Biomarkers of Contaminant Exposure in Chub (Leuciscus cephalus L.) – Biomonitoring of Major Rivers in the Czech Republic

Abstract
:1. Introduction
2. Materials and Methods
2.1 Determination of CYP 450 and EROD activity
2.2 Determination of GST activity and tripeptide GSH
2.3 Determination of PCDD/PCDF and PCB
2.4 Statistical methods
3. Results
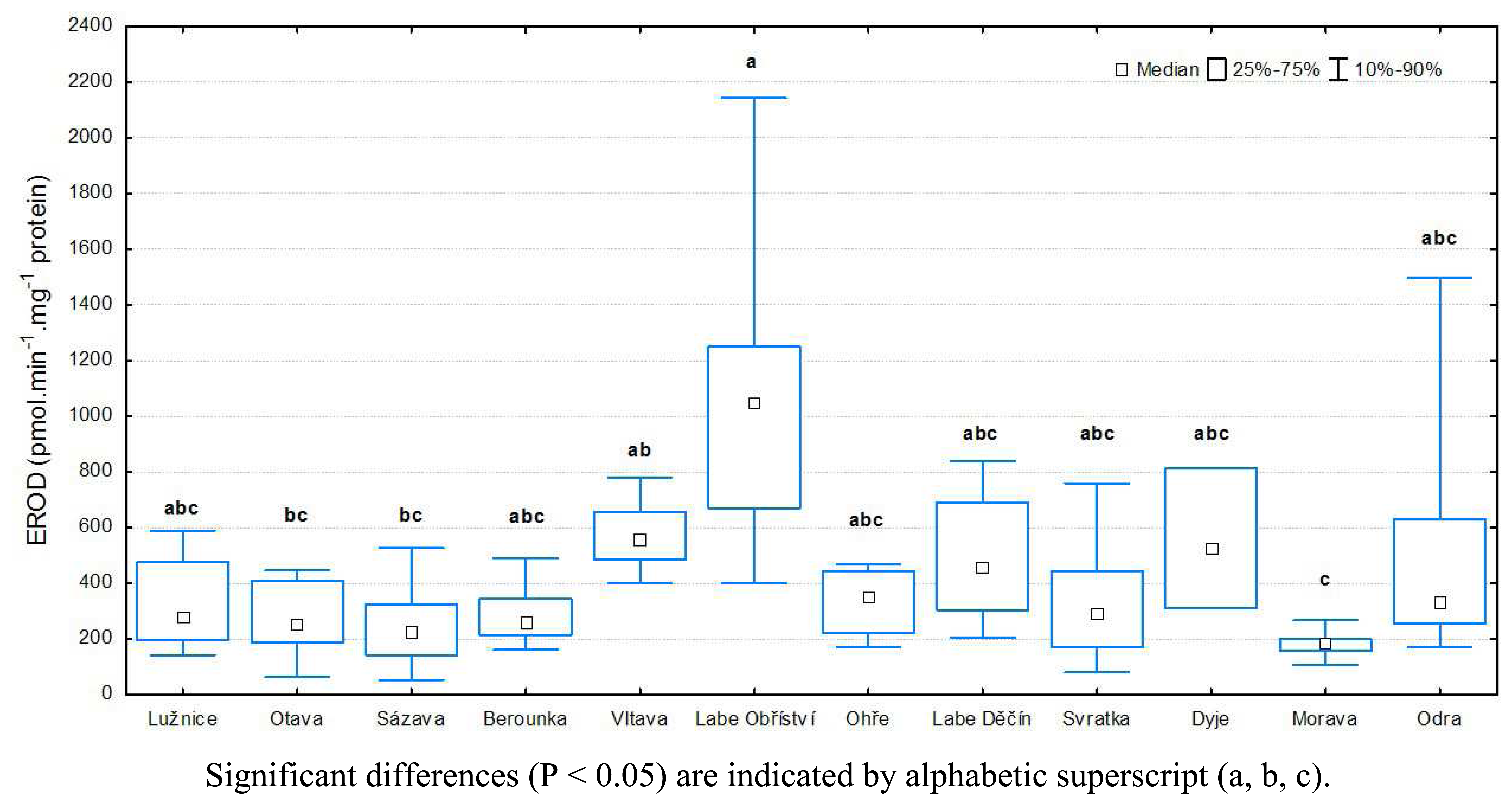
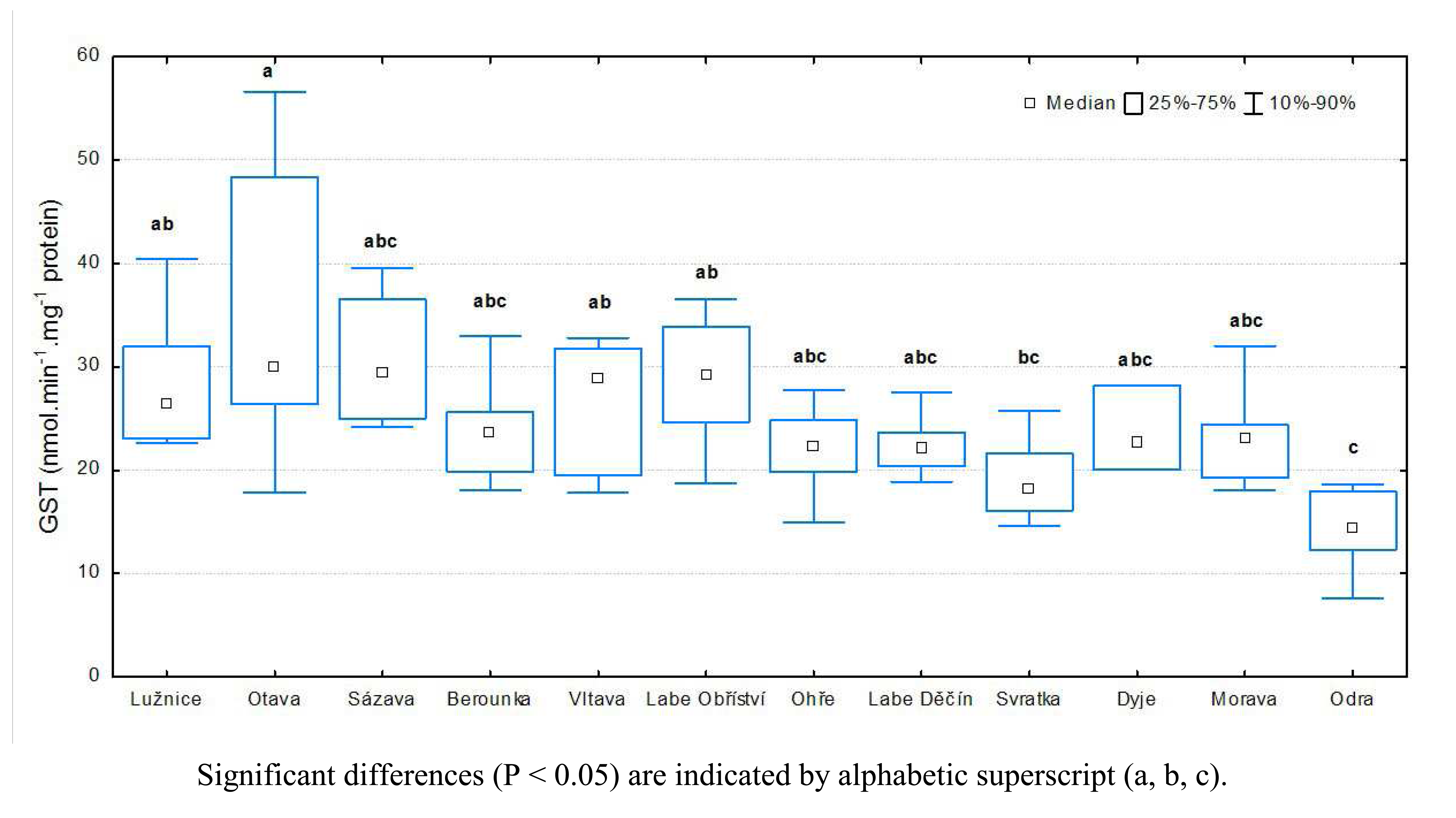
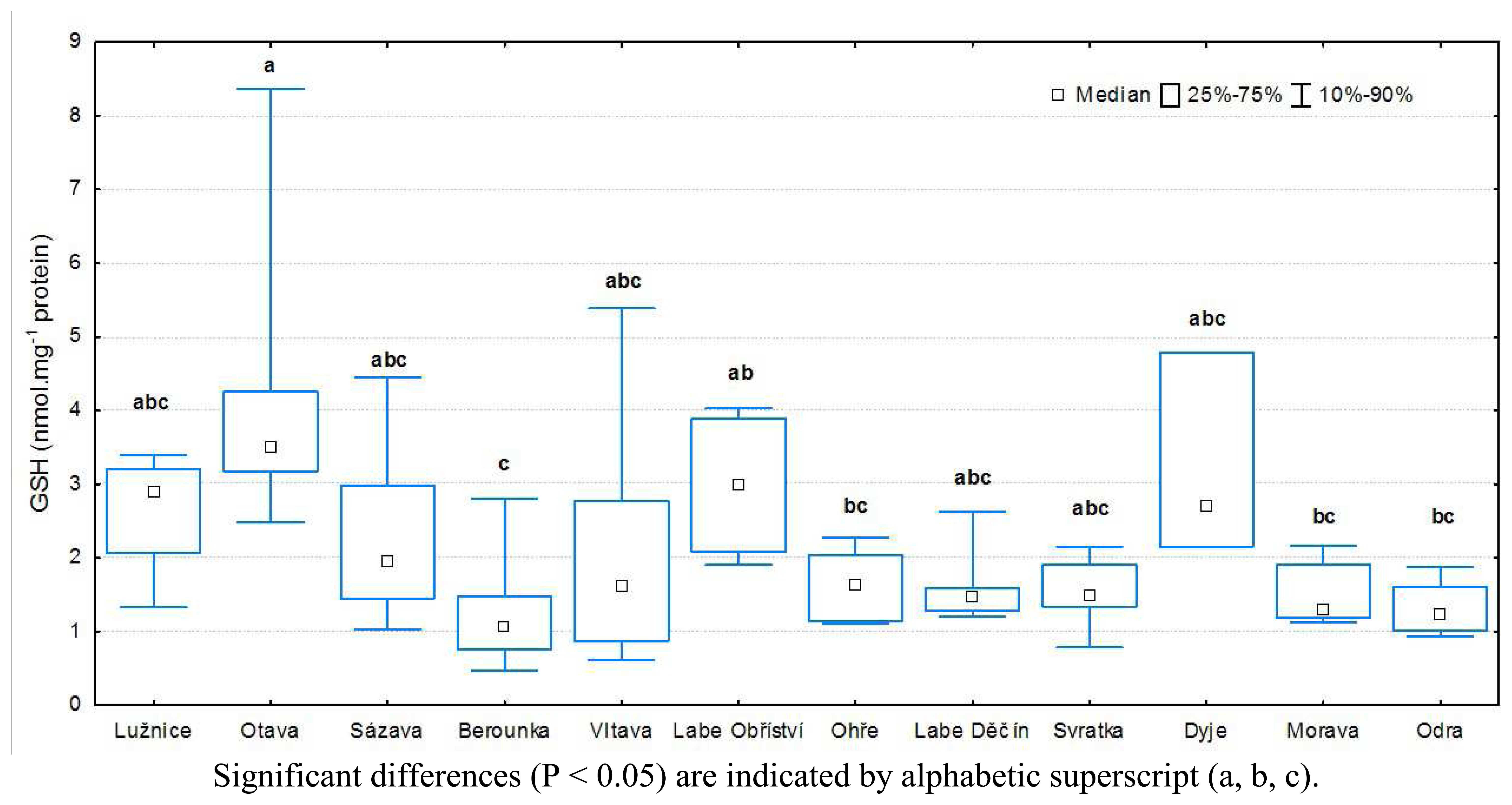
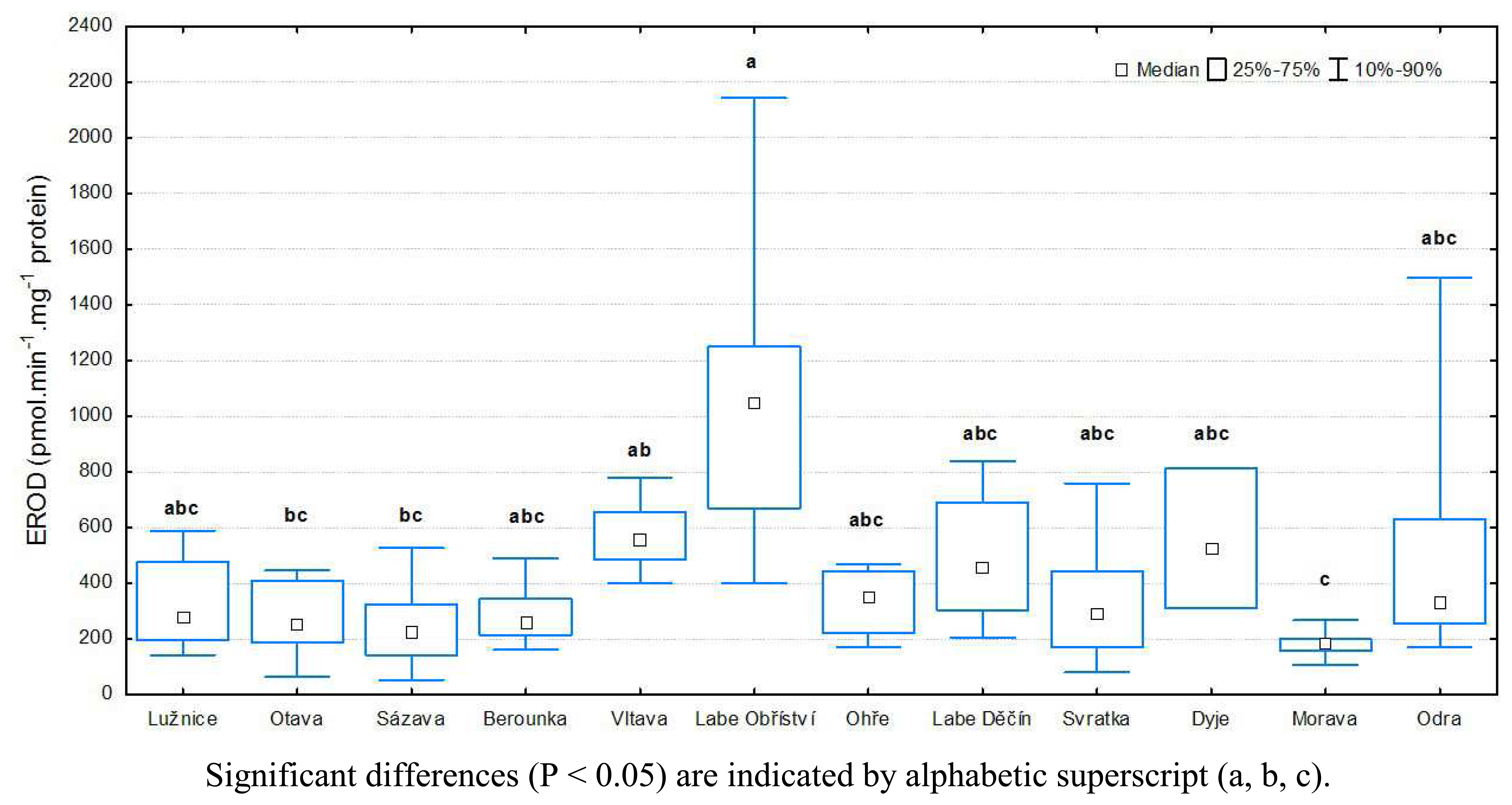
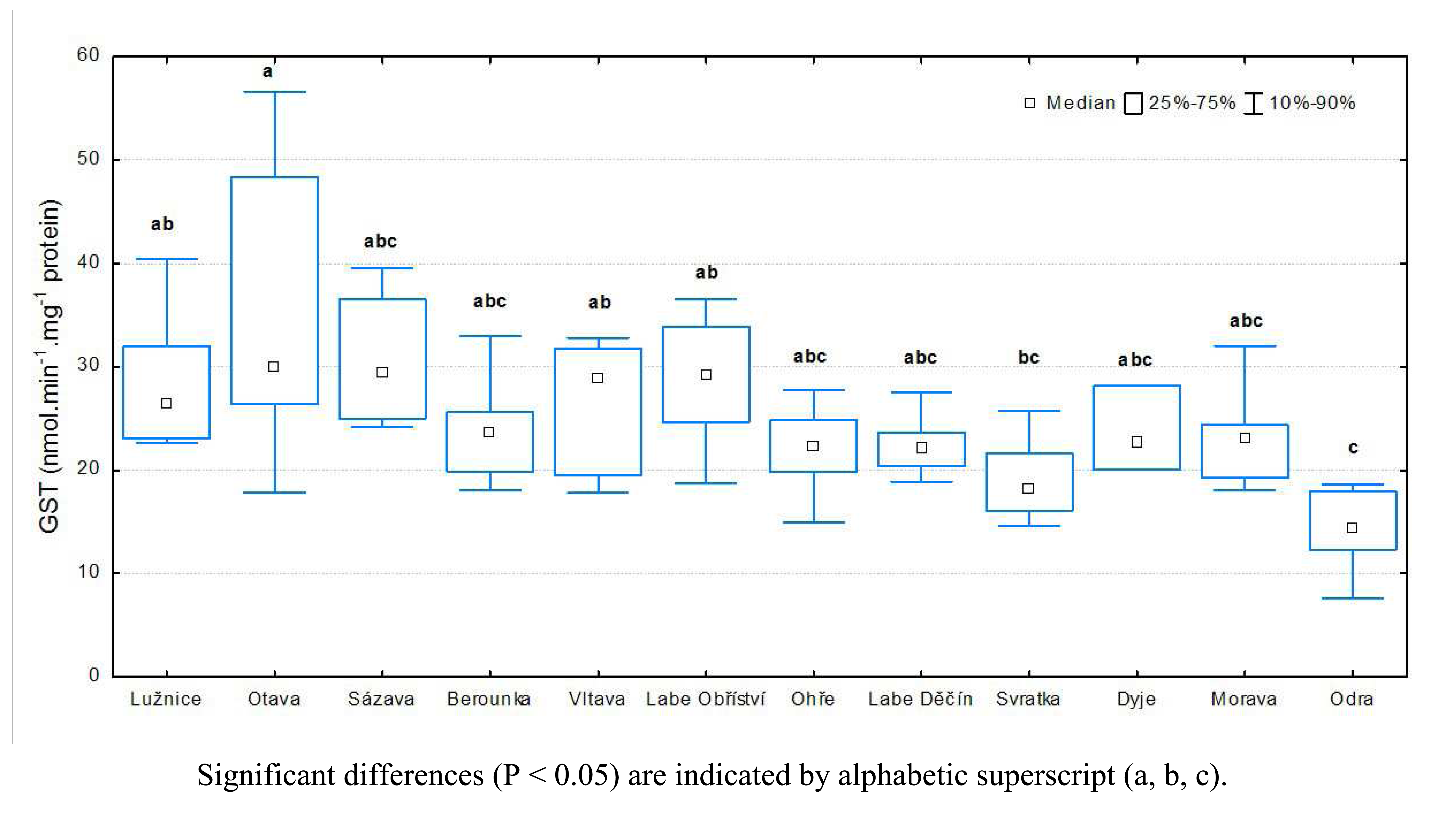
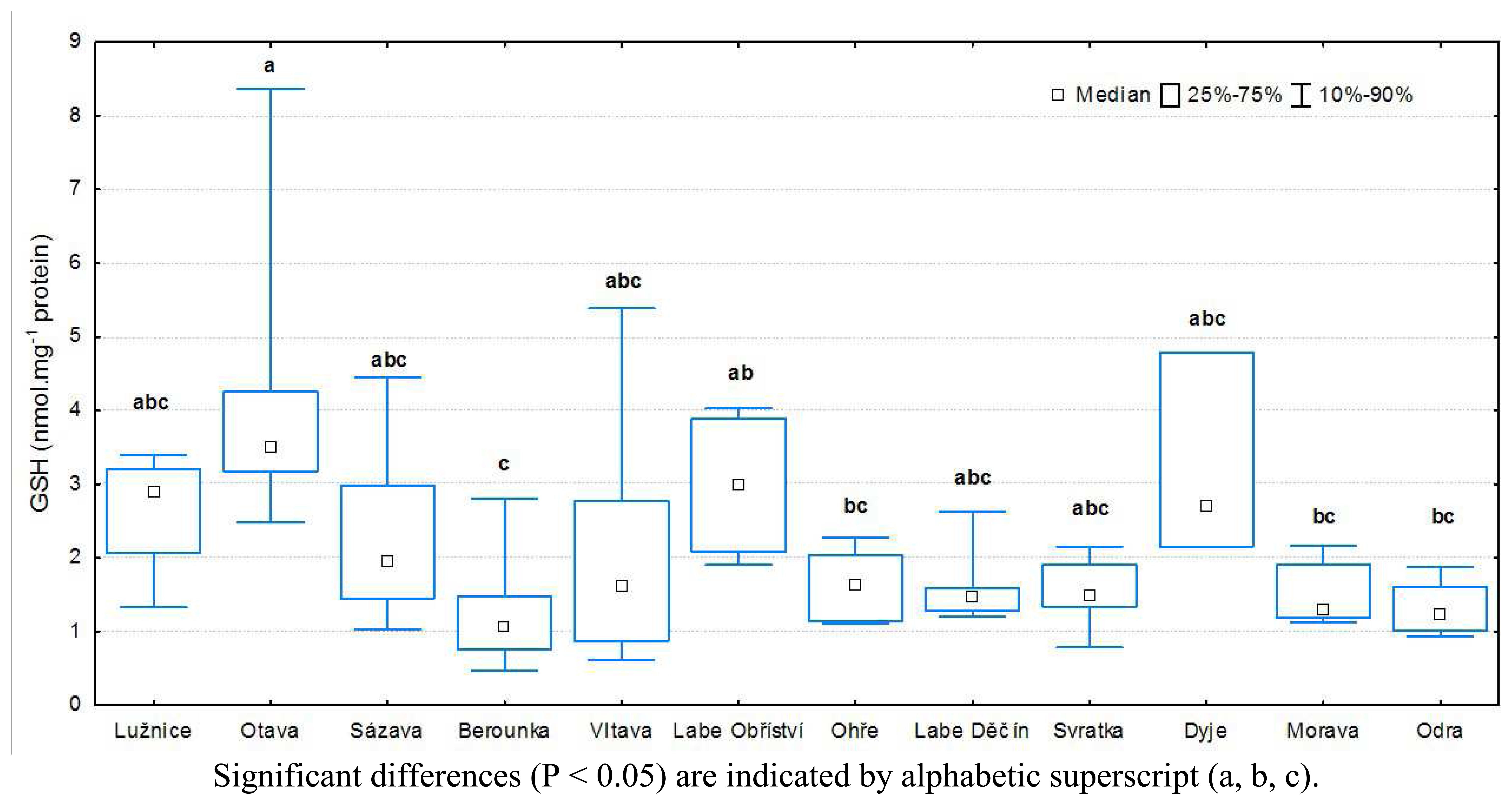
3.1 Results of biochemical analyses
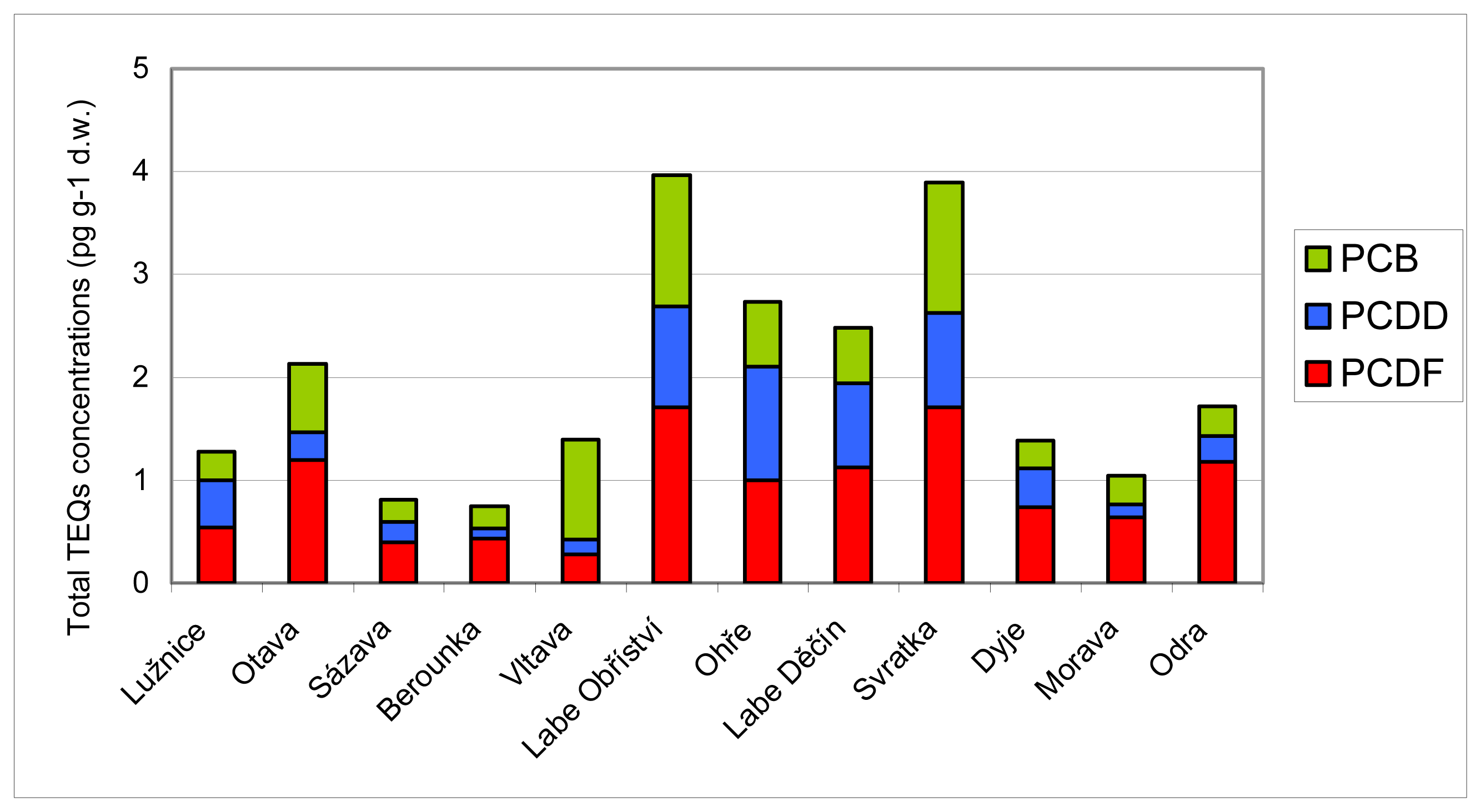
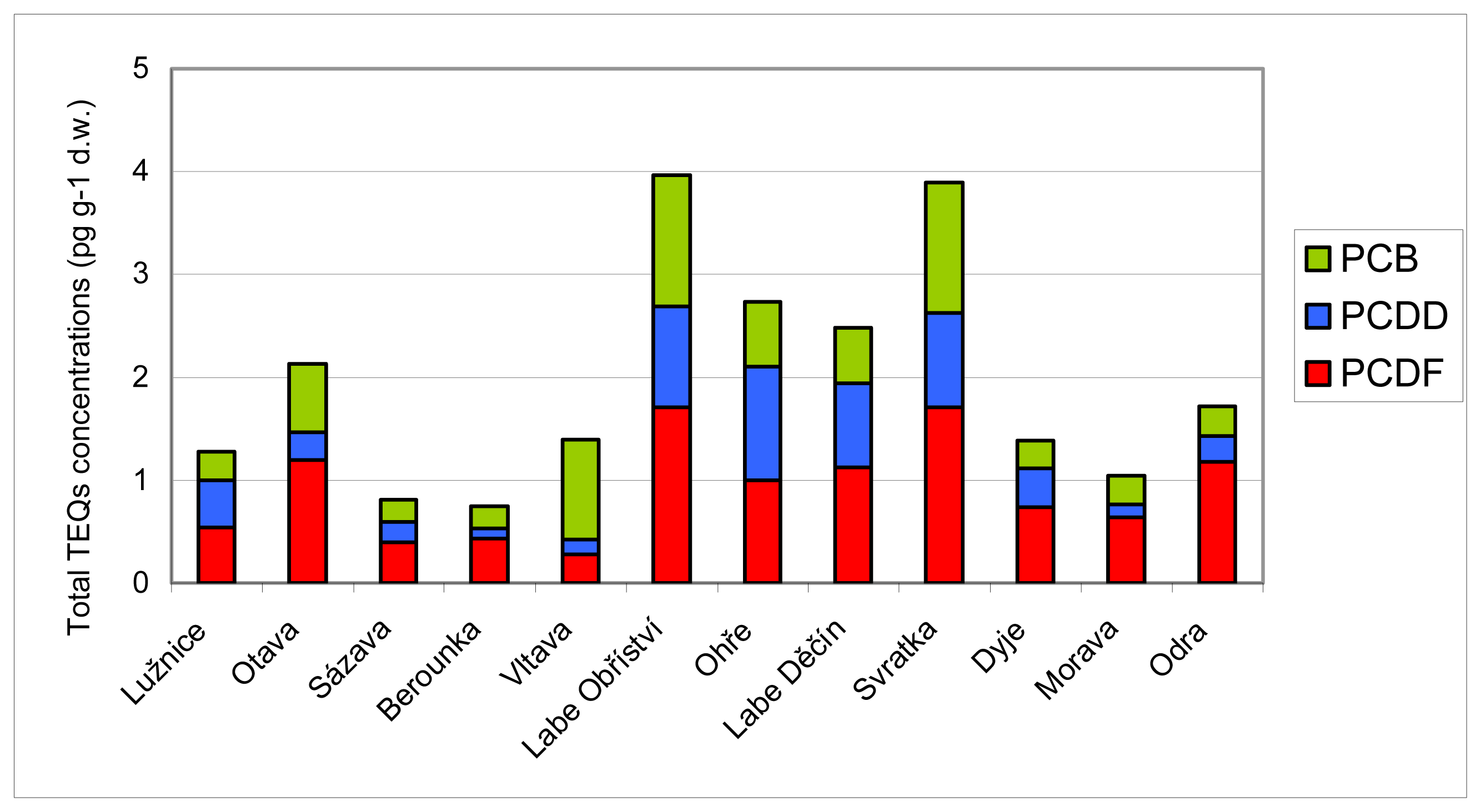
3.2 Results of chemical analyses
4. Discussion
5. Conclusions
Acknowledgments
References
- Bamgbose, O.; Jinadu, K.; Osibanjo, O. Galeoides-decadactylus - A bioindicator for chlorina hydrocarbons in the Nigerian marine-environment. J. Environ. Sci. Heal. A. 1993, 28, 321–337. [Google Scholar]
- Linde, A.R.; Arribas, P.; SanchezGalan, S.; GarciaVazquez, F. Eel (Anguilla anguilla) and brown trout (Salmo trutta) target species to assess the biological impact of trace metal pollution in freshwater ecosystems. Arch. Environ. Con. Tox. 1996, 31, 297–302. [Google Scholar]
- Ueno, D.; Iwata, H.; Tanabe, S.; Ikeda, K.; Koyama, J.; Yamada, H. Specific accumulation of persistent organochlorines in bluefin tuna collected from Japanese coastal waters. Mar. Pollut. Bull. 2002, 45, 254–261. [Google Scholar]
- Ueno, D.; Inoue, S.; Takahashi, S.; Ikeda, K.; Tanaka, H.; Subramanian, A.N.; Fillmann, G.; Lam, P.K.S.; Zheng, J.; Muchtar, M.; Prudente, M.; Chung, K.; Tanabe, S. Global pollution monitoring of butyltin compounds using skipjack tuna as a bioindicator. Environ. Pollut. 2004, 127, 1–12. [Google Scholar]
- Widianarko, B.; Van Gestel, C.A.M.; Verweij, R.A.; Van Straalen, N.M. Associations between trace metals in sediment, water, and guppy, Poecilia reticulata (Peters), from urban streams of Semarang, Indonesia. Ecotox. Environ. Safe. 2000, 46, 101–107. [Google Scholar]
