Purification and Preliminary Crystallographic Analysis of a New Lys49-PLA2 from B. Jararacussu

Abstract
:1. Introduction
2. Materials and methods
2.1. Protein purification
2.2. Platelet aggregation assays
2.3. Crystallization experiments
2.4. X-ray data collection and processing
3. Results and discussion
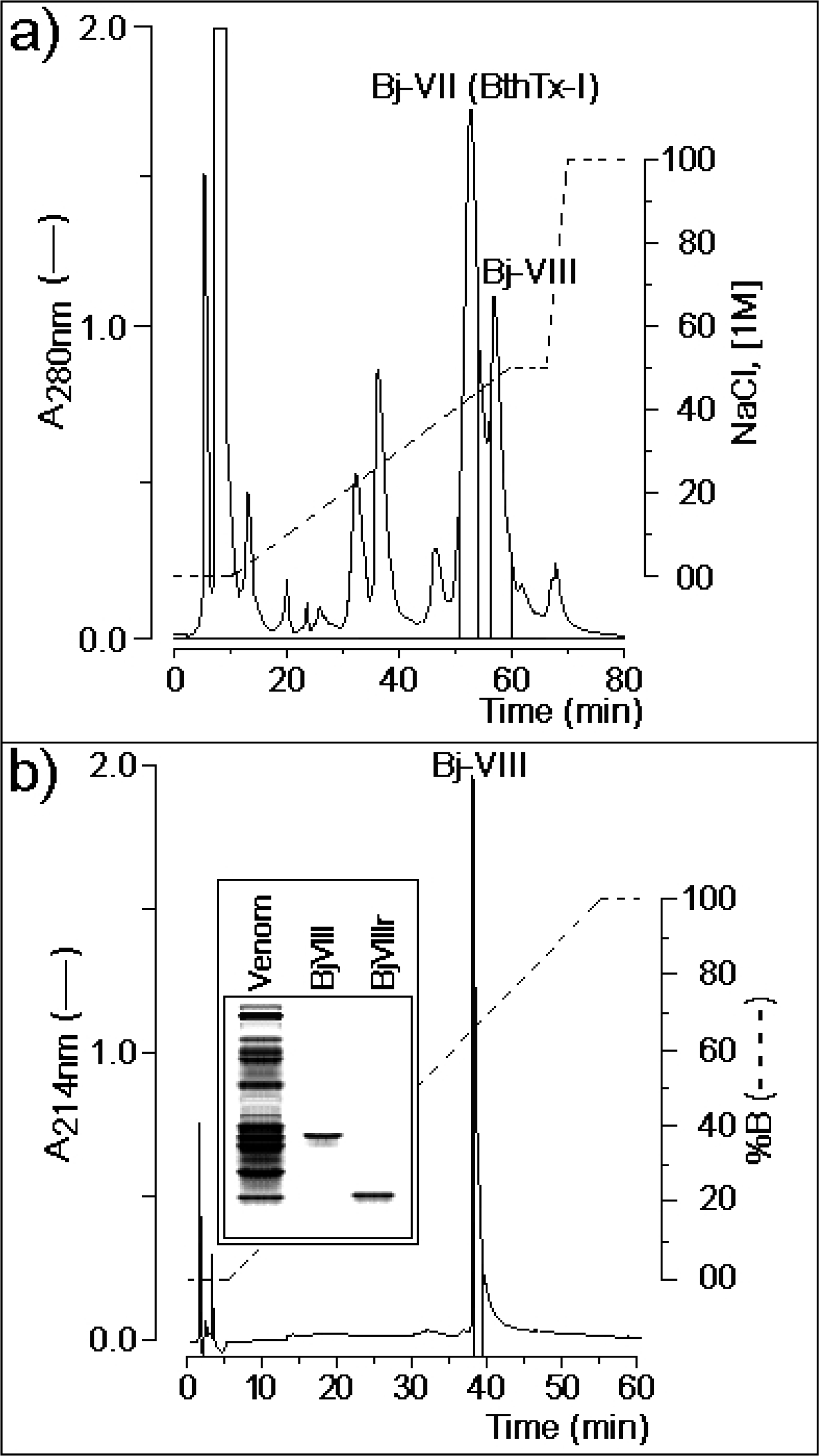
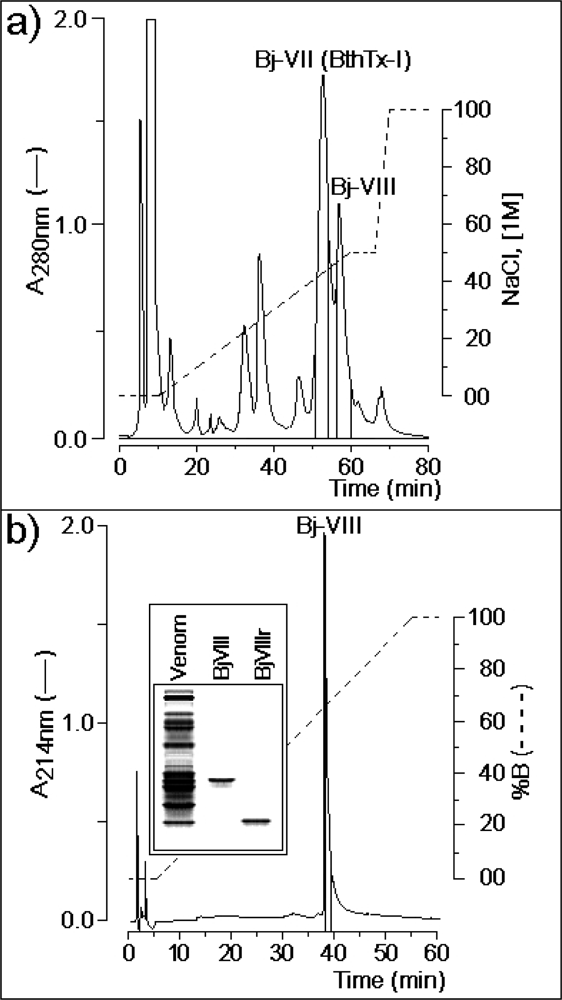
3.1. Purification of Lys49-PLA2 BjVIII
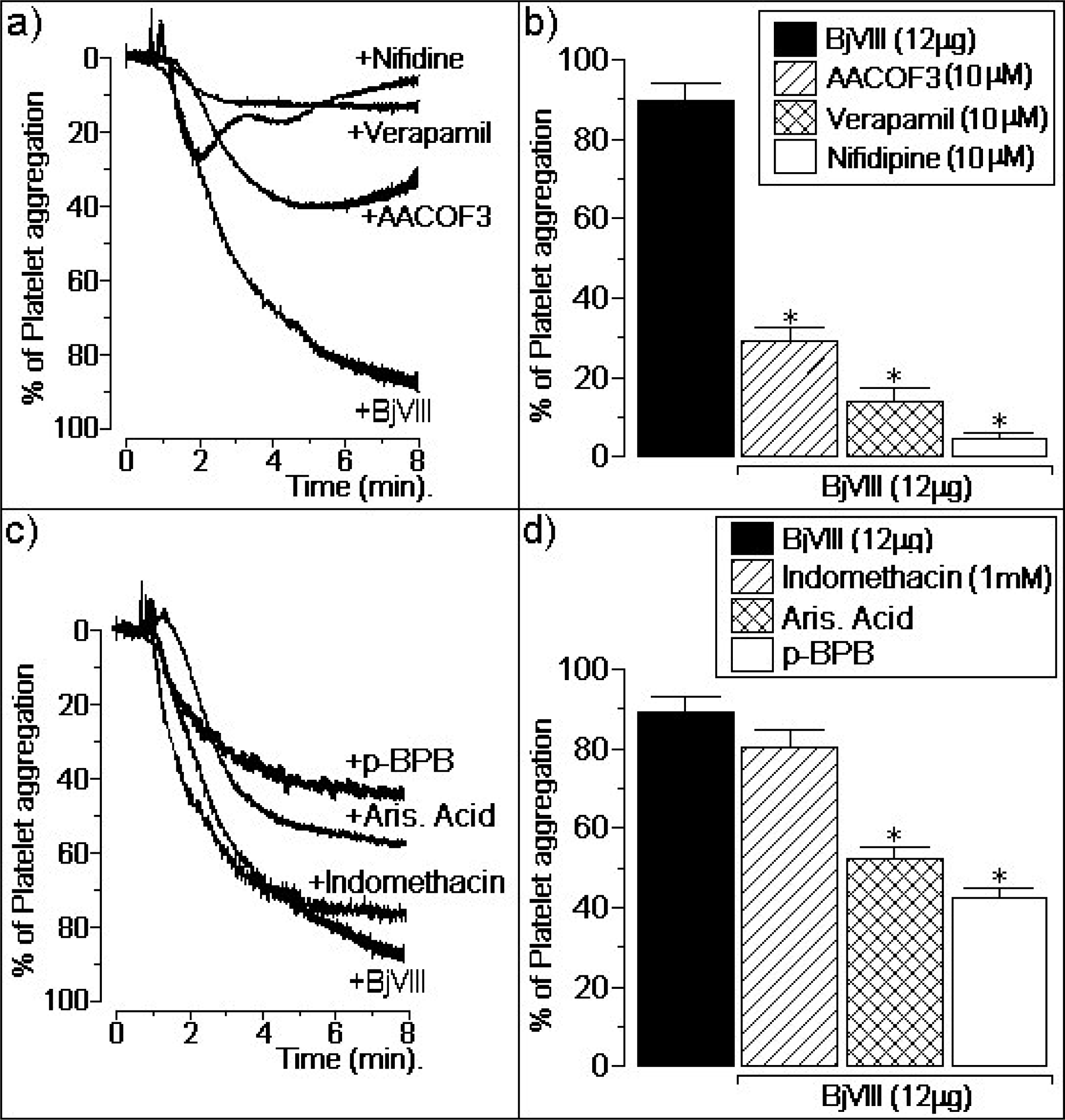
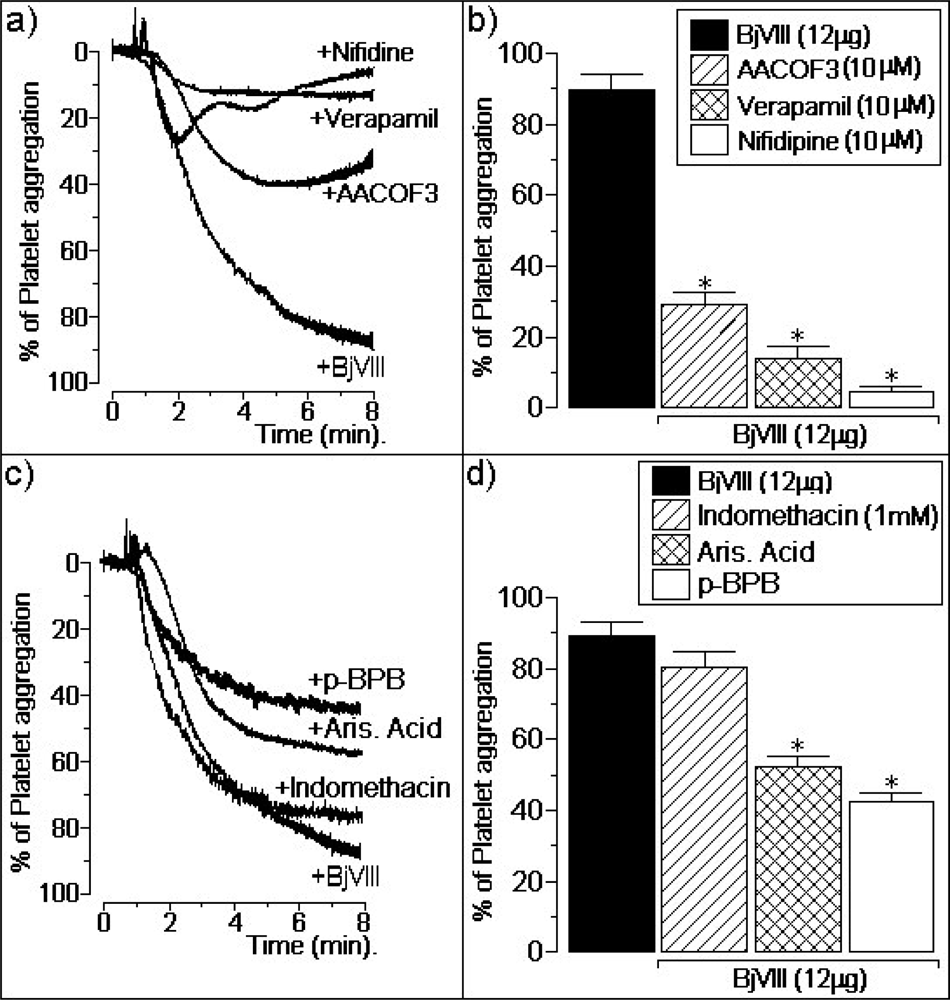
3.2. Atypical platelet aggregation activity of BjVIII
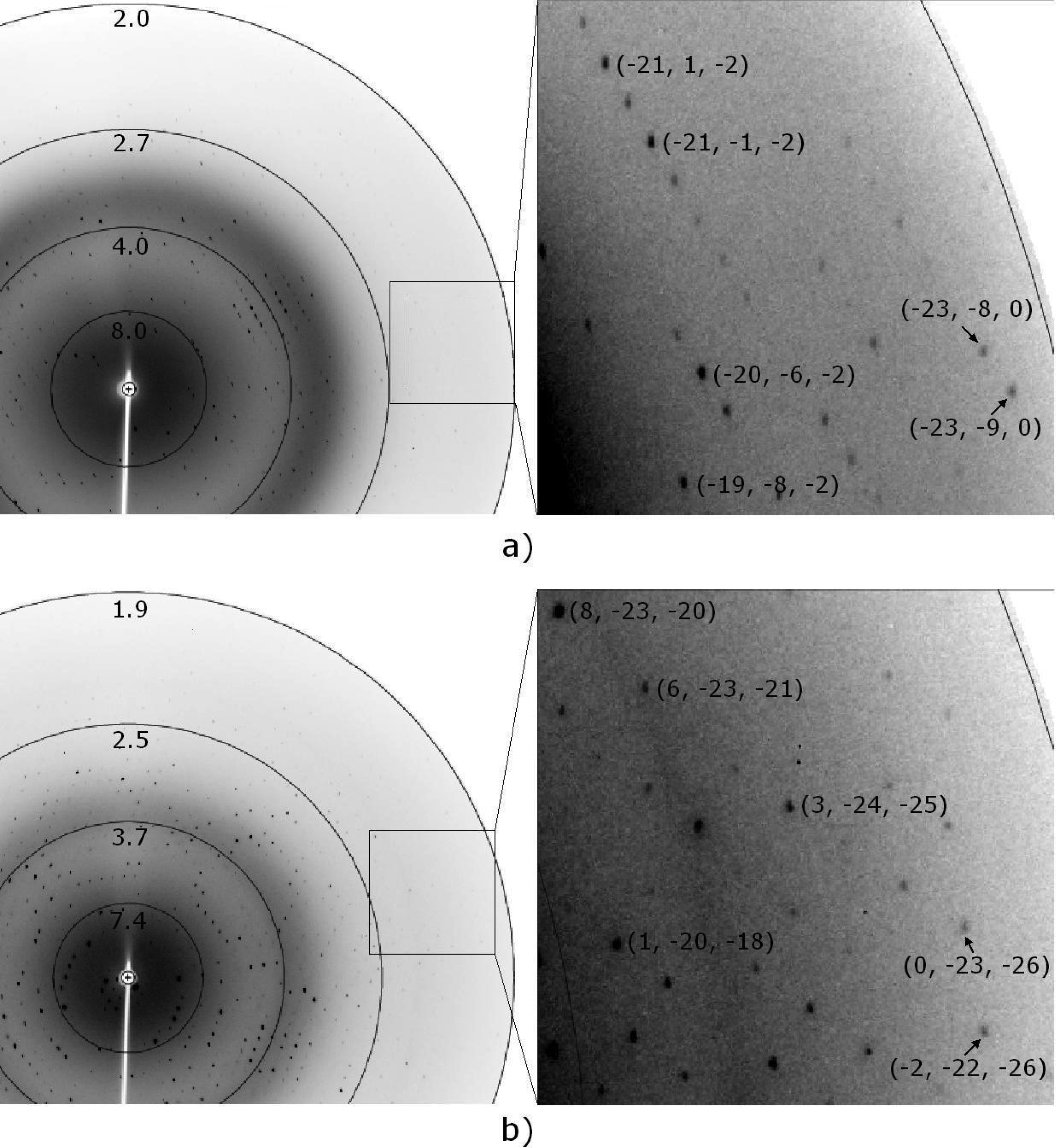
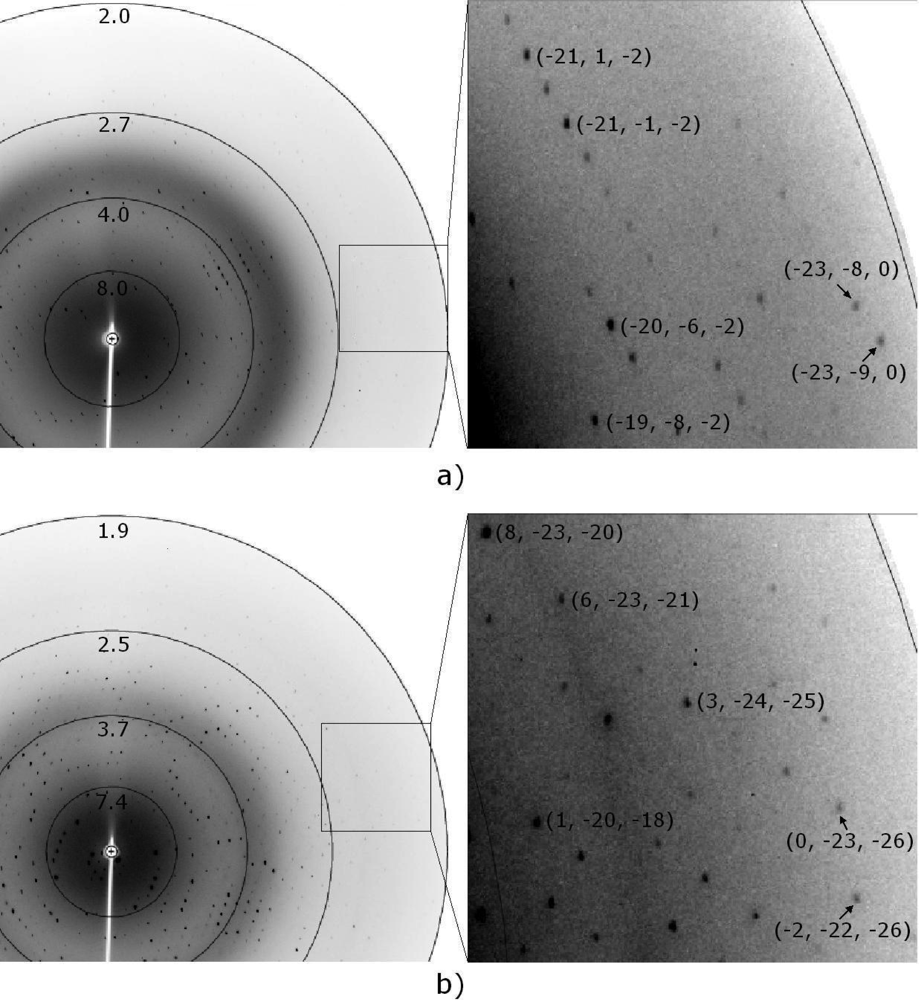
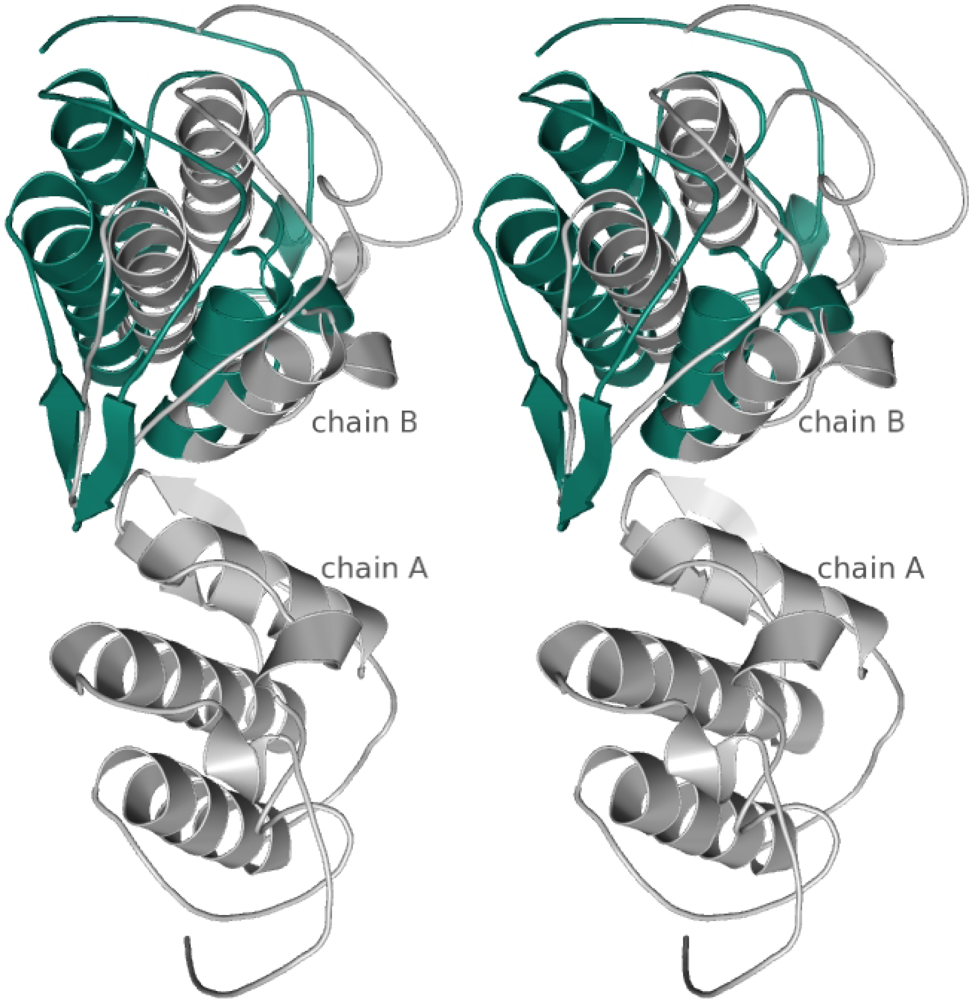
3.3. Preliminary X-ray diffraction analysis
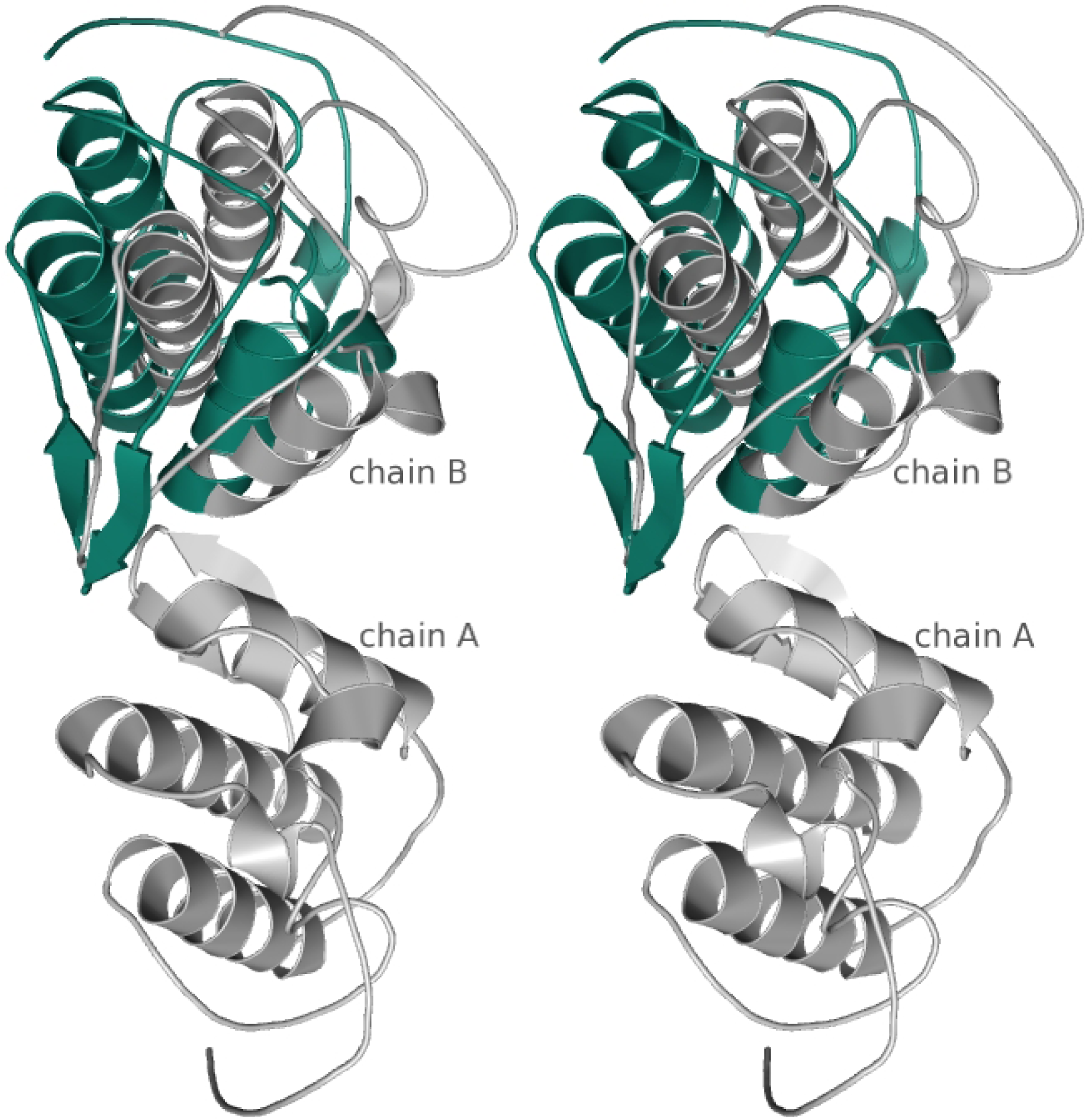
4. Concluding remarks
5. Acknowledgements
References
- van Deenen, L; de Haas, G; Heemskerk, CH. Hydrolysis of synthetic mixed-acid phosphatides by phospholipase A from human pancreas. Biochim Biophys Acta 1963, 67, 295–304. [Google Scholar]
- Higuchi, DA; Barbosa, CMV; Bincoletto, C; Chagas, JR; Magalhaes, A; Richardson, M; Sanchez, EF; Pesquero, JB; Araujo, RC; Pesquero, JL. Purification and partial characterization of two phospholipases A2 from Bothrops leucurus (white-tailed-jararaca) snake venom. Biochimie 2007, 89, 319–328. [Google Scholar]
