Protective Effect of Encapsulation in Fermentation of Limonene-contained Media and Orange Peel Hydrolyzate



Abstract
:1. Introduction
2. Results and Discussion
2.1. Enzymatic Hydrolysis
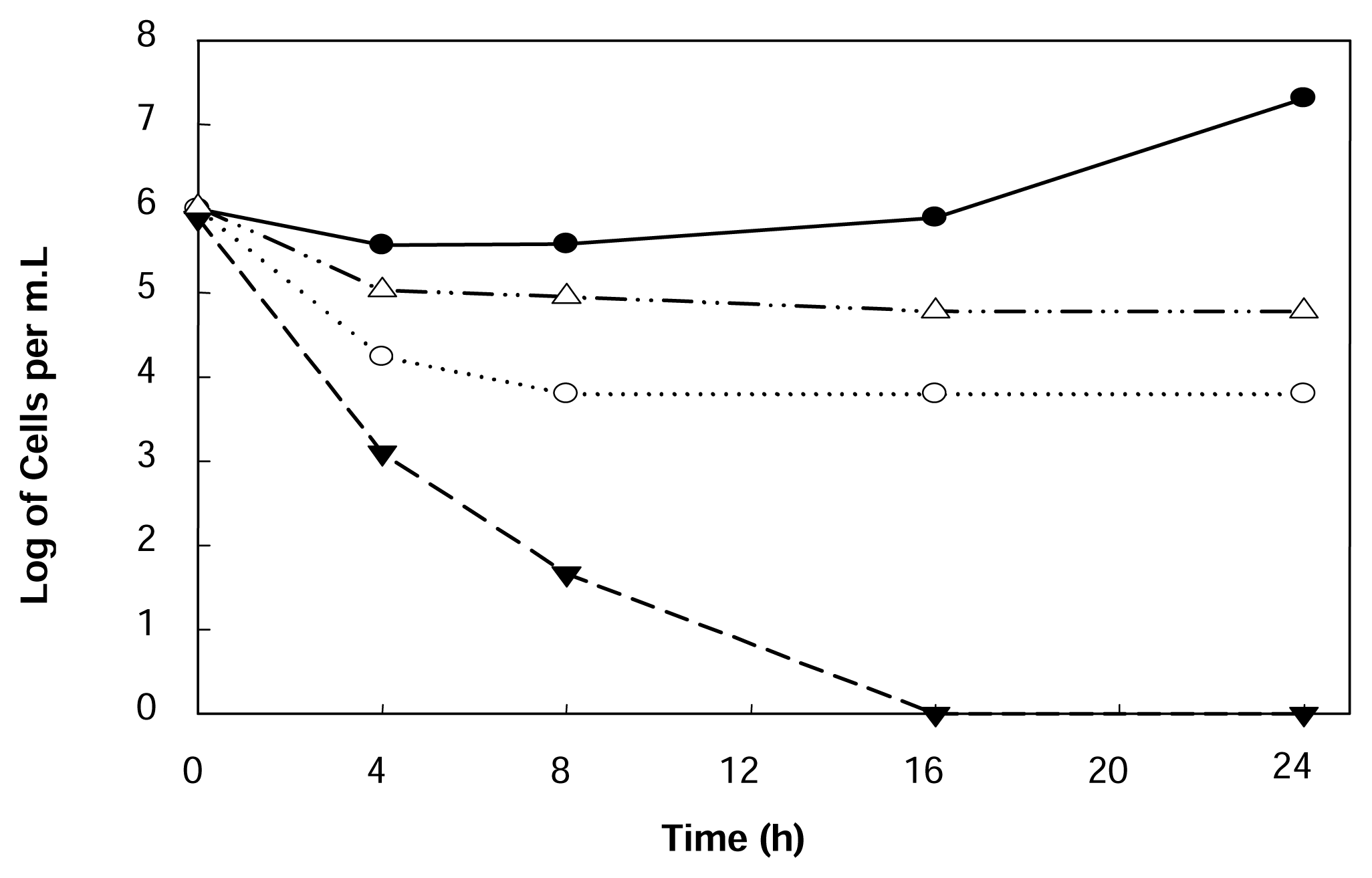
2.2. Free cell cultivation
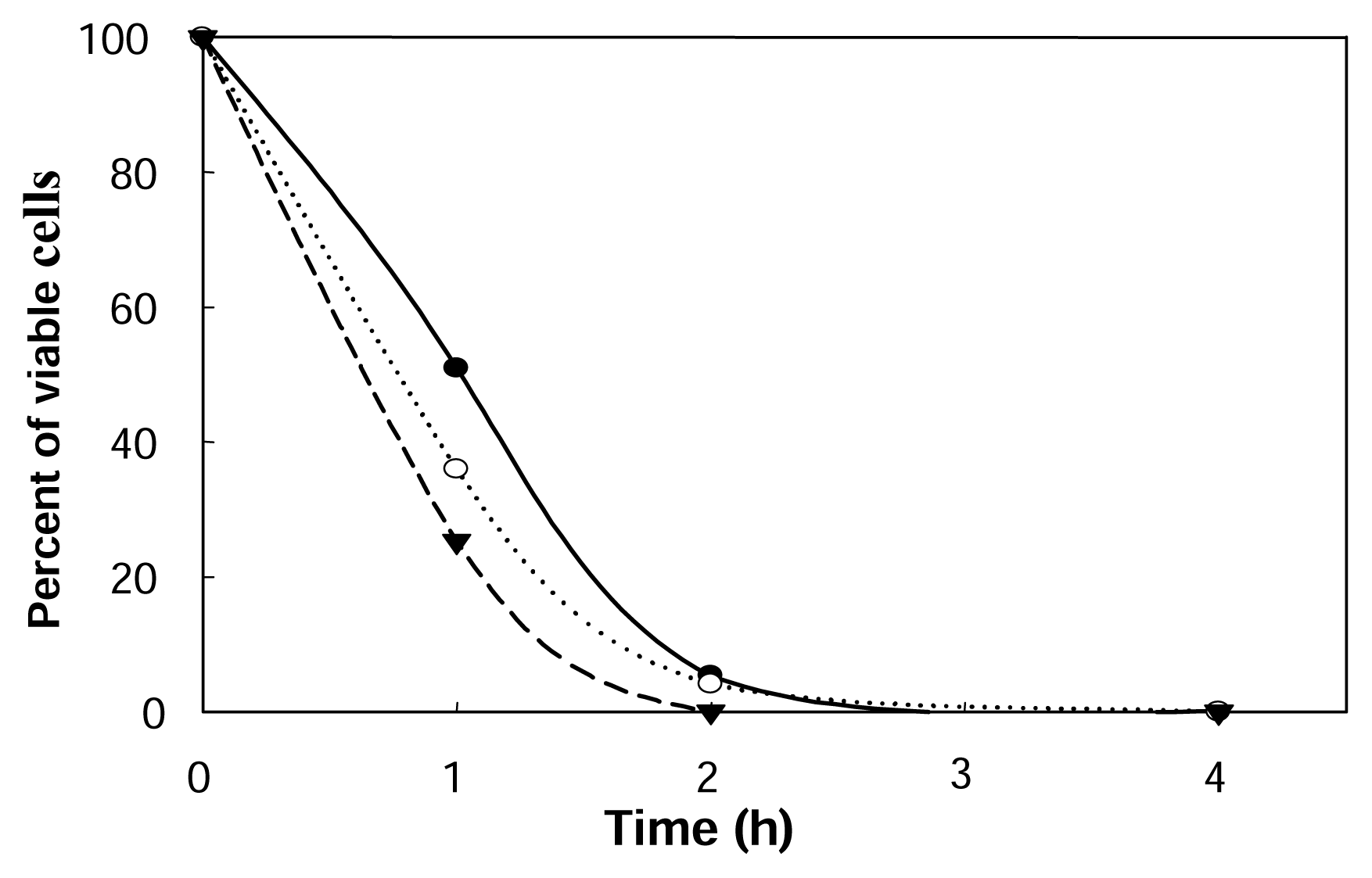
2.3. Cultivation of encapsulated cells in the presence of limonene
2.4. Yeast cultivation in peel hydrolyzate
2.5. Selectivity of capsules’ membrane
2.6. Conclusions
3. Material and Methods
3.1. Substrates and enzymes
3.2. Hydrolyses
3.3. Yeast Strain and Media
3.4. Encapsulation Method and Batch Cultivation Experiments
3.5. Selectivity of capsules’ membrane
3.6. Analytical methods





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carbohydrate | % (of total solid) |
|---|---|
| Glucose | 22.9 ± 2.4 |
| Fructose | 14.1 ± 1.3 |
| Galactose | 4.0 ± 0.2 |
| Arabinose | 7.1 ± 0.5 |
| Xylose | 0.4 ± 0.1 |
| Galacturonic acid | 19.0 ± 1.7 |
| Total | 67.5 |
| Parameter | Suspended Cells | Encapsulated cells with different limonene concentration (% v/v) of: | Peel hydrolyzate | |||
|---|---|---|---|---|---|---|
| 0 | 0.5 | 1 | 1.5 | |||
| −rS (g/L.h) | 4.0 | 8.56 | 5.96 | 5.38 | 5.58 | 5.00 |
| rE(g/L.h) | 1.6 | 3.69 | 2.57 | 2.28 | 2.33 | 2.18 |
| YSE(g/g) | 0.41(±0.03) | 0.43(±0.02) | 0.43(±0.01) | 0.42(±0.01) | 0.42(±0.01) | 0.44(±0.01) |
| YSGly(g/g) | 0.080 | 0.062 | 0.050 | 0.048 | 0.047 | 0.050 |
| YSAce(g/g) | 0.018 | 0.017 | 0.016 | 0.016 | 0.015 | 0.017 |
| YSSuc(g/g) | 0.003 | 0.003 | 0.003 | 0.003 | 0.003 | 0.003 |
