Distribution of the Genus Passiflora L. Diversity in Colombia and Its Potential as an Indicator for Biodiversity Management in the Coffee Growing Zone

Abstract
:
1. Introduction
2. Material and Methods
2.1.Geography and Climate
2.2. Species Distribution and Richness

2.3. Climatic Adaptation and Modeling
3. Results and Discussion
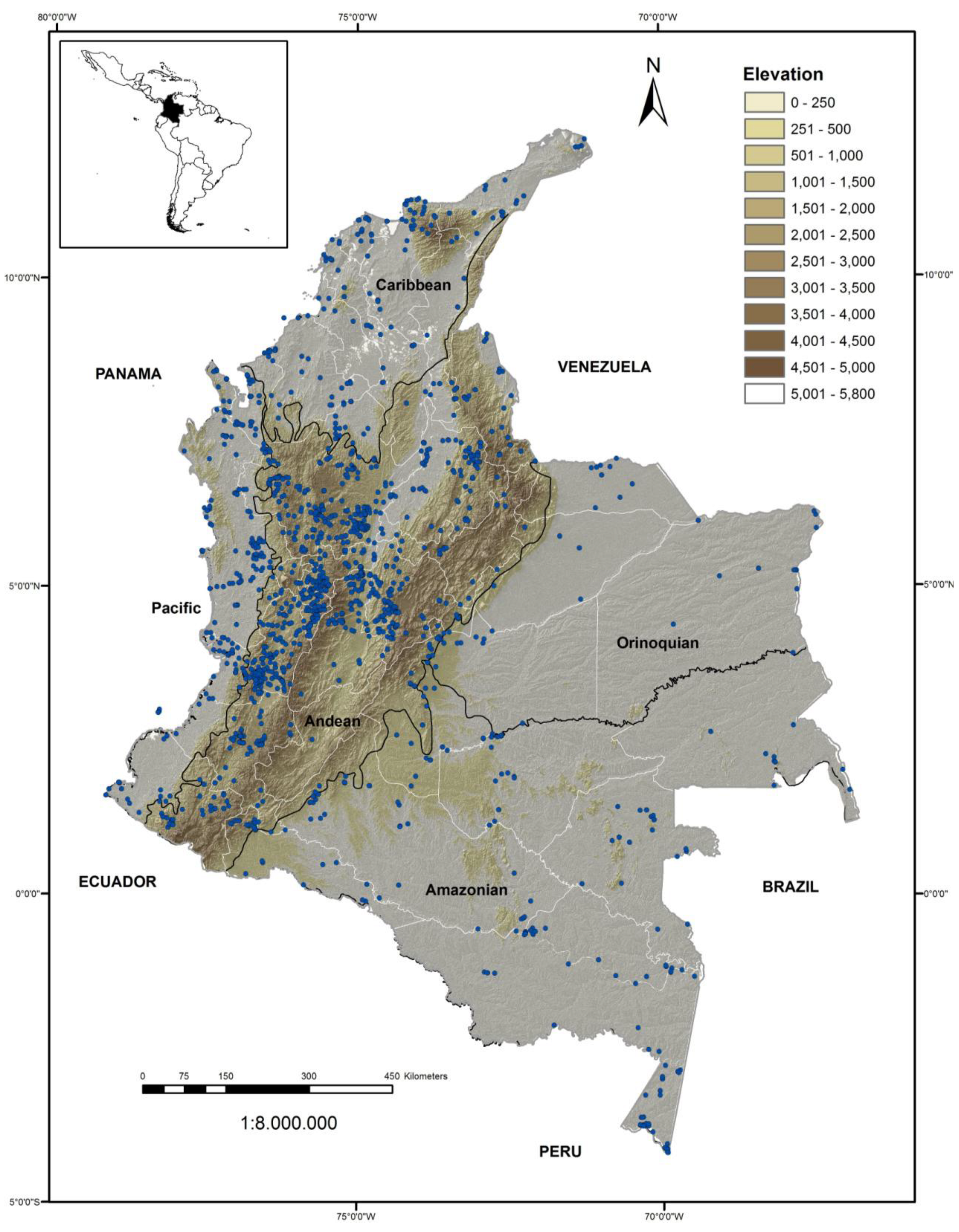
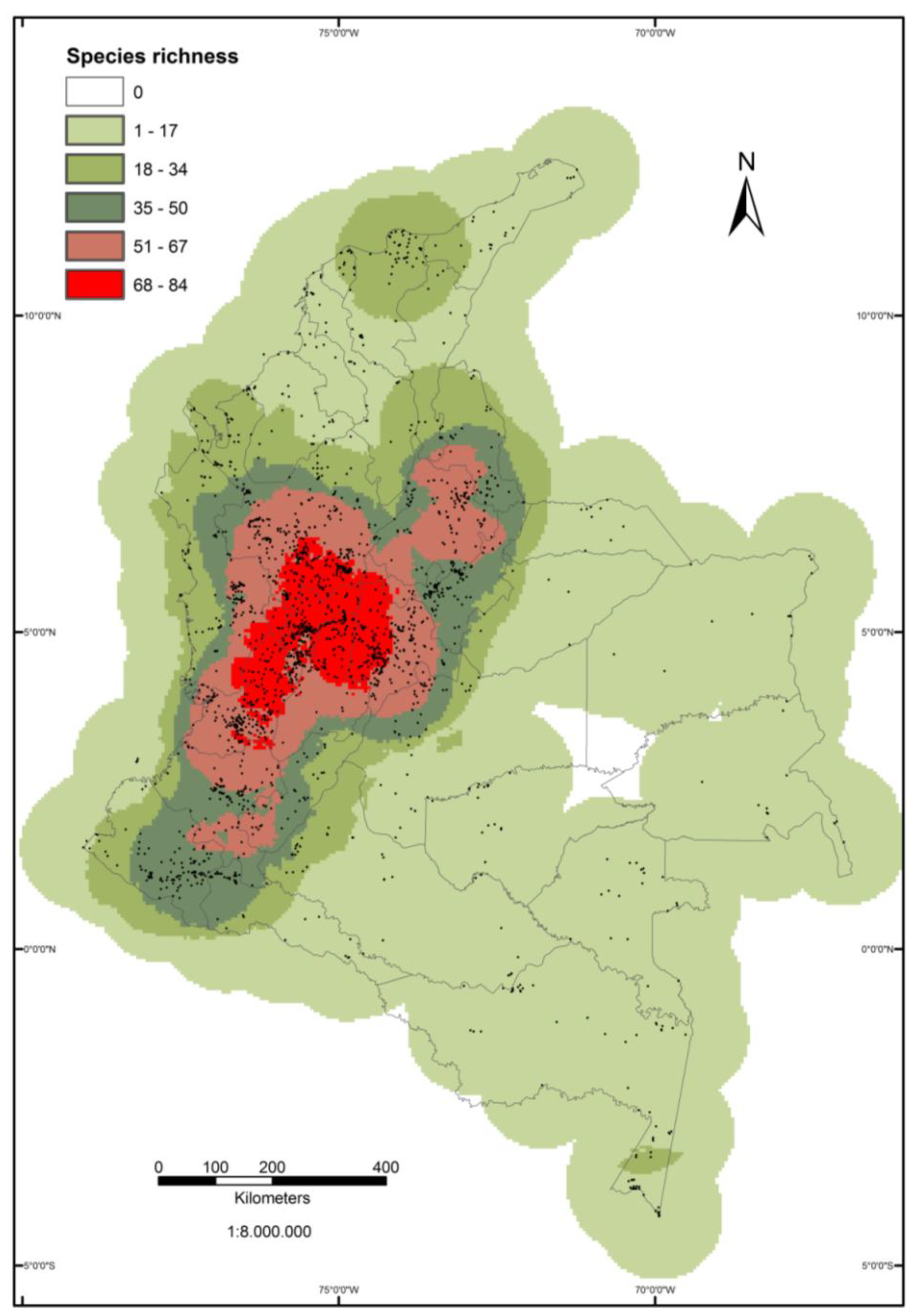
3.1. Distribution of Observations and Species Richness/Diversity


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biogeographic region/department | Area (km2) | Nb. observ. | Nb. observ./ 1,000 km2 | Total Species | Total species/ 1,000 km2 | Total species/ Log. area | Observ./ species | Rare species | Endemic species |
|---|---|---|---|---|---|---|---|---|---|
| Andean | |||||||||
| Antioquia | 62.869 | 783 | 12.45 | 68 | 1.08 | 14.171 | 11.51 | 28 | 6 |
| Boyacá | 23.012 | 145 | 6.30 | 36 | 1.56 | 7.502 | 4.03 | 14 | 1 |
| Caldas | 7.291 | 245 | 33.60 | 36 | 4.94 | 7.502 | 6.81 | 14 | 1 |
| Cundinamarca | 23.942 | 419 | 17.50 | 53 | 2.21 | 11.045 | 7.91 | 23 | 0 |
| Huila | 18.331 | 62 | 3.38 | 22 | 1.20 | 4.585 | 2.82 | 18 | 0 |
| Quindío | 1.943 | 150 | 77.20 | 38 | 19.56 | 7.919 | 3.95 | 25 | 0 |
| Norte de Santander | 22.007 | 79 | 3.59 | 36 | 1.64 | 7.502 | 2.19 | 25 | 0 |
| Risaralda | 3.592 | 68 | 18.93 | 24 | 6.68 | 5.002 | 2.83 | 20 | 0 |
| Santander | 30.537 | 207 | 6.78 | 48 | 1.57 | 10.003 | 4.31 | 31 | 3 |
