Zearalenone and Reproductive Function in Farm Animals

Abstract
:1. Introduction
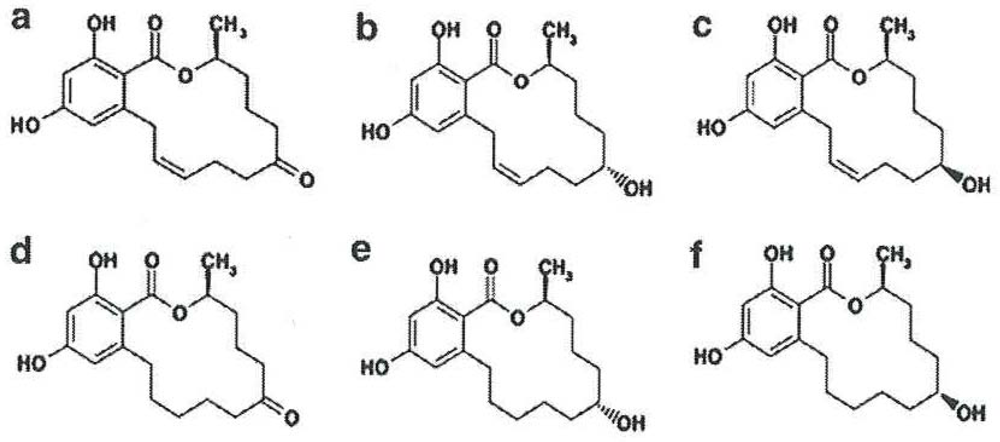
2. Metabolism of zearalenone in farm animals
- Hydroxylation resulting in the formation of α- and β-ZOL, catalyzed by 3α- and 3β- hydroxysteroid dehydrogenase (HSDs);
- Conjugation of ZEA and its reduced metabolites with glucuronic acid, catalyzed by uridine diphosphate glucuronyl transferase (UDPGT).
2.1. Pig
2.2. Ovine
2.3. Bovine
2.4. Horse
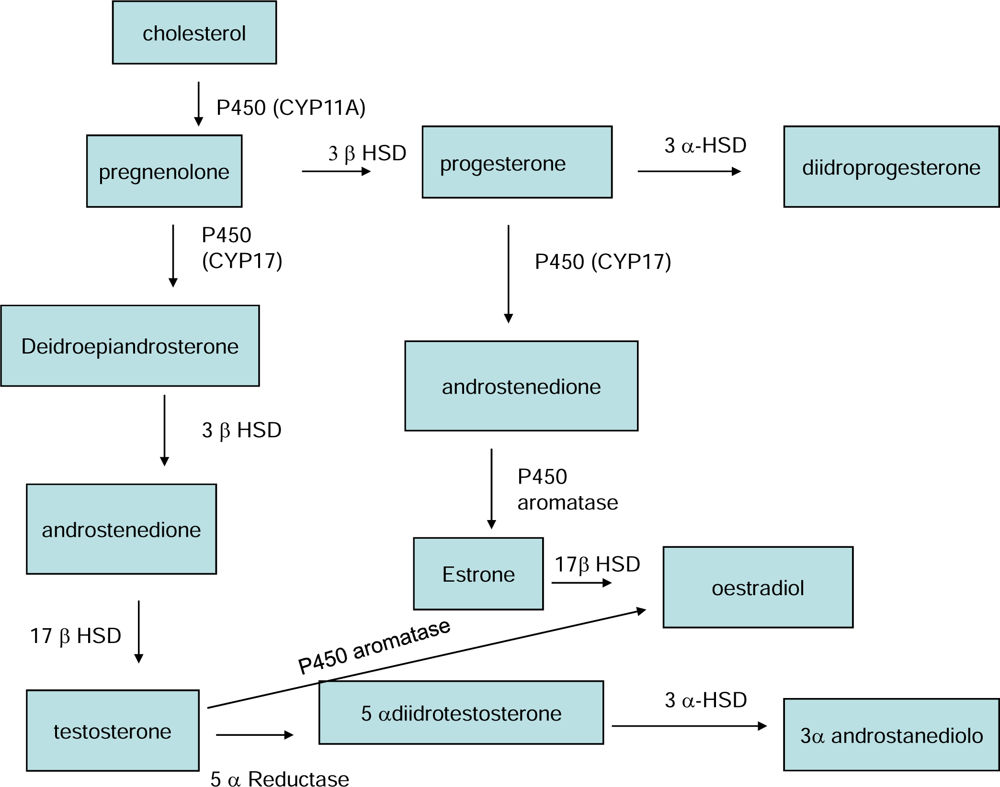
3. Action mechanisms of zearalenone and its derivatives
4 In vivo exposure of zearalenone and its derivatives
4.1. Pig
4.2. Bovine
4.2.1 Ovine
4.3. Horses
5. In vitro exposure of zearalenone and its derivatives
5.1. Follicle development, oocyte maturation and embryo development
5.1.1. Sow
5.1.2. Cattle
5.1.3. Mare
5.2. Spermatozoa parameters
5.2.1. Boar
5.2.2. Stallion
6. Conclusions
References and Notes
- Zinedine, A; Soriano, JM; Moltò, JC; Magňes, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: an estrogenic mycotoxin. Food Chem. Toxicol 2007, 45, 1–18. [Google Scholar]
- Bottalico, A; Visconti, A; Logrieco, A; Solfrizzo, M; Mirocha, CJ. Occurrence of zearalenols (diastereomeric mixture) in corn stalk rot and their production by associated Fusarium species. Appl. Environ. Microbiol 1985, 49, 547–551. [Google Scholar]
- Pittet, A. Natural occurrence of mycotoxins in food and feeds: A decade in review. In Mycotoxins and Phytotoxins in Perspective at the Turn of the Millennium. Proceedings of the Xth International IUPAC symposium on mycotoxins and phytotoxins; De Koe, WJ, Samson, RA, van Egmond, HP, Gilbert, J, Sabino, M, Eds.; Ponsen & Looyen: Wageninger, The Netherlands, 2001; pp. 153–172. [Google Scholar]
- Minervini, F; Giannoccaro, A; Cavallini, A; Visconti, A. Investigations on cellular proliferation induced by zearalenone and its derivatives in relation to the estrogenic parameters. Toxicol. Lett 2005, 159, 272–283. [Google Scholar]
- European Food Safety Authority (EFSA). Opinion of the scientific panel of contaminants in the food chain on a request from the Commission related to zearalenone as undesirable substance in animal feed. The EFSA J 2004, 8, 1–35. [Google Scholar]
