart tube, indicating that Csx/Nkx2.5 plays a critical role in cardiac morphogenesis [
3].The transcription factors of NK2 class contain highly conserved regions: a 180-bp sequence motif that encodes a helix-turn-helix DNA binding domain designated homeodomain (HD). Two other peptide domains conserved among most members are TN-Domain near the amino terminus and NK2-Specific Domain (NK2-SD), located carboxyl-terminal to the HD. Csx/Nkx2.5 belongs to the NK2 class of homebox proteins characterized by a tyrosine residue at amino acid 54 of the homeodomain (HD) and a conserved 23-amino acid NK2-SD [
4]. Molecular dissection of Csx/Nkx2.5 has revealed that, in addition to DNA binding, the HD plays critical roles in transcriptional regulation by nuclear translocation of Csx/Nkx2.5 protein [
5] and protein–protein interaction with other transcription factors [
6–
9]. Up to date, more than 26 mutations in Csx/Nkx2.5 have been reported to be associated with human congenital heart diseases (CHD). The most common phenotypes were progressive atrioventricular conduction delays (AV block) and secundum atrial septal defect (ASD).
In the majority of patients with familial ASD, Csx/Nkx2.5 mutations are situated within or around the HD and associated with AV block [
10–
12], however, recent reports have shown that most of the mutations identified in sporadic CHD were missense situated outside the HD and not associated with AV conduction disturbance [
13]. Since nearly all researches of Csx/Nkx2.5 have focused on the role of the HD, relatively little is known about the functions of TN-Domain and NK2-SD. So it becomes more interesting and important to determine the genotype–phenotype correlations in patients with Csx/Nkx2.5 outside the HD.
To better define the spectrum of biochemical properties modified by Csx/Nkx2.5 mutations, here we report the initial biochemical characterization of a series of Nkx2.5 mutants with individual domain truncation. Our study represents the comprehensive analysis of Csx/Nkx2.5 mutants and provides the structure–function relations among TN-Domain, NK2-SD and HD for further definition of molecular mechanisms of Csx/Nkx2.5 mutant protein dysfunction.
2. Materials and Methods
2.1 Construction of Plasmids
Human Csx/Nkx2.5 cDNA was amplified by RT-PCR and then subcloned into mammalian expression vector pCI-HA to be prepared as HA fusion protein. To construct a series of mutants, we replaced the wild type fragment of pCI-HA-Csx/Nkx2.5 with designed fragments generated by PCR, Their primers were listed as follows: M1(32-324)(F: 5′-cgc gga tcc atg ttc ccc agc cct gct -3′; R:5′- cgc ctc gag cta cca ggc tcg gat acc-3′), M2(1-201)( F: 5′- cgc gga tcc atg ttc ccc agc cct gct-3′;R: 5′- cgc ctc gag tgg agc tgg tgg ggc tgc), M3(43-236)( F: 5′-cgc gga tcc tcc tcc tgc atg ctg gcc-3′; R: 5′-cgc ctc gag cgt gag ttt cag cac gct-3′). M4(Δ138-197)(F1; 5′-cgc gga tcc atg ttc ccc agc cct gct-3′, F2: 5′- cgc gct gcc gcc gtc ctg gtc tga-3′; R1: 5′-gcg cga cgg cgg cag gac cag act, R2: 5′- cgc ctc gag cta cca ggc tcg gat acc-3′). HindIII-EcoRI fragment encoding either wild type Csx/Nkx2.5 or each of mutants were subcloned into His-fusion plasmid pET32a (Novagen, USA) to generated pET32a-His6-Csx (or mutant) expression plasmid. The ANF-luciferase reporter plasmid is a gift from Dr. Hiroki Aoki (Japan).
