Training for Translocation: Predator Conditioning Induces Behavioral Plasticity and Physiological Changes in Captive Eastern Hellbenders (Cryptobranchus alleganiensis alleganiensis) (Cryptobranchidae, Amphibia)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Animals
2.2. Phase 1. Predator Conditioning
2.3. Phase 2. Exposure Trials
2.4. Statistical Analyses
2.4.1. Phase 1: Predator Conditioning
2.4.2. Phase 2: Exposure Trials
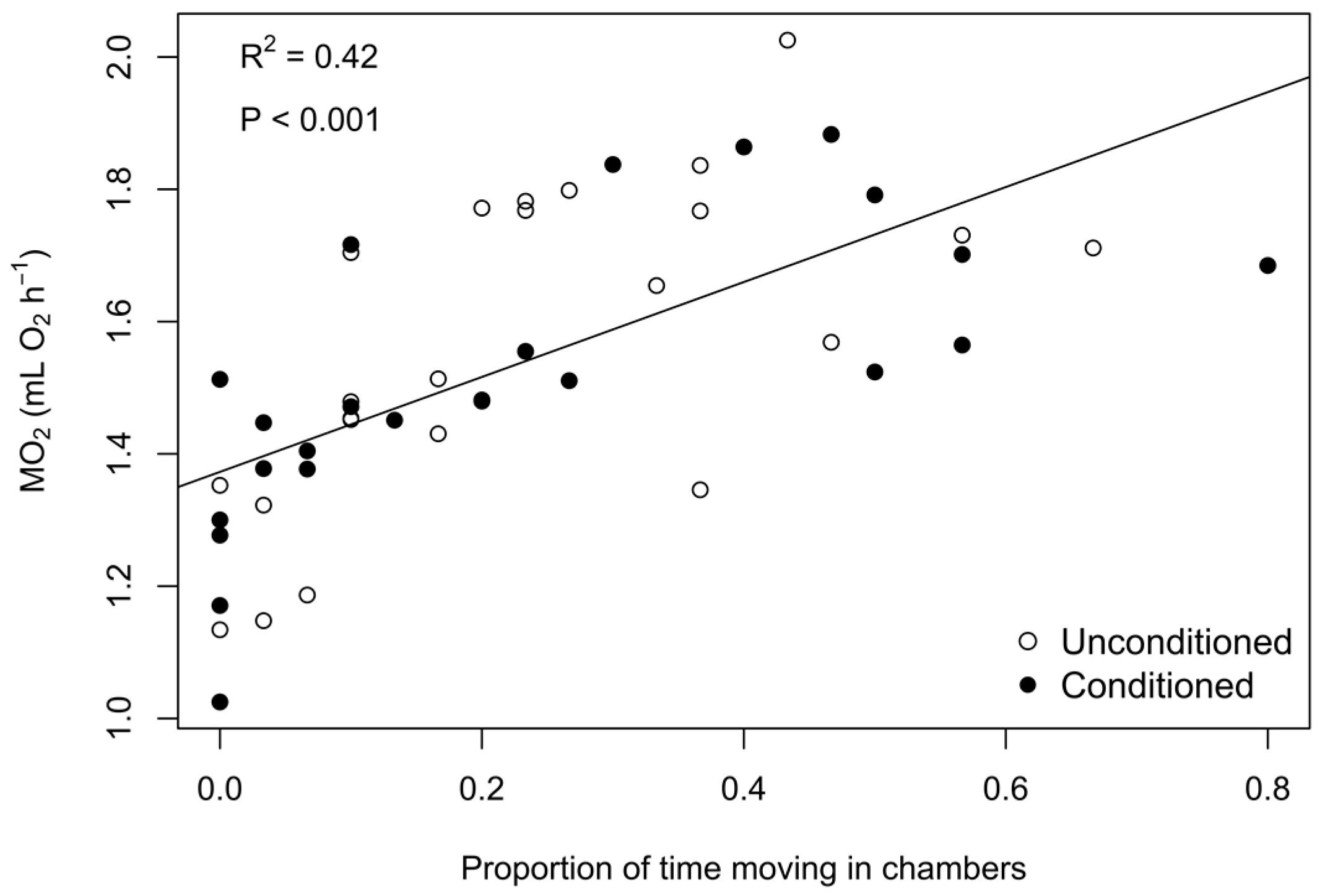
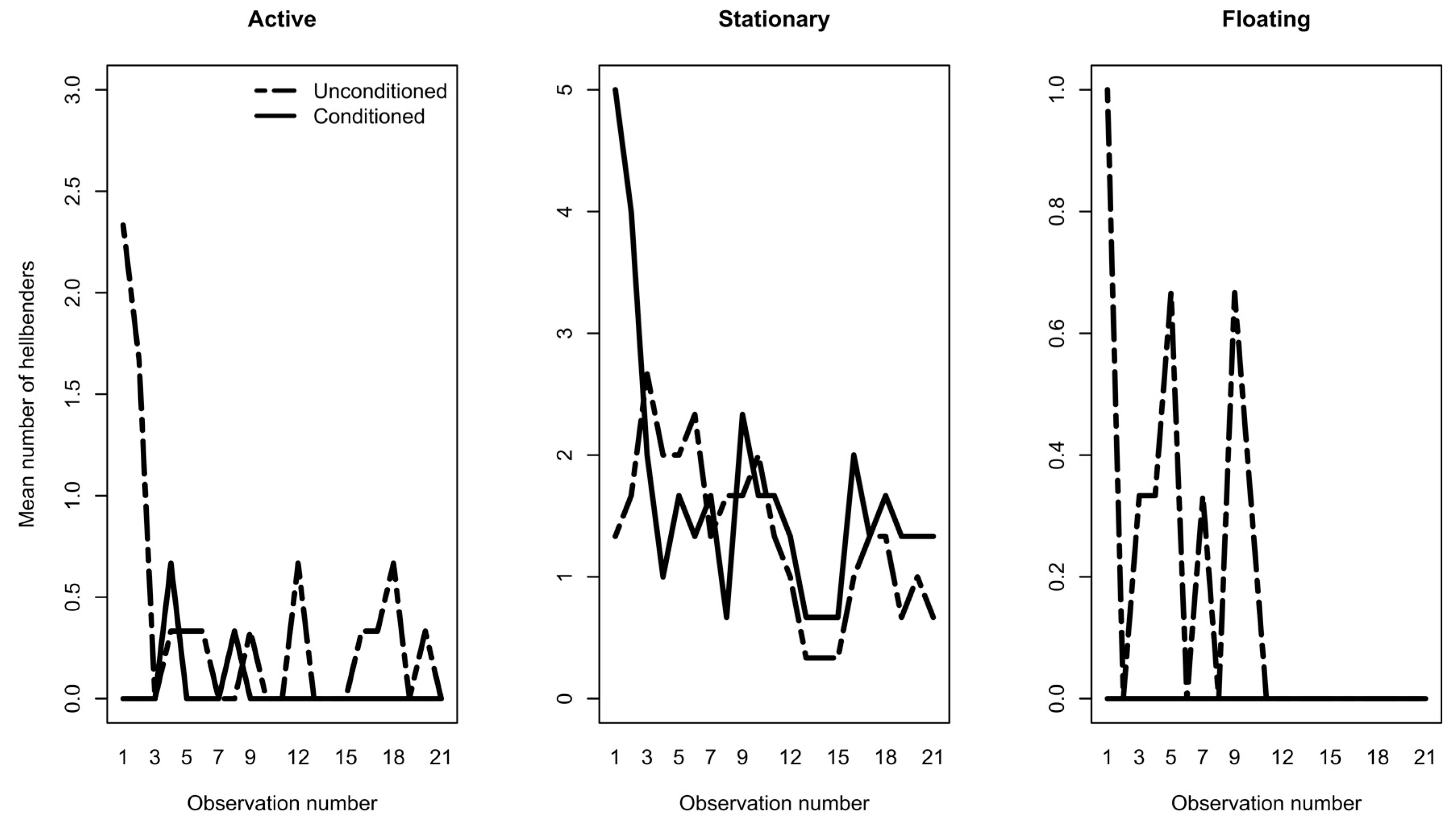
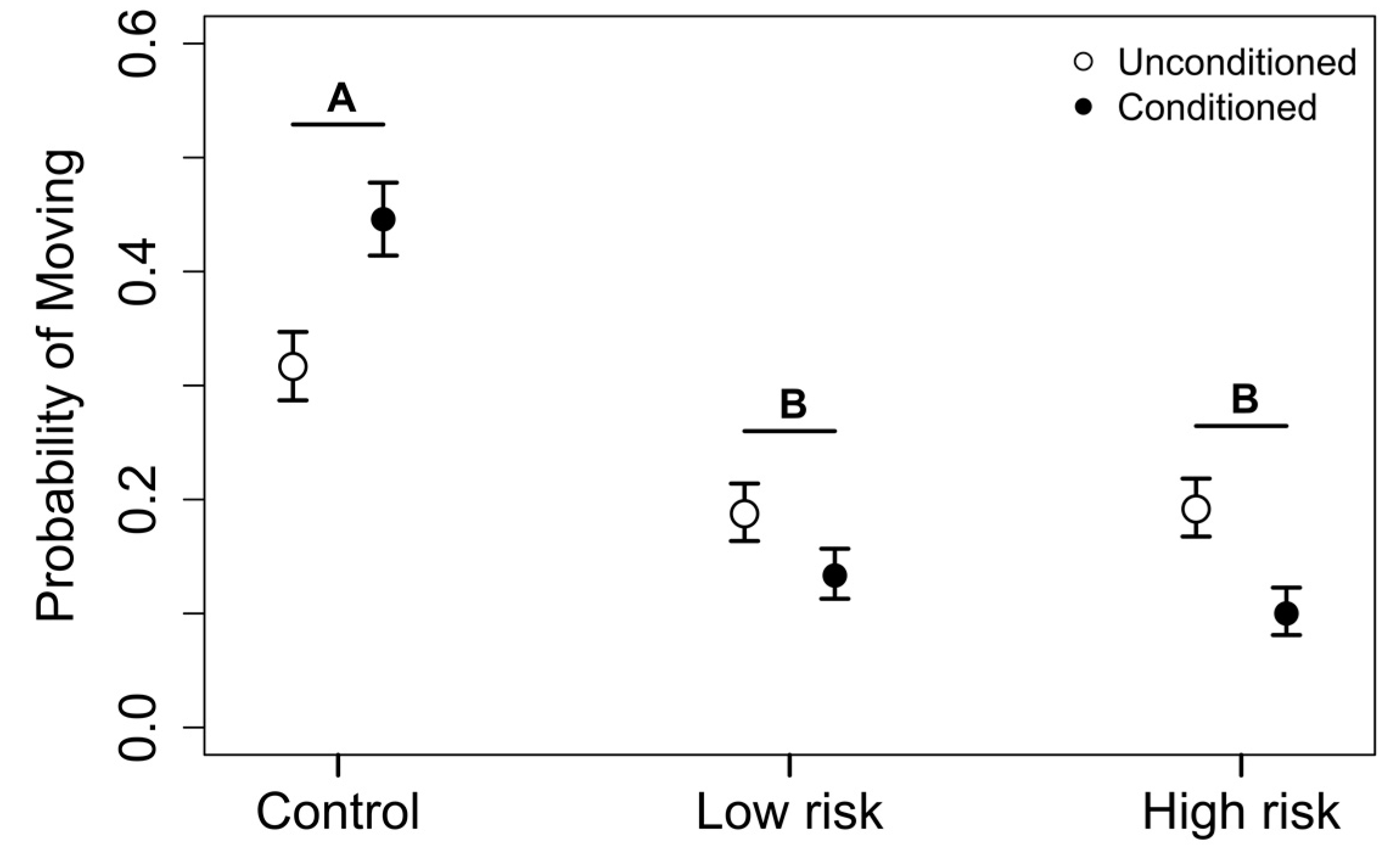
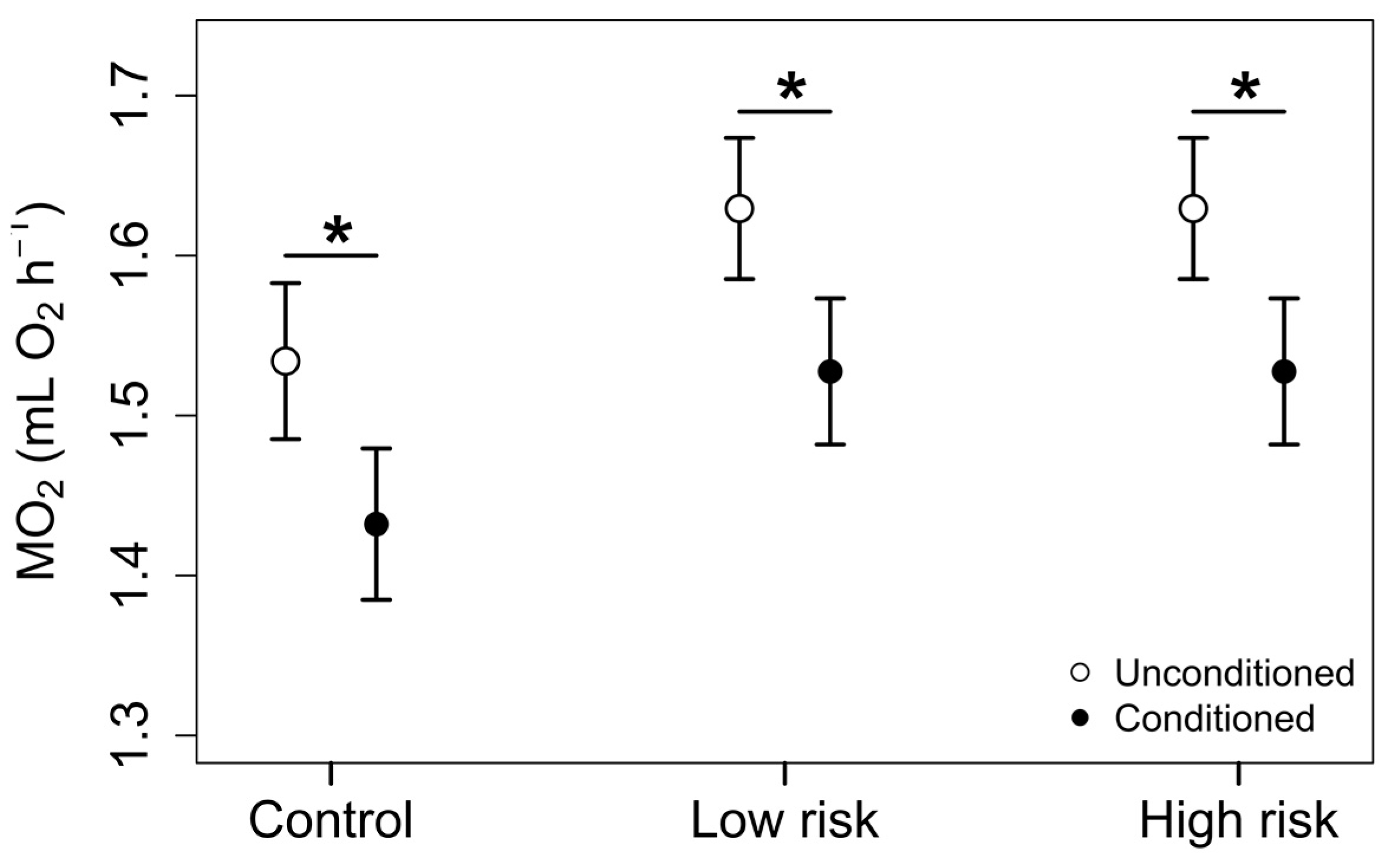
3. Results
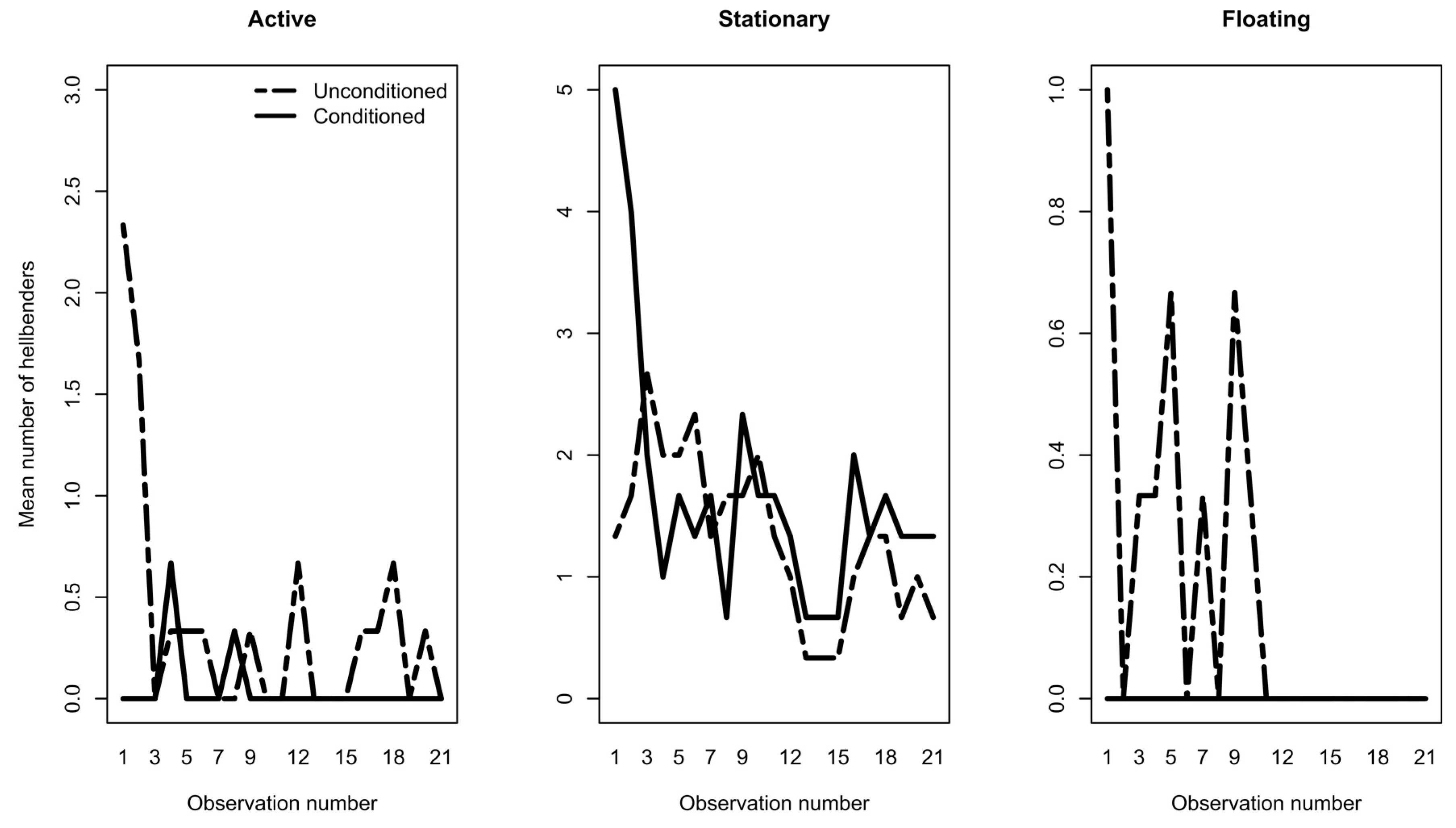
3.1. Phase 1: Predator Conditioning
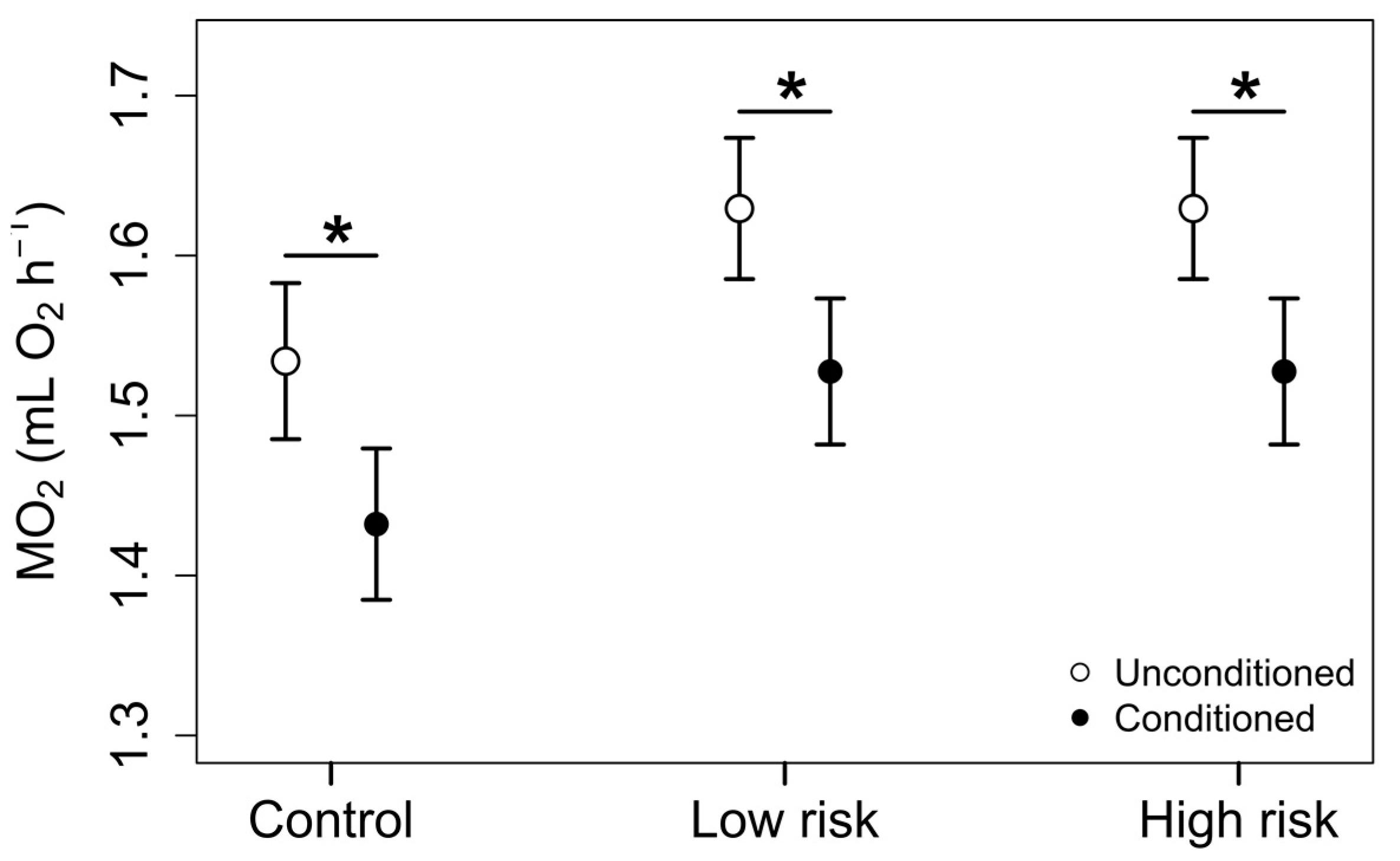
3.2. Phase 2: Exposure Trials
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Teixeira, C.P.; Schetini De Azevedo, C.; Mendl, M.; Cipreste, C.F.; Young, R.J. Revisiting translocation and reintroduction programmes : The importance of considering stress. Anim. Behav. 2007, 73, 1–13. [Google Scholar] [CrossRef]
- Dickens, M.J.; Delehanty, D.J.; Romero, L.M. Stress and translocation: Alterations in the stress physiology of translocated birds. Proc. R. Soc. Lond. B Biol. Sci. 2009, 276, 2051–2056. [Google Scholar] [CrossRef] [PubMed]
- Dickens, M.J.; Delehanty, D.J.; Michael Romero, L. Stress: An inevitable component of animal translocation. Biol. Conserv. 2010, 143, 1329–1341. [Google Scholar] [CrossRef]
- Engelsma, M.Y.; Hougee, S.; Nap, D.; Hofenk, M.; Rombout, J.H.W.M.; Van Muiswinkel, W.B.; Lidy Verburg-van Kemenade, B.M. Multiple acute temperature stress affects leucocyte populations and antibody responses in common carp, Cyprinus carpio L. Fish Shellfish Immunol. 2003, 15, 397–410. [Google Scholar] [CrossRef]
- Breves, J.P.; Specker, J.L. Cortisol stress response of juvenile winter flounder (Pseudopleuronectes americanus, Walbaum) to predators. J. Exp. Mar. Biol. Ecol. 2005, 325, 1–7. [Google Scholar] [CrossRef]
