Social Organization of Crop Genetic Diversity. The G × E × S Interaction Model

Abstract
:
1. Introduction

2. Overview of in situ Crop Genetic Diversity Studies
2.1. Sorghum Studies
2.1.1. Sorghum Diversity at the Continental Scale
2.1.2. Sorghum Diversity at the National Scale
2.2. Maize Studies
2.2.1. Maize Diversity at the Continental Scale
2.2.2. Maize Diversity at Regional Scales
3. Farmer Social Organization and Cultural Diversity
3.1. Social Embeddedness and the Orientation of Exchanges
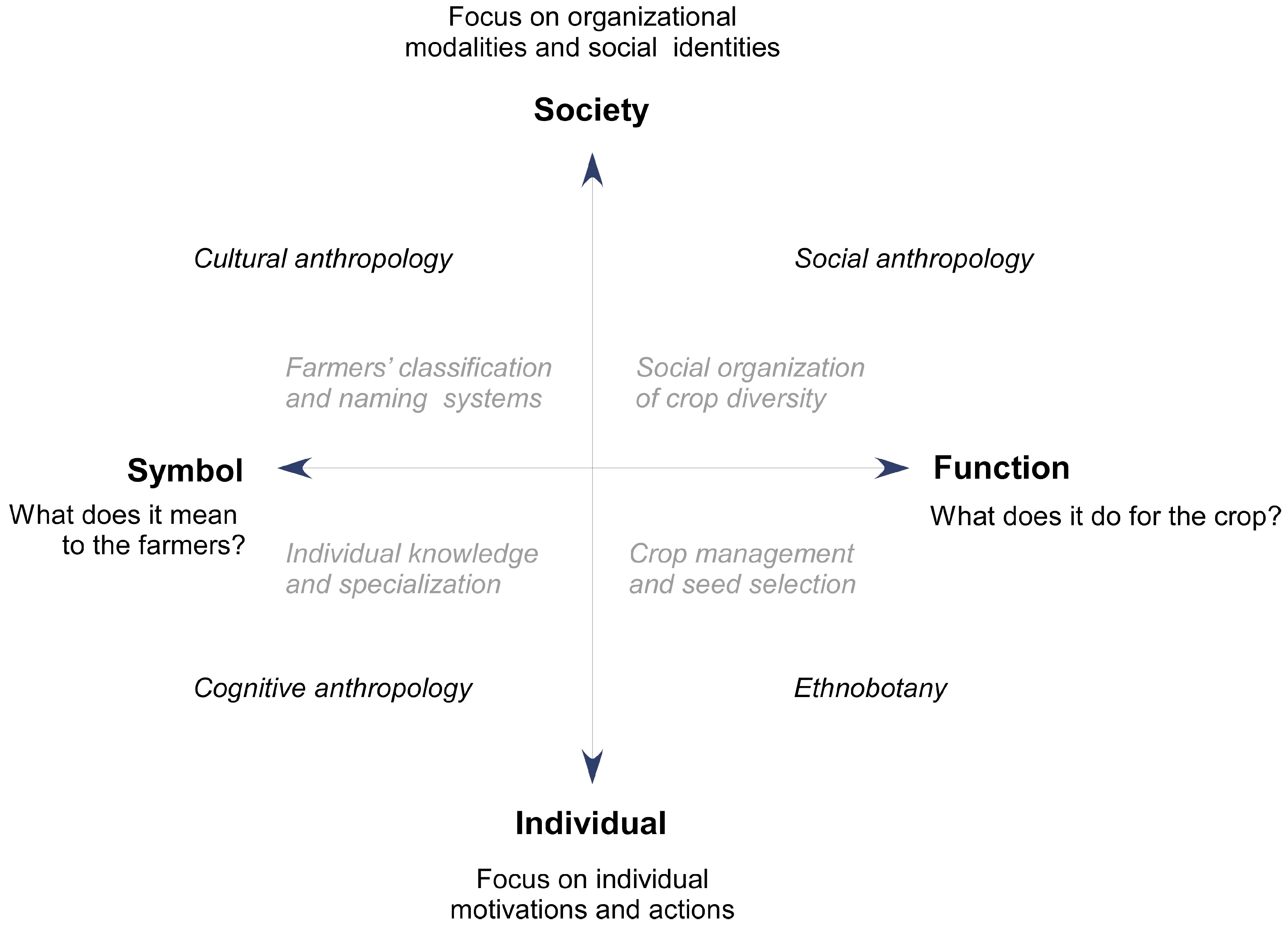
3.2. The Anthropologist’s Concepts of Cultural Diversity
3.2.1. Organizational Modalities of Human Societies
3.2.2. Cultural Diversity Explanatory Models
4. Mechanisms Leading to Social Organization of Crop Genetic Diversity
4.1. Traditional Knowledge, Perception of Crop Diversity and Conservative Selection
4.2. Seed Exchange Embedded into the Social System and Vertical Transmission
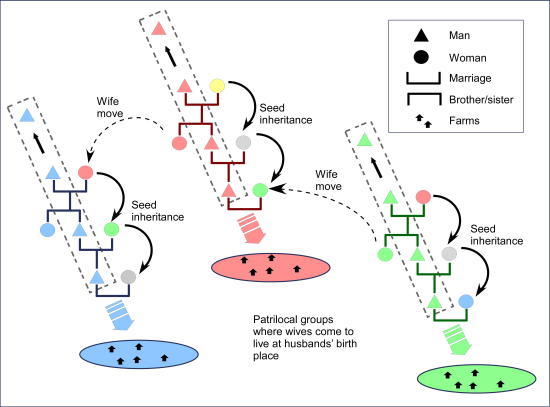
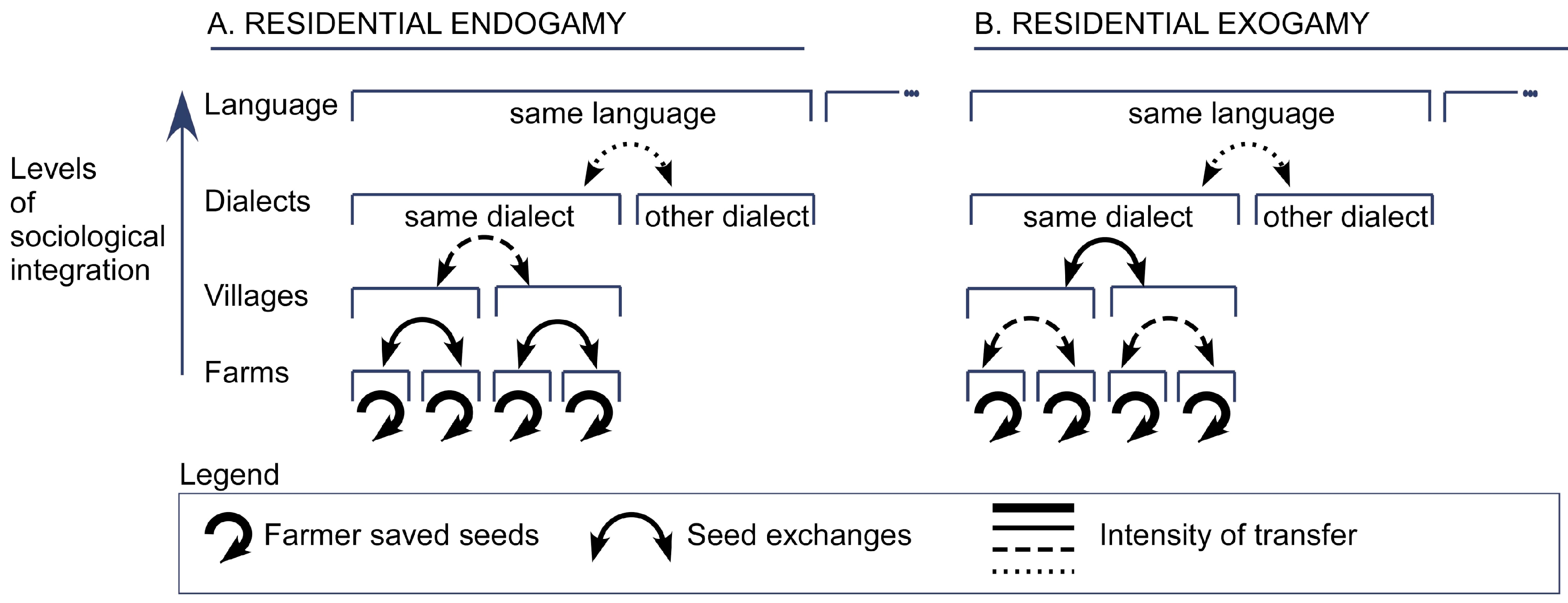
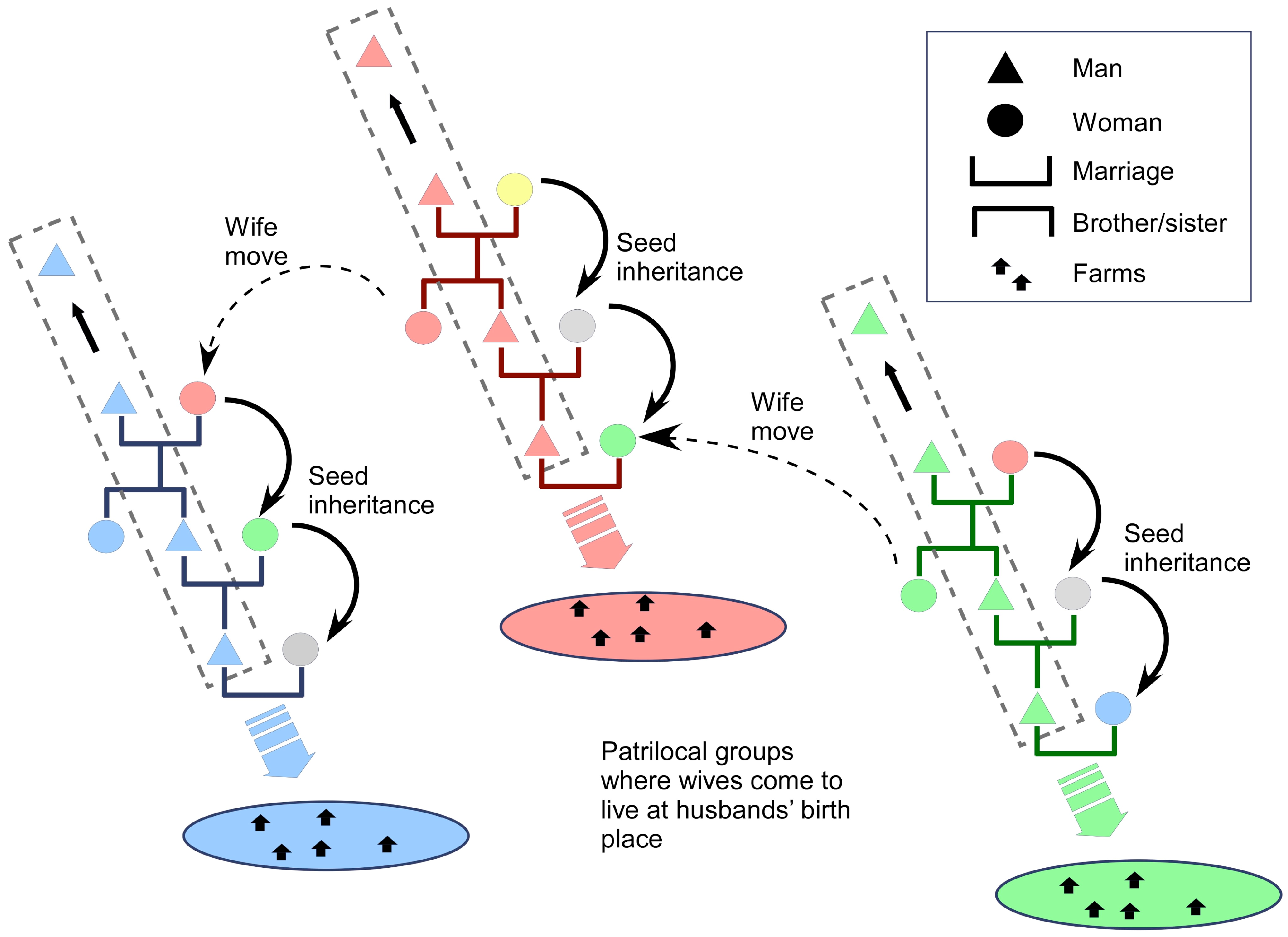
4.2.1. Basic Levels of Sociological Integration

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country | Crop | Farmer saved seeds | Family and neigbourhood | Total | References |
|---|---|---|---|---|---|
| * This percentage refers only to family members that made up 47.5% of seed providers. | |||||
| Burkina Faso | Sorghum | 70%–99% | NA | 70%–99% | [119] |
| Costa Rica | Maize | 79% | 19% | 98% | [117] |
| Costa Rica | Beans | 58% | 21% | 79% | [117] |
| Guatemala | Maize | 59% | 31% | 90% | [22] |
| Honduras | Maize | 75% | 13% | 88% | [117] |
| Honduras | Beans | 79% | 15% | 94% | [117] |
| Mexico | Maize | 90% | 9% | 99% | [120,121] |
| Mexico | Maize | 58% | 34% | 92% | [75] |
| Mexico | Maize | 79% | NA | 79% | [5] |
| Mexico | Maize | NA | 87% | 87% | [76] |
| Mexico | Maize | NA | 95% | 95% | [79] |
| Mexico | Maize (Tzeltal) | 84% | 9% | 93% | [24] |
| Mexico | Maize (Tzotzil) | 87% | 10% | 97% | [24] |
| Mexico | Maize | 76% | 11% * | 87% | [23] |
| Mexico | Maize | 76% | 21% | 97% | [84] |
| Nicaragua | Maize | 81% | 12% | 93% | [117] |
| Nicaragua | Beans | 72% | 14% | 86% | [117] |
| Peru | Potatoes/ulluco | 91% | 6% | 97% | [19] |
| Sierra Leone | Rice | 70% | NA | 70% | [89] |


4.2.2. Higher Levels of Sociological Integration
5. Conclusion
Acknowledgements
References
- Orlove, B.S.; Brush, S.B. Anthropology and the conservation of biodiversity. Annu. Rev. Anthropol. 1996, 25, 329–352. [Google Scholar]
- Soleri, D.; Smith, S.E. Conserving folk crop varieties: Different agricultures, different goals. In Ethnoecology. Situated Knowledge/Located Lives; Nazarea, V.D., Ed.; The University of Arizona Press: Tucson, AZ, USA, 1999; pp. 133–154. [Google Scholar]
- Brush, S.B. A farmer-based approach to conserving crop germplasm. Econ. Bot. 1991, 45, 153–165. [Google Scholar]
- Louette, D.; Smale, M. Genetic Diversity and Maize Seed Management in a Traditional Mexican Community: Implications for in situ Conservation of Maize; CIMMYT: El Batan, Mexico, 1996. [Google Scholar]
- Perales, H.R.; Brush, S.B.; Qualset, C.O. Dynamic management of maize landraces in Central Mexico. Econ. Bot. 2003, 57, 21–34. [Google Scholar]
- Brush, S.B. Cultural research on the origin and maintenance of agricultural diversity. In Nature Knowledge: Ethnoscience, Cognition, and Utility; Sanga, G., Ortalli, G., Eds.; Berghahn Books: New York, NY, USA, 2005; pp. 379–385. [Google Scholar]