| Tolima | 22.672 | 213 | 9.39 | 43 | 1.90 | 8.961 | 4.95 | 27 | 4 |
| Andean and Pacific | |||||||||
| Cauca | 30.985 | 161 | 5.20 | 42 | 1.36 | 8.753 | 3.83 | 24 | 1 |
| Nariño | 32.046 | 170 | 5.30 | 44 | 1.40 | 9.170 | 3.79 | 27 | 0 |
| Valle del Cauca | 21.195 | 420 | 19.82 | 56 | 2.69 | 11.670 | 7.38 | 28 | 1 |
| Pacific | |||||||||
| Chocó | 46.530 | 210 | 4.51 | 39 | 0.84 | 8.356 | 5.38 | 23 | 1 |
| Caribbean | 3.319 | 18 | 5.42 | 7 | 2.11 | 1.459 | 2.57 | 5 | 0 |
| Atlántico | |||||||||
| Bolívar | 26.469 | 33 | 1.25 | 15 | 0.57 | 3.126 | 2.20 | 9 | 1 |
| Cesar | 22.213 | 13 | 0.59 | 10 | 0.45 | 2.084 | 1.30 | 9 | 0 |
| Córdoba | 25.020 | 33 | 1.32 | 9 | 0.36 | 1.876 | 3.67 | 6 | 0 |
| La Guajira | 20.848 | 21 | 1.01 | 12 | 0.58 | 2.501 | 1.75 | 9 | 0 |
| Magdalena | 22.742 | 84 | 3.69 | 31 | 1.36 | 6.460 | 2.71 | 19 | 1 |
| S. Andrés y Providencia | 53 | 4 | 75.47 | 2 | 37.74 | 0.417 | 2.00 | 2 | 0 |
| Sucre | 10.917 | 6 | 0.55 | 3 | 0.27 | 0.625 | 2.00 | 2 | 0 |
| Orinoquian | |||||||||
| Arauca | 23.393 | 10 | 0.43 | 6 | 0.26 | 1.250 | 1.67 | 3 | 0 |
| Casanare | 44.428 | 4 | 0.09 | 4 | 0.09 | 0.834 | 1.00 | 4 | 0 |
| Meta | 85.286 | 85 | 1.00 | 24 | 0.28 | 4.930 | 3.56 | 14 | 0 |
| Vichada | 100.242 | 16 | 0.16 | 9 | 0.09 | 1.876 | 1.78 | 6 | 0 |
| Amazonian | |||||||||
| Amazonas | 109.665 | 85 | 0.75 | 16 | 0.15 | 3.175 | 5.31 | 14 | 0 |