- Poledník, L.; Řehulka, J.; Kranz, A.; Poledníková, K.; Hlaváč, V.; Kazihnitkoví, H. Physiological responses of over-wintering common carp (Cyprinus carpio) to disturbance by Eurasian otter (Lutra lutra). Fish Physiol. Biochem. 2008, 34, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Killen, S.S.; Brown, J.A. Energetic cost of reduced foraging under predation threat in newly hatched ocean pout. Mar. Ecol. Prog. Ser. 2006, 321, 255–266. [Google Scholar] [CrossRef] [Green Version]
- Van Dievel, M.; Janssens, L.; Stoks, R. Short- and long-term behavioural, physiological and stoichiometric responses to predation risk indicate chronic stress and compensatory mechanisms. Oecologia 2016, 181, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Aarts, L.; Van Schagen, I. Driving speed and the risk of road crashes: A review. Accid. Anal. Prev. 2006, 38, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Chipman, R.; Slate, D.; Rupprecht, C.; Mendoza, M. Downside risk of wildlife translocation. Dev. Biol. (Basel) 2008, 131, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Crane, A.L.; Mathis, A. Predator-recognition training: A conservation strategy to increase postrelease survival of hellbenders in head-starting programs. Zoo Biol. 2011, 30, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Kellison, G.T.; Eggleston, D.B.; Burke, J.S. Comparative behaviour and survival of hatchery-reared versus wild summer flounder (Paralichthys dentatus). Can. J. Fish. Aquat. Sci. 2000, 57, 1870–1877. [Google Scholar] [CrossRef]
- Parish, D.M.B.; Sotherton, N.W. The fate of released captive-reared grey partridges Perdix perdix: Implications for reintroduction programmes. Wildl. Biol. 2007, 13, 140–149. [Google Scholar] [CrossRef]
- Pfeiffer, W. The fright reaction of fish. Biol. Rev. 1962, 37, 495–511. [Google Scholar] [CrossRef] [PubMed]
- Werner, E.E.; Anholt, B.R. Predator-induced behavioral indirect effects: Consequences to competitive interactions in anuran larvae. Ecology 1996, 77, 157–169. [Google Scholar] [CrossRef]
- Fraser, D.F.; Gilliam, J.F. Nonlethal impacts of predator invasion: Facultative suppression of growth and reproduction. Ecology 1992, 73, 959–970. [Google Scholar] [CrossRef]
- Sapolsky, R.M. Endocrinology of the stress-response. In Behavioral Endocrinology; Becker, J.B., Breedlove, S.M., Crews, D., McCarthy, M.M., Eds.; MIT Press: Cambridge, UK, 2002; pp. 409–450. [Google Scholar]