- Caillon, S.; Degeorges, P. Biodiversity: Negotiating the border between nature and culture. Biodivers. Conserv. 2007, 16, 2919–2931. [Google Scholar]
- Granovetter, M. Economic action and social structure: The problem of embeddedness. Am. J. Sociol. 1985, 91, 481–510. [Google Scholar]
- Lacy, S.M.; Cleveland, D.; Soleri, D. Farmer choice of sorghum varieties in Southern Mali. Hum. Ecol. 2006, 34, 331–353. [Google Scholar]
- Perales, H.R.; Brush, S.B.; Qualset, C.O. Landraces of maize in Central Mexico: An altitudinal transect. Econ. Bot. 2003, 57, 7–20. [Google Scholar]
- Brush, S.B. Farmers’ Bounty. Locating Crop Diversity in the Contempory World; Yale University Press: New Haven, CT, USA; London, UK, 2004. [Google Scholar]
- Augé, M. Symbole, Fonction, Histoire. Les Interrogations de l’anthropologie; Hachette: Paris, France, 1979. [Google Scholar]
- Descola, P.; Lenclud, G.; Severi, C.; Taylor, A.-C. Les Idées de l'anthropologie; Armand Colin: Paris, France, 1988. [Google Scholar]
- Alvarez, N.; Garine, E.; Khasah, C.; Dounias, E.; Hossaert-McKey, M.; McKey, D. Farmers’ practices, metapopulation dynamics, and conservation of agricultural biodiversity on-farm: A case study of sorghum among the Duupa in sub-sahelian Cameroon. Biol. Conserv. 2005, 121, 533–543. [Google Scholar]
- Barnaud, A.; Deu, M.; Garine, E.; McKey, D.; Joly, H. Local genetic diversity of sorghum in a village in Northern Cameroon: Structure and dynamics of landraces. Theor. Appl. Genet. 2007, 114, 237–248. [Google Scholar]
- Elias, M.; McKey, D.; Panaud, O.; Anstett, M.C.; Robert, T. Traditional management of cassava morphological and genetic diversity by the Makushi Amerindian (Guyana, South America): perspective for on-farm conservation of crop genetic resources. Euphytica 2001, 120, 143–157. [Google Scholar]
- Elias, M.; Rival, L.; McKey, D. Perception and management of cassava, Manihot esculenta (Crantz) diversity among Makushi Amerindians of Guyana. J. Ethnobiol. Ethnomed. 2000, 20, 239–265. [Google Scholar]
- Thomas, M.; Dawson, J.C.; Goldringer, I.; Bonneuil, C. Seed exchanges, a key to analyze crop diversity dynamics in farmer-led on-farm conservation. Genet. Resour. Crop Evol. 2011, 58, 321–338. [Google Scholar]
- Zimmerer, K.S. Geographies of seed networks for food plants (potatoes, ulluco) and approaches to agrobiodiversity conservation in the Andean countries. Soc. Nat. Resour. 2003, 16, 583–601. [Google Scholar]
- Van Etten, J. Genetic diversity of maize (Zea mays L. ssp. mays) in communities of the western highlands of Guatemala: Geographical patterns and processes. Genet. Resour. Crop Evol. 2008, 55, 303–317. [Google Scholar] [CrossRef]
- Van Etten, J. Molding maize: The shaping of a crop diversity landscape in the western highlands of Guatemala. J. Hist. Geogr. 2006, 32, 689–711. [Google Scholar]
- Van Etten, J.; de Bruin, S. Regional and local maize seed exchange and replacement in the western highlands of Guatemala. Plant Genet. Res. Charact. Util. 2007, 5, 57–70. [Google Scholar]
- Badstue, L.B.; Bellon, M.R.; Berthaud, J.; Ramírez, A.; Flores, D.; Juárez, X. The dynamics of farmers’ maize seed supply practices in the Central Valleys of Oaxaca, Mexico. World Dev. 2007, 35, 1579–1593. [Google Scholar]
- Perales, H.R.; Benz, B.F.; Brush, S.B. Maize diversity and ethnolinguistic diversity in Chiapas, Mexico. Proc. Natl. Acad. Sci. USA 2005, 102, 949–954. [Google Scholar]