- Maraganore, JM; Merutka, G; Cho, W; Welches, W; Kézdy, FJ; Heinrikson, RL. A new class of phospholipases A2 with lysine in place of aspartate 49. Functional consequences for calcium and substrate binding. J Biol Chem 1984, 259, 13839–13843. [Google Scholar]
- Nakai, M; Nakashima, KI; Ogawa, T; Shimohigashi, Y; Hattori, S; Chang, CC; Ohno, M. Purification and primary structure of a myotoxic lysine-49 phospholipase A2 with low lipolytic activity from Trimeresurus gramineus venom. Toxicon 1995, 33, 1469–1478. [Google Scholar]
- Koh, DCI; Armugam, A; Jeyaseelan, K. Snake venom components and their applications in biomedicine. Cell Mol Life Sci 2006, 63, 3030–3041. [Google Scholar]
- Andrião-Escarso, SH; Soares, AM; Fontes, MRM; Fuly, AL; Corrêa, FMA; Rosa, JC; Greene, LJ; Giglio, JR. Structural and functional characterization of an acidic platelet aggregation inhibitor and hypotensive phospholipase A(2) from Bothrops jararacussu snake venom. Biochem Pharmacol 2002, 64, 723–732. [Google Scholar]
- Fuly, AL; Soares, AM; Marcussi, S; Giglio, JR; Guimarães, JA. Signal transduction pathways involved in the platelet aggregation induced by a D-49 phospholipase A2 isolated from Bothrops jararacussu snake venom. Biochimie 2004, 86, 731–739. [Google Scholar]