Acknowledgments
References
- Plessas, S.; Bekatorou, A.; Koutinas, A.A.; Soupioni, M.; Banat, I.M.; Marchant, R. Use of Saccharomyces cerevisiae cells immobilized on orange peel as biocatalyst for alcoholic fermentation. Bioresour. Technol 2007, 98, 860–865. [Google Scholar]
- Grohmann, K.; Baldwin, E.A. Hydrolysis of Orange Peel with Pectinase and Cellulase Enzymes. Biotechnol. Lett 1992, 14, 1169–1174. [Google Scholar]
- Tripodo, M.M.; Lanuzza, F.; Micali, G.; Coppolino, R.; Nucita, F. Citrus waste recovery: a new environmentally friendly procedure to obtain animal feed. Bioresour. Technol 2004, 91, 111–115. [Google Scholar]
- Wilkins, M.R.; Widmer, W.W.; Grohmann, K.; Cameron, R.G. Hydrolysis of grapefruit peel waste with cellulase and pectinase enzymes. Bioresour. Technol 2007, 98, 1596–1601. [Google Scholar]
- Murdock, D.I.; Allen, W.E. Germicidal Effect of Orange Peel Oil and D-Limonene in Water and Orange Juice. Food Technol 1960, 14, 441–445. [Google Scholar]
- Winniczuk, P.P.; Parish, M.E. Minimum inhibitory concentrations of antimicrobials against micro-organisms related to citrus juice. Food Microbiol 1997, 14, 373–381. [Google Scholar]
- Grohmann, K.; Baldwin, E.A.; Buslig, B.S. Production of Ethanol from Enzymatically Hydrolyzed Orange Peel by the Yeast Saccharomyces-Cerevisiae. Appl. Biochem. Biotechnol. 1994, (45–6), 315–327. [Google Scholar]
- Lane, A.G. Removal of Peel Oil from Citrus Peel Press Liquors before Anaerobic-Digestion. Environ. Technol. Lett 1983, 4, 65–72. [Google Scholar]