- Brush, S.B.; Perales, H.R. A maize landscape: Ethnicity and agro-biodiversity in Chiapas, Mexico. Agric. Ecosyst. Environ. 2007, 121, 211–221. [Google Scholar]
- Fox, R. Kinship and Marriage. An Anthropological Perspective; Cambridge University Press: Cambridge, UK, 1967. [Google Scholar]
- Van Heerwaarden, J.; van Eeuwijk, F.; Ross-Ibarra, J. Genetic diversity in a crop metapopulation. Heredity 2010, 104, 28–39. [Google Scholar]
- Wendorf, F.; Close, A.E.; Schild, R.; Wasylikowa, K.; Housley, R.A.; Harlan, J.R.; Kralik, H. Saharan exploitation of plants 8,000 years BP. Nature 1992, 359, 721–724. [Google Scholar]
- Ollitrault, P.; Noyer, J.-L.; Chantereau, J.; Glaszmann, J.-C. Structure génétique et dynamique des variétés traditionnelles de sorgho au Burkina Faso. In Gestion des Ressources Génétiques des Plantes en Afrique des Savanes; Chirat: St-Just-la-Pendue, France, 1997; pp. 231–240. [Google Scholar]
- Djé, Y.; Heuertz, M.; Ater, M.; Lefèbvre, C.; Vekemans, X. In situ estimation of outcrossing rate in sorghum landraces using microsatellite markers. Euphytica 2004, 138, 205–212. [Google Scholar]
- Barnaud, A.; Trigueros, G.; McKey, D.; Joly, H.I. High outcrossing rates in fields with mixed sorghum landraces: How are landraces maintained? Heredity 2008, 101, 445–452. [Google Scholar] [CrossRef]
- Harlan, J.R.; de Wet, J.M.J. A simplified classification of cultivated sorghum. Crop Sci. 1972, 12, 172–176. [Google Scholar]
- De Wet, J.M.J. Systematics and evolution of Sorghum Sect. Sorghum (Gramineae). Am. J. Bot. 1978, 65, 477–484. [Google Scholar] [CrossRef]
- Anderson, E.; Cutler, H.C. Races of Zea mays. I. Their recognition and classification. Ann. Mo. Bot. Gard. 1942, 29, 69–89. [Google Scholar] [CrossRef]
- Harlan, J.R.; De Wet, J.M.J. Toward a rational classification of cultivated plants. Taxon 1971, 20, 509–517. [Google Scholar]
- De Wet, J.M.J.; Huckabay, J.P. The origin of Sorghum bicolor. Distribution and domestication. Evolution 1967, 787–802. [Google Scholar]
- Stemler, A.; Harlan, J.R.; de Wet, J.M.J. Caudatum sorghums and speakers of Chari-Nile languages in Africa. J. Afr. Hist. 1975, 16, 161–183. [Google Scholar]
- Harlan, J.R.; Stemler, A. The races of sorghum in Africa. In Origins of African Plant Domestication; Harlan, J.R., de Wet, J.M.J., Stemler, A., Eds.; Mouton: The Hague, The Netherlands; Paris, France, 1976; pp. 465–478. [Google Scholar]
- Doggett, H. Sorghum; Wiley: New York, NY, USA, 1988. [Google Scholar]
- Chantereau, J.; Arnaud, M.; Ollitrault, P.; Nabaya Ogo, P.; Noyer, J.-L. Etude de la diversité morphophysiologique et classification des sorghos cultivés. Agron. Trop. 1989, 44, 223–232. [Google Scholar]
- Deu, M.; Hamon, P.; Bonnot, F.; Chantereau, J. Sorghum. In Genetic Diversity of Cultivated Tropical Plants; Hamon, P., Seguin, M., Perrier, X., Glaszmann, J.-C., Eds.; CIRAD: Montpellier, France, 2003; pp. 307–336. [Google Scholar]
- Ollitrault, P.; Arnaud, M.; Chantereau, J. Polymorphisme enzymatique des sorghos II. Organisation génétique et évolutive des sorghos cultivés. Agron. Trop. 1989, 44, 211–222. [Google Scholar]
- Deu, M.; González de León, D.; Glaszmann, J.-C.; Degremont, I.; Chantereau, J.; Lanaud, C.; Hamon, P. RFLP diversity in cultivated sorghum in relation to racial differentiation. Theor. Appl. Genet. 1994, 88, 838–844. [Google Scholar]