- Magro, AJ; Murakami, MT; Marcussi, S; Soares, AM; Arni, RK; Fontes, MRM. Crystal structure of an acidic platelet aggregation inhibitor and hypotensive phospholipase A2 in the monomeric and dimeric states: insights into its oligomeric state. Biochem Biophys Res Commun 2004, 323, 24–31. [Google Scholar]
- Toyama, MH; Carneiro, EM; Marangoni, S; Barbosa, RL; Corso, G; Boschero, AC. Biochemical characterization of two crotamine isoforms isolated by a single step RP-HPLC from Crotalus durissus terrificus (South American rattlesnake) venom and their action on insulin secretion by pancreatic islets. Biochim Biophys Acta 2000, 1474, 56–60. [Google Scholar]
- Fonseca, FV; Antunes, E; Morganti, RP; Monteiro, HSA; Martins, AMC; Toyama, DO; Marangoni, S; Toyama, MH. Characterization of a new platelet aggregating factor from crotoxin Crotalus durissus cascavella venom. Protein J 2006, 25, 183–192. [Google Scholar]
- Schägger, H; von Jagow, G. Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal Biochem 1987, 166, 368–379. [Google Scholar]
- McPherson, A. Current approaches to macromolecular crystallization. Eur J Biochem 1990, 189, 1–23. [Google Scholar]
- Polikarpov, I; Oliva, G; Castellano, EE; Garratt, RC; Arruda, P; Leite, A; Craievich, A. The protein crystallography beamline at LNLS, the Brazilian National Synchrotron Light Source. Nucl Instrum Methods Phys Res A 1998, 405, 159–164. [Google Scholar]