- Park, J.K.; Chang, H.N. Microencapsulation of microbial cells. Biotechnol. Adv 2000, 18, 303–319. [Google Scholar]
- Talebnia, F.; Niklasson, C.; Taherzadeh, M.J. Ethanol production from glucose and dilute-acid hydrolyzates by encapsulated S-cerevisiae. Biotechnol. Bioeng 2005, 90, 345–353. [Google Scholar]
- Decker, S.R.; Adney, W.S.; Jennings, E.; Vinzant, T.B.; Himmel, M.E. Automated filter paper assay for determination of cellulase activity. Appl. Biochem. Biotechnol 2003, 105, 689–703. [Google Scholar]
- Taherzadeh, M.J.; Liden, G.; Gustafsson, L.; Niklasson, C. The effects of pantothenate deficiency and acetate addition on anaerobic batch fermentation of glucose by Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol 1996, 46, 176–182. [Google Scholar]
- Taherzadeh, M.J.; Eklund, R.; Gustafsson, L.; Niklasson, C.; Liden, G. Characterization and fermentation of dilute-acid hydrolyzates from wood. Ind. Eng. Chem. Res 1997, 36, 4659–4665. [Google Scholar]
- Talebnia, F.; Taherzadeh, M.J. In situ detoxification and continuous cultivation of dilute-acid hydrolyzate to ethanol by encapsulated S. cerevisiae. J. Biotechnol 2006, 125, 377–384. [Google Scholar]
- Sipailiene, A.; Venskutonis, P.R.; Baranauskiene, R.; Sarkinas, A. Antimicrobial activity of commercial samples of thyme and marjoram oils. J. Essent. Oil Res 2006, 18, 698–703. [Google Scholar]
- Subba, M.S.; Soumithri, T.C; Rao, R.S. Antimicrobial Action of Citrus Oils. J. Food Sci 1967, 32, 225–227. [Google Scholar]