- Cui, Y.X.; Xu, G.W.; Magill, C.W.; Schertz, K.F.; Hart, G.E. RFLP-based assay of Sorghum bicolor (L.) Moench genetic diversity. Theor. Appl. Genet. 1995, 90, 787–796. [Google Scholar]
- Deu, M.; Rattunde, F.; Chantereau, J. A global view of genetic diversity in cultivated sorghum using a core collection. Genome 2006, 49, 168–180. [Google Scholar]
- Djé, Y.; Heuertz, M.; Lefèbvre, C.; Vakemans, X. Assessment of genetic diversity within and among germplasm accessions in cultivated sorghum using microsatellite markers. Theor. Appl. Genet. 2000, 100, 918–925. [Google Scholar]
- Casa, A.M.; Mitchell, S.E.; Hamblin, M.T.; Sun, H.; Bowers, J.E.; Paterson, A.H.; Aquadro, C.F.; Kresovich, S. Diversity and selection in sorghum: Simultaneous analyses using simple sequence repeats. Theor. Appl. Genet. 2005, 111, 23–30. [Google Scholar]
- Ghebru, B.; Schmidt, R.J.; Bennetzen, J.L. Genetic diversity of Eritrean sorghum landraces assessed with simple sequence repeat (SSR) markers. Theor. Appl. Genet. 2002, 105, 229–236. [Google Scholar]
- Ayana, A.; Bryngelsson, T.; Bekele, E. Genetic variation of Ethiopian and Eritrean sorghum (Sorghum bicolor (L.) Moench) germplasm assessed by random amplified polymorphic DNA (RAPD). Genet. Resour. Crop Evol. 2000, 47, 471–482. [Google Scholar] [CrossRef]
- Djé, Y.; Forcioli, D.; Ater, M.; Lefebvre, C.; Vekemans, X. Assessing population genetic structure of sorghum landraces from North-western Morocco using allozyme and microsatellite markers. Theor. Appl. Genet. 1999, 99, 157–163. [Google Scholar]
- Medraoui, L.; Ater, M.; Benlhabib, O.; Msikine, D.; Filali-Maltouf, A. Evaluation of genetic variability of sorghum (Sorghum bicolor L. Moench) in Northwestern Morocco by ISSR and RAPD markers. C. R. Biol. 2007, 330, 789–797. [Google Scholar] [CrossRef]
- Kayode, A.P.; Linnemann, A.R.; Nout, M.R.; Hounhouigan, J.D.; Stomph, T.J.; Smulders, M.J. Diversity and food quality properties of farmers’ varieties of sorghum from Benin. J. Sci. Food Agric. 2006, 86, 1032–1039. [Google Scholar]
- Abdi, A.; Bekele, E.; Asfaw, Z.; Teshome, A. Patterns of morphological variation of sorghum (Sorghum bicolor (L.) Moench) landraces in qualitative characters in North Shewa and South Welo, Ethiopia. Hereditas 2002, 137, 161–172. [Google Scholar] [CrossRef]
- Appa Rao, S.; Prasada Rao, K.E.; Mengesha, M.H.; Gopal-Reddy, V. Morphological diversity in sorghum germplasm from India. Genet. Resour. Crop Evol. 1996, 43, 559–567. [Google Scholar]
- Djé, Y.; Ater, M.; Lefèbvre, C.; Vekemans, X. Patterns of morphological and allozyme variation in sorghum landraces of Northwestern Morocco. Genet. Resour. Crop Evol. 1998, 45, 541–548. [Google Scholar]
- Djé, Y.; Heuertz, M.; Ater, M.; Lefèbvre, C.; Vekemans, X. Évaluation de la diversité morphologique des variétés traditionnelles de sorgho du Nord-ouest du Maroc. Biotechnol. Agron. Soc. 2007, 11, 39–46. [Google Scholar]
- Ayana, A.; Bekele, E. Geographical patterns of morphological variation in sorghum (Sorghum bicolor (L.) Moench) Germplasm from Ethiopia and Eritrea: Qualitative characters. Hereditas 1998, 129, 195–205. [Google Scholar] [CrossRef]
- Ayana, A.; Bekele, E. Multivariate analysis of morphological variation in sorghum (Sorghum bicolor (L.) Moench) germplasm from Ethiopia and Eritrea. Genet. Resour. Crop Evol. 1999, 46, 273–284. [Google Scholar] [CrossRef]
- Ayana, A.E.A.; Bryngelsson, T.; Bekele, E. Geographic and altitudinal allozyme variation in sorghum (Sorghum bicolor (L.) Moench) landraces from Ethiopia and Eritrea. Hereditas 2001, 135, 1–12. [Google Scholar]