- Polikarpov, I; Perles, LA; de Oliveira, RT; Oliva, G; Castellano, EE; Garratt, RC; Craievich, A. Set-up and Experimental Parameters of the Protein Crystallography Beamline at the Brazilian National Synchrotron Laboratory. J Synchrotron Radiat 1998, 5, 72–76. [Google Scholar]
- Leslie, AGW. The integration of macromolecular diffraction data. Acta Crystallogr D Biol Crystallogr 2006, 62, 48–57. [Google Scholar]
- Evans, P. Scaling and assessment of data quality. Acta Crystallogr D Biol Crystallogr 2006, 62, 72–82. [Google Scholar]
- Navaza, J. Implementation of molecular replacement in AMoRe. Acta Crystallogr D Biol Crystallogr 2001, 57, 1367–1372. [Google Scholar]
- Kozin, MB; Svergun, DI. Automated matching of high- and low-resolution structural models. J Appl Crystallogr 2001, 34, 33–41. [Google Scholar]
- CCP4. Collaborative Computational Project, Number 4. The CCP4 suite: programs for protein crystallography. Acta Crystallogr D Biol Crystallogr 1994, 50, 760–763.
- Winn, M; Dodson, EJ; Ralph, A. Collaborative computational project, number 4: Providing programs for protein crystallography. Methods Enzymol 1997, 277, 620–633. [Google Scholar]
- Spencer, PJ; Aird, SD; Boni-Mitake, M; Nascimento, N; Rogero, JR. A single-step purification of bothropstoxin-1. Braz J Med Biol Res 1998, 31, 1125–1127. [Google Scholar]
