Elevation of Inducible Nitric Oxide Synthase and Cyclooxygenase-2 Expression in the Mouse Brain after Chronic Nonylphenol Exposure

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
2.1. NP increases the NO level in the mice brain
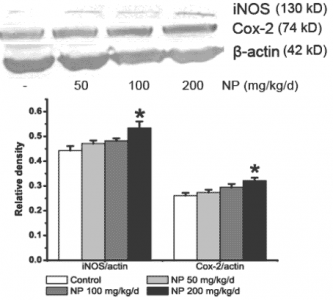
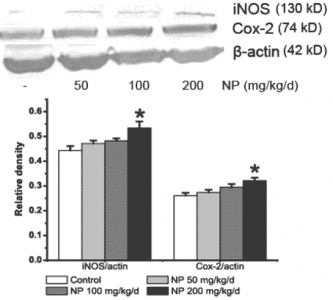
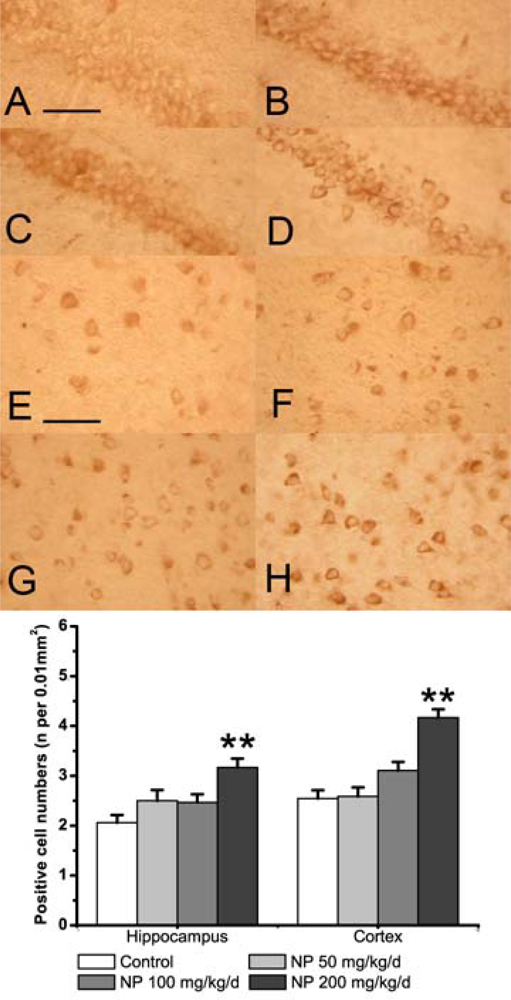
2.2. NP increases the NOS activity and the iNOS expression in the mice brain
2.3. NP increases the expression of COX-2 in the mice brain
3. Experimental Section
3.1. Animals and treatments
3.2. Brain nitrates and nitrites (NOx) assay
3.3. Assay of Nitric oxide synthase activity
3.4. Western blot
3.5. Immunohistochemistry
3.6. Statistical analysis
Acknowledgments
References
- Han, XD; Tu, ZG; Gong, Y; Shen, SN; Wang, XY; Kang, LN; Hou, YY; Chen, JX. The toxic effects of nonylphenol on the reproductive system of male rats. Reprod.Toxicol. 2004, 19, 215–221. [Google Scholar]
- Yao, G; Hou, Y. Nonylphenol induces thymocyte apoptosis through Fas/FasL pathway by mimicking estrogen in vivo. Environ. Toxicol. Pharmacol. 19–27.