- Olsen, M. Metabolism of zearalenone in farm animals. In Fusarium mycotoxins, taxonomy and pathogenicity, 1st Ed; Chelkowsi, J, Ed.; Elsevier: Amsterdam-Oxford-New York, 1989; pp. 167–177. [Google Scholar]
- Gajęcki, M. Zearalenone — undesirable substances in feed. Pol. J. Vet. Sci 2002, 5, 117–122. [Google Scholar]
- D’Mello, JPF; Placinta, CM; MacDonald, AMC. Fusarium mycotoxins: A review of global implications for animal health, welfare and productivity. Anim. Feed Sci. Technol 1999, 80, 183–205. [Google Scholar]
- Malekinejad, H; Maas-Bakker, R; Fink-Gremmels, J. Species differences in the hepatic biotransformation of zearalenone. Vet. J 2006, 172, 96–102. [Google Scholar]
- Malekinejad, H; Colenbrander, B; Fink-Gremmels, J. Hydroxysteroid dehydrogenases in bovine and porcine granulosa cells convert zearalenone into its hydroxylated metabolites α-zearalenol and β-zearalenol. Vet. Res. Commun 2006, 30, 445–453. [Google Scholar]
- Olsen, M; Malmlöf, K; Pettersson, H; Sandholm, K; Kiessling, KH. Plasma and urinary levels of zearalenone and α-zearalenol in a prepubertal gilt fed zearalenone. Acta Pharmacol. Et Toxicol 1985, 56, 239–243. [Google Scholar]
- Curtui, VG; Gareis, M; Usleber, E; Märtlbauer, E. Survey of Romanian slaughtered pigs for the occurrence of mycotoxins ochratosins A and B, and zearalenone. Food Addit. Contam 2001, 18, 730–738. [Google Scholar]
- Erasmuson, AF; Scahill, BG; West, DM. Natural zeranol (α-zearalanol) in the urine of pasture-fed animals. J. Agric. Food Chem 1994, 42, 2721–2725. [Google Scholar]
- Songsermsakul, P; Sontag, G; Cichna-Markl, M; Zentek, J; Razzazi-Fazeli, E. Determination of zearalenone and its metabolites in urine, plasma and faeces of horses by HPLC-APCI-MS. J. Chromatogr.B 2006, 843, 252–261. [Google Scholar]
- Filannino, A; Lacalandra, GM; Minervini, F; Nicassio, M; Dell’Aquila, ME; Visconti, A. Exposure of stallions to mycotoxin zearalenone and its in vitro effect on sperm chromatin structure stability (SCSA). Reprod. Dom. Anim 2008, 43(suppl 3), 212. [Google Scholar]
- Kuiper-Goodman, T; Scott, PM; Watanabe, H. Risk assessment of the mycotoxin zearalenone. Reg. Toxicol. Pharmacol 1987, 7, 253–306. [Google Scholar]
- European Commission. Opinion of the Scientific Committee on food on Fusarium Toxins: Part 2: Zearalenone (ZEA). http://ec.europa.eu/food/fs/sc/scf/out65_en.pdf, expressed June 22, 2000.
