Dispersal, Isolation, and Interaction in the Islands of Polynesia: A Critical Review of Archaeological and Genetic Evidence

1
Department of Anthropology, Southern Methodist University, P.O. Box 750336, Dallas, TX 75275-0336, USA
2
School of Geography, Archaeology and Environmental Studies, University of the Witwatersrand, Wits 2050, South Africa
*
Author to whom correspondence should be addressed.
Diversity 2017, 9(3), 37; https://doi.org/10.3390/d9030037
Submission received: 11 August 2017
/
Revised: 2 September 2017
/
Accepted: 11 September 2017
/
Published: 14 September 2017
(This article belongs to the Special Issue Ancient DNA)
Abstract
:Integration of archaeology, modern genetics, and ancient DNA holds promise for the reconstruction of the human past. We examine the advances in research on the indigenous peoples of Polynesia to determine: (1) what do archaeological and genetic data (ancient and modern DNA) tell us about the origins of Polynesians; and, (2) what evidence is there for long-distance travel and contacts between Polynesians and indigenous populations of the Americas? We note that the general dispersal pattern of founding human populations in the remote islands of the Pacific and long-distance interaction spheres continue to reflect well-established models. New research suggests that the formation of an Ancestral Polynesia Culture in Western Polynesia may have involved differential patterns of dispersal followed by significant later migrations. It has also been suggested that the pause between the settlement of Western and Eastern Polynesia was centuries longer than currently thought, followed by a remarkably rapid pulse of island colonization. Long-distance travel between islands of the Pacific is currently best documented through the sourcing of artifacts, while the discovery of admixture of Native American DNA within the genome of the people from Easter Island (Rapa Nui) is strong new evidence for sustained contacts between Polynesia and the Americas.
1. Introduction
The biogeographical metaphor of islands ‘as natural laboratories’ has been useful in focusing research on the long-term histories of the people of Polynesia and elsewhere [1,2,3]. In contemporary ecological and archaeological research, the metaphor has been replaced by one of ‘islands as model systems’ for understanding human ecodynamics [4,5]. Here, we review advances in archaeological and genetic research over the past decade to address two questions that have been central to the region: (1) what do archaeological and genetic data (ancient & modern DNA) tell us about the origins of Polynesians; and, (2) what do these same lines of evidence tell us about long-distance contacts between Polynesians and indigenous populations of the Americas before European contact? In principle, the integration of evidence from archaeology, genetics of contemporary populations, and ancient DNA holds significant promise for the reconstruction of the human past [6,7,8,9]. In practice, the inherent limitations to each of these approaches, and the large and growing technical knowledge required to read across disciplines, makes identifying and reconciling the differences problematic.
Today, we see that the initial emphasis on isolation by scholars is evolving to more explicitly include how interactions between island communities shaped human history [10]. As advances in techniques used as archaeological and genetic metrics of interaction become a regular part of how we reconstruct the human past, we find that for humanity it is not at all extraordinary to find great natural isolation paired with strong connectivity. This shift in our thinking about islands is one way for archaeology to replace popular hyper-diffusionist and other pseudoscientific theories that persist in all areas of the world with sound science on human diversity.
2. Origins of the Polynesians
In the 18th century, British Captain James Cook was awed by the peoples of the Polynesian “nation” spread across thousands of kilometers yet sharing common features, languages, and customs (Figure 1). The term “Polynesia”—meaning many islands—came later in the 19th century with racial categories for Pacific Islanders drawing lines between Melanesia, Micronesia, and Polynesia [11]. In contemporary scholarship, the term Polynesia lives on but with modifications based on historical linguistics, ethnography, and geography. The ‘Polynesian triangle’, as it is known, is the region between the Hawaiian Islands, Rapa Nui (Easter Island), and Aotearoa (New Zealand) where most of 40 island communities that speak a Polynesian language and carry on a Polynesian culture live today.
Historical linguists have long recognized shared commonalities that allow us to further divide this region into Western Polynesia, including the islands of Tonga and Sāmoa, and Eastern Polynesia. Eastern Polynesia can be further divided, based on geographic, cultural, and linguistic criteria, into Central Eastern Polynesia (Society Islands, Cook Islands, Marquesas) and Marginal Eastern Polynesia, as defined by the three island groups at the points of the triangle. In addition to these three categories (Western, Central Eastern, and Marginal Eastern) there is a group of islands outside of the triangle called the Polynesian Outliers that notably includes the island of Tikopia. At the time of European contact, these islands were home to communities speaking a Polynesian language. A small number of islands within Polynesia show signs of human settlement in the past but were unoccupied at the time of European contact; these are sometimes referred to as the ‘Mystery Islands’ [12].
The question of the origins of Polynesians in Western scholarship tracks the evolution of thinking about cultures more generally, but relevant here are two intellectual developments that have been foundational to increasingly sophisticated models of culture history based on historical linguistics, comparative ethnography, human biology, and archaeology. First, there is a broad consensus that a single archaeological culture called Lapita—ultimately related to populations in Asia and New Guinea—is responsible for pushing beyond the islands of the Western Pacific occupied during the Pleistocene era, called Near Oceania, into the more isolated islands of Remote Oceania. The geographic distribution of the Lapita sites shows a common ancestry of people across the line drawn by d’Urville between Melanesia and Polynesia. Second, Kirch and Green [13] have made the case for the development of an Ancestral Polynesian Culture (APC) as derived from the Lapita in Western Polynesia prior to the settlement of Central and then Marginal Eastern Polynesia. Strong advocates for an historical anthropology, they used a “triangulation method” for making these distinctions between homologies, analogies, and synologies (borrowing) [14], based on the tacking between historical linguistics, comparative ethnography, and archaeology.
Over the past decade, two additional hypotheses have been proposed that, if supported by the balance of evidence, would be of great significance to human diversity in the origins of Polynesian peoples. The first proposes that multiple founding groups settled Polynesia [15,16,17,18]. Most syntheses suggest that the first people to leave Near Oceania and settle the islands of Tonga and Sāmoa were part of the larger Eastern Lapita archaeological culture, and it was this community that evolved in situ to become Ancestral Polynesian Culture. Several researchers have raised the possibility that the two island groups’ early culture histories were distinct, and that in Sāmoa, initial settlement was unsuccessful, and followed by significant later migrations during the post-Lapita era from Micronesia. If this were the case, than the implication is that some of the social and cultural traits that have been attributed to local development may have been the result of migration and/or diffusion.
The second proposes that Eastern Polynesia was settled recently and rapidly [19]. A new synthesis of radiocarbon dates posits that the pause in settlement from Western to Eastern Polynesia was centuries longer than currently thought, and that the islands of Eastern Polynesia were settled in a rapid dispersals with a short 70 to 265 year long pause between Central Eastern Polynesia and Marginal Eastern Polynesia. If this were the case, than cultural, linguistic, and social diversification was also more recent and rapid than previously thought.
2.1. Archaeology
With the invention of radiocarbon dating archaeologists have used absolute dates to estimate when people first arrived in a given island group. On the broadest time scale, radiometric dating has told us that the colonization of Pacific Islands did not progress in a regular or predictable fashion, and thus in retrospect, it is sometimes described as having occurred in ‘pulses’ and ‘pauses’. The pulse-pause pattern relevant here began with the rapid colonization of the islands of Remote Oceania by Lapita peoples, followed by a pause in eastward progression in what would become Western Polynesia, and then the last pulse of movement out into Eastern Polynesia. Below, we address how this story has been modified, refined, and revised over the past ten years in the archaeology of Polynesia, beginning with a trend that we call the ‘new chronometric hygiene’.
2.1.1. The New Chronometric Hygiene
The term ‘chronometric hygiene’ entered into literature more than twenty years ago in Spriggs and Anderson’s [20] call for more strict criteria for accepting radiocarbon dates based on their context, material, and quality of the laboratory where the analysis was performed. This has shifted the bar higher for accepting new results and has been the inspiration for re-dating sites where early dates had been reported. Allen and Huebert [21] have reviewed which plant taxa are likely to yield good results. Others have continued to refine how best to calibrate marine carbon results [22,23], which are especially important given how commonly fish and shellfish are found in Oceania, and how variable the marine calibration is across space. Rieth and Athens [24] have discussed the best practices in collecting and reporting context of samples bound for radiocarbon dating and Smith [25] has proposed a method to regularize the placement of radiocarbon results within the chronology of New Zealand.
A number of islands have had thorough critical reviews of reported radiocarbon dates in order to sort results into three categories: reliable results that meet all our current standards for best practices, dates that may be correct but we cannot say for certain, and those that must be outright rejected for the Hawaiian Islands, see [26,27,28]; for Sāmoa, see [29]; and, for Rapa Nui, see [30]. By far, the largest category is the second. These are dates for which we lack the requisite information about context or material, or which have laboratory results that have large error bars, and so exist in scholarly limbo—not enough information to accept them, nor enough to outright reject them.
Better practices in selecting material to radiocarbon date, the critical review of reported dates, and the re-dating of sites once thought to be early in local sequences are in-line with the original ‘chronometric hygiene’ as advocated by Spriggs and Anderson [20]. One solution to the small number of acceptable dates—called here the ‘new chronometric hygiene’—has grown in popularity. It creates strict criteria for accepting dates and then applies a statistical model to compensate for the small sample size. Rieth and Cochrane [18] distinguish between two types of statistical models, pooling dates e.g., [19], and Bayesian models e.g.,[31]. The use of statistical reasoning is of course a part of the interpretation of all radiometric results, and is not the thing that sets these apart from the ‘re-date and revise’ response we have seen following the original call for chronometric hygiene. We begin by presenting the historical models proposed by the new chronometric hygiene and then discuss some of the underlying theories that have been driving this line of inquiry, and the counter-evidence.
The highest profile example of the new chronometric hygiene is a pooled dates study by Wilmshurst et al.’s [19] (p. 1815) proposing a “recent and rapid” colonization of Eastern Polynesia. The authors created a large database of published radiocarbon dates (n = 1434) from “at least 45 East Polynesian islands covering all of the major archipelagos” [19] (p. 1815). After rejecting 86% of these dates based on a number of criteria (context, material, error range), they accept 207 dates (Class 1). The dates are then pooled by island group and their probabilities summed. The result is a shorter ‘pause’ between the settlement of Western Polynesia and Central Eastern Polynesia, and the subsequent settlement of Marginal Eastern Polynesia (70–265 years). This same method was applied on a database of dates from Hawai’i Island [28].
Critics have pointed out that Wilmshurst et al.’s [19] approach has some fundamental flaws and yields dates that are definitively post-colonization. Mulrooney et al. [32] found inaccuracies in their summary of published radiocarbon dates, exclusion of valid colonization dates, and biases in their statistical analysis [33]. Kirch [34] (p. 16), notes their approach “depends on a fairly extreme form of chronometric hygiene”. Further, Wilmshurst et al. [19] rely heavily on New Zealand as their model for approaching the problem of dating colonization. Indeed, ~50% of Class 1 dates are from New Zealand because colonization era sites there have high archaeological visibility and new sites are regularly found; on average two to three new sites per year (Source: ArchSite, archsite.org.nz). These conditions are excellent for dating colonization no matter what method is used. For example, Dye [35], who also echoes the widely held concern that “the criteria employed by Wilmshurst et al. appear to have been too strict, resulting in an estimate of the settlement date that is too young”, recently conducted a Bayesian analysis for the colonization of New Zealand that would narrow the window to within a 40 year span (1270–1309, 95% highest posterior density). This incredible precision is not an accident; it is based on pools of pre-colonization dates (n = 10) and post-colonization dates (n = 108) that are many times larger than any other estimate for island colonization. Wilmshurst et al.’s expectation that the same analytical tool will achieve identical results on the other island groups is simply unrealistic.
2.1.2. Chronology of Settlement of Western Polynesia
It has been an active decade in the pursuit of Polynesia’s earliest communities. Not long after the publication of the recent and rapid model [19], Burley et al. [36] presented evidence for the earliest secure dates of human activity in Polynesia, specifically uranium series dates on coral artifacts (2838 ± 8 BP) and supporting radiocarbon dates from Nukuleka on Tongatapu Island in the Ha‘apai Group in Tonga [37]. Burley and colleagues have also worked on the problem of sketching out the earliest communities across Tonga, noting that there is good reason to believe colonization was simultaneous across the three major island groups within the Tonga, although Vava‘au group dates are slightly later [38], and the strongest evidence for early human presence on Nuitongatapu, at the archipelago’s northern extreme, is the presence of volcanic glass from there showing up in the earliest deposits on Tongatapu. In Tonga, founding period sites bearing Lapita pottery give way to a Polynesian Plainware phase, followed by a loss of ceramics during a formative phase, before there are clear material signs of Tongan chiefdoms.
The question of Sāmoa’s earliest communities is more problematic. In Sāmoa, the Mulifanua site remains the sole evidence for a founding Lapita population (~2500 BP), with a gap of 350 years until there is a strong signal of human occupation (~1700 BP). Rieth and Cochrane [18] (pp. 337–338) cite years of research that have failed to fill this gap as evidence that “it is more likely that the lack of Lapita and immediately post-Lapita sites is not a function of site visibility and research intensity, but represents a severely diminished or absent prehistoric population”. This position is supported by a spatial analysis by Rieth et al. [39] and by Cochrane et al.’s [17] detailed study of coastal geomorphology that suggests that the attractive coastal plains in Sāmoa may not have formed until later in its cultural history.