- Negishi, T; Kawasaki, K; Suzaki, S; Maeda, H; Ishii, Y; Kyuwa, S; Kuroda, Y; Yoshikawa, Y. Behavioral alterations in response to fear-provoking stimuli and tranylcypromine induced by perinatal exposure to bisphenol A and nonylphenol in male rats. Environ. Health Perspect. 2004, 112, 1159–1164. [Google Scholar]
- Negishi, T; Ishii, Y; Kyuwa, S; Kuroda, Y; Yoshikawa, Y. Inhibition of staurosporine-induced neuronal cell death by bisphenol A and nonylphenol in primary cultured rat hippocampal and cortical neurons. Neurosci. Lett. 2003, 353, 99–102. [Google Scholar]
- Obata, T; Kubota, S. Formation of hydroxy radicals by environmental estrogen-like chemicals in rat striatum. Neurosci. Lett. 2000, 296, 41–44. [Google Scholar]
- Dalgleish, AG; O'Byrne, KJ. Chronic immune activation and inflammation in the pathogenesis of AIDS and cancer. Adv. Cancer Res. 2002, 84, 231–276. [Google Scholar]
- Dranoff, G. Coordinated tumor immunity. J. Clin. Invest. 2003, 111, 1116–1118. [Google Scholar]
- Heneka, MT; O'Banion, MK. Inflammatory processes in Alzheimer's disease. J. Neuroimmunol. 2007, 184, 69–91. [Google Scholar]
- Tansey, MG; McCoy, MK; Frank-Cannon, TC. Neuroinflammatory mechanisms in Parkinson's disease: Potential environmental triggers, pathways, and targets for early therapeutic intervention. Exp. Neurol. 2007, 208, 1–25. [Google Scholar]
- Ekdahl, CT; Claasen, JH; Bonde, S; Kokaia, Z; Lindvall, O. Inflammation is detrimental for neurogenesis in adult brain. Proc. Natl. Acad. Sci. USA 2007, 100, 13632–13637. [Google Scholar]
- Kim, K. Differential expression of neuronal and inducible nitric oxide synthase in rat brain after subchronic administration of 3-monochloro-1,2-propanediol. Food Chem. Toxicol. 2008, 46, 960–955. [Google Scholar]
- Zhang, J; Mori, A; Chen, Q; Zhao, B. Fermented papaya preparation attenuates beta-amyloid precursor protein: beta-amyloid-mediated copper neurotoxicity in beta-amyloid precursor protein and beta-amyloid precursor protein Swedish mutation overexpressing SH-SY5Y cells. Neuroscience 2006, 143, 63–72. [Google Scholar]
- Chen, C; Magee, JC; Bazan, NG. Cyclooxygenase-2 Regulates Prostaglandin E2 Signaling in Hippocampal Long-Term Synaptic Plasticity. J. Neurophysiol. 2002, 87, 2851–2857. [Google Scholar]
- Xiang, Z; Ho, L; Valdellon, J; Borchelt, D; Kelley, K; Spielman, L; Aisen, PS; Pasinetti, GM. Cyclooxygenase (COX)-2 and cell cycle activity in a transgenic mouse model of Alzheimer's Disease neuropathology. Neurobiol. Aging 2002, 23, 327–334. [Google Scholar]
- Colborn, T. Neurodevelopment and endocrine disruption. Environ. Health Perspect. 2004, 112, 944–949. [Google Scholar]
- Schantz, SL; Widholm, JJ. Cognitive effects of endocrine-disrupting chemicals in animals. Environ. Health Perspect. 2001, 109, 1197–1206. [Google Scholar]
- Arukwe, A; Thibaut, R; Ingebrigtsen, K; Celius, T; Goksoyr, A; Cravedi, J. In vivo and in vitro metabolism and organ distribution of nonylphenol in Atlantic salmon (Salmo salar). Aquat. Toxicol. 2000, 249, 289–304. [Google Scholar]
- Doerge, DR; Twaddle, NC; Churchwell, MI; Chang, HC; Newbold, RR; Delclos, KB. Mass spectrometric determination of p-nonylphenol metabolism and disposition following oral administration to Sprague-Dawley rats. Reprod. Toxicol. 45–56.