- Adams, S.M.; Shepard, K.L.; Greeley, M.S.; Jimenez, B.D.; Ryon, M.G.; Shugart, L.R.; McCarthy, J.F.; Hinton, D.E. The use of bioindicators for assessing the effects of pollutant stress on fish. Mar. Environ. Res. 1989, 28, 459–464. [Google Scholar]
- Huska, D.; Krizkova, S.; Beklova, M.; Havel, L.; Zehnalek, J.; Diopan, V.; Adam, V.; Zeman, L.; Babula, P.; Kizek, R. Influence of cadmium(II) ions and brewery sludge on metallothionein level in earthworms (Eisenia fetida) - Biotransforming of toxic wastes. Sensors 2008, 8, 1039–1047. [Google Scholar]
- Krizkova, S.; Ryant, P.; Krystofova, O.; Adam, V.; Galiova, M.; Beklova, M.; Babula, P.; Kaiser, J.; Novotny, K.; Novotny, J.; Liska, M.; Malina, R.; Zehnalek, J.; Hubalek, J.; Havel, L.; Kizek, R. Multi-instrumental analysis of tissues of sunflower plants treated with silver(I) ions - Plants as bioindicators of environmental pollution. Sensors 2008, 8, 445–463. [Google Scholar]
- van der Oost, R.; Beyer, J.; Vermeulen, N.P.E. Fish bioaccumulation and biomarkers in environmental risk assessment: a review. Environ. Toxicol. Phar. 2003, 13, 57–149. [Google Scholar]
- Machala, M.; Nezveda, K.; Petrivalsky, M.; Jarosova, A.; Piacka, V.; Svobodova, Z. Monooxygenase activities in carp as biochemical markers of pollution by polycyclic and polyhalogenated aromatic hydrocarbons: choice of substrates and effects of temperature, gender and capture stress. Aquat. Toxicol. 1997, 37, 113–123. [Google Scholar]
- Bucheli, T.D.; Font, K. Induction of cytochrome-P450 as a biomarker for environmental contamination in aquatic ecosystems. Crit. Rev. Env. Sci. Tec. 1995, 25, 201–268. [Google Scholar]
- Goksoyr, A.; Forlin, L. The cytochrome-P-450 system in fish, aquatic toxicology and environmental monitoring. Aquat. Toxicol. 1992, 22, 287–311. [Google Scholar]
- Hahn, M.E.; Stegeman, J.J. Regulation of cytochrome P4501A1 in teleosts - sustained induction of CYP1A1 messenger-RNA, protein, and catalytic activity by 2,3,7,8 tetrachlorodibenzofuran in the marine fish. Stenotomus chrysops. Toxicol. Appl. Pharm. 1994, 127, 187–198. [Google Scholar]