One area where researchers largely agree that archaeological visibility has also proved to be an impediment has been in the identification of Ancestral Polynesian Culture (APC) [13] (p. 64). Kirch and Green’s [13] book-length treatise on the origins of the post-settlement ethno-genesis set out a method to triangulate—through historical linguistics, comparative ethnography, and archaeology—Polynesian culture history distinct from other descendants of Lapita peoples. Given the scale of their synthesis, which has direct implications for islands in Western Polynesia and knock-on effects for how we view Eastern Polynesia’s past, it is unsurprising that it has yet to be systematically tested with new evidence. Smith [40] has noted conservativeness in material culture as being at odds with the notion of an APC. However, conservativeness in material culture, such as fishing gear, is something that can be seen across the historic era [16], despite it being a period of intense social and cultural change. Conservativeness in material culture across Oceania is a particularly intractable problem in reconstructing culture histories of the Polynesian outliers. Davidson [41] (p. 1) has noted that if Nukuoro “had been uninhabited at the time of European contact it would never have been recognised as a Polynesian outlier” despite years of research. The notable exception to this is Tikopia’s Tuakamali Phase (AD 1200 to 1800) [42] where the novel introduction of Sāmoan-styled adzes gives us a material signal of new arrivals.
New research in Western Polynesia broadly, and the 350 year long gap in Sāmoa’s early record specifically, have naturally led archaeologists to question the notion that the early culture histories of Tonga and Sāmoa were “interwoven and virtually homogeneous” as they are sometimes presumed to have been [16] (p. 3). Burley and Addison [16] cite contrasting patterns in current evidence of settlement patterns, ceramics, adze style, and the use of shell as pointing to two scenarios: (1) the founding communities of Sāmoa diverged strongly and rapidly in terms of material culture from the Eastern Lapita origins that they share with Tonga’s founding communities; or, (2) that there was a second colonization event ~2500 BP that did not originate from Tonga. Here, they draw on Addison and Matisoo-Smith’s [15] recent proposition that Green’s Triple-I model (intrusion, innovation, integration) for the origins of Lapita peoples may help to explain the origins of Polynesians. The Western Polynesian Triple-I model, as they call it, begins with the intrusion of Lapita peoples into the unoccupied islands, followed by the innovation of some traits in situ. What sets the model apart is that the authors invoke an undocumented movement of people through the low islands of Micronesia ~1500 BP and credit them with introducing new lineages of dogs, rats, and chickens, as well as an increase in evidence for occupation of Sāmoa.
The animal commensal evidence used to support the Western Polynesian Triple-I model relies on several shaky premises, including that ‘patchy’ evidence of domesticates is not an artifact of taphonomic processes or sampling. In Table 1, we give a summary of all modern and ancient commensal animal genetic data for four taxa: rat (Rattus exulans), pig (Sus scrofa), chicken (Gallus gallus), and dog (Canis lupus fimiliaris) (after [43], with additions from [44,45]). There is clearly far more coverage for Melanesia and Eastern Polynesia, the two spatial-temporal endpoints of the dispersal of people in to Polynesia. The regions that are of direct interest for accepting or rejecting the hypothesis—Western Polynesia and Micronesia—are represented by only 35 ancient and modern samples, or less than 6% of published data, with no coverage for dogs. Oversampling is a problem in both ancient and modern data, for example 73% of ancient rat data is from New Zealand and 98% of modern chicken data is from Vanuatu. We also note that examples of modern and archaeological pig samples from New Zealand represent a post-European introduction. Commensal plants—(banana (Musa sp.), bottle gourd (Lagenaria siceraria), breadfruit (Artocarpus spp.), coconut (Cocos nucifera), kava (Piper methysticum), paper mulberry (Broussoneria papyrifera), sweet potato (Ipomoea batatas), taro (Colocasia esculenta), and ti (Cordyline fruticosa)—are equally problematic in that the data is exclusively from modern or herbarium specimens, with 97% to 21% of published samples from a single island or island group [43,46,47,48].
Another major study on origins, Burley’s [49] synthesis of the polygenesis of Fijian culture and society is founded on the accumulation of evidence that the division between Melanesia and Polynesia has its roots in the differential timing and source of founding populations and post-settlement movements. This study, like the West Polynesian Triple-I Model, asks archaeologists to take a hard look at the evidence to date and reject the premise that the islands of Remote Oceania have a high degree of uniformity, and once settled, were largely isolated from each other. As Kirch [50] has pointed out, we cannot maintain a Wave of Advance model for the colonization of Fiji and Western Polynesia. We also note that in Micronesia, the ‘breadfruit revolution’ hypothesis [51] adds another layer of complexity, explaining common traits seen across the region by a boom in population in the Eastern Caroline Islands following the invention of hybrid tree crops; in effect, an e pluribus unum (one out of many) or multi-phyletic origins for Micronesian cultures.
2.1.3. Chronology of Settlement of Eastern Polynesia
The flurry of publications on the settlement of Eastern Polynesia belies the wide consensus on what has been dubbed the ‘short chronology’. Archaeologists in Central Eastern Polynesia have been hard at work documenting foundation communities [52,53,54,55,56]. The settlement of island groups like the Society Islands, Marquesas, and Cook Islands are particularly important to piecing together the sequence of settlement of the remainder of unoccupied Eastern Polynesia; however, there remain large gaps in our knowledge and ambiguous results. For example, Allen [57] (p. 1) provides a thorough review of the current evidence for the settlement of the Hawaiian Islands, judging the evidence “sufficient to continue entertaining the possibility that the Marquesas Islands were a departure point for the Polynesian settlers of Hawai’i, although many uncertainties remain”.
In the furthest corners of the region called Marginal Eastern Polynesia, the date for the first New Zealanders remains unchanged, ca. AD 1250. New work at the Wairau Bar site has lent further support to the site’s status as among the earliest in New Zealand, occupied when there was movement of people from tropical Polynesia and the now extinct moa was still present on the landscape [58,59,60,61,62,63]. Anderson [64] reported that human presence on the subantarctic Auckland Islands during New Zealand’s Early Period, which together with dates from other groups (Kermadecs, Norfolk Islands, and somewhat later, the Chathams) point to a near simultaneous arrival of people in the southern extreme of Marginal Eastern Polynesia.
Athens and colleagues [31,35,65] have made much progress in specifying the date of colonization of the Hawaiian Islands from ‘after AD 800’ to between AD 940–1129 (95% highest posterior density); an estimate that has been reached by Kirch [34] as well. The precision in these estimates is in part thanks to the largely accepted hypothesis that the introduction of the Polynesian rat (Rattus exulans) caused a massive transformation of lowland vegetation by eating the seeds of indigenous palms (Pritchardia sp.). Hunt and Lipo [66] extended this to the case of Rapa Nui, arguing that their dates from the Anakena site mark the onset of human occupation and rapid demise of the palm forests there around AD 1200. This would put the settlement of Rapa Nui between the dates from Hawai’i and those from New Zealand. Not all scholars are convinced, but a date of settlement that is not a great deal earlier than Hawai’i and not a great deal later than New Zealand is likely. The ‘recent and rapid’ dispersal postulated by [19] cites the increasing the similarity in dates of colonization for Marginal Eastern Polynesia as supporting their position.
The long-standing objection by Anderson about the presumed ability of navigators [67], has entered a new chapter as more, and more refined, data are known regarding El Nino/Southern Osculation (ENSO) and how it may have influenced past sailing conditions. In a number of publications, Anderson has promoted the notion of ENSO forcing e.g., [68,69], which put simply, explains the pauses-and-pulses in colonization and long-distance voyaging by reference to climate-induced conditions that compelled people to increase (pulse) or decrease (pause) their activity, see also [70]. Computer models of ancient sailing conditions have in some cases confirmed what we have suspected based on archaeological and/or linguistic evidence, as such identifying Sāmoa as a natural bottleneck location for the pause-pulse in west-to-east colonization across the Pacific [71]. Other models have highlighted alternative source locations for settlement between sub-regions that have yet to be raised by archaeological, genetic, or linguistic based evidence [72,73]. ENSO forcing and proposed alternative routes to colonization are intriguing ideas, and with tighter chronologies and better computer models, we hope that these can either be accepted or rejected.
One study that has relevance across Polynesia is Walter et al.’s [74] application of the notion of a shift from a ‘colonizer’ to a ‘trader’ mode of interaction in New Zealand. The authors use the drop off in North Island obsidian (specifically Mayor Island Obsidian) found on South Island sites and the increase in South Island nephrite in North Island sites as indicative of a pattern seen at different timescales across the Pacific. In short, early communities naturally isolated from their external homeland interact freely and intensely with one another when establishing a viable founding population. Later, this ‘colonizer’ motivated interaction is replaced by fewer long-distance contacts and those contacts change in character to being one motivated by trade between established communities. Subsequent reexamination of South Island assemblages has found a remarkable diversity of North Island obsidians suggesting that the social divisions evident in later Māori history may be rooted in the Early Period [75,76]. New isotopic data from human bone at Wairau Bar, specifically dietary stable isotopes and 87Sr/86Sr, has produced results that have been interpreted as being indicative of high local mobility [77,78], sex-specific diets [79], and a split over whether the evidence supports the interpretation that the group includes the first generations of settlers from tropical Polynesia.
2.2. Modern DNA
Elsewhere, modern genetic data have demanded a significant revision of archaeologically derived models, not so in Polynesia. In Polynesian research, the accumulated genetic data are largely congruent with the archaeological and linguistic data. Blood proteins, globin genes, mitochondrial DNA, and recently gut bacteria e.g., [80], reinforce the standard model of human dispersal with the notable exception of previously unknown sex-specific patterns found in human genetics. Specifically, modern maternally inherited mitochondrial DNA (mtDNA) shows a strong connection with island Southeast Asia, and Taiwan, in particular, is consistent with archaeological models [81,82,83,84], with about 96% of mtDNA lineages in Polynesia being rooted in Asian populations and the rest being New Guinean in origin. In contrast, only about a third of the paternally inherited Y-chromosomes among Polynesians are Asian in origin, with the other two-thirds coming from New Guinea [85], see also [86]. Sex-biased dispersal has been documented by using genetic data to model other Neolithic expansions but we note that direct evidence from ancient DNA is far too limited to allow for a nuanced understanding of this general pattern.
Next generation sequencing of the full genome of modern people’s mitochondrial DNA has opened up new opportunities for specifying population histories that would otherwise remain hidden due to the limited variability in mtDNA in Polynesia. We note two promising discoveries but it is important to keep in mind, that, while our interest is in pre-contact trends in island colonization and interaction, that there are post-contact processes that influence the modern picture (demographic collapse, introduction of new mtDNA lineages), and there continues to be small sample sizes of mitochondrial genomes in Polynesia. At present, only 462 people’s mtDNA have been published with half from Western Polynesia (Tonga and Sāmoa, n = 99; other Western Polynesian islands, n = 118) and half from Eastern Polynesia (Central Eastern Polynesia represented by Cook Islands, n = 65; and Marginal Eastern Polynesia represented by the Hawaiian Islands, n = 160, and New Zealand, n = 20). With those caveats, the most recent revision of mtDNA haplogroups [87] clearly show an increase in the frequency of haplogroup B from Papua New Guinea and nearby islands at 60–40%, to over 80% in Fiji, and +90% among Polynesians (Figure 2). Within haplogroup B, the subgroup B4 accounts for nearly all individuals, but with the increased resolution allowed by sequencing complete mitochondrial genomes we can see that Fiji and Western Polynesia show the same frequencies of different B4 lineages, especially B4a1a, B4a1a1a and their sublineages (Figure 3). A 9-base pair deletion in mtDNA, part of a set of mutations, commonly known as the ‘Polynesian motif’, is a defining characteristic of the B4a1a1a group. Comparing Western Polynesia with Central Eastern Polynesia (Cook Islands), we see a much higher frequency of the B4a1a1m and B4a1a1m1 lineages that are found at low frequency across the Western Pacific. We also see higher frequencies of B4a1a1c, a lineage so far found only in modern Polynesia. Comparing Central Eastern Polynesia with Marginal Eastern Polynesia (the Hawaiian Islands), this lineage jumps again in frequency to account for most mtDNA. If these trends are rooted in pre-contact settlement and interaction, then they are good indicators of the effect of bottlenecks in the settlement of Eastern Polynesia out of Western Polynesia. We currently have no comparative datasets from Micronesia, however, the data as they are today can be explained without the introgression of novel mtDNA linages from other islands. In other words, modern mtDNA across Polynesia is consistent with the present model of human colonization of the Pacific.
2.3. Ancient DNA
Evidence from ancient DNA recovered from archaeological human remains from New Zealand is likewise consistent with the existing archaeologically derived model of settlement. Complete mitochondrial genome sequences of two individuals, and partial sequences of two more were recovered from human remains excavated from the site of Wairau Bar on the South Island of New Zealand [88]. These were then compared with a small study of mtDNA from 20 modern Māori (B4a1a1a3, n = 7; B4a1a1c, n = 6; B4a1a1a5, n = 4; B4a1a1a, n = 3). The resolution of mtDNA on all four sequences is sufficient to say they were members of the B4a1a1 group, the common clade of mtDNA across Marginal Eastern Polynesia. The two individuals with high DNA quality results included B4a1a1a3 and B4a1a1a, and two out of three mutations reported to be unique to modern Māori were found [89]. With such a small sample size it is difficult to interpret these beyond the generalization that some of the mtDNA sequences currently only found in Māori may have their origins either early in the settlement of New Zealand or perhaps in Central Eastern Polynesia in the generations immediately prior.
3. Long-Distance Contacts between Polynesians and Native Americas
It has been well established that there were contacts in the pre-European era between Polynesia and the Americas. The sweet potato, an American domesticated plant, has been found in Polynesia dated to AD 1000 [90], and has thus proved robust evidence in support of trans-Pacific contact see for example [91], although it should be noted that trans-Pacific natural dispersal continues to be evaluated [92]. A suite of research has made some extraordinary claims for new lines of evidence of more regular contact and a re-examination of the importance of diffusion to explain the changes in technology, culture, and society [93]. Specifically, Jones and Klar [93] present a hypothesis that Polynesian-American contact was responsible for the diffusion of a number of traits (sailing technology, fishhook style), linguistic isolate terms, and the well-established transfer of domesticated sweet potato (Ipomoea batatas). Following critiques from experts on the archaeology of Polynesia [70] and the Americas [94], the hypothesis has been developed through an edited volume [95].