- Giammarino, A; Manera, M; Robbe, D; Perugini, M; Minervini, F; Amorena, M. Influence of mycotoxins on spontaneous contraction in myometrial strips of prepubertal lamb. Res. Vet. Sci 2008, 84, 471–476. [Google Scholar]
- Monje, P; Boland, R. Characterization of membrane estrogen binding proteins from rabbit uterus. Mol. Cell. Endocrinol 1999, 147, 75–84. [Google Scholar]
- Payne, AH; Hales, DB. Overview of steroidogenic enzymes in the pathway from cholesterol to active steroid hormones. Endocr. Rev 2004, 25, 947–970. [Google Scholar]
- Stoffel-Wagner, B. Neurosteroid metabolism in the human brain. Eu. J. Endocrinol 2001, 145, 669–679. [Google Scholar]
- Penning, TM; Burczynski, ME; Jez, JM; Hung, C-F; Lin, HK; Ma, H; Moore, M; Palackal, N; Ratnam, K. Human 3α-hydroxysteroid dehydrogenase isoforms (AKR1C1-AKR1C4) of the aldo-keto reductase superfamily: functional plasticity and tissue distribution reveals roles in the inactivation and formation of male and female sex hormones. Biochem. J 2000, 351, 67–77. [Google Scholar]
- Yang, J; Zhang, Y; Wang, Y; Cui, S. Toxic effects of zearalenone and α-zearalenol on the regulation of steroidogenesis and testosterone production in mouse Leydig cells. Toxicol. In Vitro 2007, 21, 558–565. [Google Scholar]
- Haschek, WM; Haliburton, JC. Fusarium moniliforme and zearalenone toxicoses in domestic animals: A review. In Diagnosis of Mycotoxicoses; Richard, JL, Thurston, JR, Eds.; Martinus Nijhoff: Dordrecht, The Netherlands, 1986; pp. 213–235. [Google Scholar]
- Obremski, K; Gajęcki, M; Zwierzchowski, W; Zielonka, L; Otrocka-Domagala, I; Rotkiewicz, T; Mikolajczyk, A; Gajęcka, M; Polak, M. Influence of zearalenone on reproductive system cell proliferation in gilts. Pol. J. Vet. Sci 2003, 6, 239–245. [Google Scholar]
- Zwierzchowski, W; Przybylowicz, M; Obremski, K; Zielonka, L; Skorska-Wyszynska, E; Gajecka, M; Polak, M; Jakimiuk, E; Jana, B; Rybarczyk, L; Gajecki, M. Level of zearalenone in blood serum and lesions in ovarian follicles of sexually immature gilts in the course of zearalenone mycotoxicosis. Pol. J. Vet. Sci 2005, 8, 209–218. [Google Scholar]
- Alm, H; Brüssow, KP; Torner, H; Valselow, J; Tomek, W; Dänicke, S; Tiemann, U. Influence of Fusarium-toxin contaminated feed on initial quality and meiotic competence of gilt oocytes. Reprod. Toxicol 2006, 22, 44–50. [Google Scholar]
- Mirocha, CJ; Pathre, SV; Christensen, CM; Zearalenone. Mycotoxins in Human and Animal Health; Rodricks, JV, Hesseltine, CW, Mehlman, MA, Eds.; Pathotox Publisher Inc: Park Forest South, IL, USA, 1977; pp. 346–364. [Google Scholar]
- Long, GG; Turek, J; Diekman, MA; Scheidt, AB. Effect of zearalenone on days 7 to 10 post-mating on blastocyst development and endometrial morphology in sows. Vet. Pathol 1992, 29, 60–67. [Google Scholar]
- Ruhr, LP; Osweiler, GD; Foley, CW. Effect of the estrogenic mycotoxin zearalenone on reproductive potential in the boar. Am. J. Vet. Res 1983, 44, 483–485. [Google Scholar]
- Wilson, TW; Neuendorff, DA; Lewis, AW; Randel, RD. Effect of zeranol or melengestrol acetate (MGA) on testicular and antler development and aggression in farmed fallow bucks. J. Anim. Sci 2002, 80, 1433–1441. [Google Scholar]