2.2 Cell Culture, Transfection and Luciferase Reporter Assays
COS-7 cells were cultured in Dulbecco’s modified Eagle’s medium supplemented with 10% newborn calf serum. Transient transfections were performed by the standard calcium phosphate method 24 h after plating. Cells seeded on six-well plates were co-transfected with 0.5 μg of ANF-luc, 0.5 μg β-galactosidase plasmid (pON405) and 2 μg of empty pCI-HA or pCI-HA plasmids encoding wild-type or mutant Csx/Nkx2.5, and then cultured for another 48 h. Lysed cells with 200 μl of reporter lysis buffer (Promega), and assayed for luciferase activity and β-galactosidase activity. Luciferase activity was normalized against β-galactosidase activity. To examine the effects of each mutant on transcriptional activation of wild-type (WT) Csx/Nkx2.5, the plasmid amount of wild type and each mutant was adjusted at the ratio of 1:1 and then cotransfected together with the plamids listed above.
2.3 Nuclear Localization
Nuclear extracts of COS-7 cells transfected with pCI-HA-Csx (wild type or mutant) were prepared as follows: cells were washed with cold PBS buffer and soaked in 500 μl of low salt buffer (20 mM HEPES, pH 7.6, 1 mM DTT, 0.1% Triton X-100, 0.5 mM PMSF, 10 mM KCl) for 15 min on ice. Nuclei pelleted by centrifugation at 800 g for 10 min were resuspended in 100 μl of extraction buffer (20 mM HEPES, pH 7.6, 450 mM NaCl, 0.5 mM EDTA, 1 mM DTT, 25% glycerol, 0.1 mM PMSF, 1.5 mM MgCl2) and incubated for 30 min at 4°C. After centrifugation at 12,000 g for 10min, the supernatant was used as a nuclear extract and analyzed by western blot referred to the standard protocol. Antibodies against Lamin B (Zymed, USA) and Tubulin (Santa Cruz, USA) were used for validation of the cytosolic and nuclear fractions, respectively.
2.4 Immunofluorescence Staining
COS7 cells transfected with pCI-HA-Csx (wild type or mutant) using lipofectamine 2000 (Invitrogen, USA) were stained with anti-HA mAb (Santa Cruz, USA) followed by CFSE-conjugated anti-mouse IgG (Sigma, USA) and DAPI dye (Sigma, USA) as described [
14]. The fixed cells were mounted and analyzed by using Zeiss laser scanning microscope with Axiovision 3.1 software.
2.5 Electrophoretic Mobility Shift Assay (EMSA)
The purification of His6 fusion proteins with Ni-NTA-agrose (Qiagen, USA) was estimated by Coomassie-stained SDS-PAGE gel. Two pM of oligonucleotide, 5′-TCACACCTT
TGAAGTGGGGGC
CTCTTGAGGCAAATC-3′, was annealed with 5′-GATTTGC
CTCAAGAGGCCCC
CACTTCAAAGGTGTGA-3′ and labeled with [γ-
32P]dATP. The 36bp double-stranded oligonucleotide contains Csx/Nkx2.5 binding sites (underlined) from the atrial natriuretic factor (ANF) promoter and is used to examine DNA binding of Csx/Nkx2.5 or mutants [
15]. Labeled probes were incubated with wild type or mutant protein purified from E. coli as His6 fusion protein (0–33.4 × 10
−9M), 1 μg of poly(dI-dC) in 20 μl of binding buffer (50 mg of bovine serum albumin, 0.5 mg of poly(dG-dC) in 10 mM HEPES, pH 8.0, 50 mM KCl, 1 mM EGTA, 10% glycerol, 2.5 mM DTT, 7mM MgCl2) for 30 min at room temperature. The protein/DNA mixture was separated on 4% native polyacrylamide gel in 0.5× Tris borate/EDTA buffer for 2 h at 150 V. Protein-DNA binding affinity (Kd) was estimated by the protein concentration at which 50% of the DNA probe became bound [
16].