Here, we focus on three high-profile cases of new claims that support Jones and Klar’s model of Polynesian-American contact: (1) direct evidence for transfer of the domesticated chicken (Gallus gallus) from Polynesia to South America [96,97,98,99,100,101,102]; and, (2) skulls of two Polynesians within collections of human remains attributed to a Native American group in Central-Eastern Brazil (Botocudo) [103]; and, (3) full genomic study of modern Rapa Nui indicating pre-European contact admixture with Native Americans [104].
3.1. Archaeology
The physical evidence of interactions between Polynesian communities has been the focus of intense study for many years, and over the past decade we have seen a massive leap forward in the geochemical matching of artifacts—primarily volcanic stone—to their likely source [105] and advances in studies of pottery in Western Polynesia [106]. The documentation of long-distance travel between the islands of Polynesia has seen some major advances, although still remains plagued by small sample sizes, in part due to the inherent rareness of well-traveled artifacts. Beginning on the largest scale, the material linking Western and Eastern Polynesia is still uncommon, and has some significant gaps. For example, there is no material that provides an unambiguous and direct link between Marginal Eastern Polynesia and Western Polynesia. Twenty years ago, 11 basalt artifacts found in the Cook Islands (on Mangaia, n = 4; Rarotonga, n = 3; Aitutaki, n = 2; and Mauke, n = 2) [107,108,109] were shown to have been made from material naturally found in Western Polynesia (basalt from Tutulia in Sāmoa). To that group, there has been added a single tentative identification of a basalt artifact found in Tonga that has been sourced to Society Islands [110]. There are of course isolated reports of Fijian pottery in the Marquesas [111], but as has been long established, pottery production ceased in Western Polynesia before the settlement of Eastern Polynesia.
New research has helped to define regional hubs of interaction within Polynesia—one in Western Polynesia and the other in Central Eastern Polynesia—that, as noted above, were linked with one another. A recent summary of the long distance movement of artifacts within, and beyond, the islands of Western Polynesia suggests that the movement of stone tools reflects late pre-contact political integration [110]. Clark et al. [110] used the distribution and the relative frequency of adzes and adze flakes, especially Tutulia basalt from Sāmoa, as a metric for the evolution of a powerful polity sometimes called the ‘Tongan Maritime Empire’. The high volume of stone being brought to the heart of the Tongan state was unprecedented and the geographic span of adzes speaks to the fact that this social shift was not exclusive to the Sāmoa-Tonga-Fiji exchanges noted in oral tradition, but was far reaching. In the case of Sāmoa, Cochrane and Reith [112] (p. 6), observe, “... lithic artefacts were transferred rarely and in very small proportions within Sāmoa or beyond the archipelago for the first 1500 years of prehistory. Beginning about 1200 calBP, intra-archipelago transfers of basalts increase slightly in frequency and in distance, and after another 300–400 years, basalts from Sāmoa, particularly Tutuila, are transferred across the central and south-west Pacific, although they never comprise more than about 10% of a lithic assemblage, except at Lapaha, Tonga”. After 1800 calBP, ceramics follow the same pattern noted in lithic transfers.
Not all material was transferred as widely, or with as high a frequency, as basalt adzes. For example, with rare exceptions, there is much less evidence for interisland movement of pottery or volcanic glass. Burley et al. [113] have investigated the sources of volcanic glass artifacts from sites in the main Tongan archipelago (Vava’u n = 30 artifacts; Ha’apai n = 12 artifacts; and Tongatapu n = 9 artifacts), Niuatoputapu (n = 17 artifacts), and Sāmoa (n = 48 artifacts). The geochemistry of natural volcanic glass sources of Niuatoputapu (located between Tonga and Sāmoa) come from Kirch’s excavations at the Nt 90 site (Kirch 1988, n = 75 grab samples across excavation context) and Sāmoa natural source material comes from sites on Tutuila (n = 10 samples) and ′Upolu (n = 38). Artifacts made from Niuatoputapu glass were found in small quantities in archaeological sites across Tonga; a distance of 600 km. No glass from this source was found in Sāmoa, however, one piece of Sāmoan volcanic glass was found on one of the northern islands of the main Tongan island groups (Vava’u group), accounting for 1 out of 9 samples of geochemically assayed artifacts from the site of Vana on the island of Pangaimotu. This artifact was found in a post-600 BP context (aceramic), and attributed to transfer during the heyday of the Tongan Maritime Empire. Research on pottery in Sāmoa [114] and Tonga [106] further underline the limited evidence for the movement of goods between islands.
In addition to Western Polynesia, we have also seen advances in defining the interaction within a second geographic sub-section of Polynesia; the islands in Central Eastern Polynesia. Allen and colleagues have identified the Eiao Island basalt source from the Marquesas Islands as far away as the Line Islands, Southern Cook Islands, and Mangareva, and across the Tuamotus [41]. Collerson and Weisler [115] used surface collected adzes in the Tuamotu Islands to document the presence of material from virtually all the surrounding island groups in Central Eastern Polynesia (e.g., Austral Islands, Pitcairn Group), as well as a single adze (C7727) linked to the island of Kaho’olawe in the Hawaiian archipelago, but made in the 3A adze style not otherwise found in Hawai’i. Anderson [116] and others have pointed out the inherent difficulty in using these surface finds to make sweeping statements about long-distance travel. In a forum in the Journal of Island and Coastal Archaeology, Anderson frames the problem in terms of what he calls ‘traditionalism’ versus ‘historicity’; the former broadly drawing upon oral traditions referring to distant islands, and the latter narrowly focusing on documentary evidence.
Thanks to new research in the Society Islands [117] and Austral Islands [54,118], we have the first documented cases of movement of volcanic stone artifacts between neighboring islands in Central Eastern Polynesia. On the small island of Mo’orea in the Society Islands, Kahn et al. [99] used a two step methodology where 198 volcanic stone artifacts were screened using non-destructive XRF (X-ray Fluorescence) to divide them into two groups, and then a sub-set analyzed with the WD-XRF (Wavelength Dispersive X-ray Fluorescence) method (n = 47). The results, when compared with other published geochemistry, showed that one geochemical group (Group B) was local (i.e., from the island of Mo’orea), and the other (Group A) included local stone and non-local stone from the two closest islands. These non-local samples (n = 14) constituted 30% of the sub-sample tested, that most likely came from Tahiti (26%, ~19 km away) with many fewer from Ra’itea (4%, ~190 km away). Interestingly, there was no correlation between the relative rank of the households where non-local stone artifacts where found, however, there was dramatic bias toward sites of religious ritual, accounting for 67% of non-local stone. The authors suggest this is “indicative of exchange among both social elites and ritual elites, such as priests and craft specialists (tahua) who were often of the upper classes” [117] (p. 1201). They also note the lack of extra-regional stone from the Marquesas or Sāmoa at these post- AD 1350 sites is consistent with the evolution of a local prestige economy after long-distance travel had declined. Rollet et al. [118] also apply a destructive WD-XRF method on samples representing the Vitaria Adze Quarry on Rarutu in the Austral Islands along with three adzes from the Peabody Museum. Based on their results, and a re-consideration of the identity of previously published samples, we now have artifacts that represent movement between islands (n = 2; from Raratu or Tubuai to Raivavae), artifacts created from Austral volcanic stone found in neighboring archipelagos (n = 1 in Tuamotus; n = 2 in the Cook Islands), and an artifact imported to the Austral Islands from outside of the island group (n = 1, Marquesas to Tubuai). Ditchfield et al.’s [119] work in the Cook Islands demonstrates the promise of examining reduction sequences and to identify periods of greater and less frequent production.
There is at present, no strong artifact based evidence for trans-Pacific contacts. Ramírez-Aliaga [120] reviewed several distinctly Polynesian artifact classes that have at different times been proposed as having been transferred to South America: obsidian tools (mata’a), clubs (patu), and stone adzes (toki). The parallels in the form of artifacts center on comparisons of the Mapuche of coastal Chile and Marginal Eastern Polynesians, especially the archaeology of Rapa Nui and Aotearoa (New Zealand). While there are uncanny similarities, and some linguistic evidence to suggest a Polynesian source, the evidence in South America remains unconvincing. In the case of obsidian artifacts, “none…have a secure provenance or have been found during well-controlled excavations” (p. 99). Clubs lack “secure archaeological context” (p. 100). What the Mapuche call toki kura are, “axes of biplane to biconvex section, with few adzes” (p. 104)—the Polynesian tradition is the opposite, adzes with few axes—and the long-term local history of axe/adze technology in Americas remains undescribed.
3.2. Modern DNA
The strongest non-archaeological evidence of contacts between Polynesians and Native Americans in the pre-European contact era is a genomic study of modern Rapa Nui indicating admixture with Native Americans. This study focused on genetic data derived from 27 modern Rapa Nui. By assaying for genetic variability across the nuclear genome Moreno-Mayar et al. [104] were able to disentangle portions of the genomes showing shared sequences with Polynesia, as expected, as well as the Americas and Europe. Further, by modeling demographic scenarios, the authors were able to estimate that the likely timing of the admixture between Rapa Nui and Native Americans pre-dated Europeans. Admixture levels were not high, resulting in only about 6% of modern Rapa Nui genomes deriving from Native American sources, but they seem to indicate sufficiently extensive contact to result in offspring of Rapa Nui and Native American parentage. More recent admixture with European populations was more extensive, resulting in about 16% of modern Rapa Nui genomes originating among recent European populations. Calibrating the genetic evidence to metrics of pre-contact interaction is of course problematic, which brings us to the topic of the artifact evidence of travel and social interaction.
3.3. Ancient DNA
Much of the scholarly attention has been on a claim of direct evidence for the domesticated chicken (Gallus gallus) in South America in cal AD 1304–1424 [81]. Major critiques, and the replies to those critiques, center on two issues: the reliability of the radiocarbon date and the question of contamination in the molecular identification of the bones at the Chilean site of El Arenal-1 [98,99,102,103,129]. Others have pointed to alternative models for the evidence at hand [93,100,130]. In our view, the original study’s authors have answered the question of marine carbon contribution but have yet to address the question that the genetic sequences reported are the results of DNA contamination.
Recent ancient DNA studies of human skulls in the collections of the National Museum of Brazil (Museu Nacional)/Universidade Federal do Rio de Janeiro (UFRJ), attributed to a Native American group in Eastern-Central Brazil called the Botocudo have been shown to have mtDNA linages (B4a1a1a and B4a1a1) [103] and nuclear DNA typical of Polynesians [131]. The skulls were accessioned in 1890 and direct radiocarbon dates clearly show that these individuals died post-Columbian contact with the New World (AD 1492), and post-Magellan contact (AD 1521) in Oceania (Individual identified as Bot15, OxA-27184, 408 ± 24 BP and AAR-17522, 417 ± 25 BP; and the individual identified as Bot17, AAR-17657, 487 ± 25 BP). Malaspinas et al. [103] point out that there is a high probability that these two people died before the traffic in EuroAmerican ships increased in AD 1760, but do not think it likely these were people who have traveled on early Western voyages. They suggest that these remains are evidence of possible pre-contact expansion of people out of Polynesia. The authors do, however, concede that there is no independent supporting evidence to connect Native Americans living in Eastern-Central Brazil to Polynesia. We think it more likely that these are the skulls of two people who died in Polynesia sometime early in the period of European voyaging, and whose graves were robbed by later visitors, and then mistakenly grouped in collections with the remains of Native Americans.
4. Patterns of Dispersal, Interaction, and Isolation
Evidence from archaeology, modern genetics, and ancient DNA reflects the general dispersal pattern of founding human populations in the remote islands of the Pacific and the long-distance interaction spheres. New research, taken together, support well-established models [132] (Figure 1), and may help refine patterns of dispersal, interaction, and isolation from long-distance contacts. Figure 4 shows the general pattern of human dispersal and known long-distance interaction spheres. One proposed hypothesis suggests that the formation of an Ancestral Polynesia Culture in Western Polynesia may have involved differential patterns of dispersal, specifically a failed early settlement of Sāmoa, followed by significant later migrations, possibly from Micronesia [15]. It has also been suggested that the pause between the settlement of Western and Eastern Polynesia was centuries longer than currently thought, followed by a remarkably rapid pulse of island colonization [19]. Both explain the current gaps in our current body of knowledge, and as such, only time will tell if these gaps will persist and/or new evidence presented in their support, or if these hypotheses will be falsified by the discovery of countervailing evidence. Our review of long-distance travel between islands of the Pacific clearly shows that best documented cases at present are based on sourcing of artifacts, with the exception of the plant genetic and linguistic evidence of the transfer of the sweet potato to Polynesia from the Americas (see also [72,73,74]). The discovery of admixture of Native American DNA within the genome of the people of Easter Island (Rapa Nui) is strong new evidence for sustained contacts between Polynesia and the Americas. All other evidence presented to date, in our view, requires further research to confirm and eliminate alternative explanations.
5. Conclusions
With the growth of the field over the past decade we have seen lively debates on topics that have a long pedigree in Polynesia. The question of the origins of Polynesian peoples has evolved to a stage where we can consider how differences in the early culture histories of Tonga and Sāmoa contributed to the ethnogenesis, and next generation DNA sequencing of full mtDNA and nuclear DNA has exposed variability impossible to see using short-sequences of control regions. Statistical approaches to dating settlement by Bayesian models and pooled radiocarbon probabilities—called here the new chronometric hygiene—have taken center stage as the number of dates appropriate for resolving the question of the arrival of people remains frustratingly small. This greater reliance on statistical modeling has been polarizing in the case of Wilmshurt et al.’s [19] push to revise settlement to periods well after we have good site-based and paleo-environmental proxy evidence for the presence of people. In other cases, it has produced results that independently support more conventional methods of estimating the arrival of people. It is perhaps unnecessary to say that more securely dated archaeological deposits will improve the current picture.