| Caquetá | 91.725 | 46 | 0.50 | 17 | 0.20 | 3.425 | 2.71 | 13 | 0 |
| Guainía | 70.691 | 16 | 0.23 | 10 | 0.14 | 2.084 | 1.60 | 9 | 0 |
| Guaviare | 55.391 | 27 | 0.49 | 14 | 0.25 | 5.418 | 1.93 | 11 | 0 |
| Putumayo | 24.885 | 56 | 2.25 | 26 | 1.04 | 2.918 | 2.15 | 20 | 0 |
| Vaupés | 54.135 | 34 | 0.63 | 19 | 0.36 | 4.014 | 1.79 | 10 | 0 |
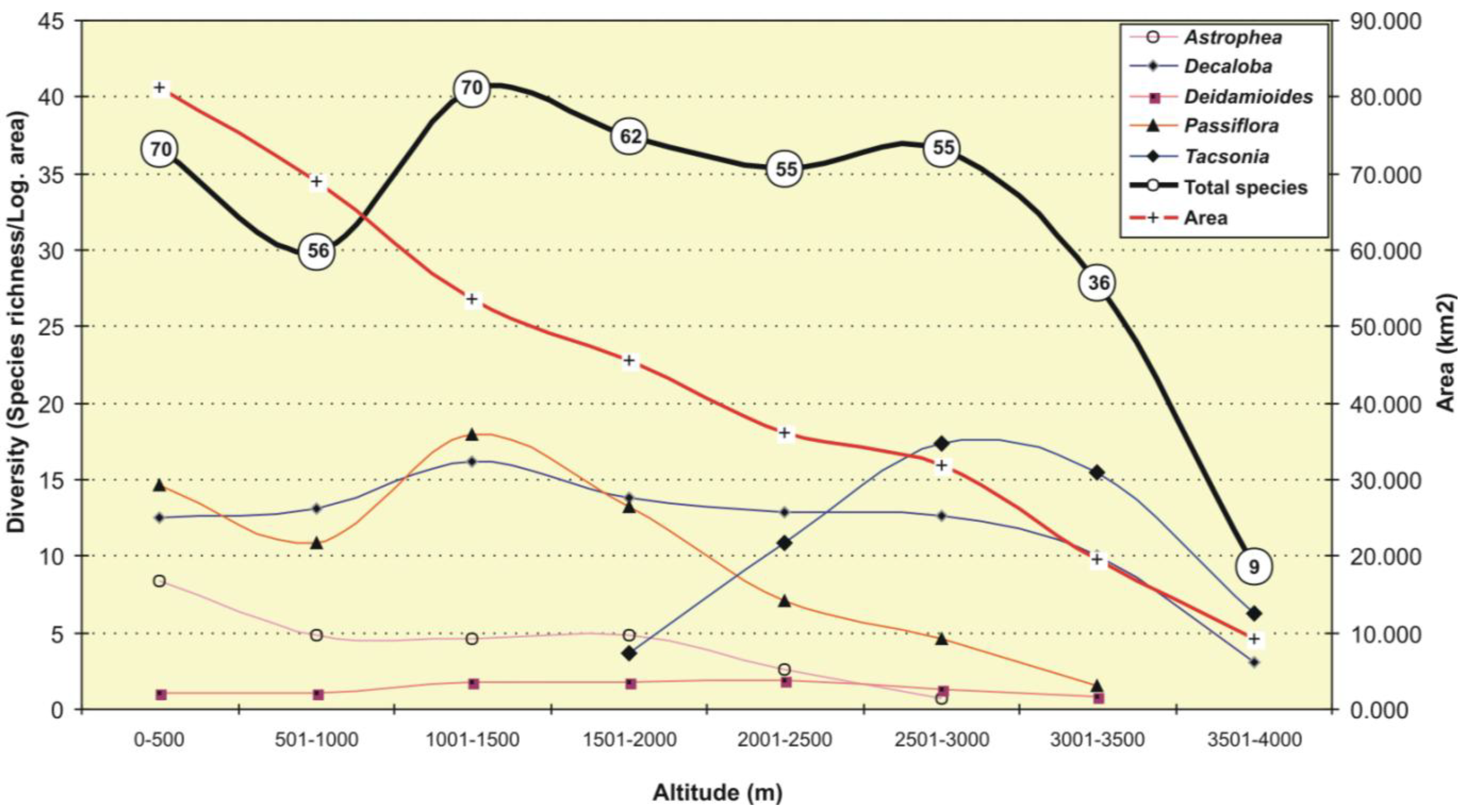
3.2. Altitudinal Distribution
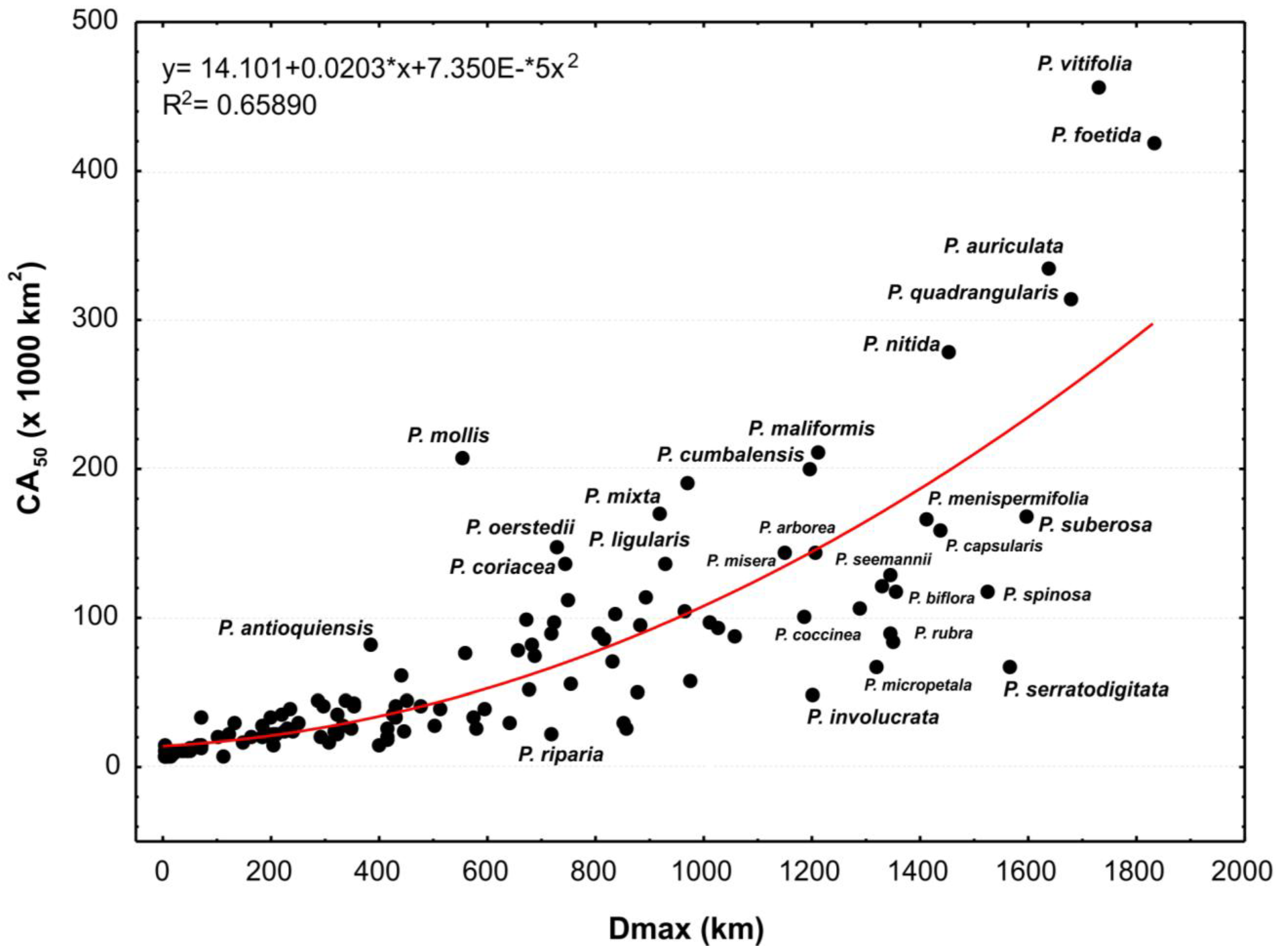
3.3. Climatic Requirements

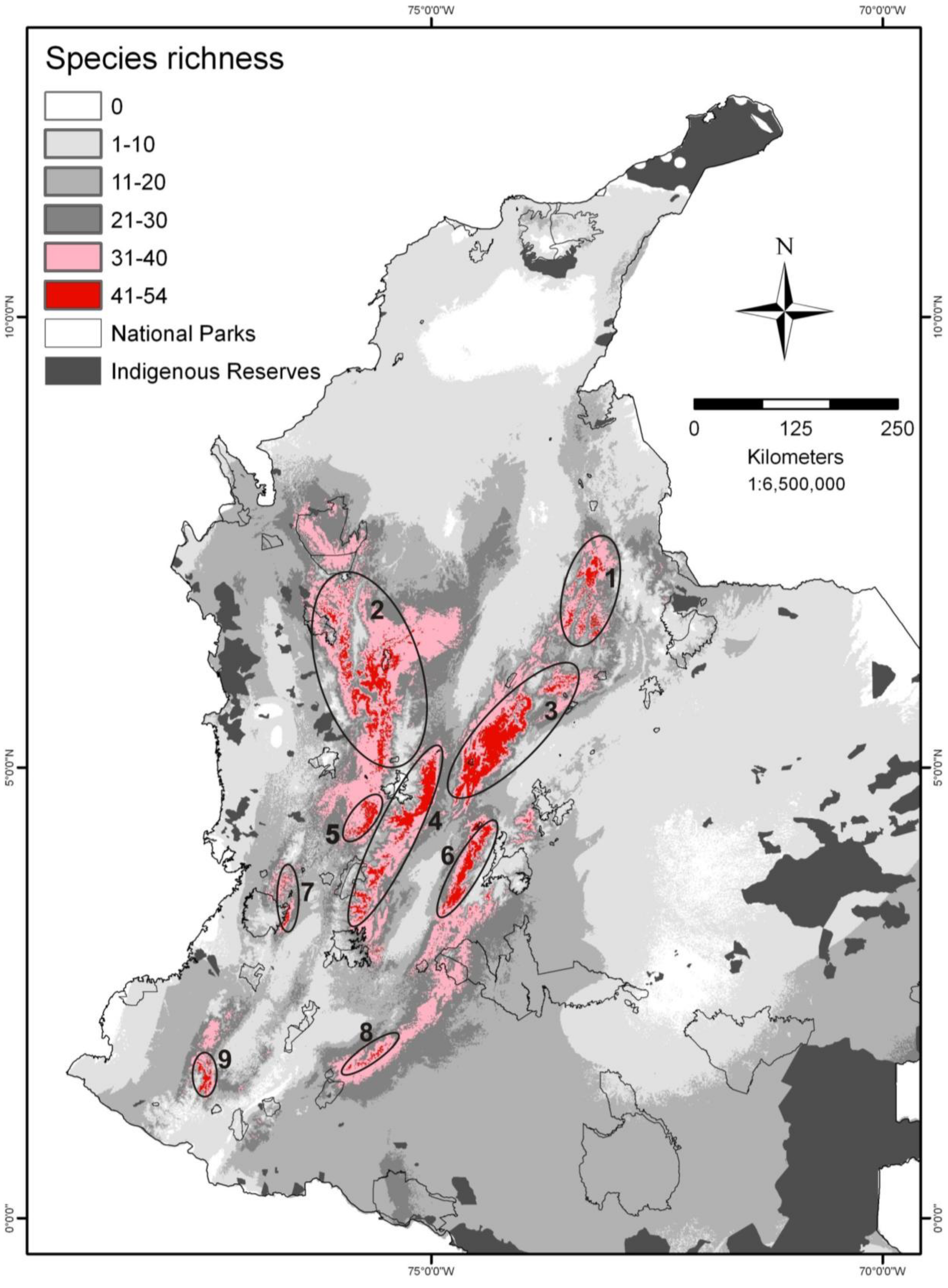
3.4. Areas of Distribution and Endemism


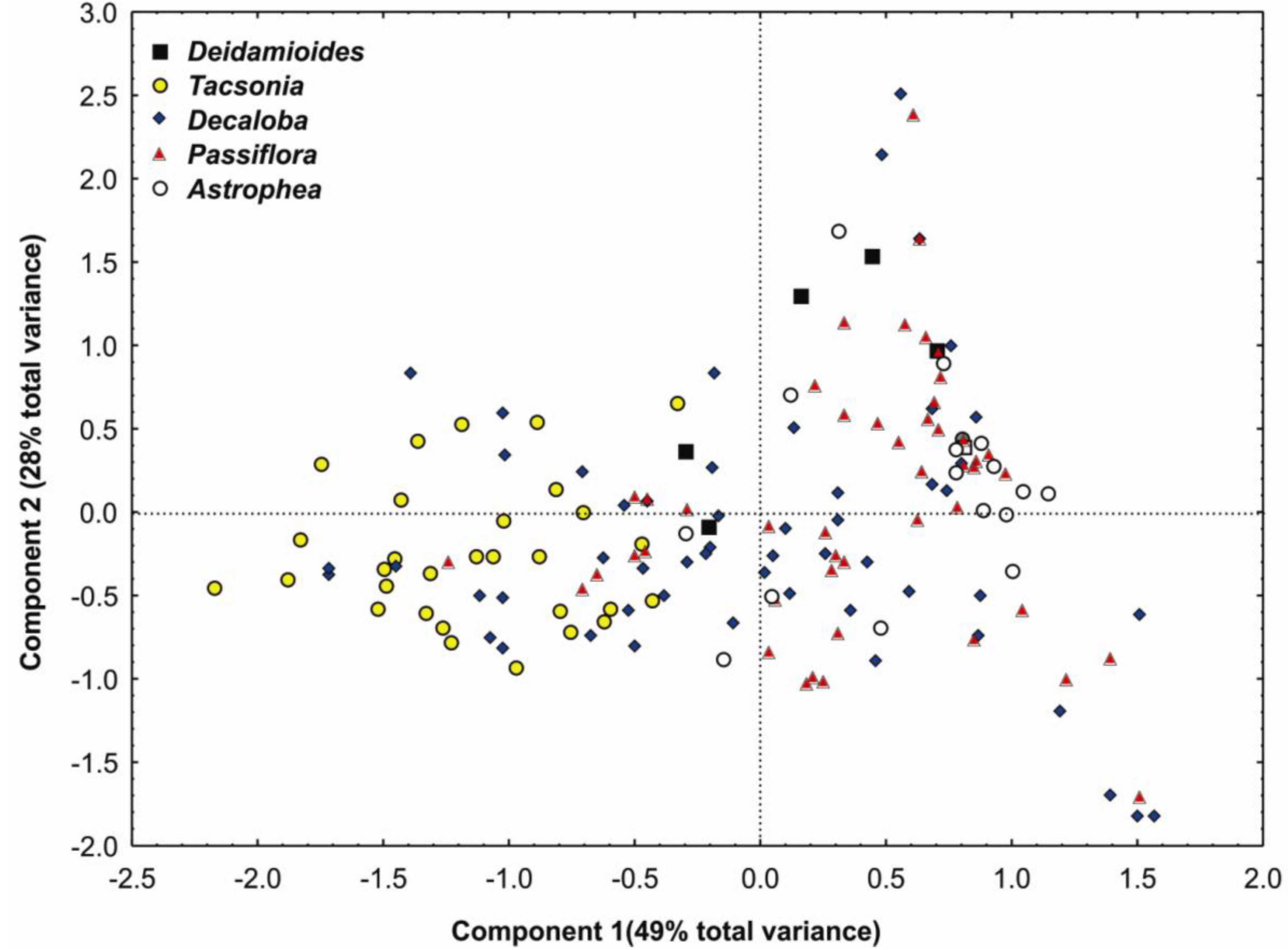
3.5. Modeling Distributions and Species Assemblages
| Bioclim Parameters | Principal components | |||
|---|---|---|---|---|
| Annual Mean Temperature | 0.98 | 0.17 | 0.09 | −0.03 |
| Mean Monthly Temperature Range | 0.08 | −0.21 | −0.16 | −0.96 |
| Isothermality | 0.00 | 0.06 | −0.95 | −0.01 |
| Temperature Seasonality | 0.45 | 0.03 | 0.77 | −0.18 |
| Max, Temperature of Warmest Month | 0.97 | 0.16 | 0.12 | −0.12 |
| Min, Temperature of Coldest Month | 0.98 | 0.20 | 0.06 | 0.04 |
| Temp, Annual Range | 0.08 | −0.22 | 0.37 | −0.89 |
| Mean Temperature of Wettest Quarter | 0.98 | 0.17 | 0.09 | −0.02 |
| Mean Temperature of Driest Quarter | 0.98 | 0.18 | 0.10 | −0.04 |
| Mean Temperature of Warmest Quarter | 0.98 | 0.17 | 0.11 | −0.04 |
| Mean Temperature of Coldest Quarter | 0.98 | 0.17 | 0.07 | −0.03 |
| Annual Precipitation | 0.24 | 0.96 | 0.04 | 0.10 |