- Slos, S.; Stoks, R. Predation risk induces stress proteins and reduces antioxidant defense. Funct. Ecol. 2008, 22, 637–642. [Google Scholar] [CrossRef]
- Steiner, U.K.; Van Buskirk, J. Predator-induced changes in metabolism cannot explain the growth/predation risk tradeoff. PLoS ONE 2009, 4, e6160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lankford, S.E.; Adams, T.E.; Miller, R.A.; Cech, J.J., Jr. The cost of chronic stress: Impacts of a nonhabituating stress response on metabolic variables and swimming performance in sturgeon. Physiol. Biochem. Zool. 2005, 78, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, H.F.; Bodie, B.L.; Tauchi, M.; Dolgas, C.M.; Herman, J.P. Stress integration after acute and chronic predator stress: Differential activation of central stress circuitry and sensitization of the hypothalamo-pituitary-adrenocortical axis. Endocrinology 2003, 144, 5249–5258. [Google Scholar] [CrossRef] [PubMed]
- Petersson, E.; Valencia, A.C.; Järvi, T. Failure of predator conditioning: An experimental study of predator avoidance in brown trout (Salmo trutta). Ecol. Freshw. Fish 2015, 24, 329–337. [Google Scholar] [CrossRef]
- Chivers, D.P.; Mirza, R.S. Importance of predator diet cues in responses of larval wood frogs to fish and invertebrate predators. J. Chem. Ecol. 2001, 27, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.; Gardner, C.; Braithwaite, V.A. Differential stress responses in fish from areas of high- and low-predation pressure. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2005, 175, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Woodward, J.J.; Smith, L.S. Exercise training and the stress response in rainbow trout, Salmo gairdneri Richardson. J. Fish Biol. 1985, 26, 435–447. [Google Scholar] [CrossRef]
- Young, P.S.; Cech, J.J.J. Effects of exercise conditioning on stress response and recovery in cultured and wild young-of-the-year striped bass, Morone saxatilis. Can. J. Fish. Aquat. Sci. 1993, 50, 2094–2099. [Google Scholar] [CrossRef]