- Zalko, D; Costagliola, R; Dorio, C; Rathahao, E; Cravedi, J. In Vivo Metabolic Fate of the Xeno-Estrogen 4-n-Nonylphenol in Wistar Rats. Drug Metab. Dispos. 2003, 31, 168–178. [Google Scholar]
- Nagao, T; Wada, K; Marumo, H; Yoshimura, S; Ono, H. Reproductive effects of nonylphenol in rats after gavage administration: a two-generation study. Reprod. Toxicol. 2001, 15, 293–315. [Google Scholar]
- Leong, SK; Ruan, RS; Zhang, Z. A critical assessment of the neurodestructive and neuroprotective effects of nitric oxide. Ann. NY Acad. Sci. 2002, 962, 161–181. [Google Scholar]
- Guix, FX; Uribesalgo, I; Coma, M; Munoz, FJ. The physiology and pathophysiology of nitric oxide in the brain. Prog. Neurobiol. 2005, 76, 126–152. [Google Scholar]
- Beckman, JS; Beckman, TW; Chen, J; Marshall, PA; Freeman, BA. Apparent hydroxyl radical production by peroxynitrite: implications for endothelial injury from nitric oxide and superoxide. Proc. Natl. Acad. Sci. USA 1990, 87, 1624–1620. [Google Scholar]
- Radi, R; Beckman, JS; Bush, KM; Freeman, BA. Peroxynitrite oxidation of sulfhydryls: The cytotoxic potential of superoxide and nitric oxide. J. Biol. Chem. 1991, 266, 4250–4244. [Google Scholar]
- Dawson, VL; Dawson, TM. Nitric oxide neurotoxicity. J. Chem. Neuroanat. 1996, 10, 179–190. [Google Scholar]
- Ischiropoulos, H; Beckman, JS. Oxidative stress and nitration in neurodegeneration: cause, effect, or association? J. Clin. Invest. 2003, 11, 163–169. [Google Scholar]
- Wei, T; Chen, C; Hou, J; Xin, W; Mori, A. Nitric oxide induces oxidative stress and apoptosis in neuronal cells. Biochim. Biophys. Acta 2000, 1498, 72–79. [Google Scholar]
- Brune, B; von Knethen, A; Sandau, KB. Nitric oxide and its role in apoptosis. Eur. J. Pharmacol. 1998, 351, 272–261. [Google Scholar]
- Heneka, MT; Feinstein, DL. Expression and function of inducible nitric oxide synthase in neurons. J. Neuroimmunol. 2001, 114, 8–18. [Google Scholar]
- Kim, JY; Choi, CY; Lee, KJ; Shin, DW; Jung, KS; Chung, YC; Lee, SS; Shin, JG; Jeong, HG. Induction of inducible nitric oxide synthase and proinflammatory cytokines expression by o,p-DDT in macrophages. Toxicol. Lett. 2004, 14, 269–261. [Google Scholar]
- Hewett, SJ; Silakova, JM; Hewett, JA. Oral Treatment with Rofecoxib Reduces Hippocampal Excitotoxic Neurodegeneration. J. Pharmacol. Exp. Ther. 2006, 319, 1219–1224. [Google Scholar]
- Minghetti, L. Cyclooxygenase-2 (COX-2) in Inflammatory and Degenerative Brain Diseases. J. Neuropathol. Exp. Neurol. 2004, 63, 901–910. [Google Scholar]
- Kadoyama, K; Takahashi, Y; Higashida, H; Tanabe, T; Yoshimoto, T. Overexpression of cyclooxygenase-2 stimulates amyloid beta-peptide production in neuronal cells. Int. Congr. Ser. 2002, 1233, 349–353. [Google Scholar]
- Tracey, WR; Tse, J; Carter, G. Lipopolysaccaride induced changes in plasma nitrite andnitrate concentrations in rats and mice: Pharmacological evaluation of nitric oxide synthase inhibitors. J. Pharmacol. Exp. Ther. 1995, 282, 1011–1015. [Google Scholar]
- Fernandez-Cancio, M; Fernandez-Vitos, EM; Centelles, JJ; Imperial, S. Sources of interference in the use of 2,3-diaminonaphthalene for the fluorimetric determination of nitric oxide synthase activity in biological samples. Clin. Chim. Acta 2001, 312, 205–212. [Google Scholar]




© 2008 by MDPI This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, Y.-Q.; Mao, Z.; Zheng, Y.-L.; Han, B.-P.; Chen, L.-T.; Li, J.; Li, F. Elevation of Inducible Nitric Oxide Synthase and Cyclooxygenase-2 Expression in the Mouse Brain after Chronic Nonylphenol Exposure. Int. J. Mol. Sci. 2008, 9, 1977-1988. https://doi.org/10.3390/ijms9101977
Zhang Y-Q, Mao Z, Zheng Y-L, Han B-P, Chen L-T, Li J, Li F. Elevation of Inducible Nitric Oxide Synthase and Cyclooxygenase-2 Expression in the Mouse Brain after Chronic Nonylphenol Exposure. International Journal of Molecular Sciences. 2008; 9(10):1977-1988. https://doi.org/10.3390/ijms9101977
Chicago/Turabian StyleZhang, Yan-Qiu, Zhen Mao, Yuan-Lin Zheng, Bao-Ping Han, Ling-Tong Chen, Jing Li, and Fei Li. 2008. "Elevation of Inducible Nitric Oxide Synthase and Cyclooxygenase-2 Expression in the Mouse Brain after Chronic Nonylphenol Exposure" International Journal of Molecular Sciences 9, no. 10: 1977-1988. https://doi.org/10.3390/ijms9101977