3. Results
3.1 Expression and purification of wt Csx/Nkx2.5 and its mutants
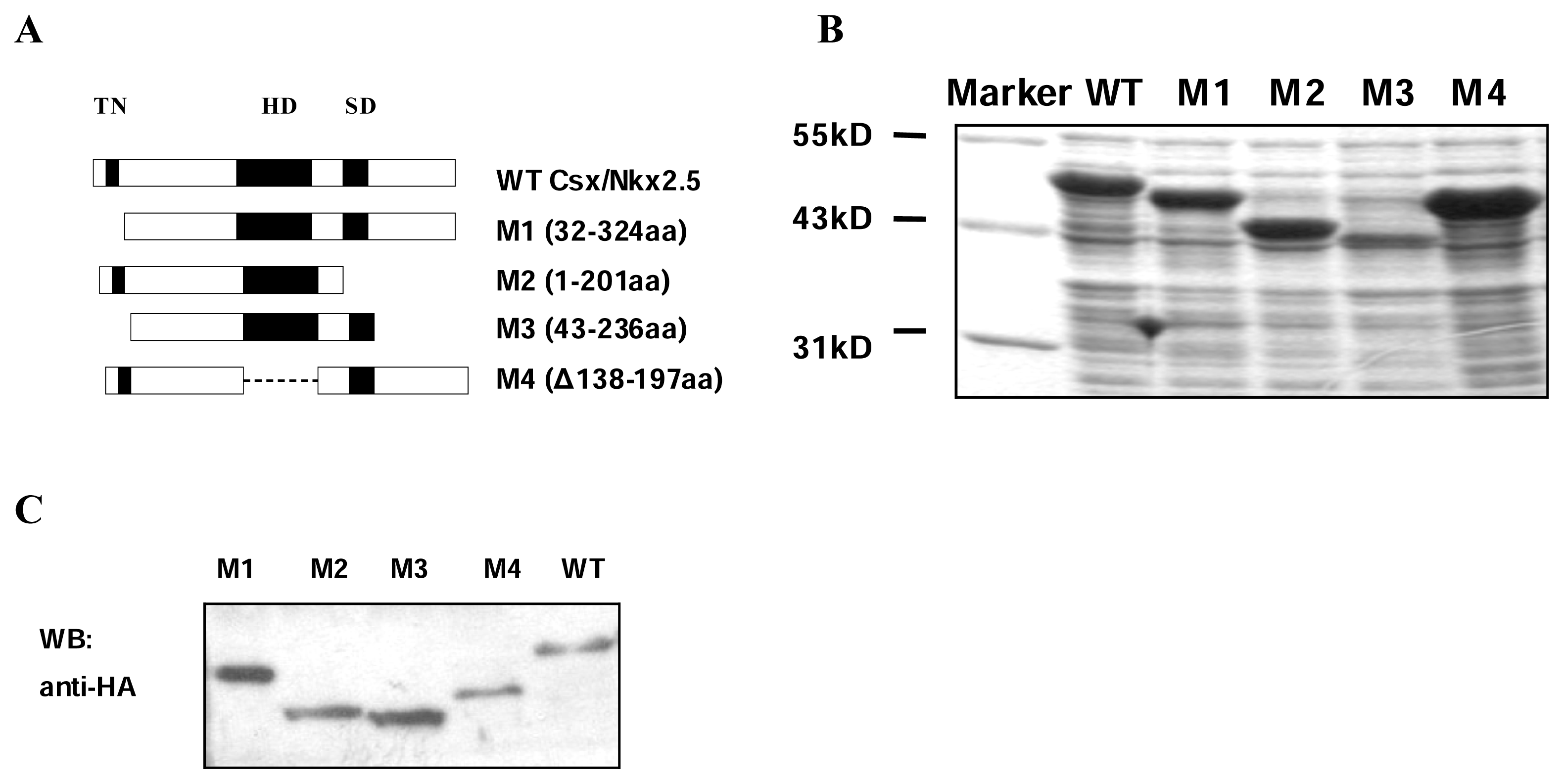
Human Csx/Nkx2.5 is a 324-amino protein, including three domains: TN, HD and NK2-SD. To gain insights into the function of TN and NK2-SD domain and their relation to HD domain, expression vectors for wild-type(WT) Csx/Nkx2.5 and a series of deletion mutants were constructed: M1(32-324) lacking TN domain, M2(1-201) missing NK2-SD domain and carboxyl terminal tail, M3(43-236) lacking both N-terminal and C-terminal domain, and M4(Δ138-197) missing HD domain of Csx/Nkx2.5 (
Figure 1A). We expressed each protein by either fused with His6-tag in E. coli BL21(DE3) or fused with HA-tag in COS-7 cells. The expression levels of all the fusion proteins in bacteria were analyzed by Coomassie blue-stained SDS-PAGE, the molecular weight of each fusion protein was estimated according to the protein markers (
Figure 1B). Western blot analysis using whole cell lysates revealed that all mutants as well as WT were expressed in COS-7 cells at comparable levels (
Figure 1C).