- Bodinof, C.M.; Briggler, J.T.; Junge, R.E.; Mong, T.; Beringer, J.; Wanner, M.D.; Schuette, C.D.; Ettling, J.; Millspaugh, J.J. Survival and body condition of captive-reared juvenile Ozark hellbenders (Cryptobranchus alleganiensis bishopi) following translocation to the wild. Copeia 2012, 2012, 150–159. [Google Scholar] [CrossRef]
- Boerner, J.A. Comparison of Movement Patterns in Captive-Released Eastern Hellbenders (Cryptobranchus alleganiensis alleganiensis) Using Three Different Release Methods; State University of New York Buffalo: Buffalo, NY, USA, 2014. [Google Scholar]
- Kraus, B.T.; McCallen, E.B.; Williams, R.N. Evaluating the survival of translocated adult and captive-reared, juvenile eastern hellbenders (Cryptobranchus alleganiensis alleganiensis). Herpetologica 2017, 73, 271–276. [Google Scholar] [CrossRef]
- Carnahan, D.P. Blue River Fisheries Survey and Game Fish Population Estimates in Crawford, Harrison, and Washington Counties; Indiana Department of Natural Resources: Indianapolis, Indiana, 2001. [Google Scholar]
- Gall, B.G. Predator-Prey Interactions between Hellbenders (Cryptobranchus alleganiensis alleganiensis) and (CA. Bishopi) and Native and Nonnative Fishes. Master’s Thesis, Missouri State University, Springfield, MO, USA, 2008. [Google Scholar]
- Gall, B.G.; Mathis, A. Innate predator recognition and the problem of introduced trout. Ethology 2010, 116, 47–58. [Google Scholar] [CrossRef]
- Burgmeier, N.G.; Unger, S.D.; Sutton, T.M.; Williams, R.N. Population status of the eastern hellbender (Cryptobranchus alleganiensis alleganiensis) in Indiana. J. Herpetol. 2011, 45, 195–201. [Google Scholar] [CrossRef]
- Kats, L.B.; Petranka, J.W.; Sih, A. Antipredator defenses and the persistence of amphibian larvae with fishes. Ecology 1988, 69, 1865–1870. [Google Scholar] [CrossRef]
- Barbosa, P.; Castellanos, I. Ecology of Predator-Prey Interactions; Oxford University Press: Oxford, UK, 2005; ISBN 9780195171204. [Google Scholar]
- Ferland-Raymond, B.; Murray, D.L. Predator diet and prey adaptive responses: Can tadpoles distinguish between predators feeding on congeneric vs. conspecific prey? Can. J. Zool. 2008, 86, 1329–1336. [Google Scholar] [CrossRef]
- Schoeppner, N.M.; Relyea, R.A. When should prey respond to consumed heterospecifics? Testing hypotheses of perceived risk. Copeia 2009, 2009, 190–194. [Google Scholar] [CrossRef]