The physical evidence for long-distance interaction has advanced with a new geochemical matching of artifacts to their natural source highlighting interaction spheres stretching across much of Polynesia. We note that not all of the material types moved between islands with the same distance or frequency, with basalt adzes traveling a great deal more than artifacts made of volcanic glass or pottery. We now have more evidence linking Western Polynesia and Central Eastern Polynesia [110], artifacts representing movements between the islands of Central Eastern Polynesia [118], and a single artifact from Marginal Eastern Polynesia found in Central Eastern Polynesia [115]. There is also evidence of travel between the islands of Western Polynesia and a number of islands in Micronesia and Melanesia, including Polynesian Outlier islands [110]. The most widespread metric of travel beyond Western Polynesia is currently basalt sourced to Sāmoa. While small assemblage sizes and poorly controlled archaeological context continue to be problems for quantifying the degree of interaction and isolation of island communities, what is beginning to emerge is a picture of linked regional hubs in Western Polynesia and Central Eastern Polynesia connecting most of the remote islands in this area of the Pacific. We see much less frequent long-distance travel originating, or ending, at the points of the Polynesian triangle, and no artifact-based evidence for travel to or from the Americas.
Modern human genetics—which we should take care not to over-interpret since it is the by-product of both pre- and post-European contact historical processes—suggest that there were sex-specific patterns in the settlement of Polynesia with larger cohorts of male linages traced back to Near Oceania and nearly all female linages back to Asia. At present we can confidently bracket the earliest presence of people to Tonga in 2838 ± 8 BP dated by uranium series [37], supported by a Bayesian model of radiocarbon dates from Lapita sites to 2863–2835 cal BP [38]. On the late end of the settlement of Polynesia, New Zealand is dated through the Bayesian model of radiocarbon dates to AD 1270–1309 [35].
Of the material evidence of contact with the Americas, direct AMS radiocarbon dates on sweet potato remain our most secure archaeological signal of contact [see 90]. The exciting announcement of the discovery of the Polynesian chicken in a pre-contact era site in South America (El-Arenel, Chile) has unfortunately been blunted by the possibility of laboratory contamination and post-European contact era. Polynesian remains reported in Brazilian collections, in our view, do not speak to pre-contact Polynesian-American contact. This is not to say that there were no trans-Pacific long-distance contacts; indeed the discovery of what appears to be pre-European contact admixture of Native American DNA within the genome of Rapa Nui people is strong evidence for a period of sustained contacts.
Acknowledgments
We thank the guest editors of this special issue, Anna Linderholm and Ben Krause-Kyora, for the invitation to write this review. We are grateful to three anonymous reviewers for their comments and help in revising the manuscript. Thank you to all our many colleagues who have helped us navigate the issues discussed here, and especially to Roger Green and Patrick Kirch for their mentorship and for championing a holistic approach to anthropology. KAH thanks the Downey Family Award for Faculty Excellence for financial support of this work.
Author Contributions
K. Ann Horsburgh and Mark D. McCoy conceived and wrote the paper.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Keegan, W.F.; Diamond, J.M. Colonization of Islands by Humans: A biogeographic perspective. Adv. Archaeol. Method Theory 1987, 10, 49–92. [Google Scholar]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Mead, M. Introduction to Polynesia as a laboratory for the development of models in the study of cultural evolution. J. Polyn. Soc. 1957, 66, 145. [Google Scholar]
- Kirch, P.V. Hawaii as a model system for human ecodynamics. Am. Anthropol. 2007, 109, 8–26. [Google Scholar] [CrossRef]
- Vitousek, P.M. Oceanic islands as model systems for ecological studies. J. Biogeogr. 2002, 29, 573–582. [Google Scholar] [CrossRef]
- Slatkin, M.; Racimo, F. Ancient DNA and human history. Proc. Natl. Acad. Sci. USA 2016, 113, 6380–6387. [Google Scholar] [CrossRef] [PubMed]
- Haber, M.; Mezzavilla, M.; Xue, Y.; Tyler-Smith, C. Ancient DNA and the rewriting of human history: Be sparing with Occam’s razor. Genome Biol. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Horsburgh, K.A. Molecular anthropology: The judicial use of genetic data in archaeology. J. Archaeol. Sci. 2015, 56, 141–145. [Google Scholar] [CrossRef]
- Horsburgh, K.A.; Orton, J.; Klein, R.G. Beware the springbok in sheep’s clothing: How secure are the faunal identifications upon which we build our models? Afr. Archaeol. Rev. 2016, 33, 353–361. [Google Scholar] [CrossRef]
- Fitzpatrick, S.M.; Anderson, A. Islands of isolation: Archaeology and the power of aquatic perimeters. J. Isl. Coast. Archaeol. 2008, 3, 4–16. [Google Scholar] [CrossRef]
- D’Urville, J.S.C.D. Sur les iles du Grand Ocean. Bulletin de la Societe de Geographie 1832, 17, 1–21. [Google Scholar]
- Bellwood, P. Man’s Conquest of the Pacific: The Prehistory of Southeast Asia and Oceania; Collins Publishers Ltd.: Auckland, New Zealand, 1978. [Google Scholar]
- Kirch, P.V.; Green, R.C. Hawaiki, Ancestral Polynesia: An Essay in Historical Anthropology; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Boyd, R.; Mulder, M.B.; Durham, W.H.; Richerson, P.J. Are cultural phylogenies possible? In Human by Nature: Between Biology and the Social Sciences; Weingart, P., Mitchell, S.D., Richerson, P.J., Maasen, S., Eds.; Lawrence Erlbaum Associates: London, UK, 1997; pp. 355–386. [Google Scholar]
- Addison, D.J.; Matisoo-Smith, E. Rethinking Polynesians origins: A West-Polynesia Triple-I Model. Archaeol. Ocean. 2010, 45, 1–12. [Google Scholar] [CrossRef]
- Burley, D.V.; Addison, D.J. Tonga and Sāmoa in Oceanic prehistory: Contemporary debates and personal perspectives. In The Oxford Handbook of Prehistoric Oceania; Cochrane, E., Hunt, T., Eds.; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Cochrane, E.E.; Kane, H.; Fletcher, C., III; Horrocks, M.; Mills, J.; Barbee, M.; Morrison, A.E.; Tautunu, M.M. Lack of suitable coastal plains likely influenced Lapita (~2800 cal. BP) settlement of Sāmoa: Evidence from south-eastern ′Upolu. Holocene 2015, 26, 126–135. [Google Scholar] [CrossRef]
- Rieth, T.M.; Cochrane, E.E. The Chronology of Colonization in Remote Oceania. In The Oxford Handbook of Prehistoric Oceania; Cochrane, E., Hunt, T., Eds.; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Wilmshurst, J.M.; Hunt, T.L.; Lipo, C.P.; Anderson, A.J. High-precision radiocarbon dating shows recent and rapid initial human colonization of East Polynesia. Proc. Natl. Acad. Sci. USA 2011, 108, 1815–1820. [Google Scholar] [CrossRef] [PubMed]
- Spriggs, M.; Anderson, A. Late colonization of East Polynesia. Antiquity 1993, 67, 200–217. [Google Scholar] [CrossRef]
- Allen, M.S.; Huebert, J.M. Short-Lived Plant Materials, Long-Lived Trees, and Polynesian 14C Dating: Considerations for 14C Sample Selection and Documentation. Radiocarbon 2014, 56, 257–276. [Google Scholar] [CrossRef]
- Petchey, F.; Allen, M.A.; Addison, D.J.; Anderson, A. Stability in the South Pacific surface marine 14C reservoir over the last 750 years. Evidence from American Samoa, the southern Cook Islands and the Marquesas. J. Archaeol. Sci. 2009, 36, 2234–2243. [Google Scholar] [CrossRef]
- Weisler, M.I.; Hua, Q.; Zhao, J. Late Holocene C-14 marine reservoir ages in Hawaii from U-series dated corals. Radiocarbon 2009, 51, 955–968. [Google Scholar] [CrossRef]
- Rieth, T.M.; Athens, J.S. Suggested best practices for the application of radiocarbon dating to Hawaiian archaeology. Hawaii. Arch. 2013, 13, 3–29. [Google Scholar]
- Smith, I. Protocols for organising radiocarbon dated assemblages from New Zealand archaeological sites for comparative analysis. J. Pac. Archaeol. 2010, 1, 184–187. [Google Scholar]
- Carson, M. Chronology of Kaua’i: Colonisation, land use demography. J. Polyn. Soc. 2006, 115, 173–186. [Google Scholar]
- McCoy, M.D. A revised late Holocene culture history for Moloka’i Island. Hawai’i Radiocarb. 2007, 49, 1273–1322. [Google Scholar] [CrossRef]
- Rieth, T.M.; Hunt, T.L.; Lipo, C.; Wilmshurst, J.M. The 13th century Polynesian colonization of Hawai’i Island. J. Archaeol. Sci. 2011, 38, 2740–2749. [Google Scholar] [CrossRef]
- Rieth, T.M.; Hunt, T.L. A radiocarbon chronology for Sāmoan prehistory. J. Archaeol. Sci. 2008, 35, 1901–1927. [Google Scholar] [CrossRef]
- Mulrooney, M.A. An island-wide assessment of the chronology of settlement and land use on Rapa Nui (Easter Island) based on radiocarbon data. J. Archaeol. Sci. 2013, 40, 4377–4399. [Google Scholar] [CrossRef]
- Athens, J.S.; Rieth, T.M.; Dye, T.S. A paleoenvironmental and archaeological model-based age estimate for the colonization of Hawai’i. Am. Antiq. 2014, 79, 144–155. [Google Scholar] [CrossRef]
- Mulrooney, M.A.; Bickler, S.H.; Allen, M.S.; Ladefoged, T.N. High-precision dating of colonization and settlement in East Polynesia. Proc. Natl. Acad. Sci. USA 2011, 108, E192–E194. [Google Scholar] [CrossRef] [PubMed]
- Wilmshurst, J.M.; Hunt, T.L.; Lipo, C.P.; Anderson, A.J. Reply to Mulrooney et al.: Accepting lower precision radiocarbon dates results in longer colonization chronologies for East Polynesia. Proc. Natl. Acad. Sci. USA 2011, 108, E195. [Google Scholar] [CrossRef]
- Kirch, P.V. When did the Polynesians settle Hawaii? A review of 150 years of scholarly inquiry and a tentative answer. Hawaii. Arch. 2011, 12, 3–26. [Google Scholar]
- Dye, T.S. Dating human dispersal in Remote Oceania: A Bayesian view from Hawai‘i. World Archaeol. 2015, 47, 661–676. [Google Scholar] [CrossRef]
- Burley, D.; Weisler, M.I.; Zhao, J. High precision U/Th dating of first Polynesian settlement. PLoS ONE 2012, 7, e48769. [Google Scholar] [CrossRef] [PubMed]
- Burley, D.V.; Barton, A.; Dickinson, W.R.; Connaughton, S.P.; Taché, K. Nukuleka as a founder colony for West Polynesian settlement: New insights from recent excavations. J. Pac. Archaeol. 2010, 1, 128–144. [Google Scholar]
- Burley, D.; Edinborough, K.; Weisler, M.; Zhao, J. Bayesian modeling and chronological precision for Polynesian settlement of Tonga. PLoS ONE 2015, 10, e0120795. [Google Scholar] [CrossRef] [PubMed]
- Rieth, T.M.; Morrison, A.E.; Addison, D.J. The temporal and spatial patterning of the initial settlement of Sāmoa. J. Isl. Coast. Archaeol. 2008, 3, 214–239. [Google Scholar] [CrossRef]
- Smith, A. An Archaeology of West Polynesian Prehistory; Australian National University Press: Canberra, Australia, 2002. [Google Scholar]
- Davidson, J.M. Intrusion, integration and innovation on small and not-so-small islands with particular reference to Samoa. Archaeol. Ocean. 2012, 47, 1–13. [Google Scholar] [CrossRef]
- Kirch, P.V.; Yen, D.E. Tikopia: The prehistory and ecology of a Polynesian Outlier; Bishop Museum Press: Honolulu, HI, USA, 1982. [Google Scholar]
- Storey, A.A.; Clarke, A.C.; Ladefoged, T.; Robins, J.; Matisoo-Smith, E. DNA and Pacific Commensal Models: Construction, limitations, and future prospects. J. Isl. Coast. Archaeol. 2013, 8, 37–65. [Google Scholar] [CrossRef]