| Precipitation of Wettest Month | 0.29 | 0.91 | 0.15 | 0.10 |
| Precipitation of Driest Month | 0.09 | 0.91 | −0.28 | 0.13 |
| Precipitation Seasonality | 0.23 | −0.55 | 0.60 | 0.00 |
| Precipitation of Wettest Quarter | 0.28 | 0.91 | 0.17 | 0.09 |
| Precipitation of Driest Quarter | 0.09 | 0.93 | −0.25 | 0.13 |
| Precipitation of Warmest Quarter | 0.10 | 0.87 | −0.20 | 0.12 |
| Precipitation of Coldest Quarter | 0.29 | 0.89 | 0.05 | 0.02 |
| Eigenvalue | 9.24 | 5.35 | 1.74 | 1.50 |
| Percentage of variance | 48.71 | 28.28 | 9.13 | 7.95 |
3.6. Conservation of Passiflora species and their Habitat


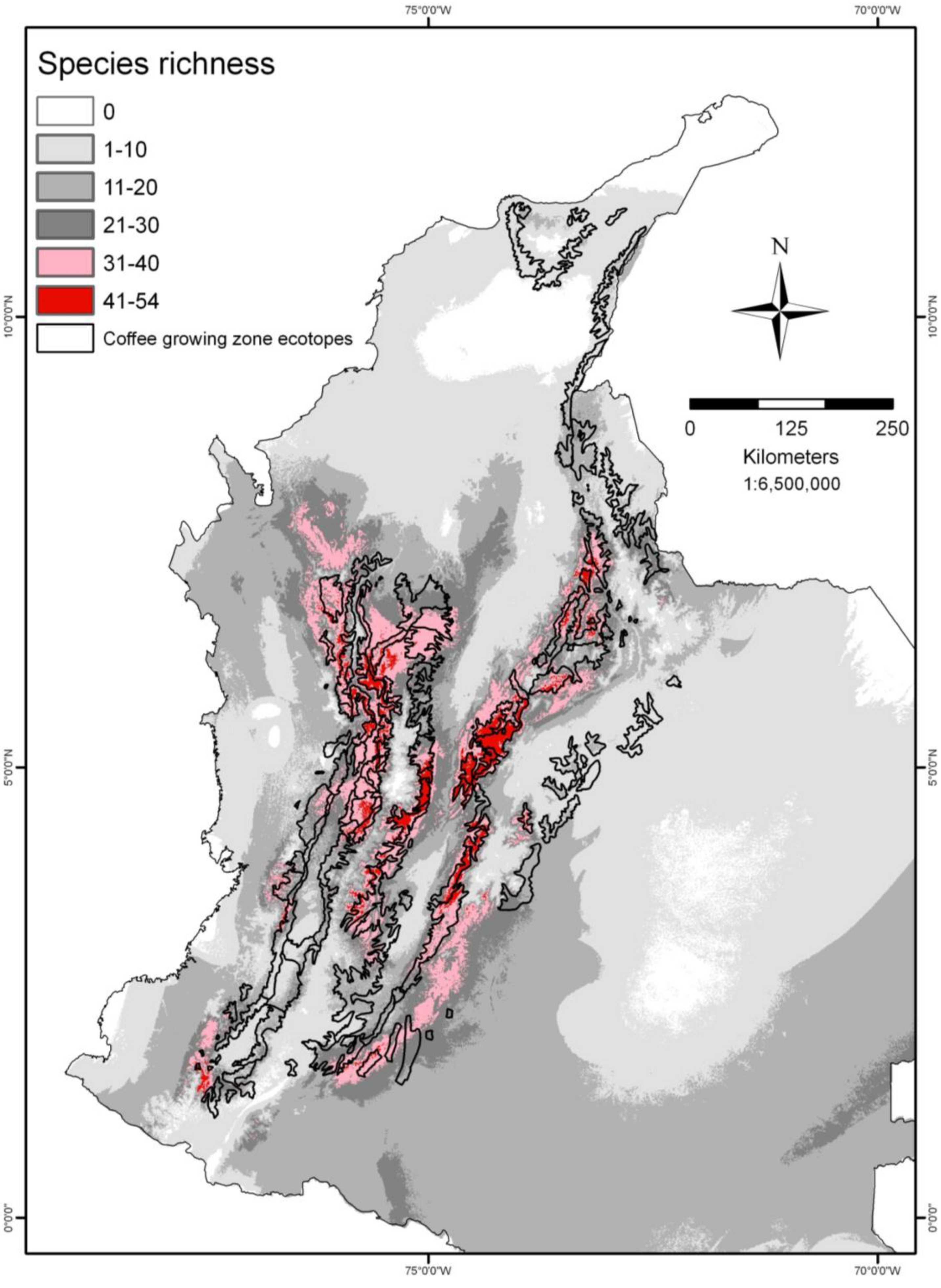
3.7. Passiflora as Indicators of Biodiversity
4. Conclusions
Acknowledgements
References
- Hernández, J.; Hurtado, A.; Ortiz, R.; Walschuburger, T. Unidades biogeográficas de Colombia. In Estado de la biodiversidad en Colombia; Hernández, J., Ortiz, R., Walschuburger, T., Hurtado., Eds.; Colciencias: Bogotá, Colombia, 1991; pp. 15–17. [Google Scholar]
- Gentry, A.H. Endemism in tropical versus temperate plant communities. In Conservation Biology; Soulé, M.E., Ed.; Sinauer Associates: Sunderland, MA, USA, 1986; pp. 153–181. [Google Scholar]
- MacNeely, J.A.; Miller, K.R.; Reid, N.A.; Mittemer, R.A; Wainer, T.B. Conserving the World’s Biological Diversity; International Union for the Conservation of Nature and Natural Resources, World Resources Institute, Conservation International, WWF and World Banks: Washington, DC, USA, 1990; p. 174. [Google Scholar]