- Altmann, J. Observational study of behavior: Sampling methods. Behaviour 1974, 49, 227–267. [Google Scholar] [CrossRef] [PubMed]
- Maher, J.M.; Werner, E.E.; Denver, R.J. Stress hormones mediate predator-induced phenotypic plasticity in amphibian tadpoles. Proc. R. Soc. B Biol. Sci. 2013, 280, 1–9. [Google Scholar] [CrossRef]
- Hall, A.E.; Clark, T.D. Seeing is believing: Metabolism provides insight into threat perception for a prey species of coral reef fish. Anim. Behav. 2016, 115, 117–126. [Google Scholar] [CrossRef]
- Burraco, P.; Duarte, L.J.; Gomez-Mestre, I. Predator-induced physiological responses in tadpoles challenged with herbicide pollution. Curr. Zool. 2013, 59, 475–484. [Google Scholar] [CrossRef]
- Alvarez, D.; Cano, J.M.; Nicieza, A.G. Microgeographic variation in metabolic rate and energy storage of brown trout: Countergradient selection or thermal sensitivity? Evol. Ecol. 2006, 20, 345–363. [Google Scholar] [CrossRef]
- Relyea, R.A. Fine-tuned phenotypes: Tadpole plasticity under 16 combinations of predators and competitors. Ecology 2004, 85, 172–179. [Google Scholar] [CrossRef]
- Orlofske, S.A.; Hopkins, W.A. Energetics of metamorphic climax in the pickerel frog (Lithobates palustris). Comp. Biochem. Physiol. Part A 2009, 154, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Kearney, B.D.; Byrne, P.G.; Reina, R.D. Short-and long-term consequences of developmental saline stress: Impacts on anuran respiration and behaviour. R. Soc. Open Sci. 2016, 3, 150640. [Google Scholar] [CrossRef] [PubMed]
- Srean, P.; Almeida, D.; Rubio-Gracia, F.; Luo, Y.; Garcia-Berthou, E. Effects of size and sex on swimming performance and metabolism of invasive mosquitofish Gambusia holbrooki. Ecol. Freshw. Fish 2017, 26, 424–433. [Google Scholar] [CrossRef]
- Guimond, R.W.; Hutchison, V.H. Aquatic respiration: An unusual strategy in the hellbender Cryptobranchus alleganiensis alleganiensis (Daudin). Science 1973, 182, 1263–1265. [Google Scholar] [CrossRef] [PubMed]