- Greig, K.; Walter, R.; Matisoo-Smith, E.A. Dogs and people in Southeast Asia and the Pacific. In Routledge Handbook of Bioarchaeology in Southeast Asia and the Pacific Islands; Oxenham, M., Buckley, H., Eds.; Routledge: Abingdon, UK, 2016; pp. 462–482. [Google Scholar]
- Larson, G.; Cucchi, T.; Fujita, M.; Matisoo-Smith, E.; Robins, J.; Anderson, A.; Rolett, B.; Spriggs, M.; Dolman, G.; Kim, T.; et al. Phylogeny and ancient DNA of Sus provides insights into neolithic expansion in Island Southeast Asia and Oceania. Proc. Natl. Acad. Sci. USA 2007, 104, 4834–4839. [Google Scholar] [CrossRef] [PubMed]
- Clarke, A. Origins and Dispersal of the Sweet Potato and Bottle Gourd in Oceania: Implications for Prehistoric Human Mobility. Ph.D. Thesis, Massey University, Palmerston North, New Zealand, 2009. [Google Scholar]
- Roullier, C.; Duputié, A.; Wennekes, P.; Benoit, L.; Fernández Bringas, V.M.; Rossel, G.; Tay, D.; McKey, D.; Lebot, V. Disentangling the Origins of Cultivated Sweet Potato (Ipomoea batatas (L.) Lam.). PLoS ONE 2013, 8, e62707. [Google Scholar] [CrossRef]
- Roullier, C.; Kambouo, R.; Paofa, J.; McKey, D.; Lebot, V. On the origin of sweet potato (Ipomoea batatas (L.) Lam.) genetic diversity in New Guinea, a secondary centre of diversity. Heredity 2013, 110, 594–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burley, D.V. Fijian Polygenesis and the Melanesian/Polynesian divide. Curr. Anthropol. 2013, 54, 436–462. [Google Scholar] [CrossRef]
- Nunn, P. Comment on “Fijian Polygenesis and the Melanesian/Polynesian divide” by David V. Burley. Curr. Anthropol. 2013, 54, 453–454. [Google Scholar]
- Petersen, G. Micronesia’s breadfruit revolution and the evolution of a culture area. Archaeol. Ocean. 2006, 41, 82–92. [Google Scholar] [CrossRef]
- Allen, M.S.; McAlister, A. Early Marquesan settlement patterns of interaction: New insights from Hatiheu Valley, Nuku Hiva Island. J. Pac. Archaeol. 2013, 4, 90–109. [Google Scholar]
- Allen, M.S.; Wallace, R. New evidence from the East Polynesian gateway: Substantive and methodological results from Aitutaki, southern Cook Islands. Radiocarbon 2007, 49, 1163–1179. [Google Scholar] [CrossRef]
- Bollt, R. Peva: The Archaeology of an Austral Island Settlement. Bulletins in Anthropology; Bishop Museum Press: Honolulu, HI, USA, 2008. [Google Scholar]
- Conte, E.; Molle, G. Reinvestigating a key site for Polynesian prehistory: New results from the Hane dune site, Ua Huka (Marquesas). Archaeol. Ocean. 2014, 49, 121–136. [Google Scholar] [CrossRef]
- Molle, G. Ua Huka, une île dans l’Histoire. Histoire pré-et post-Européenne d’une Société Marquisienne. Ph.D. Thesis, University of French Polynesia, Tahiti, France, 2 December 2011. [Google Scholar]
- Allen, M.S. Marquesan colonization chronologies and post-colonisation interaction: Implications for Hawaiian origins and the ‘Marquesan Homeland’ hypothesis. J. Pac. Archaeol. 2004, 5, 1–17. [Google Scholar]
- Brooks, E.; Jacomb, C.; Walter, R. Archaeological investigations at Wairau Bar. Archaeol. N. Z. 2009, 52, 259–268. [Google Scholar]
- Brooks, E.; Walter, R.; Jacomb, C. History of excavations at Wairau Bar. Rec. Canterb. Mus. 2011, 25, 13–58. [Google Scholar]
- Davidson, J.; Findlater, A.; Fyfe, R.; MacDonald, J.; Marshall, B. Connections with Hawaiki: The evidence of a shell tool from Wairau Bar, Marlborough, New Zealand. J. Pac. Archaeol. 2011, 2, 93–102. [Google Scholar]
- Jacomb, C.; Holdaway, R.; Allentoft, M.E.; Bunce, M.; Oskam, C.; Walter, R. High-precision dating and ancient DNA profiling of moa (Aves: Dinornithiformes) eggshell documents a complex feature at Wairau Bar and refines the chronology of New Zealand settlement by Polynesians. J. Archaeol. Sci. 2014, 50, 24–30. [Google Scholar] [CrossRef]
- Jacomb, C.; Walter, R.; Brooks, E. Living on Pipi (Paphies. australis): Specialised shellfish harvest in a marginal environment at Karamea, West Coast, New Zealand. J. Pac. Archaeol. 2010, 1, 36–52. [Google Scholar]
- Oskam, C.L.; Jacomb, C.; Allentoft, M.E.; Walter, R.; Scofield, R.P.; Haile, J.; Holdaway, R.N.; Bunce, M. Molecular and morphological analyses of avian eggshell excavated from a late thirteenth century earth oven. J. Archaeol. Sci. 2011, 38, 2589–2595. [Google Scholar] [CrossRef]
- Anderson, A. Subpolar settlement in South Polynesia. Antiquity 2005, 79, 791–800. [Google Scholar] [CrossRef]
- Dye, T.S. A model-based age estimate for Polynesian colonization of Hawai’i. Archaeol. Ocean. 2011, 46, 130–138. [Google Scholar] [CrossRef]
- Hunt, T.L.; Lipo, C.P. Late colonization of Easter Island. Science 2006, 311, 1603–1606. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A. Slow boats from China: Issues in the prehistory of Indo-Pacific seafaring. In East of Wallace’s Line: Studies of Past and Present Maritime Cultures of the Indo-Pacific Region; O’Connor, S., Veth, P., Eds.; A Balkema: Rotterdam, The Netherlands, 2000; pp. 13–51. [Google Scholar]
- Anderson, A.; Chappell, J.; Gagan, M.; Grove, R. Prehistoric maritime migration in the Pacific Islands: An hypothesis of ENSO forcing. Holocene 2006, 16, 1–6. [Google Scholar] [CrossRef]
- Goodwin, I.D.; Browning, S.A.; Anderson, A.J. Climate windows for Polynesian voyaging to New Zealand and Easter Island. Proc. Natl. Acad. Sci. USA 2014, 111, 14716–14721. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A. Polynesian seafaring and American horizons: A response to Jones and Klar. Am. Antiq. 2006, 71, 759–763. [Google Scholar] [CrossRef]
- Di Piazza, A.; Di Piazza, P.; Pearthree, E. Sailing virtual canoes across Oceania: Revisiting island accessibility. J. Archaeol. Sci. 2007, 34, 1219–1225. [Google Scholar] [CrossRef]
- Montenegro, Á.; Callaghan, R.T.; Fitzpatrick, S.M. Using seafaring simulations and shortest-hop trajectories to model the prehistoric colonization of Remote Oceania. Proc. Natl. Acad. Sci. USA 2016, 113, 12685–12690. [Google Scholar] [CrossRef] [PubMed]
- Montenegro, A.; Callaghan, R.T.; Fitzpatrick, S.M. From west to east: Environmental influences on the rate and pathways of Polynesian colonization. Holocene 2014, 24, 242–256. [Google Scholar] [CrossRef]
- Walter, R.; Jacomb, C.; Boron-Muth, S. Colonisation, mobility and exchange in New Zealand prehistory. Antiquity 2010, 84, 497–513. [Google Scholar] [CrossRef]
- McCoy, M.D.; Robles, H.R. The Geographic Range of Interaction Spheres during the Colonization of New Zealand (Aotearoa): New evidence for obsidian circulation in Southern New Zealand. J. Isl. Coast. Archaeol. 2015. [Google Scholar] [CrossRef]
- Lawrence, M.; McCoy, M.D.; Barber, I.; Walter, R. Geochemical Sourcing Obsidian from the Purakaunui Site, South Island, New Zealand. Archaeol. Ocean. 2014, 49, 158–163. [Google Scholar] [CrossRef]
- Kinaston, R.L.; Shaw, B.; Gray, A.R.; Walter, R.K.; Jacomb, C.; Brooks, E.; Halcrow, S.E.; Buckley, H.R. Critical Review of Brown and Thomas “The First New Zealanders? An Alternative Interpretation of the Stable Isotope Data from Wairau Bar”. PLoS ONE 2015, 10, e0137616. [Google Scholar] [CrossRef] [PubMed]
- Kinaston, R.L.; Walter, R.K.; Jacomb, C.; Brooks, E.; Tables, N.; Halcrow, S.E.; Stirling, C.; Reid, M.; Gray, A.R.; Spinks, J.; et al. The First New Zealanders: Patterns of Diet and Mobility Revealed through Isotope Analysis. PLoS ONE 2013, 8, e64580. [Google Scholar] [CrossRef]
- Brown, A.A.; Thomas, T. The First New Zealanders? An Alternative Interpretation of Stable Isotope Data from Wairau Bar, New Zeal. PLoS ONE 2015. [Google Scholar] [CrossRef] [PubMed]
- Moodley, Y.; Linz, B.; Yamaoka, Y.; Windsor, H.M.; Breurec, S.; Wu, Y.; Maady, A.; Bernhöft, S.; Thiberge, J.; Phuanukoonnon, S.; et al. The Peopling of the Pacific from a Bacterial Perspective. Science 2009, 323, 527–530. [Google Scholar] [CrossRef] [PubMed]
- Hagelberg, E.; Clegg, J.B. Genetic polymorphism in prehistoric Pacific islanders determined by analysis of ancient bone DNA. Proc. R. Soc. Lond. B Biol. Sci. 1993, 252, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Friedlaender, J.; Schurr, T.; Gentz, F.; Koki, G.; Friedlaender, F.; Horvat, G.; Babb, P.; Cerchio, S.; Kaestle, F.; Schanfield, M.; et al. Expanding Southwest Pacific Mitochondrial Haplogroups P and Q. Mol. Biol. Evol. 2005, 22, 1506–1517. [Google Scholar] [CrossRef] [PubMed]
- Melton, T.; Clifford, S.; Martinson, E.; Batzer, M.; Stoneking, M. Genetic evidence for the proto-Austronesian homeland in Asia: MtDNA and nuclear DNA variation in Taiwanese aboriginal tribes. Am. J. Hum. Genet. 1998, 63, 1807–1823. [Google Scholar] [CrossRef] [PubMed]
- Redd, A.J.; Takezaki, N.; Sherry, S.T.; McGarvey, S.T.; Sofro, A.S.M.; Stoneking, M. Evolutionary history of the COII/tRHA Lys intergenic 9 base pair deletion in human mitochondrial DNAs from the Pacific. Mol. Biol. Evol. 1995, 12, 604–615. [Google Scholar] [PubMed]
- Kayser, M.; Brauer, S.; Cordaux, R.; Casto, A.; Lao, O.; Zhivotovsky, L.A.; Moyse-Faurie, C.; Rutledge, R.B.; Schiefenhoevel, W.; Gil, D.; et al. Melanesian and Asian origins of Polynesians: MtDNA and Y chromosome gradients across the Pacific. Mol. Biol. Evol. 2006, 23, 2234–2244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirabal, S.; Herrera, K.J.; Gaydena, T.; Regueiro, M.; Underhill, P.A.; Garcia-Bertrand, R.L.; Herrera, R.J. Increased Y-chromosome resolution of haplogroup O suggests genetic ties between the Ami aborigines of Taiwan and the Polynesian Islands of Samoa and Tonga. Gene 2010, 494, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Kayser, M.; van Oven, M. Updated comprehensive phylogenetic tree of global human mitochondrial DNA variation. Hum. Mutat. 2009, 30, E386–E394. [Google Scholar]
- Knapp, M.; Horsburgh, K.A.; Prost, S.; Stanton, J.A.; Buckley, H.R.; Walter, R.K.; Matisoo-Smith, E.A. Complete mitochondrial DNA genome sequences from the first New Zealanders. Proc. Natl. Acad. Sci. USA 2012, 109, 18350–18354. [Google Scholar] [CrossRef] [PubMed]
- Benton, M.; Macartney-Coxson, D.; Eccles, D.; Griffiths, L.; Chambers, G.; Lea, R. Complete Mitochondrial Genome Sequencing Reveals Novel Haplotypes in a Polynesian Population. PLoS ONE 2012, 7, e35026. [Google Scholar] [CrossRef]
- Hather, J.G.; Kirch, P.V. Prehistoric sweet potato (Ipomoea batatas) from Mangaia Island, central Polynesia. Antiquity 1991, 65, 887–893. [Google Scholar] [CrossRef]
- Ladefoged, T.N.; Graves, M.W.; Coil, J.H. The introduction of sweet potato in Polynesia: Early remains in Hawai’i. J. Polyn. Soc. 2005, 114, 359–374. [Google Scholar]
- Montenegro, A.; Avis, C.; Weaver, A.J. Modeling the prehistoric arrival of the sweet potato in Polynesia. J. Archaeol. Sci. 2008, 35, 355–367. [Google Scholar] [CrossRef]
- Jones, T.L.; Klar, K.A. Diffusionism reconsidered: Linguistic and archaeological evidence for prehistoric Polynesian contact with southern California. Am. Antiq. 2005, 70, 457–484. [Google Scholar] [CrossRef]
- Arnold, J.E. Credit Where Credit is Due: The history of the Chumash oceangoing plank canoe. Am. Antiq. 2007, 72, 196–209. [Google Scholar] [CrossRef]
- Jones, T.L.; Storey, A.A.; Matisoo-Smith, E.A.; Ramírez-Aliaga, J.M. (Eds.) Polynesians in America: Pre-Columbian Contacts with the New World; AltaMira Press: New York, NY, USA, 2011. [Google Scholar]
- Storey, A.A.; Ramírez, J.M.; Quiroz, D.; Burley, D.V.; Addison, D.J.; Walter, R.; Anderson, A.J.; Hunt, T.L.; Athens, J.S.; Huynen, L.; et al. Radiocarbon and DNA evidence for a pre-Columbian introduction of Polynesian chickens to Chile. Proc. Natl. Acad. Sci. USA 2007, 104, 10335–10339. [Google Scholar] [CrossRef] [PubMed]