- Soares, AM; Guerra-Sá, R; Borja-Oliveira, CR; Rodrigues, VM; Rodrigues-Simioni, L; Rodrigues, V; Fontes, MRM; Lomonte, B; Gutiérrez, JM; Giglio, JR. Structural and functional characterization of BnSP-7, a Lys49 myotoxic phospholipase A2 homologue from Bothrops neuwiedi pauloensis venom. Arch Biochem Biophys 2000, 378, 201–209. [Google Scholar]
- Arni, RK; Ward, RJ; Gutiérrez, JM; Tulinsky, A. Structure of a calcium-independent phospholipase-like myotoxic protein from Bothrops asper venom. Acta Crystallogr D Biol Crystallogr 1995, 51, 311–317. [Google Scholar]
- Murakami, MT; Viçoti, MM; Abrego, JRB; Lourenzoni, MR; Cintra, ACO; Arruda, EZ; Tomaz, MA; Melo, PA; Arni, RK. Interfacial surface charge and free accessibility to the PLA2-active site-like region are essential requirements for the activity of Lys49 PLA2 homologues. Toxicon 2007, 49, 378–387. [Google Scholar]
- Toyama, MH; Soares, AM; Vieira, CA; Novello, JC; Oliveira, B; Giglio, JR; Marangoni, S. Amino acid sequence of piratoxin-I, a myotoxin from Bothrops pirajai snake venom, and its biological activity after alkylation with p-bromophenacyl bromide. J Protein Chem 1998, 17, 713–718. [Google Scholar]
- Barbosa, PSF; Martins, AMC; Alves, RS; Amora, DN; Martins, RD; Toyama, MH; Havt, A; Nascimento, NRF; Rocha, VLC; Menezes, DB; Fonteles, MC; Monteiro, HSA. The role of indomethacin and tezosentan on renal effects induced by Bothrops moojeni Lys49 myotoxin I. Toxicon 2006, 47, 831–837. [Google Scholar]