- Barro-Kondombo, C.; Sagnard, F.; Chantereau, J.; Deu, M.; vom Brocke, K.; Durand, P.; Gozé, E.; Zongo, J.D. Genetic structure among sorghum landraces as revealed by morphological variation and microsatellite markers in three agroclimatic regions of Burkina Faso. Theor. Appl. Genet. 2010, 120, 1511–1523. [Google Scholar]
- Zongo, J.; Gouyon, P.; Sarr, A.; Sandmeier, M. Genetic diversity and phylogenic relations among Sahelian sorghum accessions. Genet. Resour. Crop Evol. 2005, 52, 869–878. [Google Scholar]
- Deu, M.; Sagnard, F.; Chantereau, J.; Calatayud, C.; Hérault, D.; Mariac, C.; Pham, J.-L.; Vigouroux, Y.; Kapran, I.; Traore, P.S.; et al. Niger-wide assessment of in situ sorghum genetic diversity with microsatellite markers. Theor. Appl. Genet. 2008, 103, 903–913. [Google Scholar]
- Brown, C.H. Development of agriculture in prehistoric Mesoamerica: The linguistic evidence. In Pre-Columbian Foodways; Staller, J.E., Carrasco, M., Eds.; Springer: Berlin, Germany, 2009; pp. 71–107. [Google Scholar]
- Benz, B.F. Maize in the Americas. In Histories of Maize. Multidisciplinary Approaches to the Prehistory, Linguistics, Biogeography, Domestication, and Evolution of Maize; Staller, J.E., Tykot, R.H., Benz, B.F., Eds.; Elsevier: San Diego, CA, USA, 2006; pp. 9–20. [Google Scholar]
- Matsuoka, Y.; Vigouroux, Y.; Goodman, M.M.; Sánchez, J.; Buckler, E.; Doebley, J. A single domestication for maize shown by multilocus microsatellite genotyping. Proc. Natl. Acad. Sci. USA 2002, 99, 6080–6084. [Google Scholar]
- Blake, M. Dating the initial spread of Zea mays. In Histories of Maize. Multidisciplinary Approaches to the Prehistory, Linguistics, Biogeography, Domestication, and Evolution of Maize; Staller, J.E., Tykot, R.H., Benz, B.F., Eds.; Elsevier: San Diego, CA, USA, 2006; pp. 55–72. [Google Scholar]
- Doolittle, W.E.; Mabry, J.B. Environmental mosaics, agricultural diversity, and the evolutionary adoption of maize in the American Southwest. In Histories of Maize. Multidisciplinary Approaches to the Prehistory, Linguistics, Biogeography, Domestication, and Evolution of Maize; Staller, J.E., Tykot, R.H., Benz, B.F., Eds.; Elsevier: San Diego, CA, USA, 2006; pp. 109–121. [Google Scholar]
- Vigouroux, Y.; Glaubitz, J.C.; Matsuoka, Y.; Goodman, M.M.; Sanchez, J.; Doebley, J. Population structure and genetic diversity of New World maize races assessed by DNA microsatellites. Am. J. Bot. 2008, 95, 1240–1253. [Google Scholar]
- Wellhausen, E.; Roberts, L.M.; Hernández, E. Races of Maize in Mexico: Their Origin, Characteristics and Distribution; Bussey Institute, Harvard University: Cambridge, MA, USA, 1952; p. 223. [Google Scholar]
- Mera, L.M. Diversificación y distribución reciente del maíz en México. In Origen y Diversificación del Maíz: Una Revisión Analítica; Kato, T.A., Mapes, C., Mera, L.M., Serratos, J.A., Bye, R.A., Eds.; Universidad Nacional Autónoma de México, Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: México City, Mexico, 2009; pp. 69–81. [Google Scholar]
- Hernández, E.; Alanís, G. Estudio morfológico de cinco nuevas razas de maíz de la Sierra Madre Occidental de México: Implicaciones citogenéticas y filogenéticas. Agrociencia 1970, 5, 3–30. [Google Scholar]
- Doebley, J. Molecular evidence and the evolution of maize. Econ. Bot. 1990, 44, 6–27. [Google Scholar]