- Storey, A.A.; Ladefoged, T.; Matisoo-Smith, E.A. Counting your chickens: Density and distribution of chicken remains in archaeological sites of Oceania. Int. J. Osteoarchaeol. 2008, 18, 240–261. [Google Scholar] [CrossRef]
- Gongora, J.; Rawlence, N.J.; Mobegi, V.A.; Jianlin, H.; Alcalde, J.A.; Matus, J.T.; Hanotte, O.; Moran, C.; Austin, J.J.; Ulm, S.; et al. Indo-European and Asian origins for Chilean and Pacific chickens revealed by mtDNA. Proc. Natl. Acad. Sci. USA 2008, 105, 10308–10313. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, S.M.; Callaghan, R. Examining dispersal mechanisms for the translocation of chicken (Gallus gallus) from Polynesia to South America. J. Archaeol. Sci. 2009, 36, 214–223. [Google Scholar] [CrossRef]
- Storey, A.A.; Athens, J.S.; Bryant, D.; Carson, M.; Emery, K.; deFrance, S.; Higham, C.; Huynen, L.; Intoh, M.; Jones, S.; et al. Investigating the global dispersal of chickens in prehistory using ancient mitochondrial DNA signatures. PLoS ONE 2012, 7, e39171. [Google Scholar] [CrossRef] [PubMed]
- Thomson, V.A.; Lebrasseur, O.; Austin, J.J.; Hunt, T.L.; Burney, D.A.; Denham, T.; Rawlence, N.J.; Wood, J.R.; Gongora, J.; Flink, L.G.; et al. Reply to Beavan, Bryant, and Storey and Matisoo-Smith: Ancestral Polynesian “D” haplotypes reflect authentic Pacific chicken lineages. Proc. Natl. Acad. Sci. USA 2014, 111, E3585–E3586. [Google Scholar] [CrossRef] [PubMed]
- Beavan, N. No evidence for sample contamination or diet offset for pre-Columbian chicken dates from El Arena 1. Proc. Natl. Acad. Sci. USA 2014, 111, E3582. [Google Scholar] [CrossRef] [PubMed]
- Malaspinas, A.; Oscar, L.; Schroeder, H.; Rasmussen, R.; Raghavan, M.; Moltke, I.; Campos, P.F.; Sagredo, F.S.; Rasmussen, S.; Gonçalves, V.F.; et al. Two ancient human genomes reveal Polynesian ancestry among the indigenous Botocudos of Brazil. Curr. Biol. 2014, 24, R1035–R1037. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Mayar, J.V.; Rasmussen, S.; Seguin-Orlando, A.; Rasmussen, M.; Liang, M.; Flåm, S.T.; Lie, B.A.; Gilfillan, G.D.; Nielsen, R.; Thorsby, E.; et al. Genome-wide Ancestry Patterns in Rapanui Suggest Pre-European Admixture with Native Americans. Curr. Biol. 2014, 24, 2518–2525. [Google Scholar] [CrossRef] [PubMed]
- Mills, P.R.; Lundblad, S.P. Current perspectives on Hawai‘i’s stone tool economies. J. Pac. Archaeol. 2014, 5, 30–39. [Google Scholar]
- Burley, D.V.; Dickinson, W.R. Among Polynesia’s first pots. J. Archaeol. Sci. 2010, 37, 1020–1026. [Google Scholar] [CrossRef]
- Allen, M.S.; Johnson, K.T.M. Tracking ancient patterns of interaction: Recent geochemical studies in the southern Cook Islands. In Prehistoric Long-Distance Interaction in Oceania: An Interdisciplinary Approach; Weisler, M.I., Ed.; NZAA: Auckland, New Zealand, 1997; pp. 111–113. [Google Scholar]
- Sheppard, P.; Walter, R.; Parker, R. Basalt sourcing and the development of Cook Island exchange systems. In Prehistoric Long-distance Interaction in Oceania: An Interdisciplinary Approach; Weisler, M.I., Ed.; NZAA: Auckland, New Zealand, 1997; pp. 85–110. [Google Scholar]
- Weisler, M.I. Hard Evidence for Prehistoric Interaction in Polynesia. Curr. Anthropol. 1998, 39, 521–532. [Google Scholar] [CrossRef]
- Clark, G.R.; Reepmeyer, C.; Melekiola, N.; Woodhead, J.; Dickinson, W.R.; Martinsson-Wallin, H. Stone tools from the ancient Tongan state reveal prehistoric interaction centers in the Central Pacific. Proc. Natl. Acad. Sci. USA 2014, 111, 10491–10496. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.S.; Dickinson, W.R.; Huebert, J.M. The anomaly of Marquesan ceramics: A fifty year retrospective. J. Pac. Archaeol. 2012, 3, 90–104. [Google Scholar]
- Cochrane, E.E.; Rieth, T.M. Sāmoan artefact provenance reveals limited artefact transfer within and beyond the archipelago. Archaeol. Ocean. 2016. [Google Scholar] [CrossRef]
- Burley, D.V.; Sheppard, P.J.; Simonin, M. Tongan and Samoan volcanic glass: PXRF analysis and implications for constructs of ancestral Polynesian society. J. Archaeol. Sci. 2011, 38, 2625–2632. [Google Scholar] [CrossRef]
- Eckert, S.L.; James, W.D. Investigating the production and distribution of plain ware pottery in the Samoan archipelago with laser ablation-inductively coupled plasma-mass spectrometry (LA-ICP-MS). J. Archaeol. Sci. 2011, 38, 2155–2170. [Google Scholar] [CrossRef]
- Collerson, K.D.; Weisler, M.I. Stone adze compositions and the extent of ancient Polynesian voyaging and trade. Science 2007, 307, 1907–1911. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A. Traditionalism, Interaction, and Long-Distance Seafaring in Polynesia. J. Isl. Coast. Archaeol. 2008, 3, 240–250. [Google Scholar] [CrossRef]
- Kahn, J.G.; Sinton, J.; Mills, P.R.; Lundblad, S.P. X-ray fluorescence analysis and intra-island exchange in the Society Island archipelago (Central Eastern Polynesia). J. Archaeol. Sci. 2013, 40, 1194–1202. [Google Scholar] [CrossRef]
- Rolett, B.V.; West, E.W.; Sinton, J.M.; Iovita, R. Ancient East Polynesian voyaging spheres: New evidence from the Vitaria adze quarry (Rurutu, Austral Islands). J. Archaeol. Sci. 2015, 53, 459–471. [Google Scholar] [CrossRef]
- Ditchfield, K.; Holdaway, S.; Allen, M.S.; McAlister, A. Measuring stone artifact transport: The experimental demonstration and pilot application of a new method to a prehistoric adze workshop, southern Cook Islands. J. Archaeol. Sci. 2014, 50, 512–523. [Google Scholar] [CrossRef]
- Ramírez-Aliaga, J.M. The Mapuche Connection. In Polynesians in America: Pre-Columbian contacts with the New World; Jones, T.L., Storey, A.A., Matisoo-Smith, E.A., Ramírez-Aliaga, J.M., Eds.; Alta Mira Press: Lanham, MD, USA, 2011; pp. 95–110. [Google Scholar]
- Roullier, C.; Benoit, L.; McKey, B.D.; Lebot, V. Historical collections reveal patterns of diffusion of sweet potato in Oceania obscured by modern plant movements and recombination. Proc. Natl. Acad. Sci. USA 2013, 110, 2205–2210. [Google Scholar] [CrossRef] [PubMed]
- Clarke, A.C.; Burtenshaw, M.K.; McLenachan, P.A.; Erickson, D.L.; Penny, D. Reconstructing the Origins and Dispersal of the Polynesian Bottle Gourd (Lagenaria siceraria). Mol. Biol. Evol. 2006, 23, 893–900. [Google Scholar] [CrossRef] [PubMed]
- Erickson, D.L.; Smith, B.D.; Clarke, A.C.; Sandweiss, D.H.; Tuross, N. An Asian origin for a 10,000-year-old domesticated plant in the Americas. Proc. Natl. Acad. Sci. USA 2005, 102, 18315–18320. [Google Scholar] [CrossRef] [PubMed]
- Kistler, L.; Montenegro, A.; Smith, B.D.; Gifford, J.A.; Greene, R.E.; Newsom, L.A.; Shapiro, B. Transoceanic drift and the domestication of African bottle gourds in the Americas. Proc. Natl. Acad. Sci. USA 2014, 111, 2937–2941. [Google Scholar] [CrossRef] [PubMed]
- Moncada, X.; Payacan, C.; Arriaza, F.; Lobos, S.; Seelenfreund, D.; Seelenfreund, A. DNA extraction and amplification from contemporary Polynesian bark-cloth. PLoS ONE 2013, 8, e56549. [Google Scholar] [CrossRef] [PubMed]
- Seelenfreund, D.; Clarke, A.C.; Oyanedel, N.; Piña, R.; Lobos, S.; Matisoo-Smith, E.A.; Seelenfreund, A. Paper mulberry (Broussonetia papyrifera) as a commensal model for human mobility in Oceania: Anthropological, botanical and genetic considerations. N. Z. J. Bot. 2010, 48, 231–247. [Google Scholar] [CrossRef]
- Zerega, N.; Wiesner-Hanks, T.; Ragone, D.; Irish, B.; Scheffler, B.; Simpson, S.; Zee, F. Diversity in the breadfruit complex (Artocarpus, Moraceae): Genetic characterization of critical germplasm. Tree Genet. Genomes 2015, 11, 4. [Google Scholar] [CrossRef]
- Greig, K.; Boocock, J.; Prost, S.; Horsburgh, K.A.; Jacomb, C.; Walter, R.; Matisoo-Smith, E. Complete mitochondrial genomes of New Zealand’s first dogs. PLoS ONE 2015, 10, e0138536. [Google Scholar] [CrossRef] [PubMed]
- Storey, A.A.; Matisoo-Smith, E. No evidence against Polynesian dispersal of chickens to pre-Columbian South America. Proc. Natl. Acad. Sci. USA 2014, 111, E3583. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A. Islands of exile: Ideological motivation in maritime migration. J. Isl. Coast. Archaeol. 2006, 1, 33–47. [Google Scholar] [CrossRef]
- Goncalves, V.F.; Stenderup, J.; Rodrigues-Carvalho, C.; Silva, H.P.; Goncalves-Dornelas, H.; Liryo, A.; Kivisild, T.; Malaspinas, A.-S.; Campus, P.F.; Rasmussen, M.; et al. Identification of Polynesian mtDNA haplogroups in remains of Botocudo Amerindians from Brazil. Proc. Natl. Acad. Sci. USA 2013, 110, 6465–6469. [Google Scholar] [CrossRef] [PubMed]
- Kirch, P.V. On the Road of the Winds: An Archaeological History of the Pacific Islands before Europe; University of California Press: Oakland, CA, USA, 2002. [Google Scholar]
- Kirch, P.V. The Evolution of Polynesian Chiefdoms; Cambridge University Press: Cambridge, UK, 1984. [Google Scholar]
Figure 1.
The Human Colonization of the Islands of Polynesia. This map shows the general pattern of pulse-and-pause settlement of the remote islands of Polynesia: (1) movement of Lapita peoples with ancestry in Southeast Asia and New Guinea to the island groups that would become Western Polynesia (WP); (2) the dispersal from WP to Central Eastern Polynesia (CEP); and, (3) the settlement of the Polynesian Outliers (PO) in Melanesia and Micronesia from WP, and the settlement of islands of Marginal East Polynesia (MEP) from CEP. Islands by regions and sub-regions mentioned in the text (list is not exhaustive for sub-regions): Melanesia (Bougainville, Solomon Islands, New Britain, Fiji), Western Polynesia (Tonga, Sāmoa, Niue, Tuvalu, Wallis and Futuna), Polynesian Outliers (Tikopia), Central Eastern Polynesia (Society Islands, Cook Islands, Marquesas, Austral Islands, the Pitcairn Group), Marginal Eastern Polynesia (in the north, the Hawaiian Islands; in the east, Rapa Nui (Easter Island); in the southwest, Aotearoa (New Zealand), Chatham Islands, Norfolk Islands, Kermadec Islands, Auckland Islands). Source (base map): OpenStreetMap.
Figure 1.
The Human Colonization of the Islands of Polynesia. This map shows the general pattern of pulse-and-pause settlement of the remote islands of Polynesia: (1) movement of Lapita peoples with ancestry in Southeast Asia and New Guinea to the island groups that would become Western Polynesia (WP); (2) the dispersal from WP to Central Eastern Polynesia (CEP); and, (3) the settlement of the Polynesian Outliers (PO) in Melanesia and Micronesia from WP, and the settlement of islands of Marginal East Polynesia (MEP) from CEP. Islands by regions and sub-regions mentioned in the text (list is not exhaustive for sub-regions): Melanesia (Bougainville, Solomon Islands, New Britain, Fiji), Western Polynesia (Tonga, Sāmoa, Niue, Tuvalu, Wallis and Futuna), Polynesian Outliers (Tikopia), Central Eastern Polynesia (Society Islands, Cook Islands, Marquesas, Austral Islands, the Pitcairn Group), Marginal Eastern Polynesia (in the north, the Hawaiian Islands; in the east, Rapa Nui (Easter Island); in the southwest, Aotearoa (New Zealand), Chatham Islands, Norfolk Islands, Kermadec Islands, Auckland Islands). Source (base map): OpenStreetMap.

Figure 2.
Modern human mtDNA (full-genome) haplogroup frequencies across the Pacific. Note the increased frequency of haplogroup-B (black) from west (left) to east (right).
Figure 2.
Modern human mtDNA (full-genome) haplogroup frequencies across the Pacific. Note the increased frequency of haplogroup-B (black) from west (left) to east (right).
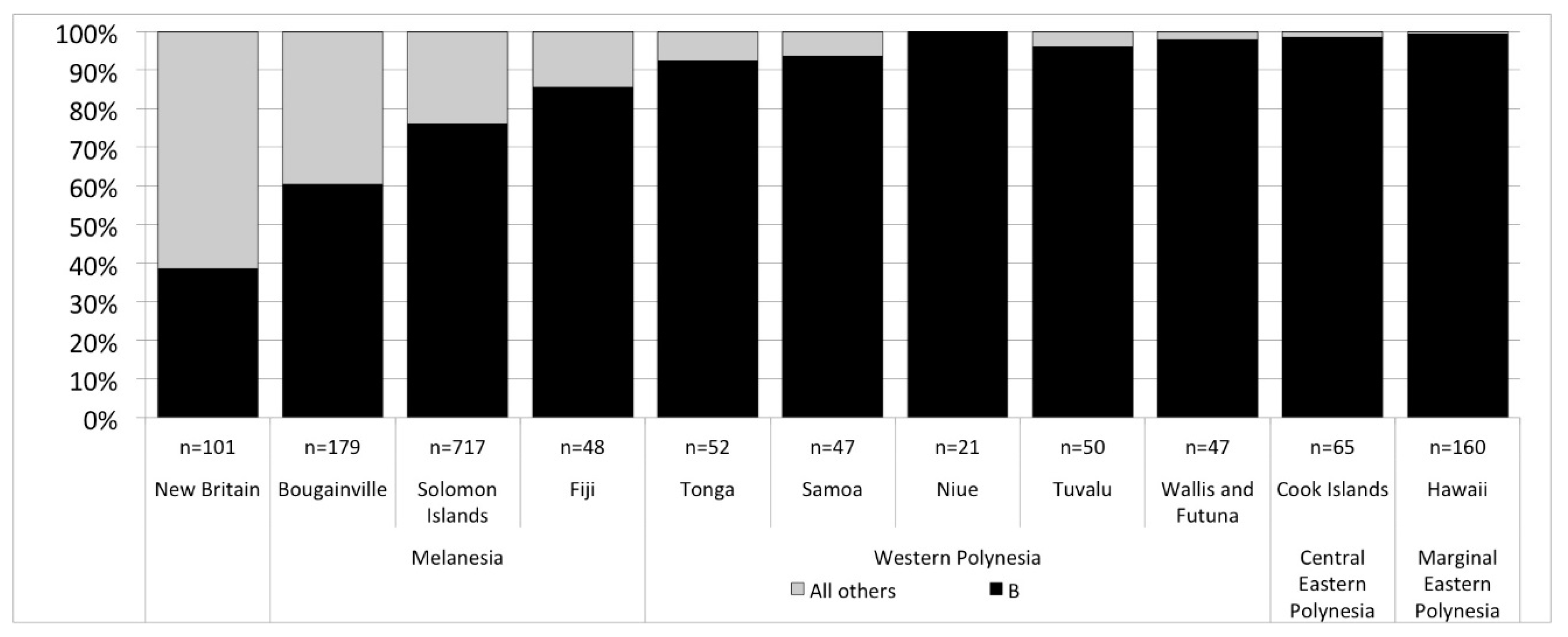
Figure 3.