- May, R.M. How many species inhabit the Earth? Sci. Amer. 1992, 267, 18–24. [Google Scholar]
- Groombridge, B.; Jenkins, M.D. World Atlas of Biodiversity, Earth’s Living Resources in the 21st Century; University of California Press: Berkeley, CA, USA, 2002; p. 340. [Google Scholar]
- Villegas, B.; Sesana, L.; Hurtado-García, A.; Nieto, F. Parques Naturales Nacionales de Colombia; Villegas, Editores., Ed.; Villegas Editores—Parques Nacionales—Instituto de Investigaciones Biológicas Alexander Von Humboldt: Bogotá, Colombia, 2006; p. 448. [Google Scholar]
- Olson, D.M.; Dinerstein, E. The Global 2000: A representative approach to conserving the earth’s most biologically valuable ecoregions. Conserv. Biol. 1998, 12, 502–515. [Google Scholar]
- Midgley, G.F.; Hannah, L.; Millar, D.; Rutherford, M.C.; Powrie, L.W. Assessing the vulnerability of species richness to anthropogenic climate change in a biodiversity hotspot. Global Ecol. Biogeog. 2002, 11, 445–451. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Word Wildlife Fund (WWW). Biodiversity Vision for the Northern Andes Ecoregional Complex; S. Palminteri: Santiago de Cali, Colombia, 2001; pp. 1–88. [Google Scholar]