- Benz, B.F. Diversidad y distribución prehispánica del maíz mexicano. Arqueología Mexicana 1997, 5, 17–23. [Google Scholar]
- Bird, R.M. Maize evolution from 500 B.C. to the present. Biotropica 1980, 12, 30–41. [Google Scholar] [CrossRef]
- Louette, D.; Charrier, A.; Berthaud, J. In situ conservation of maize in Mexico: Genetic diversity and maize seed management in a traditional community. Econ. Bot. 1997, 51, 20–38. [Google Scholar]
- Berthaud, J.; Pressoir, G.; Ramírez-Corona, F.; Bellon, M.R. Farmers management of maize landrace diversity. A case study in Oaxaca and beyond. In Proceedings of The Seventh International Symposium on the Biosafety of Genetically Modified Organisms, Beijing, China, 10–16 October 2002; International Society for Biosafety Research, China National Center for Biotechnology Development: Beijing, China, 2002; pp. 79–88. [Google Scholar]
- Pressoir, G.; Berthaud, J. Patterns of population structure in maize landraces from the Central Valleys of Oaxaca in Mexico. Heredity 2004, 92, 88–94. [Google Scholar]
- Pressoir, G.; Berthaud, J. Population structure and strong divergent selection shape phenotypic diversification in maize landraces. Heredity 2004, 92, 95–101. [Google Scholar]
- Benz, B.F.; Perales, H.R.; Brush, S.B. Tzeltal and Tzotzil farmer knowledge and maize diversity in Chiapas, Mexico. Curr. Anthropol. 2007, 48, 289–300. [Google Scholar]
- Bellon, M.R.; Brush, S.B. Keepers of maize in Chiapas, Mexico. Econ. Bot. 1994, 48, 196–209. [Google Scholar]
- Louette, D.; Smale, M. Farmers’ seed selection practices and traditional maize varieties in Cuzalapa, Mexico. Euphytica 2000, 113, 24–41. [Google Scholar]
- Dyer, G.A.; Taylor, J.E. A crop population perspective on maize seed systems in Mexico. Proc. Natl. Acad. Sci. USA 2008, 105, 470–475. [Google Scholar]
- Patal, F.; García, P.O.; Espantzay, C. Rujunamaxik ri Kaqchikel Chi'. Variación Dialectal en Kaqchikel; Cholsamaj: Antigua, Guatemala, 2000. [Google Scholar]
- Bellon, M.R.; Hodson, D.; Hellinc, J. Assessing the vulnerability of traditional maize seed systems in Mexico to climate change. Proc. Natl. Acad. Sci. USA 2011, 108. [Google Scholar] [CrossRef]
- Polanyi, K. The Great Transformation: The Political and Economic Origins of Our Time; Beacon Press: Boston, MA, USA, 1944. [Google Scholar]
- Laville, J.-L. Granovetter et la nouvelle sociologie économique. In Sociologie Economique; Granovetter, M., Ed.; Le Seuil: Paris, France, 2008; pp. 11–32. [Google Scholar]
- Granovetter, M. A Theoretical agenda for economic sociology. In The New Economic Sociology: Developments in an Emerging Field; Guillen, M., Collins, R., England, P., Meyer, M., Eds.; Russell Sage Foundation: New York, NY, USA, 2002; pp. 35–60. [Google Scholar]
- Granovetter, M. The strength of weak ties. Am. J. Sociol. 1973, 78, 1360–1380. [Google Scholar]
- Longley, C. A social life of seeds: Local management of crop variability in North-Western Sierra Leone. Ph.D. Dissertation, University of London, London, UK, 2000. [Google Scholar]
- Barth, F. Ethnic groups and Boundaries; Little, Brown & Co: Boston, MA, USA, 1969. [Google Scholar]
- Murdock, G.P.; White, D.R. Standard Cross-Cultural Sample; Working Papers Series; Social Dynamics and Complexity, Institute for Mathematical Behavioral Sciences, University of California, Irvine: Irvine, CA, USA, 2006, On-line edition pp. 329–369. Available online: http://eclectic.ss.uci.edu/~drwhite/worldcul/SCCS1969.pdf (accessed on 1 November 2011).