3.2 Nuclear transport of Csx/Nkx2.5 mutants
Csx/Nkx2.5 is a nuclear phosphoprotein whose NLS requires the amino-terminal basic residues of the HD [
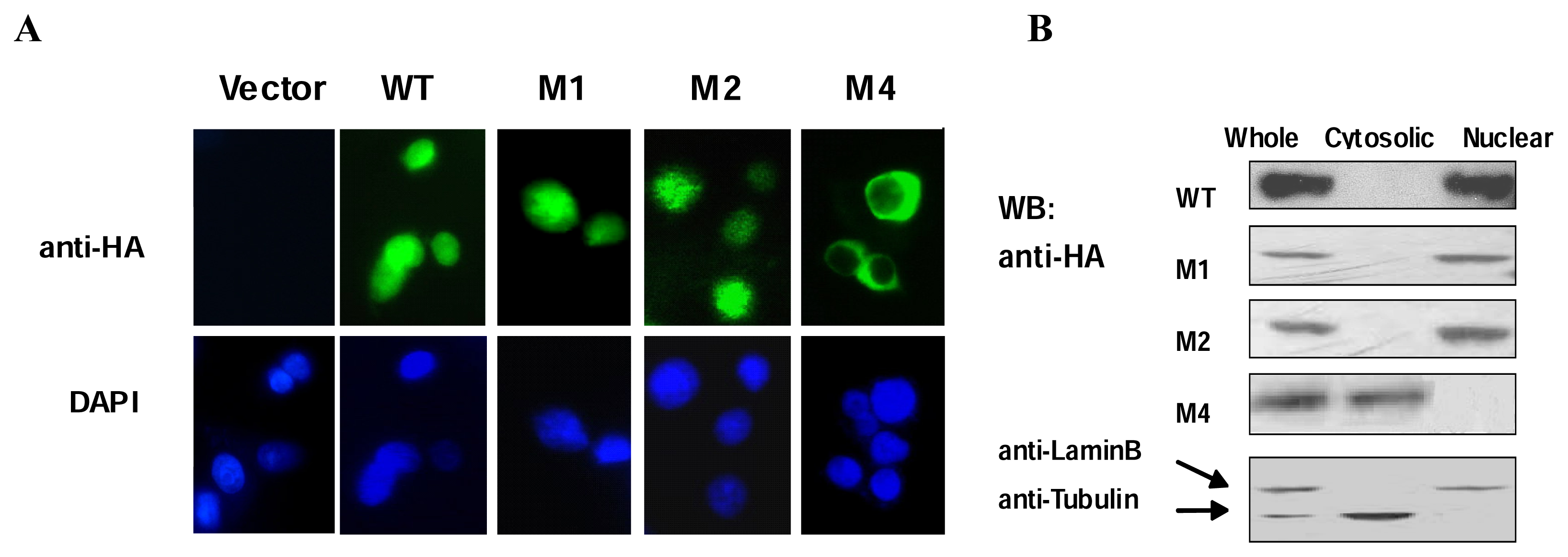
5]. To investigate whether or not the domains outside the HD of Csx/Nkx2.5 are necessary for nuclear transport, the mutant constructs were transfected into COS-7 cells and their subcellular distributions were visualized by immunofluorescent staining (
Figure 2A). Only the M4 mutant lacking HD showed cytoplasmic localization, in contrast, WT and other mutant proteins exhibited a strong nuclear localization. These results were further confirmed by immunochemical analysis of the nuclear and cytosolic fractions of COS-7 cells over-expressing WT Csx/Nkx2.5 or mutants respectively (
Figure 2B). Consistent with previous reports, the localization of Csx/Nkx2.5 is dependent on the HD. Both TN and NK2-SD deletion mutants we constructed showed no significant effect on the nuclear transport.