- Kramer, RM; Roberts, EF; Manetta, JV; Hyslop, PA; Jakubowski, JA. Thrombin-induced phosphorylation and activation of Ca(2+)-sensitive cytosolic phospholipase A2 in human platelets. J Biol Chem 1993, 268, 26796–26804. [Google Scholar]
- Puri, RN. Phospholipase A2: its role in ADP- and thrombin-induced platelet activation mechanisms. Int J Biochem Cell Biol 1998, 30, 1107–1122. [Google Scholar]
- Soares, AM; Andrião-Escarso, SH; Angulo, Y; Lomonte, B; Gutiérrez, JM; Marangoni, S; Toyama, MH; Arni, RK; Giglio, JR. Structural and functional characterization of myotoxin I, a Lys49 phospholipase A(2) homologue from Bothrops moojeni (Caissaca) snake venom. Arch Biochem Biophys 2000, 373, 7–15. [Google Scholar]
- Singh, N; Jabeen, T; Pal, A; Sharma, S; Perbandt, M; Betzel, C; Singh, TP. Crystal structures of the complexes of a group IIA phospholipase A2 with two natural anti-inflammatory agents, anisic acid, and atropine reveal a similar mode of binding. Proteins 2006, 64, 89–100. [Google Scholar]
- Kini, RM; Evans, HJ. A model to explain the pharmacological effects of snake venom phospholipases A2. Toxicon 1989, 27, 613–635. [Google Scholar]
- Gutiérrez, JM; Lomonte, B. Phospholipase A2 myotoxins from Bothrops snake venoms. Toxicon 1995, 33, 1405–1424. [Google Scholar]