- Payne, J.F.; Fancey, L.L.; Rahimtula, A.D.; Porter, E.L. Review and perspective on the use of mixed-function oxygenase enzymes in biological monitoring. Comp. Biochem. Phys. C. 1987, 86, 233–245. [Google Scholar]
- Eaton, D.L.; Bammler, T.K. Concise review of the glutathione S-transferases and their significance to toxicology. Toxicol. Sci. 1999, 49, 156–164. [Google Scholar]
- Gadagbui, B.K.M.; Goksoyr, A. CYP1A and other biomarker responses to effluents from a textile mill in the Volta River (Ghana) using caged tilapia (Oreochromis niloticus) and sediment-exposed mudfish (Clarias anguillaris). Biomarkers 1996, 1, 252–261. [Google Scholar]
- Otto, D.M.E.; Moon, T.W. Phase I and II enzymes and antioxidant responses in different tissues of brown bullheads from relatively polluted and non-polluted systems. Arch. Environ. Con. Tox. 1996, 31, 141–147. [Google Scholar]
- Pedrajas, J.R.; Peinado, J.; LopezBarea, J. Oxidative stress in fish exposed to model xenobiotics. Oxidatively modified forms of Cu,Zn-superoxide dismutase as potential biomarkers. Chem-Biol. Interact. 1995, 98, 267–282. [Google Scholar]
- Bello, S.M.; Franks, D.G.; Stegeman, J.J.; Hahn, M.E. Acquired resistance to Ah receptor agonists in a population of Atlantic killifish (Fundulus heteroclitus) inhabiting a marine superfund site: In vivo and in vitro studies on the inducibility of xenobiotic metabolizing enzymes. Toxicol. Sci. 2001, 60, 77–91. [Google Scholar]
- Celander, M.; Leaver, M.J.; George, S.G.; Forlin, L. Induction of cytochrome P450-1A1 and conjugating enzymes in rainbow-trout (Oncorhynchus-mykiss) liver - a time-course study. Comp. Biochem. Phys. C. 1993, 106, 343–349. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Siroka, Z.; Krijt, J.; Randak, T.; Svobodova, Z.; Peskova, G.; Fuksa, J.; Hajslova, J.; Jarkovsky, J.; Janska, M. Organic pollutant contamination of the River Elbe as assessed by biochemical markers. Acta Vet. Brno 2005, 74, 293–303. [Google Scholar]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases. First enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar]