- Korotayev, A. Form of marriage, sexual division of labor, and postmarital residence in cross-cultural perspective: A reconsideration. J. Anthropol. Res. 2003, 59, 69–89. [Google Scholar]
- Holden, C.; Mace, R. Spread of cattle led to the loss of matrilineal descent in Africa: A coevolutionary analysis. Proc. R. Soc. B 2003, 270, 2425–2433. [Google Scholar]
- Cavalli-Sforza, L.L.; Feldman, M. Cultural Transmission and Evolution: A Quantitative Approach; Princeton University Press: Princeton, NJ, USA, 1981. [Google Scholar]
- Mace, R.; Holden, C.J. A phylogenetic approach to cultural evolution. Trends Ecol. Evol. 2005, 20, 116–121. [Google Scholar]
- Holden, C. Bantu language trees reflect the spread of farming across Sub-saharan Africa: A maximum parsimony analysis. Proc. R. Soc. B 2002, 269, 793–799. [Google Scholar]
- Gray, R.D.; Greenhill, S.J.; Ross, R.M. The pleasures and perils of Darwinizing culture (with phylogenies). Biol. Theory 2007, 2, 360–375. [Google Scholar]
- Grimes, B.F. Ethnologue; SIL International: Dallas, TX, USA, 2000. [Google Scholar]
- Hewlett, B.S.; Silvestri, A.D.; Guglielmino, C.R. Semes and genes in Africa. Curr. Anthropol. 2002, 43, 313–321. [Google Scholar]
- Steward, J.H. Theory of Culture Change, the Methodology of Multilinear Evolution; University of Illinois Press: Urbana, IL, USA, 1955. [Google Scholar]
- Reyes-García, V.; Broesch, J.; Calvet-Mir, L.; Fuentes-Peláez, N.; McDade, T.W.; Parsa, S.; Tanner, S.; Huanca, T.; Leonard, W.R.; Martínez-Rodríguez, M. Cultural transmission of ethnobotanical knowledge and skills: An empirical analysis from an Amerindian society. Evol. Hum. Behav. 2009, 30, 274–585. [Google Scholar]
- Lozada, M.; Ladio, A.; Weigandt, M. Cultural transmission of ethnobotanical knowledge in a rural community of Northwestern Patagonia, Argentina. Econ. Bot. 2006, 60, 374–385. [Google Scholar]
- Ohmagari, K.; Berkes, F. Transmission of indigenous knowledge and bush skills among the Western James Bay Cree women of Subartic Canada. Hum. Ecol. 1977, 25, 197–222. [Google Scholar]
- Hewlett, B.S.; Cavalli-Sforza, L.L. Cultural transmission among Aka Pygmies. Am. Anthropol. 1986, 88, 922–934. [Google Scholar]
- Jarvis, D.I.; Brown, A.H.D.; Cuong, P.H.; Collado-Panduro, L.; Latournerie-Moreno, L.; Gyawali, S.; Tanto, T.; Sawadogo, M.; Mar, I.; Sadiki, M.; et al. A global perspective of the richness and evenness of traditional crop-variety diversity maintained by farming communities. Proc. Natl. Acad. Sci. USA 2008, 105, 5326–5331. [Google Scholar]
- Romney, A.K.; Weller, S.C.; Batchelder, W.H. Culture as consensus: A theory of culture and informant accuracy. Am. Anthropol. 1986, 88, 313–338. [Google Scholar]
- Reyes-García, V.; Martí, N.; McDade, T.; Tanner, S.; Vadez, V. Concepts and methods in studies measuring individual ethnobotanical knowledge. J. Ethnobiol. Ethnomed. 2007, 27, 182–203. [Google Scholar]
- Boster, J.S. Selection of perceptual distinctiveness: Evidence from Aguaruna cultivars of Manihot esculenta. Econ. Bot. 1985, 39, 310–325. [Google Scholar]
- Duputié, A.; Massol, F.; David, P.; Haxaire, C.; McKey, D. Traditional Amerindian cultivators combine directional and ideotypic selection for sustainable management of cassava genetic diversity. J. Evol. Biol. 2009, 22, 1317–1325. [Google Scholar]
- Berlin, B. Folk systematics in relation to biological classification and nomenclature. Annu. Rev. Ecol. Syst. 1973, 4, 259–271. [Google Scholar]
- Berlin, B.; Breedlove, D.E.; Raven, P.H. The Principles of Tzeltal Plant Classification; Academic Press: New-York, NY, USA; London, UK, 1974. [Google Scholar]
- Berlin, B.; Breedlove, D.E.; Raven, P.H. General principles of classification and nomenclature in folk biology. Am. Anthropol. 1973, 75, 214–242. [Google Scholar]
- Martin, M.A. L’ethnobotanique, science per se? A propos d’un livre de B. Berlin, D.E. Breedlove, P.H. Raven “The principles of Tzeltal plant classification”. J. Agric. Trop. Bot. Appl. 1975, 22, 237–276. [Google Scholar]
- Bourdeix, R.; Leclerc, C.; Thampan, P.H.; Baudouin, L.; Joly, H.I. Modern and natural coconut hybrids in southern India : Natural, technical and social facts. J. Ethnobiol. 2008, 28, 39–54. [Google Scholar]