- Conover, W.J. Practical Nonparametric Statistics; Wiley: Hoboken, NJ, USA, 1999; ISBN 0471160687. [Google Scholar]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Norrdahl, K.; Korpimäki, E. Fear in farmlands: How much does predator avoidance affect bird community structure? J. Avian Biol. 1998, 29, 79–85. [Google Scholar] [CrossRef]
- Kenison, E.K.E.K.; Weldy, P.Y.P.Y.; Williams, R.N.R.N. There must be something in the water: Assessing the behavioral responses of rusty crayfish (Orconectes rusticus) to fish and amphibian predator kairomones. J. Ethol. 2017, 1–8. [Google Scholar] [CrossRef]
- Bleckmann, H.; Zelick, R. Lateral line system of fish. Integr. Zool. 2009, 4, 13–25. [Google Scholar] [CrossRef] [PubMed]
- McPeek, M.A. The growth/predation risk trade-off: So what is the mechanism? Am. Nat. 2004, 163, E88–E111. [Google Scholar] [CrossRef] [PubMed]
- Holopainen, I.J.; Aho, J.; Vornanen, M.; Huuskonen, H. Phenotypic plasticity and predator effects on morphology and physiology of crucian carp in nature and in the laboratory. J. Fish Biol. 1997, 50, 781–798. [Google Scholar] [CrossRef]
- Brönmark, C.; Miner, J.G. Predator-induced phenotypical change in body morphology in crucian carp. Science 1992, 258, 1348–1350. [Google Scholar] [CrossRef] [PubMed]
- Repka, S.; Walls, M.; Kettle, M. Neck spine protects Daphnia Pulex from predation by Chaoborus, but individuals with longer tail spine are at a greater risk. J. Plankton Res. 1995, 17, 393–403. [Google Scholar] [CrossRef]
- Hoverman, J.T.; Auld, J.R.; Relyea, R.A. Putting prey back together again: Integrating predator-induced behavior, morphology, and life history. Oecologia 2005, 144, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Haskell, A.; Graham, T.E.; Griffin, C.R.; Hestbeck, J.B. Size related survival of headstarted redbelly turtles (Pseudemys rubriventris) in Massachusetts. J. Herpetol. 1996, 30, 524–527. [Google Scholar] [CrossRef]
- Kats, L.B.; Dill, L.M. The scent of death: Chemosensory assessment of predation risk by prey animals. Écoscience 1998, 5, 361–394. [Google Scholar] [CrossRef]
- Epp, K.J.; Gabor, C.R. Innate and learned predator recognition mediated by chemical signals in Eurycea nana. Ethology 2008, 114, 607–615. [Google Scholar] [CrossRef]