- Villareal, H.; Álvarez, M.; Córdoba, S.; Escobar, F.; Fagua, G.; Gast, F.; Mendoza, H.; Ospina, M.; Umaña, A.M. Manual de métodos para el desarrollo de inventarios de biodiversidad, Programa de Inventarios de Biodiversidad, 2nd ed.; Instituto de Investigación de Recursos Biológicos Alexander von Humboldt: Bogotá, Colombia, 2006; p. 210. [Google Scholar]
- Ocampo, J.; Coppens d’Eeckenbrugge, G.; Restrepo, M.; Jarvis, A.; Salazar, M.; Caetano, C. Diversity of Colombian Passifloraceae: biogeography and an updated list for conservation. Biota Colombiana 2007, 8, 1–45. [Google Scholar]
- Hernández, A.; Garcia, N. Las Pasifloras. In Libro rojo de las plantas de Colombia: Las bromelias, las labiadas, y las pasifloras; Garcia, N., Galeano, G., Eds.; Instituto Alexander von Humboldt—Instituto de Ciencias Naturales de la Universidad Nacional de Colombia: Bogotá, Colombia, 2006; pp. 583–567. [Google Scholar]
- Ulmer, T.; MacDougal, J. Passiflora, Passionflowers of the World, 1st ed.; Timber Press: Portland, OR, USA, 2004; p. 430. [Google Scholar]
- Martin, F.W.; Nakasone, H.Y. The edible species of Passiflora. Econ. Bot. 1970, 24, 333–343. [Google Scholar] [CrossRef]
- Coppens d’Eeckenbrugge, G.; Segura, S.D.; Hodson De Jaramillo, E.; Góngora., G.A. Passion Fruits. In Tropical Plant Breeding; Charrier, A., Jacquot, M., Hamon, S., Nicolas, D., Eds.; Enfield Sciences: Montpellier, France, 2001; pp. 381–401. [Google Scholar]
- Passion fruit. Supply and demand. iTi tropicals Inc. 2010. Available online: http://passionfruitjuice.com (accessed on 26 July 2010).