3.3 DNA binding affinity of Csx/Nkx2.5 mutants
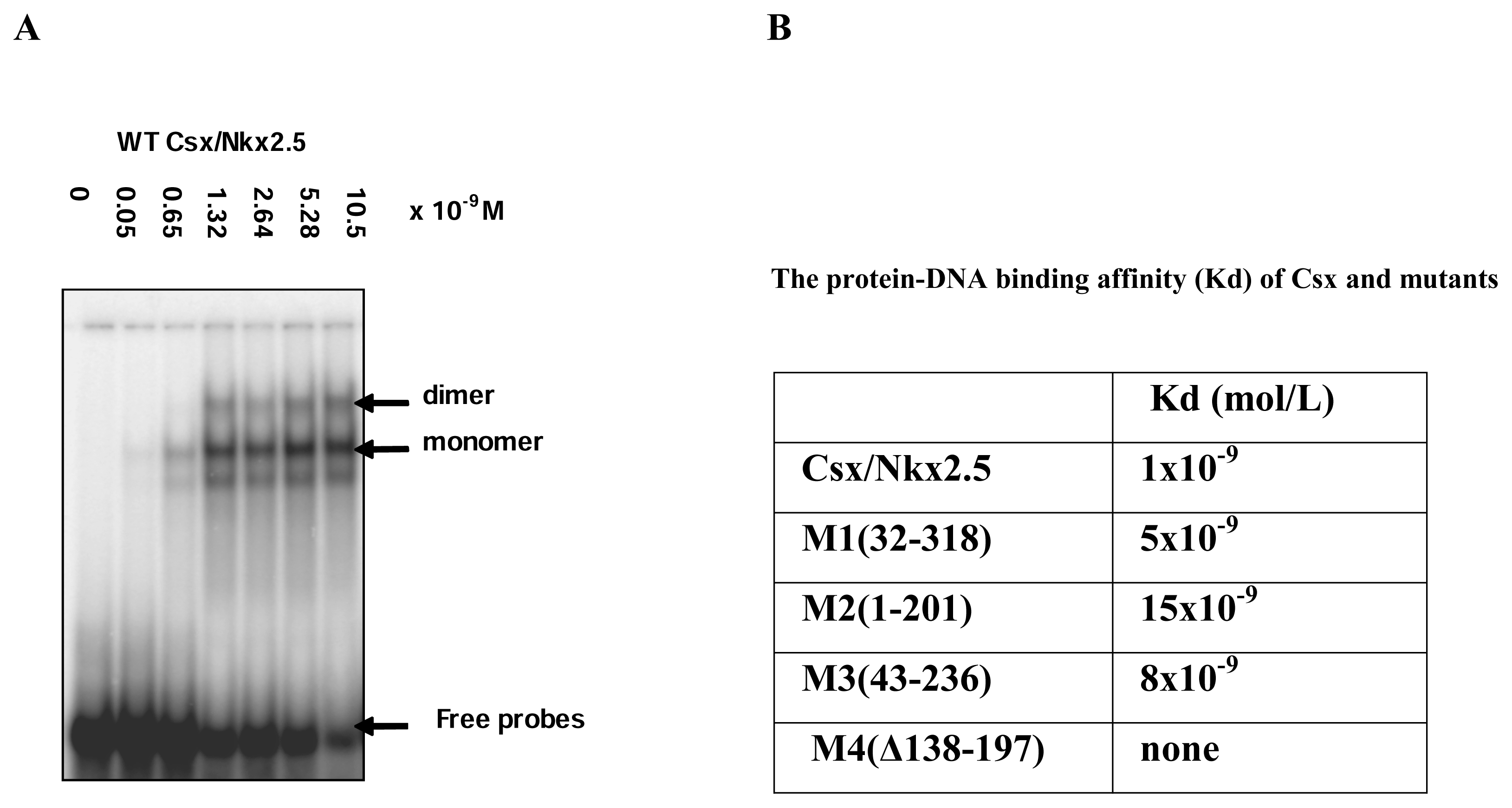
A very limited number of studies have addressed the biological function of the missense mutation situated outside the HD. We next examined DNA binding capability of Csx/Nkx2.5 mutants by EMSA and compared the differences of the mutants in DNA binding affinities to target sites. The atrial natriuretic factor (ANF) promoter contains three specific binding sites for Csx/Nkx2.5. The upstream of transcription start site at positions -242 which contains two binding sequences spaced by 5 nucleotides has higher affinity for Csx/Nkx2.5. Using 32p-labeled 36 bp oligonucleotides of the ANF promoter (−264 bp to −227 bp) and His-Csx/Nkx2.5 fusion protein purified from E. coli BL21 in EMSA, WT Csx/Nkx2.5 exhibited two specific shifted bands, the faster migrating band corresponding to Csx/Nkx2.5 monomer and the slower to dimmer (
Figure 3A). The intensity of both bands was dependent on the protein concentration of WT Csx/Nkx2.5 in the reaction. The protein-DNA binding affinity (Kd) of WT was estimated with a Kd ≈1×10-9 M within the affinity range for the HD protein (Kd in the range of 10-9 to 10-10 M). In the same way, the Kd values of all Csx/Nkx2.5 truncated mutants as His-tagged fusion protein were calculated by EMSA and shown in