- Schoeppner, N.M.; Relyea, R.A. Damage, digestion, and defence: The roles of alarm cues and kairomones for inducing prey defences. Ecol. Lett. 2005, 8, 505–512. [Google Scholar] [CrossRef] [PubMed]
- DeWitt, T.J.; Sih, A.; Wilson, D.S. Cost and limits of phenotypic plasticity. Trends Ecol. Evol. 1998, 13, 77–81. [Google Scholar] [CrossRef]
- Van Buskirk, J.; Relyea, R.A. Selection for phenotypic plasticity in Rana sylvatica tadpoles. Biol. J. Linn. Soc. 1998, 65, 301–328. [Google Scholar] [CrossRef]
- Schoeppner, N.M.; Relyea, R.A. Interpreting the smells of predation: How alarm cues and kairomones induce different prey defences. Funct. Ecol. 2009, 23, 1114–1121. [Google Scholar] [CrossRef]
- Schoeppner, N.M.; Relyea, R.A. Detecting small environmental differences: Risk-response curves for predator-induced behavior and morphology. Oecologia 2008, 154, 743–754. [Google Scholar] [CrossRef] [PubMed]
- Van Buskirk, J.; Muller, C.; Portmann, A.; Surbeck, M. A test of the risk allocation hypothesis: Tadpole responses to temporal change in predation risk. Behav. Ecol. 2002, 13, 526–530. [Google Scholar] [CrossRef]
- Lima, S.L.; Bednekoff, P.A. Temporal variation in danger drives antipredator behavior: The predation risk allocation hypothesis. Am. Nat. 1999, 153, 649–659. [Google Scholar] [CrossRef]
- Sih, A.; McCarthy, T.M. Prey responses to pulses of risk and safety: Testing the risk allocation hypothesis. Anim. Behav. 2002, 63, 437–443. [Google Scholar] [CrossRef]
- Barry, M.J.; Syal, S. Metabolic responses of tadpoles to chemical predation cues. Hydrobiologia 2013, 700, 267–276. [Google Scholar] [CrossRef]
- Reed, D.H.; Lowe, E.H.; Briscoe, D.A.; Frankham, R. Fitness and adaptation in a novel environment: Effect of inbreeding, prior environment, and lineage. Evolution (N. Y.) 2003, 57, 1822–1828. [Google Scholar] [CrossRef]
- Handelsman, C.A.; Broder, E.D.; Dalton, C.M.; Ruell, E.W.; Myrick, C.A.; Reznick, D.N.; Ghalambor, C.K. Predator-induced phenotypic plasticity in metabolism and rate of growth: Rapid adaptation to a novel environment. Integr. Comp. Biol. 2013, 53, 975–988. [Google Scholar] [CrossRef] [PubMed]