- Hussein, HS; Brasel, JM. Toxicity, metabolism and impact of mycotoxins on humans and animals. Toxicology 2001, 167, 101–134. [Google Scholar]
- Milano, GD; Odriozola, E; Lopèz, TA. Lack of effect of a diet containing zearalenone on spermatogenesis in rams. Vet. Rec 1991, 13, 33–34. [Google Scholar]
- Gimeno, A; Quintavilla, JA. Analytical and mycotoxicological study of a natural outbreak of zearalenone mycotoxicosis in horse. Proceeding of International Symposium on Mycotoxins, National Research Centre, Cairo, Egypt; 1983; pp. 387–392. [Google Scholar]
- Aurich, JE; Hoppen, HO; Trampler, R; Zentek, J; Boehm, J; Razzazi-Fazeli, E; Aurich, C. Effects of mycotoxins on reproductive function in mares. Anim. Reprod. Sci 2006, 94, 238–241. [Google Scholar]
- Juhàsz, J; Nagy, P; Kulcsàr, M; Szigeti, G; Reiczigel, J; Huszenicza, GY. Effect of low-dose zearalenone exposure on luteal function, follicular activity and uterine oedema in cycling mares. Acta Vet. Hung 2001, 49, 211–222. [Google Scholar]
- Powell, DG; Troppman, A; Tobin, T. Proceedings of the First Workshop on Mare Reproductive Loss Syndrome; Kentucky Agricultural Experimental Station, University of Kentucky, 2003.
- Alm, H; Greising, T; Brüssow, KP; Torner, H; Tiemann, U. The influence of the mycotoxins deoxynivalenol and zearalenol on in vitro maturation of pig oocytes and in vitro culture of pig zygotes. Toxicol. In Vitro 2002, 16, 643–648. [Google Scholar]
- Tiemann, U; Dänicke, S. In vivo and in vitro effects of the mycotoxins zearalenone and deoxynivalenol on different non-reproductive and reproductive organs in female pigs: A review. Food Addit. Contam 2007, 24, 306–314. [Google Scholar]
- Malekinejad, H; Schoevers, EJ; Daemen, IJJM; Zijlstra, C; Colenbrander, B; Fink-Gremmels, J; Roelen, BAJ. Exposure of oocyte to the Fusarium toxins zearalenone and deoxynivalenol causes aneuploidy and abnormal embryo development in pigs. Biol. Reprod 2007, 77, 840–847. [Google Scholar]
- Tiemann, U; Tomek, W; Schneider, F; Vancelow, J. Effects of the mycotoxins α- and β-zearalenol on regulation of progesterone synthesis in cultured granulose cells from porcine ovaries. Reprod. Toxicol 2003, 17, 673–681. [Google Scholar]
- Ranzenigo, G; Caloni, F; Cremonesi, F; Aad, PY; Spicer, LJ. Effects of Fusarium mycotoxins on steroid production by porcine granulosa cells. Anim. Reprod. Sci 2008, 107, 115–130. [Google Scholar]
- Minervini, F; Dell’Aquila, ME; Maritato, F; Minoia, P; Visconti, A. Toxic effects of the mycotoxin zearalenone and its derivatives on in vitro maturation of bovine oocytes and 17β-estradiol levels in mural granulosa cell cultures. Toxicol. In Vitro 2001, 15, 489–495. [Google Scholar]
- Minervini, F; Giannoccaro, A; Fornelli, F; Dell’Aquila, ME; Minoia, P; Visconti, A. Influence of mycotoxin zearalenone and its derivatives (alpha and beta-zearalenol) on apoptosis and proliferation of cultured granulosa cells from equine ovaries. Reprod. Biol. Endocrinol 2006, 4. doi:10.1186/1477-7827-4-62. [Google Scholar]