- Ownby, CL; de Araujo, HSS; White, SP; Fletcher, JE. Lysine 49 phospholipase A2 proteins. Toxicon 1999, 37, 411–445. [Google Scholar]
- Yang, CC. In; Venom Phospholipase A2 enzymes: Structure, Function and Mechanism; Kini, RM, Ed.; John Wiley & Sons: UK, 1997; Chapter Chemical modification and functional sites of phospholipases A2, p 185. [Google Scholar]
- Stiles, BG; Choumet, V. In; Venom Phospholipase A2 enzymes: Structure, Function and Mechanism; Kini, RM, Ed.; John Wiley & Sons: UK, 1997; Chapter Antibodies studies with venom phospholipases A2, p 223. [Google Scholar]
- Matthews, BW. Solvent content of protein crystals. J Mol Biol 1968, 33, 491–497. [Google Scholar]
- Altschul, SF; Madden, TL; Schäffer, AA; Zhang, J; Zhang, Z; Miller, W; Lipman, DJ. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 1997, 25, 3389–3402. [Google Scholar]
- Magro, AJ; Soares, AM; Giglio, JR; Fontes, MRM. Crystal structures of BnSP-7 and BnSP-6, two Lys49-phospholipases A(2): quaternary structure and inhibition mechanism insights. Biochem Biophys Res Commun 2003, 311, 713–720. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crystal form | Orthorhombic | Trigonal |
|---|---|---|
| Wavelength used (Å) | 1.425 | 1.425 |
| Space group | P212121 | P3121 |
| Unit-cell parameters | ||
| a (Å) | 48.4 | 55.7 |
| b (Å) | 65.3 | 55.7 |
| c (Å) | 84.3 | 127.9 |
| Unit-cell volume (Å3) | 266666.7 | 344131.9 |