- Grabic, R.; Novak, J.; Pacakova, V. Optimization of a GC-MS/MS method for the analysis of PCDDs and PCDFs in human and fish tissue. HRC J. High. Res. Chromatogr. 2000, 23, 595–599. [Google Scholar]
- Van den Berg, M.; Birnbaum, L.; Bosveld, A.T.C.; Brunström, B.; Cook, P.; Feeley, M.; Giesy, J.P.; Hanberg, A.; Hasegawa, R.; Kennedy, S.W.; Kubiak, T.; Larsen, J.C.; van Leeuwen, F.X.R.; Liem, A.K.D.; Nolt, C.; Peterson, R.E.; Poellinger, L.; Safe, S.; Schrenke, D.; Tillitt, D.; Tysklind, M.; Younes, M.; Wærn, F.; Zacharewski, T. Toxic Equivalency Factors (TEFs) for PCBs, PCDDs, PCDFs for humans and wildlife. Environ. Health. Persp. 1989, 106, 775–792. [Google Scholar]
- Agradi, E.; Baga, R.; Cillo, F.; Ceradini, S.; Heltai, D. Environmental contaminants and biochemical response in eel exposed to Po river water. Chemosphere 2000, 41, 1555–1562. [Google Scholar]
- Schlezinger, J.J.; Stegeman, J.J. Induction of cytochrome P450 1A in the American Eel by model halogenated and non-halogenated aryl hydrocarbon receptor agonists. Aquat. Toxicol. 2000, 50, 375–386. [Google Scholar]
- Stagg, R.M.; Rusin, J.; McPhail, M.E.; McIntosh, A.D.; Moffat, C.F.; Craft, J.A. Effects of polycyclic aromatic hydrocarbons on expression of CYP1A in salmon (Salmo salar) following experimental exposure and after the Braer oil spill. Environ. Toxicol. Chem. 2000, 19, 2797–2805. [Google Scholar]
- Vanderweiden, M.E.J.; Vanderkolk, J.; Bleumink, R.; Seinen, W.; Vandenberg, M. Concurrence of P450-1A1 induction and toxic effects after administration of a low-dose of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) in the rainbow-trout (Oncorhynchus-mykiss). Aquat. Toxicol. 1992, 24, 123–142. [Google Scholar]
- Al-Arabi, S.A.M.; Adolfsson-Erici, M.; Waagbo, R.; Ali, M.S.; Goksoyr, A. Contaminant accumulation and biomarker responses in caged fish exposed to effluents from anthropogenic sources in the Karnaphuly River, Bangladesh. Environ. Toxicol. Chem. 2005, 24, 1968–1978. [Google Scholar]