Linages and Sublinages of haplogroup-B in modern human mtDNA (full-genome). The increased frequencies of some lineages (B4a1a1c; and B4a1a and B4a1a1a and sublineages) we attribute to a serial bottleneck with colonization of Eastern Polynesia. mtDNA reported in Benton et al. [87] and Knapp et al. [88] not shown here.
Figure 3.
Linages and Sublinages of haplogroup-B in modern human mtDNA (full-genome). The increased frequencies of some lineages (B4a1a1c; and B4a1a and B4a1a1a and sublineages) we attribute to a serial bottleneck with colonization of Eastern Polynesia. mtDNA reported in Benton et al. [87] and Knapp et al. [88] not shown here.
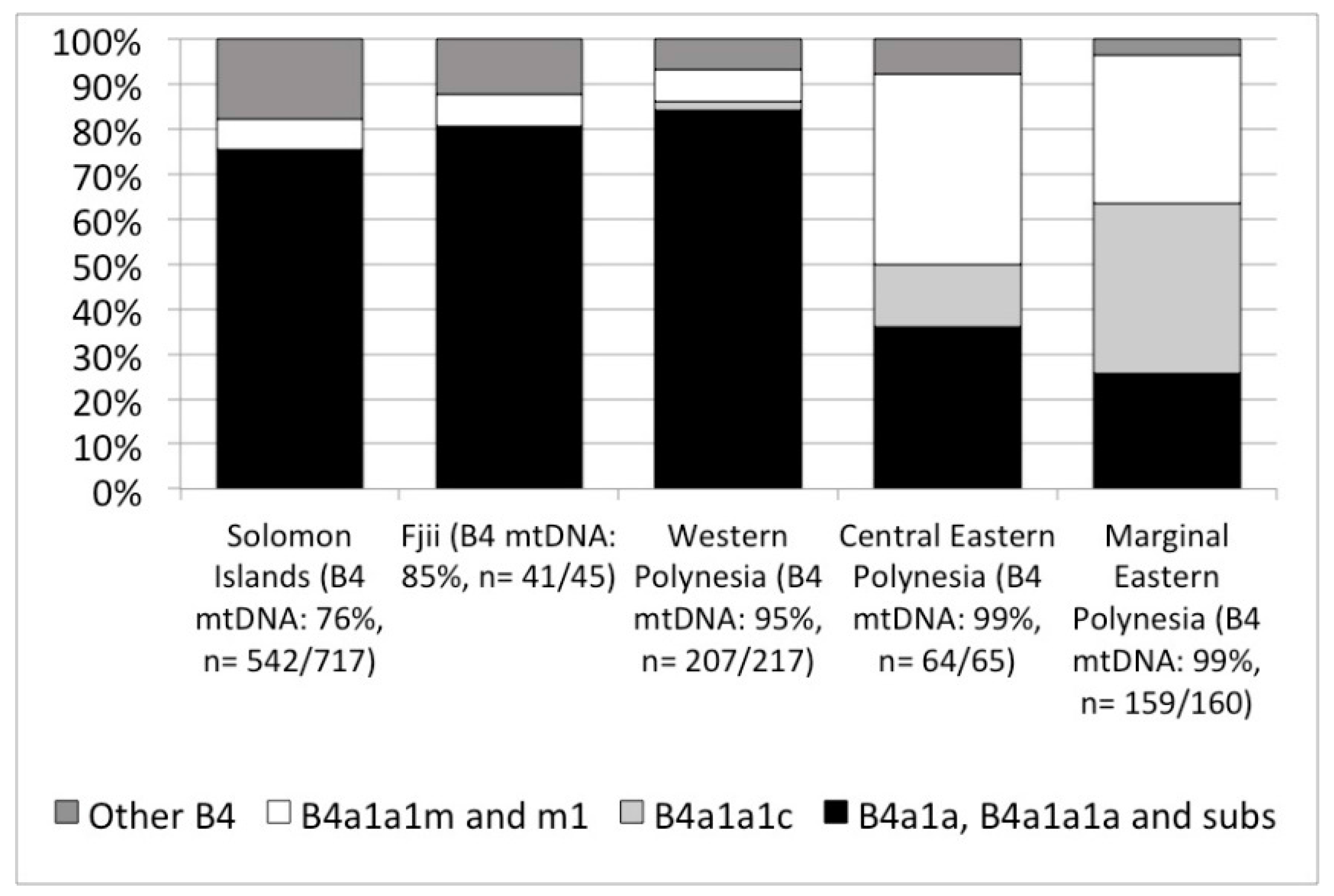
Figure 4.
A model of cultural differentiation in Polynesia. In this general model, modified from Kirch [133], the sequence of colonization is shown beginning with movements of people with ancestry from Asia and Melanesia (Lapita Culture) to islands across the Pacific. An Ancestral Polynesian Culture formed in the island groups that would become Western Polynesia. A dispersal of people out from this region to Central Eastern Polynesia, Marginal Eastern Polynesia, and the Polynesian Outliers created an interaction sphere that spanned a vast region of the Pacific, and likely included the Pacific coast of the Americas. At the time of European contact, the farthest points in this network (Marginal Eastern Polynesia and the Americas) have dropped out. Later long-distance interaction is often attributed to the action of what has been called the Tongan ‘maritime polity’.
Figure 4.
A model of cultural differentiation in Polynesia. In this general model, modified from Kirch [133], the sequence of colonization is shown beginning with movements of people with ancestry from Asia and Melanesia (Lapita Culture) to islands across the Pacific. An Ancestral Polynesian Culture formed in the island groups that would become Western Polynesia. A dispersal of people out from this region to Central Eastern Polynesia, Marginal Eastern Polynesia, and the Polynesian Outliers created an interaction sphere that spanned a vast region of the Pacific, and likely included the Pacific coast of the Americas. At the time of European contact, the farthest points in this network (Marginal Eastern Polynesia and the Americas) have dropped out. Later long-distance interaction is often attributed to the action of what has been called the Tongan ‘maritime polity’.