- Bishop, J.R.P.; Nelson, G.; Lamb, J. Microencapsulation in yeast cells. J. Microencapsul 1998, 15, 761–773. [Google Scholar]
- Kimura, K.; Yamaoka, M.; Kamisaka, Y. Inhibition of Lipid Accumulation and Lipid Body Formation in Oleaginous Yeast by Effective Components in Spices, Carvacrol, Eugenol, Thymol, and Piperine. J. Agric. Food Chem 2006, 54, 3528–3534. [Google Scholar]
- Chang, H.N.; Seong, G.H.; Yoo, I.K.; Park, J.K.; Seo, J.H. Microencapsulation of recombinant Saccharomyces cerevisiae cells with invertase activity in liquid-core alginate capsules. Biotechnol. Bioeng 1996, 51, 157–162. [Google Scholar]
© 2007 by MDPI Reproduction is permitted for noncommercial purposes.
Share and Cite
Pourbafrani, M.; Talebnia, F.; Niklasson, C.; Taherzadeh, M.J. Protective Effect of Encapsulation in Fermentation of Limonene-contained Media and Orange Peel Hydrolyzate. Int. J. Mol. Sci. 2007, 8, 777-787. https://doi.org/10.3390/i8080777
Pourbafrani M, Talebnia F, Niklasson C, Taherzadeh MJ. Protective Effect of Encapsulation in Fermentation of Limonene-contained Media and Orange Peel Hydrolyzate. International Journal of Molecular Sciences. 2007; 8(8):777-787. https://doi.org/10.3390/i8080777
Chicago/Turabian StylePourbafrani, Mohammad, Farid Talebnia, Claes Niklasson, and Mohammad J. Taherzadeh. 2007. "Protective Effect of Encapsulation in Fermentation of Limonene-contained Media and Orange Peel Hydrolyzate" International Journal of Molecular Sciences 8, no. 8: 777-787. https://doi.org/10.3390/i8080777