- Killip, E.P. The American Species of Passifloraceae; Botanical Series 19; Field Museum of Natural History Publication: Chicago, IL, USA, 1938; p. 613. [Google Scholar]
- Escobar, L.K. Passifloraceae, Flora de Colombia 10; Instituto de Ciencias Naturales, Universidad Nacional de Colombia: Bogotá, Colombia, 1988; p. 143. [Google Scholar]
- Escobar, L.K. A new subgenus and five new species in Passiflora (Passifloraceae) from South America. Ann. Missouri Bot. Gard. 1989, 76, 877–885. [Google Scholar] [CrossRef]
- MacDougal, J.M. Revision of Passiflora section Decaloba, Pseudodysosmia (Passifloraceae). Syst. Bot. Monogr. 1994, 14, 146. [Google Scholar]
- Feuillet, C.; MacDougal, J. A new infrageneric classification of Passiflora L. (Passifloraceae). Passiflora 2003, 13, 34–38. [Google Scholar]
- Muschner, V.; Lorenz-Lemke, A.; Cervi, A.C.; Bonatto, S.; Souza-Chies, T.; Salzano, F.; Freitas, L. A first molecular phylogenetic analysis of Passiflora (Passifloraceae). Am. J. Bot. 2003, 90, 1229–1238. [Google Scholar] [CrossRef]
- Yockteng, R.; Nadot, S. Phylogenetic relationships among Passiflora species based on the glutamine synthetase nuclear gene expressed in chloroplast (ncpGS). Mol. Phyl. Evol. 2004, 31, 379–396. [Google Scholar] [CrossRef]
- Hansen, A.K.; Lawrence, G.; Simpson, B.B.; Downie, S.R.; Stephen, S.; Cervi, A.C.; Jansen, R.K. Phylogenetic relationships and chromosome number evolution in Passiflora. Syst. Bot. 2006, 31, 138–150. [Google Scholar] [CrossRef]
- Ocampo, J. Study of the Diversity of Genus Passiflora L. (Passifloraceae) and its Distribution in Colombia. Ph.D. Dissertation, Sciences Agronomiques; Ecole Nationale Supérieure Agronomique, de Montpellier, France, 2007; p. 268. [Google Scholar]
- Gilbert, L.E. The evolution of a butterfly and a vine, Heliconious butterflies. Sci. Am. 1982, 247, 10–121. [Google Scholar]
- Radhamani, T.R.; Sudarshana, L.; Krishnan, R. Defence and carnivory: dual roles of bracts in Passiflora foetida L. J. Biosci. 1995, 20, 657–664. [Google Scholar] [CrossRef]
- Apple, J.; Feener, D. Ant visitation of extrafloral nectaries of Passiflora: the effects of nectary attributes and ant behavior on patterns in facultative ant-plant mutualisms. Oecologia 2001, 127, 409–416. [Google Scholar] [CrossRef]
- Büchert, A.; Mogens, J. The fragility of extreme specialization: Passiflora mixta and its pollinating hummingbird Ensifera ensifera. J. Trop. Ecol. 2001, 17, 323–329. [Google Scholar]
- Sazima, M.; Sazima, I. Bat pollination of the passion flower, Passiflora mucronata, in southeastern Brazil. Biotropica 1978, 10, 100–109. [Google Scholar] [CrossRef]
- Debouck, D.G.; Libreros-Ferla, D. Neotropical montane forests: a fragile home of genetic resources of wild relatives of New World crops. In Biodiversity and Conservation of Neotropical Montane Forests, Proceedings of a Symposium; Biodiversity and Conservation of Neotropical Montane Forests, Proceedings of a Symposium, New York Botanical Garden, New York, NY, USA, 21–26 June 1993; Churchill, S.P., Balslev, H., Forero, E., Luteyn, J.L., Eds.; pp. 561–577.
- Maxted, N.; van Slageren, M.W.; Rihan, J.R. Ecogeographic surveys. In Collecting Plant Genetic Diversity, Technical Guidelines; Guarino, L., Ramanatha Rao, V., Reid, R., Eds.; CAB International: Wallingford, Australia, 1995; pp. 255–286. [Google Scholar]
- Pearson, D.L. Selecting indicator taxa for the quantitative assessment of biodiversity. Phil. Trans. Roy. Soc. Lond. 1994, 345, 75–79. [Google Scholar] [CrossRef]
- Skov, F. Potential plant distribution mapping based on climatic similarity. Taxon 2000, 49, 503–515. [Google Scholar] [CrossRef]
- Lehmann, A.; Leathwick, J.R.; Overton, J.M. Assessing New Zealand fern diversity from spatial predictions of species assemblages. Biodivers. Conserv. 2002, 11, 2217–2238. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Spooner, D.M. Geography of wild potato species. Am. J. Bot. 2001, 88, 2101–2112. [Google Scholar] [CrossRef]
- Scheldeman, X.; Willemen, L.; Coppens d'Eeckenbrugge, G.; Romeijn-Peeters, E.; Restrepo, M.T.; Romero Motoche, J.; Jiménez, D.; Lobo, M.; Medina, C.I.; Reyes, C.; Rodríguez, D.; Ocampo, J.A.; Van Damme, P.; Goetgebeur, P. Distribution, diversity and crop ecology of highland papayas (Vasconcellea spp.) and papaya (Carica papaya L.) in Latin America. Biodivers. Conserv. 2006, 16, 1867–1884. [Google Scholar]
- Jarvis, A.; Ferguson, M.E.; Williams, D.E.; Guarino, L.; Jones, P.G.; Stalker, H.T.; Valls, J.F.M.; Pittman, R.N.; Simpson, C.E.; Bramel, P. Biogeography of wild Arachis: assessing conservation status and setting future priorities. Crop Sci. 2003, 43, 1100–1108. [Google Scholar] [CrossRef]
- Kingston, N.; Waldren, S. A conservation appraisal of the rare and endemic vascular plants of Pitcairn Island. Biodivers. Conserv. 2005, 14, 781–800. [Google Scholar] [CrossRef]
- Zaharieva, M.; Prosperi, J.M.; Monneveux, P. Ecological distribution and species diversity of Aegilops L. genus in Bulgaria. Biodivers. Conserv. 2004, 13, 2319–2337. [Google Scholar] [CrossRef]