- Behrens, A.; Segner, H. Cytochrome P4501A induction in brown trout exposed to small streams of an urbanised area: results of a five-year-study. Environ. Pollut. 2005, 136, 231–242. [Google Scholar]
- Curtis, L.R.; Carpenter, H.M.; Donohoe, R.M.; Williams, D.E.; Hedstrom, O.R.; Deinzer, M.L.; Bellstein, M.A.; Foster, E.; Gates, R. Sensitivity of cytochrome-P450-1A1 induction in fish as a biomarker for distribution of TCDD and TCDF in the Willamette River, Oregon. Environ. Sci. Technol. 1993, 27, 2149–2157. [Google Scholar]
- Monod, D.; Devaux, A.; Riviere, J.L. Effects of chemical pollution on the activities of hepatic xenobiotic metabolizing enzymes in fish from the River Rhone. Sci. Total. Environ. 1988, 73, 189–201. [Google Scholar]
- Adams, M.S.; Kausch, H.; Gaumert, T.; Kruger, K.E. The effect of the reunification of Germany on the water chemistry and ecology of selected rivers. Environ. Conserv. 1996, 23, 35–43. [Google Scholar]
- Gandrass, J.; Zoll, M. Chlorinated hydrocarbons in sediments of the Elbe catchment area -Analytical methods and status of pollution. Acta Hydroch. Hydrob. 1996, 24, 212–217. [Google Scholar]
- Heininger, P.; Pelzer, J. Trends and patterns in the contamination of sediments from federal waterways in Eastern Germany. Acta Hydroch. Hydrob. 1998, 26, 218–225. [Google Scholar]
- Lehmann, A.; Rode, M. Long-term behaviour and cross-correlation water quality analysis of the river Elbe, Germany. Water. Res. 2001, 35, 2153–2160. [Google Scholar]
- Oetken, M.; Stachel, B.; Pfenninger, M.; Oehlmann, J. Impact of a flood disaster on sediment toxicity in a major river system - the Elbe flood 2002 as a case study. Environ. Pollut. 2005, 134, 87–95. [Google Scholar]
- Havelkova, M.; Randak, T.; Zlabek, V.; Krijt, J.; Kroupova, H.; Pulkrabova, J.; Svobodova, Z. Biochemical markers for assessing aquatic contamination. Sensors 2008, 7, 2599–2611. [Google Scholar]