- Haudricourt, A.G. Nature et culture dans la civilisation de l'igname: L'origine des clones et des clans. Homme 1964, 4, 93–104. [Google Scholar]
- Nuijten, E.; Almekinders, C.J.M. Mechanisms explaining variety naming by farmers and name consistency of rice varieties in the Gambia. Econ. Bot. 2008, 62, 148–160. [Google Scholar]
- Almekinders, C.J.M.; Louwaars, N.P.; Bruijn, G.H. Local seed systems and their importance for an improved seed supply in developing contries. Euphytica 1994, 78, 207–216. [Google Scholar]
- Mauss, M. Essai sur le don, forme et raison de l'échange dans les sociétés archaïques. In Sociologie et Anthropologie; Mauss, M., Ed.; Presses Universitaires de France: Paris, France, 1950; pp. 143–279. [Google Scholar]
- Delaunay, S.; Tescar, R.-P.; Oualbego, A.; Vom Brocke, K.; Lançon, J. La culture du coton ne bouleverse pas les échanges traditionnels de semences de sorghos. Cah. Agric. 2008, 17, 189–194. [Google Scholar]
- Smale, M.; Aguirre, A.; Bellon, M.R.; Mendoza, J.; Rosas, I.M. Farmer Management of Maize Diversity in the Central Valleys of Oaxaca, Mexico; CIMMYT/INIFAP: Mexico D.F., Mexico, 1999. [Google Scholar]
- Badstue, L.B.; Bellon, M.R.; Berthaud, J.; Juárez, X.; Rosas, I.M.; Solano, A.M.; Ramírez, A. Examining the role of collective action in an informal seed system: A case study from the Central Valleys of Oaxaca, Mexico. Hum. Ecol. 2006, 34, 249–273. [Google Scholar]
- McGuire, S.J. Vulnerability in farmer seed systems: farmer practices for coping with seed insecurity for sorghum in Eastern Ethiopia. Econ. Bot. 2007, 616, 211–222. [Google Scholar]
- Janssen, W. Bean Seed Supply Systems for Small Farmers: The Need for Primary Data in Institutional Design; CIAT: Cali, Colombia, 1989. [Google Scholar]
- Philippson, G.; Bahuchet, S. Cultivated crops and bantu migrations in Central and Eastern Africa: A Linguistic Approach. In The Growth of Farming Communities in Africa from the Equator Southwards; Sutton, J., Ed.; British Institute in Eastern Africa: Nairobi, Kenya, 1994; pp. 103–120. [Google Scholar]
- Zerega, N.J.C.; Ragon, D.; Motley, T.J. Complex origins of breadfruit (Artocarpus altilis, Moraceae): Implications for human migrations in Oceania. Am. J. Bot. 2004, 91, 760–766. [Google Scholar]
- Perrier, X.; de Langhe, E.; Donohue, M.; Lentfer, C.; Vrydaghs, L.; Bakry, F.; Carreel, F.; Hippolyte, I.; Horry, J.-P.; Jenny, C.; et al. Multidisciplinary perspectives on banana (Musa spp.) domestication. Proc. Natl. Acad. Sci. USA 2011, 108, 11311–11318. [Google Scholar]
- Hanotte, O.; Bradley, D.G.; Ochieng, J.W.; Verjee, Y.; Hill, E.W.; Rege, J.E.O. African pastoralism: Genetic imprints of origins and migrations. Science 2002, 296, 336–339. [Google Scholar]
- Cymbron, T.; Freeman, A.R.; Malheiro, M.I.; Vigne, J.-D.; Bradley, D.G. Microsatellite diversity suggests different histories for Mediterranean and Northern European cattle populations. Proc. R. Soc. B 2005, 272, 1837–1843. [Google Scholar]
- Zeder, M.A.; Emshwiller, E.; Smith, B.D.; Bradley, D.G. Documenting domestication: The intersection of genetics and archeology. Trends Genet. 2006, 22, 139–155. [Google Scholar]
- Maffi, L. Linguistic, cultural, and biological diversity. Annu. Rev. Anthropol. 2005, 34, 599–617. [Google Scholar]
- Saussure De, F. Cours de Linguistique Générale; Grande Bibliotèque Payot: Paris, France, 1916. [Google Scholar]
- Sagnard, F.; Barnaud, A.; Deu, M.; Barro, C.; Luce, C.; Billot, C.; Rami, J.-F.; Bouchet, S.; Dambele, D.; Pomies, V.; et al. Analyse multiéchelle de la diversité génétique des sorghos: Compréhension des processus évolutifs pour la conservation in-situ. Cah. Agric. 2008, 17, 114–121. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Leclerc, C.; Coppens d’Eeckenbrugge, G. Social Organization of Crop Genetic Diversity. The G × E × S Interaction Model. Diversity 2012, 4, 1-32. https://doi.org/10.3390/d4010001
Leclerc C, Coppens d’Eeckenbrugge G. Social Organization of Crop Genetic Diversity. The G × E × S Interaction Model. Diversity. 2012; 4(1):1-32. https://doi.org/10.3390/d4010001
Chicago/Turabian StyleLeclerc, Christian, and Geo Coppens d’Eeckenbrugge. 2012. "Social Organization of Crop Genetic Diversity. The G × E × S Interaction Model" Diversity 4, no. 1: 1-32. https://doi.org/10.3390/d4010001