Figure 3B. In comparison with full-length Csx/Nkx2.5, the binding affinity of M1, M2 and M3 mutant proteins obviously decreased 5, 15 and 8 times respectively and M4 could not bind DNA due to the absence of HD. M2 mutant, which lacking both NK2-SD and C-terminus, was observed markedly reduced in DNA affinity compared with TN or C-terminal truncated mutants, implying NK2-SD is more potential regulator for the DNA binding and seems to facilitate the DNA-protein interaction in the process of Csx/Nkx2.5 bound to ANF promoter. These findings confirmed that additional protein domains outside HD also play an important role in the regulation of DNA binding.
3.4 Transcriptional activity of Csx/Nkx2.5 mutants
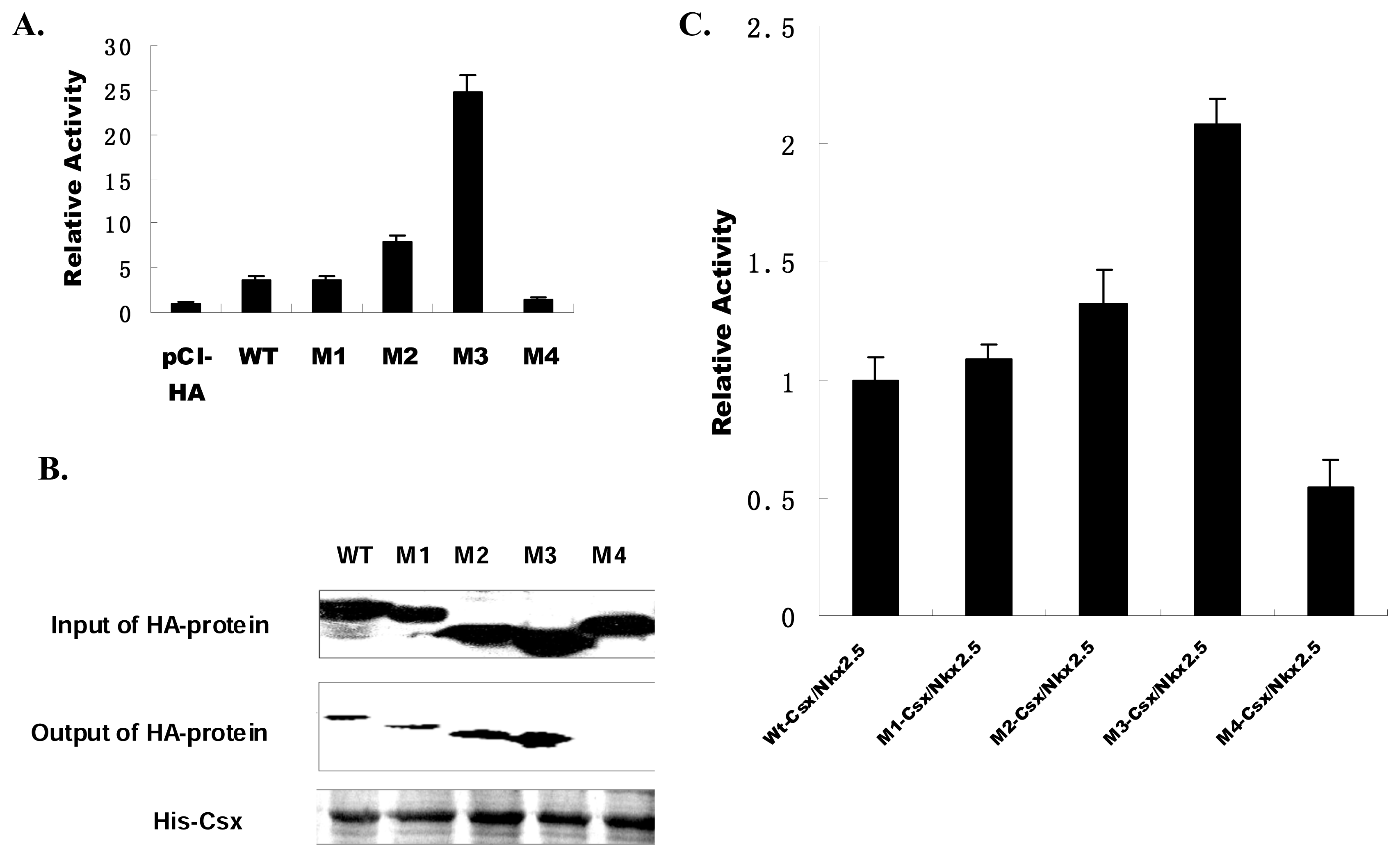
In addition to DNA binding, the Csx/Nkx2.5 plays critical roles in transcriptional regulation and gene expression. To evaluate the transcriptional activation capability of these mutant truncates of Csx/Nkx2.5, we transfected WT or each of these mutants with the ANF-luc promoter-driven reporter into COS-7 cells. WT activated the ANF promoter by ~3-fold. The ANF promoter was also activated by M1, M2 and M3 but not by M4 (
Figure 4A). M1 exhibited similar activity with WT Csx/Nkx2.5, suggesting TN domain as a DNA binding defective mutant was less efficient to affect the transcription of Csx/Nkx2.5. Interestingly, M3 mutant, which lacks the carboxyl terminus and TN domain, more strongly stimulated the transcription of ANF promoter activity by ~7-fold enhancement than WT Csx/Nkx2.5. Regardless of the effect from TN domain, it means the carboxyl terminus outside of the HD is an inhibitory domain for Csx/Nkx2.5 transcriptional activity, similarly to the previous report [
14]. M2 mutant with the deletion of carboxyl terminus and NK2-SD displayed slightly enhanced transcriptional ability than the WT. NK2-SD has been reported as a potential interface of protein-protein interaction [
17]. If Csx/Nkx2.5 is expected to transactivate in combination with other cooperative factors, unlike M3 which strongly enhanced the transcriptional activity, M2’s slight activation of ANF promoter can be attributed to the loss of interaction domain with other transcription factors. It implies that DNA binding ability may not enough to reflect the transcription efficiency of Csx/Nkx2.5, its ability of interaction with other protein or transcriptional factor also essentially contribute to the regulation of its transcription ability. As a dominant negative mutant, M4 showed no any transcriptional activity because of its inability of binding to target DNA. The different characteristic of individual domain in Csx/Nkx2.5 makes these Csx/Nkx2.5 mutants to display different transcriptional activities.