- Peterson, A.T. Predictability of the geography of species invasions via ecological niche modeling. Quart. Rev. Biol. 2001, 78, 419–433. [Google Scholar] [CrossRef]
- Peterson, A.T.; Robins, C.R. When endangered meets invasive: ecological niche modeling predicts double trouble for spotted owls, Strix occidentalis. Conserv. Biol. 2003, 17, 1161–1165. [Google Scholar] [CrossRef]
- Leimberck, R.; Valencia, R.; Balslev, H. Landscape diversity patterns and endemism of Araceae in Ecuador. Biodivers. Conserv. 2004, 13, 1755–1779. [Google Scholar] [CrossRef]
- Peterson, A.T. Predicting species geographic distribution based on ecological niche modeling. Condor 2001, 10, 599–605. [Google Scholar] [CrossRef]
- Rodríguez, D.; Marín, C.; Quecán, H.; Ortiz, R. Áreas potenciales para colectas del género Vasconcellea Badillo en Venezuela. Bioagro 2005, 17, 3–10. [Google Scholar]
- Jarvis, A.; Lane, A.; Hijmans, R.J. The effect of climate change on crop wild relatives. Agricult., Ecosyst. Environm. 2008, 126, 13–23. [Google Scholar]
- Segura, S.; Coppens d’Eeckenbrugge, G.; López, L.; Grum, M.; Guarino, L. Mapping the potential distribution of five species of Passiflora in Andean countries. Gen. Res. Crop. Evol. 2003, 50, 555–566. [Google Scholar]
- Killip, E.P. Supplemental Notes on the American Species of Passifloraceae with Descriptions of New Species; Bulletin of the United States National Museum—Herbarium; Volume 35, Part 1; Smithsonian Institution: Washington, DC, USA, 1960; p. 29. [Google Scholar]
- Uribe, L. Pasifloráceas y Begoniáceas de la Real Expedición Botánica del Nuevo Reino de Granada; Ediciones Cultura Hispánica 26: Madrid, España, 1955; p. 98. [Google Scholar]
- Escobar, L.K. Two new species and a key to Passiflora subg. Astrophea. Syst. Bot. 1994, 19, 203–210. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Schreuder, M.; De la Cruz, M.; Rojas, E. Computer tools for spatial analysis of plant genetic resources data: DIVA-GIS. Pl. Gen. Res. Newsl. 2001, 27, 15–19. [Google Scholar]
- Busby, J.R. BIOCLIM - A bioclimate prediction system. In Nature Conservation: Cost Effective Biological Surveys and Data Analysis; Margules, C.R., Austin, M.P., Eds.; CSIRO: Melbourne, Australia, 1991; pp. 4–68. [Google Scholar]
- Hijmans, R.J.; Graham, C.H. The ability of climate envelope models to predict the effect of climate change on species distributions. Glob. Chan. Biol. 2006, 12, 2272–2281. [Google Scholar] [CrossRef]
- Rebelo, A.G.R. Iterative selection procedures: centers of endemism and optimal placement of reserves. Strelitzia 1994, 1, 231–257. [Google Scholar]
- Jørgensen, P.M. Passifloraceae. In Catalogue of the Vascular Plants of Ecuador; Jørgensen, P.M., León-Yánez, S., Eds.; Monographs in Systematic Botany 75; Missouri Botanical Garden: St. Louis, MO, USA, 1999; pp. 779–783. [Google Scholar]
- Kessler, M. The elevational gradient of Andean plant endemism: varying influences of taxon-specific traits and topography at different taxonomic levels. J. Biogeogr. 2002, 29, 1159–1165. [Google Scholar] [CrossRef]
- IUCN. Guidelines for Application of IUCN Red List Criteria at Regional Levels: Version 3.0; IUCN Species Survival Commission; IUCN: Gland, Switzerland and Cambridge, UK, 2003; p. 26. [Google Scholar]
- Contreras-Medina, R.; Luna-Vega, I. Species richness, endemism and conservation of Mexican gymnosperms. Biodivers. Conserv. 2007, 16, 1803–1821. [Google Scholar] [CrossRef]
- Werner, U.; Buszko, J. Detecting diversity hot spots using species-area and endemics-area relationships: the case of butterflies. Biodiv. Conser. 2005, 14, 1977–1988. [Google Scholar] [CrossRef]
- Jaramillo, M.A. Using Piper species diversity to identify conservation priorities in the Chocó Region of Colombia. Biodivers. Conserv. 2006, 15, 1695–1712. [Google Scholar] [CrossRef]
- Federación Nacional de Cafeteros de Colombia. El Café. In Atlas de Colombia, 5a ed.; Instituto Agustín Codazzi: Bogotá, Colombia, 2003; p. 252. [Google Scholar]
- Perfecto, I.; Mas, A.; Dietsch, T.; Vandermeer, J. Conservation of biodiversity in coffee agroecosystems: a tri-taxa comparison in southern Mexico. Biodivers. Conserv. 2003, 12, 1239–1252. [Google Scholar] [CrossRef]
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ocampo, J.; Coppens D’Eeckenbrugge, G.; Jarvis, A. Distribution of the Genus Passiflora L. Diversity in Colombia and Its Potential as an Indicator for Biodiversity Management in the Coffee Growing Zone. Diversity 2010, 2, 1158-1180. https://doi.org/10.3390/d2111158
Ocampo J, Coppens D’Eeckenbrugge G, Jarvis A. Distribution of the Genus Passiflora L. Diversity in Colombia and Its Potential as an Indicator for Biodiversity Management in the Coffee Growing Zone. Diversity. 2010; 2(11):1158-1180. https://doi.org/10.3390/d2111158
Chicago/Turabian StyleOcampo, John, Geo Coppens D’Eeckenbrugge, and Andy Jarvis. 2010. "Distribution of the Genus Passiflora L. Diversity in Colombia and Its Potential as an Indicator for Biodiversity Management in the Coffee Growing Zone" Diversity 2, no. 11: 1158-1180. https://doi.org/10.3390/d2111158