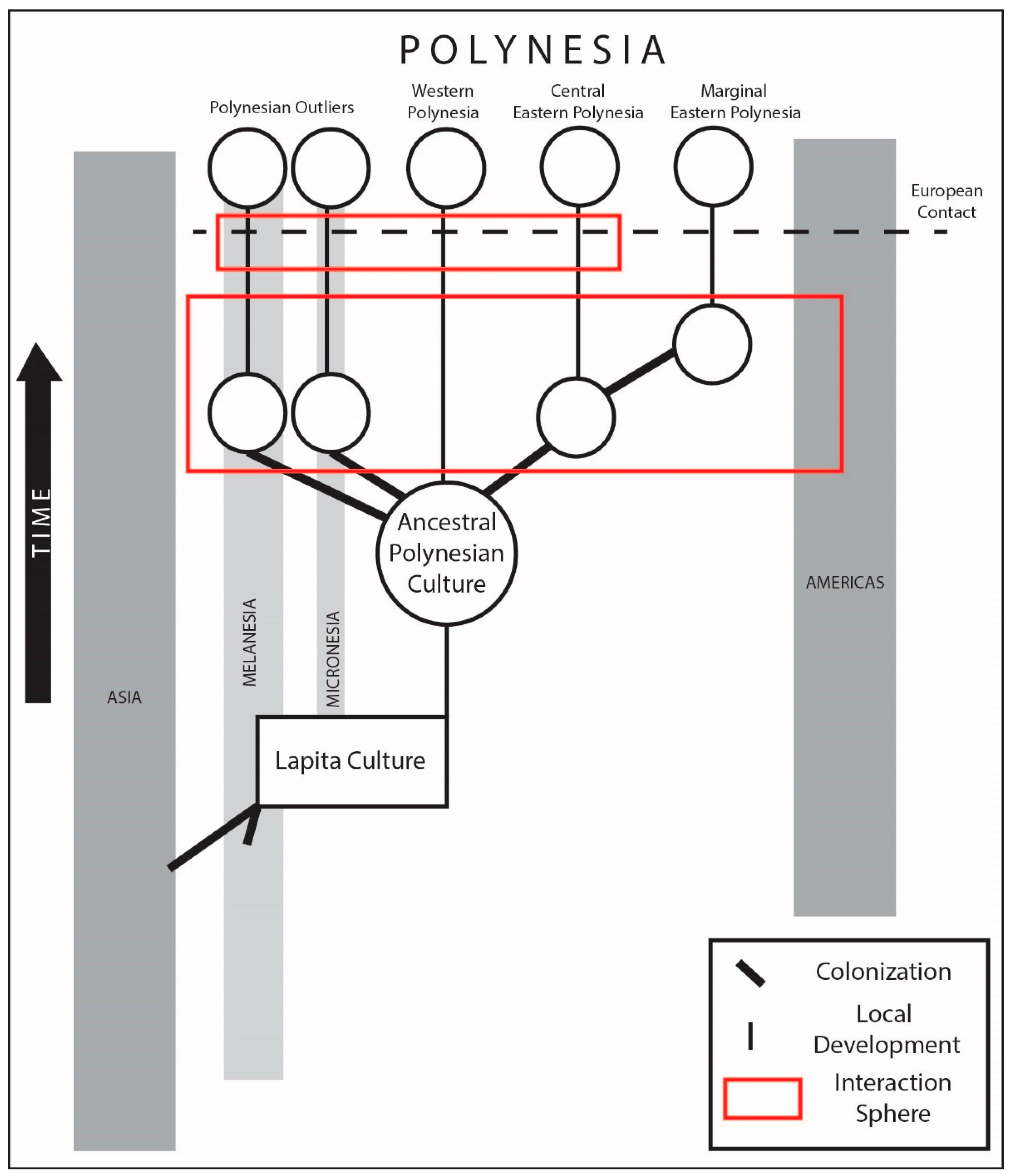

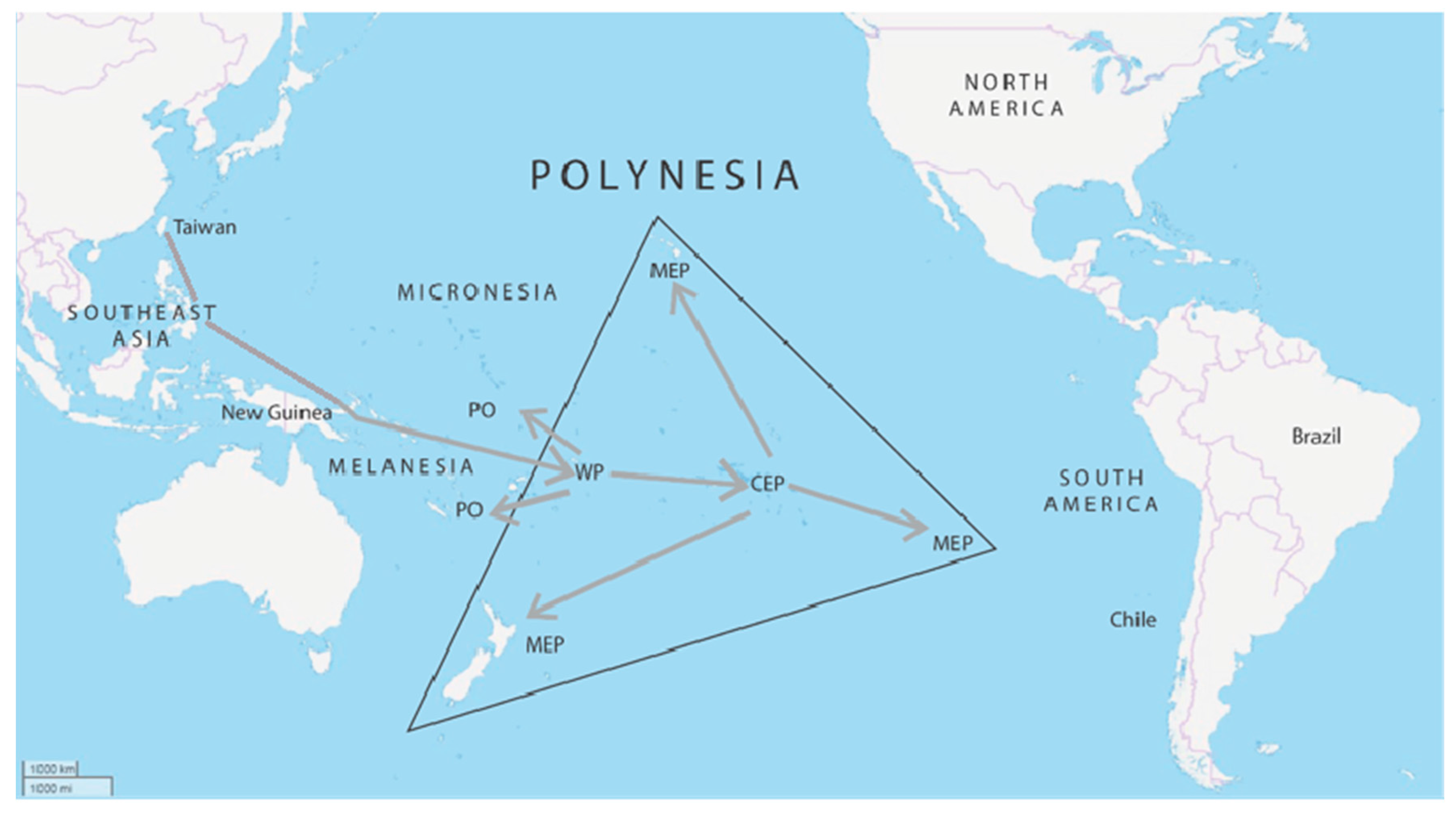{kind=link}
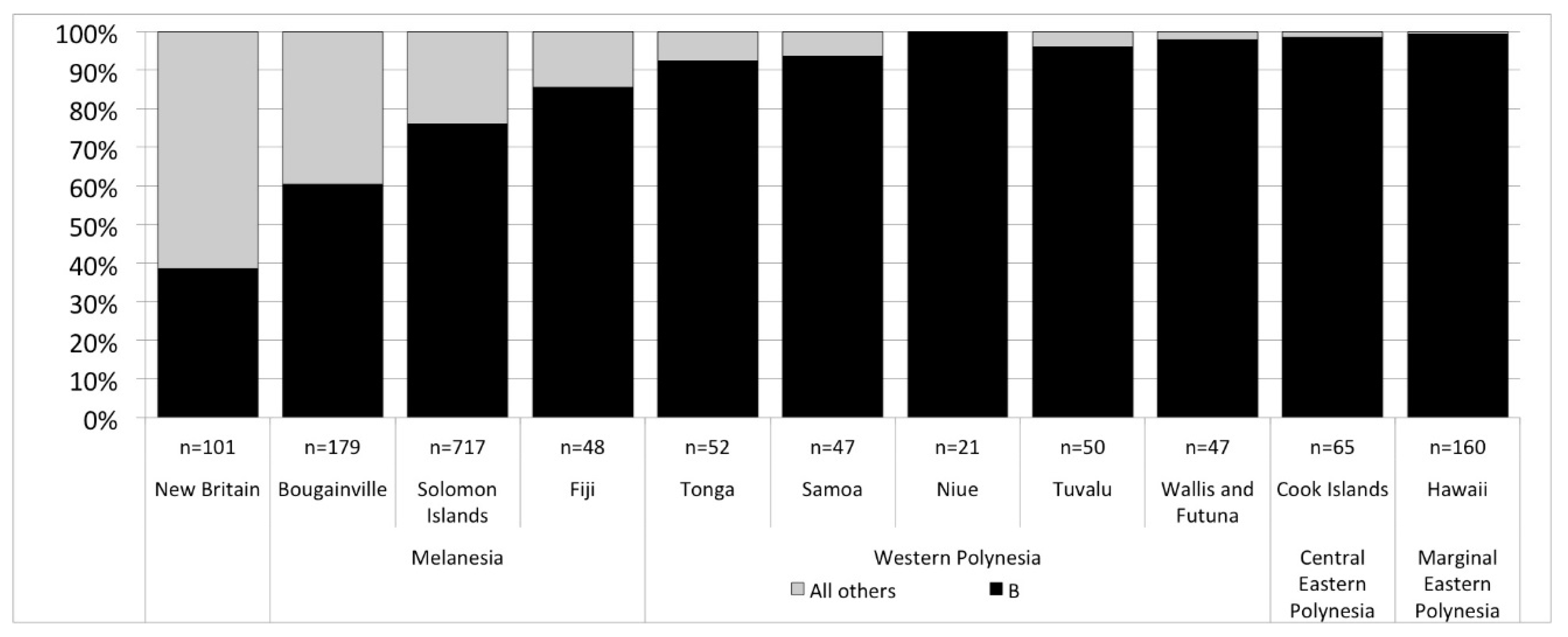{kind=link}
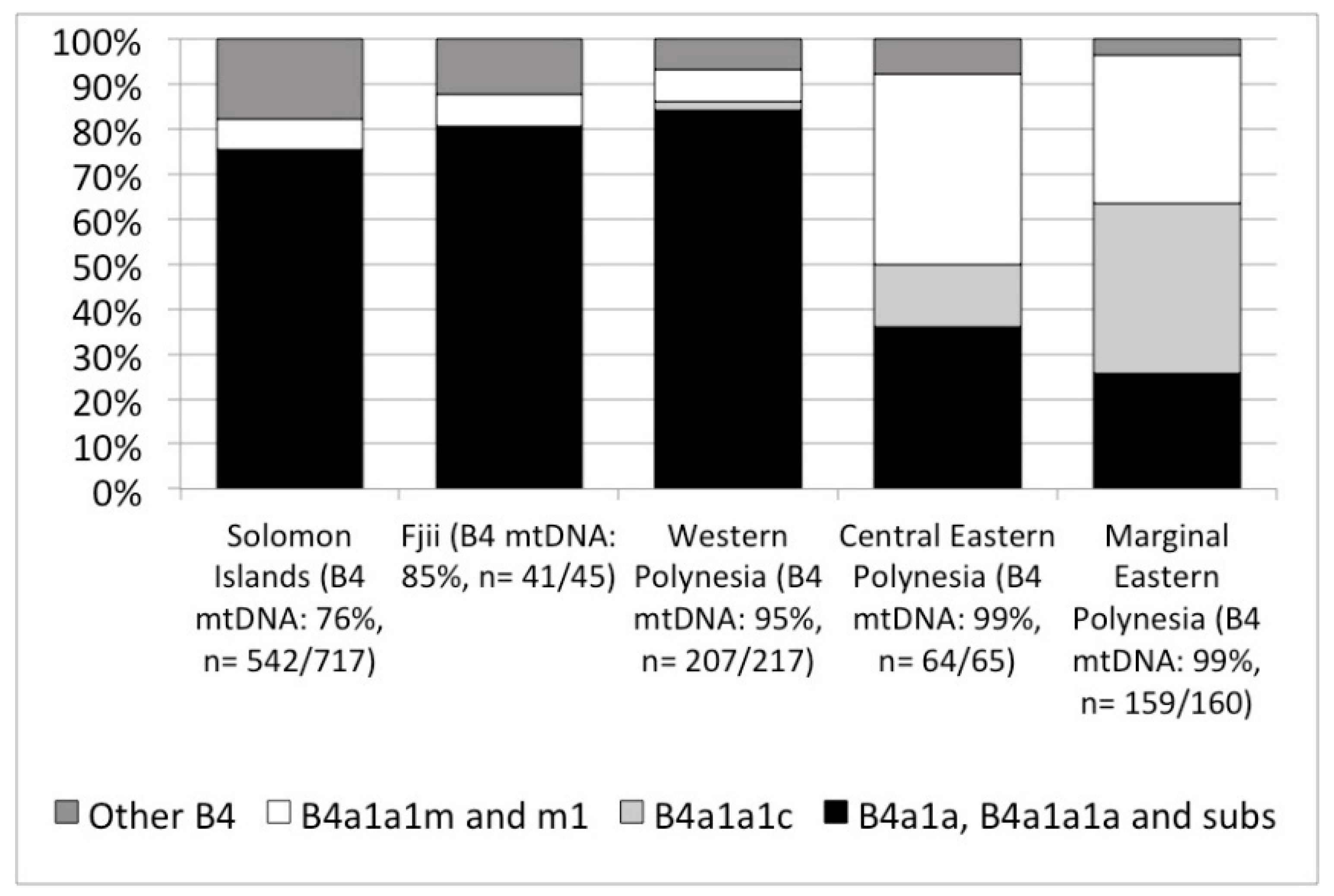{kind=link}
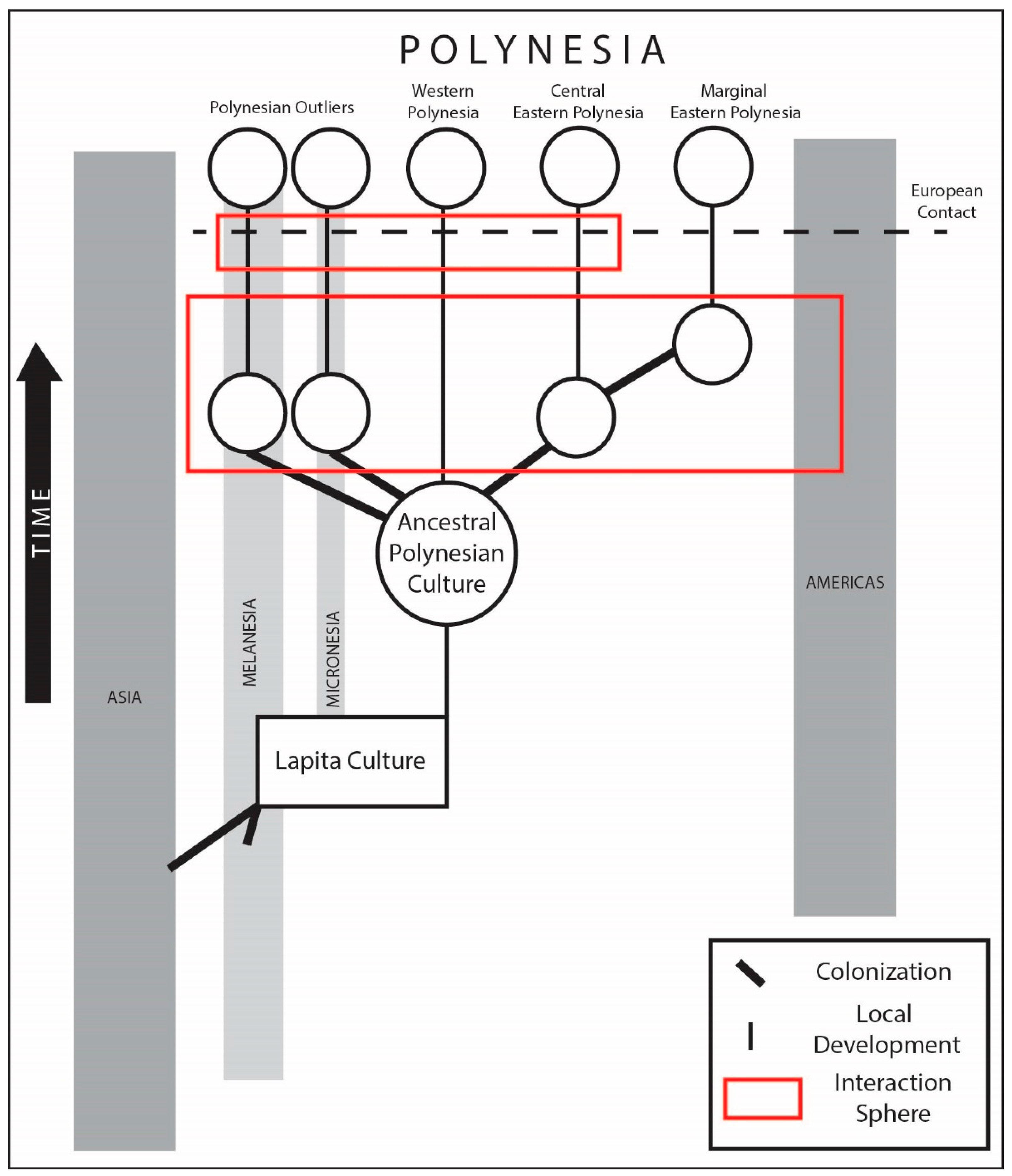{kind=link}
Table 1.
Summary of Genetic Evidence from Commensal Animals (Modern/Ancient). A total of 596 individuals—408 modern and 188 ancient—have been published; in most cases portions mtDNA. After [43].
Table 1.
Summary of Genetic Evidence from Commensal Animals (Modern/Ancient). A total of 596 individuals—408 modern and 188 ancient—have been published; in most cases portions mtDNA. After [43].
| Taxa | Melanesia | Micronesia | Western Polynesia | Polynesian Outliers | Eastern Polynesia |
|---|---|---|---|---|---|
| Rat (Rattus exulans) Modern: 266 Ancient: 115 Total: 381 | 86/16 | 3/3 | 2/3 | 0/5 | 175/88 |
| Pig (Sus scrofa) Modern: 88 Ancient: 6 Total: 94 | 47/0 | 5/5 | 0/1 | 0/1 | 36/4 |
| Chicken (Gallus gallus) Modern: 48 Ancient: 31 Total: 79 | 43/3 | 5/4 | 0/4 | 0/2 | 0/18 |
| Dog (Canis lupus fimiliaris) Modern: 6 Ancient: 36 Total: 40 | 6/1 | 0/0 | 0/0 | 0/0 | 0/33 |
| Modern: 408 Ancient: 188 | 182/20 | 13/12 | 2/8 | 0/8 | 211/143 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Horsburgh, K.A.; McCoy, M.D. Dispersal, Isolation, and Interaction in the Islands of Polynesia: A Critical Review of Archaeological and Genetic Evidence. Diversity 2017, 9, 37. https://doi.org/10.3390/d9030037
AMA Style
Horsburgh KA, McCoy MD. Dispersal, Isolation, and Interaction in the Islands of Polynesia: A Critical Review of Archaeological and Genetic Evidence. Diversity. 2017; 9(3):37. https://doi.org/10.3390/d9030037
Chicago/Turabian StyleHorsburgh, K. Ann, and Mark D. McCoy. 2017. "Dispersal, Isolation, and Interaction in the Islands of Polynesia: A Critical Review of Archaeological and Genetic Evidence" Diversity 9, no. 3: 37. https://doi.org/10.3390/d9030037
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.