- Tsakmakidis, IA; Lymberopoulos, AG; Alexopoulos, C; Boscos, CM; Kyriakis, SC. In vitro effect of zearalenone and α-zearalenol on boar sperm characteristics and acrosome reaction. Reprod. Dom. Anim 2006, 41, 394–401. [Google Scholar]
- Tsakmakidis, IA; Lymberopoulos, AG; Khalifa, TAA; Boscos, CM; Saratsi, A; Alexopoulos, C. Evaluation of zearalenone and α-zearalenol toxicity on boar sperm DNA integrity. J. Appl. Toxicol 2007. DOI: 10.1002/jat.1322. [Google Scholar]
- Benzoni, E; Minervini, F; Giannoccaro, A; Fornelli, F; Vigo, D; Visconti, A. Influence of in vitro exposure to mycotoxin zearalenone and its derivatives on swine sperm quality. Reprod. Toxicol 2008, 25, 461–467. [Google Scholar]
- Rajkovic, A; Uyttendaele, M; Debevere, J. Computer aided boar semen motility analysis for cereulide detection in different food matrices. Int. J. Food. Microb 2007, 114, 92–99. [Google Scholar]
- D’Occhio, MJ; Hengstberger, KJ; Johnston, SD. Biology of sperm chromatin structure and relationship to male fertility and embryonic survival. Anim. Reprod. Sci 2007, 101, 1–17. [Google Scholar]
- Filannino, A; Giannoccaro, A; Nicassio, M; Lacalandra, GM; Dell’Aquila, ME; Visconti, A; Minervini, F. Influence of Fusarium mycotoxins on stallion spermatozoa viability and chromatin structure stability after in vitro exposure. Proceedings of the 12st European Society for Domestic Animal Reproduction, Utrecht University, The Netherlands, 20–23, November 2008.
- Love, CC; Thompson, JA; Lowry, VK; Varner, DD. Effect of storage time and temperature on stallion sperm DNA and fertility. Theriogenology 2002, 57, 1135–1142. [Google Scholar]



{kind=link}
{kind=link}
| Sow | ||||
|---|---|---|---|---|
| Target | Oocyte | Granulosa cells | Hormonal status | Ovary |
| In vivo | Oocytes with degenerated meiotic chromatin [27] | No modification of progesterone synthesis [27]
↓ FSH [24] | Stimulation of primary and secondary follicles [28]
Ovarian follicle atresia [25,26] Delay of maturation of ovarian follicles during preovulatory stage [24] | |
| In vitro | Alteration of spindle formation during meiosis [39]
Delay of TI and MII oocyte stage and nuclear malformation [8] | ↓ FSH or foskolin- enhanced progesterone [41]
Modulation dose- related IGF-induced oestradiol, progesterone and of FSH-induced progesterone [41] | ||
| Boar | ||||
|---|---|---|---|---|
| Target | Hormonal status | Sperm motility | Sperm viability | Chromatin structure |
| In vivo | ↓ testosterone [8] | ↓ [28] | ||
| In vitro | ↓ [44,46] | ↓ [44,46] | ↓ [44,46] | |
© 2008 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/). This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Minervini, F.; Dell’Aquila, M.E. Zearalenone and Reproductive Function in Farm Animals. Int. J. Mol. Sci. 2008, 9, 2570-2584. https://doi.org/10.3390/ijms9122570
Minervini F, Dell’Aquila ME. Zearalenone and Reproductive Function in Farm Animals. International Journal of Molecular Sciences. 2008; 9(12):2570-2584. https://doi.org/10.3390/ijms9122570
Chicago/Turabian StyleMinervini, Fiorenza, and Maria Elena Dell’Aquila. 2008. "Zearalenone and Reproductive Function in Farm Animals" International Journal of Molecular Sciences 9, no. 12: 2570-2584. https://doi.org/10.3390/ijms9122570