- Nishimoto, M.; Leeberhart, B.T.; Sanborn, H.R.; Krone, C.; Varanasi, U.; Stein, J.E. Effects of a complex mixture of chemical contaminants on hepatic glutathione, L-cysteine and gamma-glutamylcysteine synthetase in English sole (Pleuronectes vetulus). Environ. Toxicol. Chem. 1995, 14, 461–469. [Google Scholar]
- Eufemia, N.A.; Collier, T.K.; Stein, J.E.; Watson, D.E.; DiGiulio, R.T. Biochemical responses to sediment-associated contaminants in brown bullhead (Ameriurus nebulosus) from the Niagara River ecosystem. Ecotoxicology 1997, 6, 13–34. [Google Scholar]
- Petrivalsky, M.; Machala, M.; Nezveda, K.; Piacka, V.; Svobodova, Z.; Drabek, P. Glutathione-dependent detoxifying enzymes in rainbow trout liver: Search for specific biochemical markers of chemical stress. Environ. Toxicol. Chem. 1997, 16, 1417–1421. [Google Scholar]
- Vanderoost, R.; Vangastel, L.; Worst, D.; Hanraads, M.; Satumalay, K.; Vanschooten, F.J.; Heida, H.; Vermeulen, N.P.E. Biochemical markers in feral roach (Rutilus-rutilus) in relation to the bioaccumulation of organic trace pollutants. Chemosphere 1994, 29, 801–817. [Google Scholar]
- Vigano, L.; Arillo, A.; Melodia, F.; Bagnasco, M.; Bennicelli, C.; Deflora, S. Hepatic and biliary biomarkers in rainbow-trout injected with sediment extracts from the River Po (Italy). Chemosphere 1995, 30, 2117–2128. [Google Scholar]
- Armknecht, S.L.; Kaattari, S.L.; Van Veld, P.A. An elevated glutathione S-transferase in creosote-resistant mummichog (Fundulus heteroclitus). Aquat. Toxicol. 1998, 41, 1–16. [Google Scholar]
- Lenartova, V.; Holovska, K.; Javorsky, P. The influence of environmental pollution on the SOD and GST-isoenzyme patterns. Water. Sci. Technol. 2000, 42, 209–214. [Google Scholar]
- Tuvikene, A.; Huuskonen, S.; Koponen, K.; Ritola, O.; Mauer, U.; Lindstrom-Seppa, P. Oil shale processing as a source of aquatic pollution: Monitoring of the biologic effects in caged and feral freshwater fish. Environ. Health. Persp. 1999, 107, 745–752. [Google Scholar]