- D’Anna, G.; Giacalone, V.M.; Vega Fernández, T.; Vaccaro, A.M.; Pipitone, C.; Mirto, S.; Mazzola, S.; Badalamenti, F. Effects of predator and shelter conditioning on hatchery-reared white seabream Diplodus sargus (L., 1758) released at sea. Aquaculture 2012, 356–357, 91–97. [Google Scholar] [CrossRef]
- Mirza, R.S.; Chivers, D.P. Predator-recognition training enhances survival of brook trout: Evidence from laboratory and field- enclosure studies. Can. J. Zool. 2000, 2208, 2198–2208. [Google Scholar] [CrossRef]
- Griffin, A.S.; Blumstein, D.T.; Evans, C.S. Review: Training captive-bred or translocated animals to avoid predators. Conserv. Biol. 2000, 14, 1317–1326. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MANOVA | Df | Pillai | f Value | p Value |
|---|---|---|---|---|
| Treatment | 1, 122 | 0.128 | 5.88 | <0.001 |
| Date | 1, 122 | 0.318 | 18.61 | <0.001 |
| Treatment*Date | 1, 122 | 0.087 | 3.81 | 0.012 |
| REPEATED MEASURES ANOVA | Active | Stationary | Floating | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Df | f value | p value | Df | f value | p value | Df | f value | p value | |
| Treatment | 1 | 9.43 | 0.003 | 1 | 3.70 | 0.057 | 1 | 11.21 | 0.001 |
| Treatment*Date | 1 | 5.83 | 0.017 | 1 | 0.80 | 0.372 | 1 | 5.41 | 0.022 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kenison, E.K.; Williams, R.N. Training for Translocation: Predator Conditioning Induces Behavioral Plasticity and Physiological Changes in Captive Eastern Hellbenders (Cryptobranchus alleganiensis alleganiensis) (Cryptobranchidae, Amphibia). Diversity 2018, 10, 13. https://doi.org/10.3390/d10010013
Kenison EK, Williams RN. Training for Translocation: Predator Conditioning Induces Behavioral Plasticity and Physiological Changes in Captive Eastern Hellbenders (Cryptobranchus alleganiensis alleganiensis) (Cryptobranchidae, Amphibia). Diversity. 2018; 10(1):13. https://doi.org/10.3390/d10010013
Chicago/Turabian StyleKenison, Erin K., and Rod N. Williams. 2018. "Training for Translocation: Predator Conditioning Induces Behavioral Plasticity and Physiological Changes in Captive Eastern Hellbenders (Cryptobranchus alleganiensis alleganiensis) (Cryptobranchidae, Amphibia)" Diversity 10, no. 1: 13. https://doi.org/10.3390/d10010013