| VM (Å3 Da −1) | 2.44 | 2.10 |
| Solvent content (%) | 49.69 | 41.53 |
| ASU contents (molecules) | 2 | 2 |
| Resolution range (Å) | 84.2−2.0 (2.1−2.0) | 48.3−1.9 (2.0−1.9) |
| No. of images | 197 | 237 |
| No. of measured reflections | 136881 (14411) | 103843 (11729) |
| No. of unique reflections | 19116 (2226) | 20906 (2525) |
| Completeness (%) | 96.5 (78.6) | 97.2 (81.8) |
| Multiplicity | 7.2 (6.5) | 5.0 (4.6) |
| 〈I/σ(I)〉 | 20.0 (4.6) | 22.5 (6.4) |
| † (%) | 7.9 (35.4) | 4.9 (17.7) |
Share and Cite
Dos Santos, M.L.; Fagundes, F.H.R.; Teixeira, B.R.F.; Toyama, M.H.; Aparicio, R. Purification and Preliminary Crystallographic Analysis of a New Lys49-PLA2 from B. Jararacussu. Int. J. Mol. Sci. 2008, 9, 736-750. https://doi.org/10.3390/ijms9050736
Dos Santos ML, Fagundes FHR, Teixeira BRF, Toyama MH, Aparicio R. Purification and Preliminary Crystallographic Analysis of a New Lys49-PLA2 from B. Jararacussu. International Journal of Molecular Sciences. 2008; 9(5):736-750. https://doi.org/10.3390/ijms9050736
Chicago/Turabian StyleDos Santos, Marcelo L., Fábio H. R. Fagundes, Bruno R. F. Teixeira, Marcos H. Toyama, and Ricardo Aparicio. 2008. "Purification and Preliminary Crystallographic Analysis of a New Lys49-PLA2 from B. Jararacussu" International Journal of Molecular Sciences 9, no. 5: 736-750. https://doi.org/10.3390/ijms9050736