- Vigano, L.; Arillo, A.; Melodia, F.; Arlati, P.; Monti, C. Biomarker responses in cyprinids of the middle stretch of the River Po, Italy. Environ. Toxicol. Chem. 1998, 17, 404–411. [Google Scholar]
- Hamed, R.R.; Farid, N.M.; Elowa, S.H.E.; Abdalla, A.M. Glutathione related enzyme levels of freshwater fish as bioindicators of pollution. Environmentalist 2003, 23, 313–322. [Google Scholar]
- Mannervik, B.; Danielson, U.H. Glutathione transferases - structure and catalytic activity. CRC Cr. Rev. Bioch. Mol. 1988, 23, 283–337. [Google Scholar]
- Bozcaarmutlu, A.; Arinc, E. Inhibitory effects of divalent metal ions on liver microsomal 7-ethoxyresorufin-O-deethylase (EROD) activity of leaping mullet. Mar. Environ. Res. 2004, 58, 521–524. [Google Scholar]
- Fent, K.; Bucheli, T.D. Inhibition of hepatic-microsomal monooxygenase system by organotins in-vitro in fresh-water fish. Aquat. Toxicol. 1994, 28, 107–126. [Google Scholar]
- Forlin, L.; Haux, C.; Karlsson-Norrgren, L.; Runn, P.; Larsson, A. Biotransformation enzyme-activities and histopathology in rainbow trout, Salmo-Gairdneri, treated with cadmium. Aquat. Toxicol. 1986, 8, 51–64. [Google Scholar]
- Risso-de Faverney, C.; Lafaurie, M.; Girard, J.P.; Rahmani, R. Effects of heavy metals and 3-methylcholanthrene on expression and induction of CYP1A1 and metallothionein levels in trout (Oncorhynchus mykiss) hepatocyte cultures. Environ. Toxicol. Chem. 2000, 19, 2239–2248. [Google Scholar]
- Stien, X.; Risso, C.; GnassiaBarelli, M.; Romeo, M.; Lafaurie, M. Effect of copper chloride in vitro and in vivo on the hepatic EROD activity in the fish Dicentrarchus labrax. Environ. Toxicol. Chem. 1997, 16, 214–219. [Google Scholar]
- Flammarion, P.; Garric, J. Cyprinids EROD activities in low contaminated rivers: A relevant statistical approach to estimate reference levels for EROD biomarker? Chemosphere 1997, 35, 2375–2388. [Google Scholar]