3.5 Effects of mutants on the activity of wild-type Csx/Nkx2.5
Csx/Nkx2.5 physically interacts with each other in vitro as well as in cells, the complex protein-protein interaction plays a role in regulation of its transcriptional function. Therefore, it is critical to examine whether Csx/Nkx2.5 and its mutant proteins can homodimerize to regulate their transcriptional activity. Using Pull-down assays to examine the dimer formation of the mutant proteins with WT Csx/Nkx2.5, we found that M1, M2 and M3 mutants displayed similar interaction with WT, but M4 mutant exhibited very slight or even no interaction with WT Csx/Nkx2.5 (
Figure 4B), in consistent with the previous report that HD is essential for dimerization [
3,
15]. Accordingly, the effect of each mutant on WT Csx/Nkx2.5 was validated by co-transfection of the expression plasmid encoding each of mutants with WT Csx/Nkx2.5. We examined the function of transcriptional activation using the ANF-luc reporter. Luciferase activation after co-transfection of WT-Csx/Nkx2.5 was defined as 100% (1-fold). Compared with WT, M3-Csx/Nkx2.5 displayed approximately 200% increase (>2-fold) in activation of ANF-luciferase reporter (
Figure 4C), but which is far lower than the transcriptional activity of M3 alone (
Figure 4A). According to our previous conclusion that C-terminus of Csx/Nkx2.5 is a transcription inhibitory domain, but in M3/WT dimmer WT holds intact C-terminus, so WT can partially hamper the enhanced transcriptional activation capability of M3 mutant. Reduction in luciferase activity was observed in M4 due to its lack of HD. M1-Csx/Nkx2.5 and M2-Csx/Nkx2.5 did not exhibit inhibitory effects, rather transactivate a little bit higher than WT. In other words, the regions beside of HD did not significantly affect the dimmer formation, but could exert some influence on the transcriptional activity of WT through protein-protein interaction.
4. Discussion
Mutation of human Csx/Nkx2.5 has been associated with various congenital heart diseases such as ASD, ventricular septal defect (VSD). Some mutations identified in sporadic CHD were missense situated outside the HD. Currently, two peptide domains: TN-domain and NK2-SD, in addition to the HD, which are conserved within NK2 class, have not been shown their clear functions. In this study, we constructed four truncated mutants, respectively deleted TN-domain, NK2-SD, HD and C-terminus, and examined the function of these truncated mutants in order to gain insights into the nature of the regions outsides HD.
Previous studies have revealed that Csx/Nkx2.5 has the capabilities of nuclear localization, DNA binding, transcriptional activation, and protein-protein interactions. So all the truncated mutants had been examined their individual biochemical properties and their effect on the functions of Csx/Nkx2.5. Our result showed that the mutant lacking the HD loses its nuclear localization signal. TN-domain and NK2-SD have no effect on the nuclear distribution.
Noticeably, the greatest similarity between all the mutant proteins studied here was the markedly reduced DNA binding affinity. The protein saturation concentration of M2 was ~15 fold higher than WT Csx/Nkx2.5, implying NK2-SD domain may take a more important role in protein-DNA interaction. Based on the observed DNA binding reduction, we analyzed the transcriptional activity of mutants accordingly. All the mutants were expected to have little or no transcriptional activation function, but surprisingly, M3 mutant strongly activated the ANF promoter up to ~24-fold, and M1 and M2 showed a weakly enhanced activation. The transcription activity results appeared disagreed with their ability of DNA binding. It may be attributed to that Csx/Nkx2.5 required the protein-protein interaction to activate the transcriptional activity far more than the protein-DNA interaction. The similar results were obtained by co-transfection of WT Csx/Nkx2.5 with each of mutants to assay the transcriptional activity. By His pull-down assay, these mutants, except M4 lacking HD, were found to associated with WT Csx/Nkx2.5 and showed no obviously reduced dimer formation. It suggested that these mutants did not act in a dominant inhibitory manner and the domains besides HD represented as regulator during Csx/Nkx2.5 transcription. We also found the C-terminus deletion mutant of Csx/Nkx2.5 had a significantly increased transcriptional activity on ANF promoter compared with WT. It indicated that the C-terminus region may contain transcriptional repression domain. Moreover, NK-SD may operate as an interface of protein-protein interaction to regulate the transcriptional activity.
In summary, mutants located outside the HD retained intact nuclear localization and had nearly normal or increased transcriptional activity but impaired DNA binding capability. The C-terminus region exhibited an inhibitory function on transcriptional activity of WT Csx/Nkx2.5. The NK2-SD was likely to facilitate both DNA binding and protein-protein interaction, TN-domain seemed no precise function. In the current study, Csx/Nkx2.5 deletion mutant in HD domain displayed extremely different biological appearance from the mutants with deletion outside of the HD, these may explain the clinical observation that patients with missense situated outside the HD were not associated with AV conduction disturbance [
13].

{kind=link}
{kind=link}
{kind=link}
{kind=link}