- Koivusaari, U.; Harri, M.; Hanninen, O. Seasonal-variation of hepatic biotransformation in female and male rainbow-trout (Salmo-gairdneri). Comp. Biochem. Phys. C. 1981, 70, 149–157. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location River (Municipality) (distance from site to mouth of river) | n | Weight±SD (g) | Age (years) mean (range) |
|---|---|---|---|
| Lužnice (Bechyně) (11 kms) | 8 | 614.4±155.0 | 4.9 (4–6) |
| Otava (Topělec) (20 kms) | 9 | 360.6±253.0 | 4.2 (3–6) |
| Sázava (Nespeky) (27.5 kms) | 8 | 278.1±63.9 | 3.8 (3–4) |
| Berounka (Srbsko) (29 kms) | 8 | 292.5±99.5 | 3.6 (3–5) |
| Vltava (Zelčín) (5 kms) | 10 | 383.5±199.3 | 4.0 (3–7) |
| Labe (Obříství) (122 kms)* | 8 | 306.3±144.9 | 4.1 (3–6) |
| Ohře (Terezín) (3 kms) | 10 | 540.5±201.8 | 4.3 (3–7) |
| Labe (Děčín) (21 kms)* | 8 | 546.3±211.0 | 4.8 (4–7) |
| Svratka (Židlochovice) (23 kms) | 8 | 243.8±94.2 | 3.5 (3–4) |
| Dyje (Pohansko) (16 kms) | 3 | 626.7±558.2 | 4.7 (3–7) |
| Morava (Lanžhot) (9.5 kms) | 8 | 304.4±110.6 | 3.1 (2–4) |
| Odra (Bohumín) (9 kms)** | 8 | 149.4±78.1 | 2.8 (2–4) |
© 2008 by MDPI (http://www.mdpi.org). Reproduction is permitted for noncommercial purposes.
Share and Cite
Havelkova, M.; Blahova, J.; Kroupova, H.; Randak, T.; Slatinska, I.; Leontovycova, D.; Grabic, R.; Pospisil, R.; Svobodova, Z. Biomarkers of Contaminant Exposure in Chub (Leuciscus cephalus L.) – Biomonitoring of Major Rivers in the Czech Republic. Sensors 2008, 8, 2589-2603. https://doi.org/10.3390/s8042589
Havelkova M, Blahova J, Kroupova H, Randak T, Slatinska I, Leontovycova D, Grabic R, Pospisil R, Svobodova Z. Biomarkers of Contaminant Exposure in Chub (Leuciscus cephalus L.) – Biomonitoring of Major Rivers in the Czech Republic. Sensors. 2008; 8(4):2589-2603. https://doi.org/10.3390/s8042589
Chicago/Turabian StyleHavelkova, Marcela, Jana Blahova, Hana Kroupova, Tomas Randak, Iveta Slatinska, Drahomira Leontovycova, Roman Grabic, Richard Pospisil, and Zdenka Svobodova. 2008. "Biomarkers of Contaminant Exposure in Chub (Leuciscus cephalus L.) – Biomonitoring of Major Rivers in the Czech Republic" Sensors 8, no. 4: 2589-2603. https://doi.org/10.3390/s8042589