1. Introduction
More than 130 different pesticides are used worldwide for the production of tobacco, with approximately 25 compounds used on a regular basis. They belong to different chemical groups, such as organophosphates, carbamates, organochlorine and heterocyclic pesticides, nitro compounds, pyrethroids and amides. Many of these compounds can cause moderate to severe respiratory and neurological damage or act as genotoxic and carcinogenic agents, therefore increasing the health risks associated with smoking. Organophosphate (such as chlorpyrifos) and carbamate pesticides (such as carbaryl) in particular, affect the nervous system by inhibiting acetylcholine esterase (AChE), an enzyme that is important for the transmission of nerve impulses [
1,
2]. Furthermore, compared to other crops, tobacco leaves have more surface to weight ratio [
3] and with excessive use of pesticides there is always a chance for accumulation of pesticide residues in the leaf tissue.
The conventional analysis of pesticide residues in agricultural commodities is a labor-intensive procedure, since it is necessary to cover a wide range of different chemicals, using a single procedure [
4]. Standard analysis methods include extensive sample pretreatment (with solvent extraction and partitioning phases) and determination by gas chromatography (GC), high-pressure liquid chromatography (HPLC) and mass spectrometry (MS) to achieve the necessary selectivity and sensitivity for the different classes of compounds under detection [
5]. As a consequence, current methods of analysis provide a limited sample analysis capacity, on a day/instrument basis. Therefore, rapid pesticide residue testing is necessary. In a region-specific pattern, this results to a general lack of resources for implementation and enforcement of environmental, pesticide and labour regulations in tobacco-producing countries.
A biosensor is a device that detects, transmits and records information regarding a physiological or biochemical change. In recent years there has been a rapid increase in the number of diagnostic applications based on biosensors, including live, intact cells and -in some cases- tissues, organs or whole organisms. A cell-based sensor design employs the physiological responses of whole living cells as the sensing component, namely oxygen consumption, surface chemical or electrical potential, mobility or genetic activity [
6].
A number of biological methods and protocols for toxicity measurement (including pesticide residues) have been developed, based on microrganisms such as bacteria and yeast. The advantages of toxicity bioassays include their low cost, easy detection protocol, reproducibility and rapid response. In addition, bioassays offer direct toxicity information on tested chemicals or samples that analytical methods cannot provide. Cairns and Mount [
7] stated that chemical concentrations can be measured with an instrument, but living organisms can measure toxicity only. The number of toxic compounds that can be assayed by cell-based sensors is virtually endless. Current cell-based sensors for environmental monitoring can detect heavy metals or their salts, organic compounds, various carcinogens and pesticides such as diuron, atrazine, organotin compounds, organophosphates, glyphosate and various antibiotics (for a review see [
8,
9]). However, it must be mentioned that cell sensors commonly are non-selective, not being able to discriminate between individual bioactive compounds. Also, cell-based sensors must provide for an environment maintaining sufficient cell viability (e.g. CO
2-enriched culture atmosphere).
Commercial environmental cell biosensors utilize luminescence from natural or genetically modified microbial species, whereas specificity of the response against an analyte is based on the fusion of reporter genes, with a metabolic operon induced by its respective analyte. Bioluminescence-based systems have been extensively used to monitor cellular stress responses, either by inducing the response at low analyte concentrations or by inhibiting the luminescence at higher concentrations [
5,
10-
15].
Fluorescence-based systems are not always suitable for quantitative determinations. They are also far more expensive than amperometric techniques [
16]. Fluorescent dyes may also have considerable side-effects on cellular physiology, such as uncoupling or respiratory inhibition and membrane permeabilization [
17,
18] and may interfere with the cell parameter that they assay [
19].
Approached from different angles, electrically active cells can be interfaced with microelectrodes which allow the capture of extracellular spikes or impedance changes associated with cellular or tissue response. An example is the Bioelectric Recognition Assay (BERA) which has been originally developed for the detection of viruses on the basis of their specific interaction with appropriately immobilized, mammalian cells and the measurement of the change of the electric potential that is caused by the aforementioned interaction [
9,
20,
21].
In the present study, we developed a cellular biosensor based on the BERA working principle for detecting organophosphate and carbamate pesticide residues in tobacco. The sensor is based on neuroblastoma N2a cells. The presence of pesticidal compounds is detected by the degree of inhibition of AChE, which is directly associated with changes of the immobilized cell membrane potential after administration of acetylcholine (ACh).
2. Experimental Section
2.1. Materials
Mouse neuroblastoma (N2a) cell cultures were originally provided from LGC Promochem (UK). Acetylcholine iodide (ACh) was purchased from Sigma Chem. Co. (USA). Fresh stock solutions of 14 mM S-acetylthiocholine-iodide (ATCi; Alfa Aesar Gmbh & Co KG) were made weekly in distilled water and kept at 4°C in the dark. Dithiobisnitrobenzoic acid (DTNB (Ellman's reagent; Aldrich) was used as an indicator for ATCi hydrolysis. Fresh stock solutions were made monthly in P-buffer (0.1 M KH2PO4/K2HPO4; pH = 7,4) and kept at 4°C in the dark. Clorpyrifos (O,O-diethyl O-3,5,6-trichloro-2-pyridyl phosphorothioate; CAS [2921-88-2]; Mw = 350.59; DowAgroSciences, USA) was used as a standard organophosphate insecticide and carbaryl (1-naphthyl methylcarbamate; CAS [63-25-2]; Mw = 201.221 Bayer Cropscience, France) as a standard carbamate insecticide. Pesticide mixes which contained 10 μM of each pesticide, were prepared daily in acetone solution. All other reagents were purchased from Fluka (Switzerland).
2.2. Sensor fabrication from N2a cells
Cells were cultured in Dulbecco's medium with 10% heat-inactivated foetal calf serum (FCS), 10% antibiotics (streptomycin) and 10% l-glutamine. After cell detachment from the culture vessel by adding trypsine/EDTA for 10 min at 37°C and cell concentration by centrifugation (6 min, 1200 rpm, 25 °C), 1 ml of cells (at a density of 2.5×106/ml) were mixed with 2ml of 4% (w/v) sodium alginate solution and then the mixture was added drop wise, by means of a 22G syringe, in 0.8M CaCl2. Each of the resulting calcium alginate beads had an approximate diameter of 2mm and contained approximately 50×103 cells.
2.3. Sample preparation
Tobacco leaves (of the oriental cultivar Basmas) were kindly provided by the local tobacco processing company SEKE S.A. in blended dry form. Leaves were derived from plants grown at thirteen different locations in Northern Greece.
Control samples contained strictly organically-grown tobacco, which was free of pesticide residues.
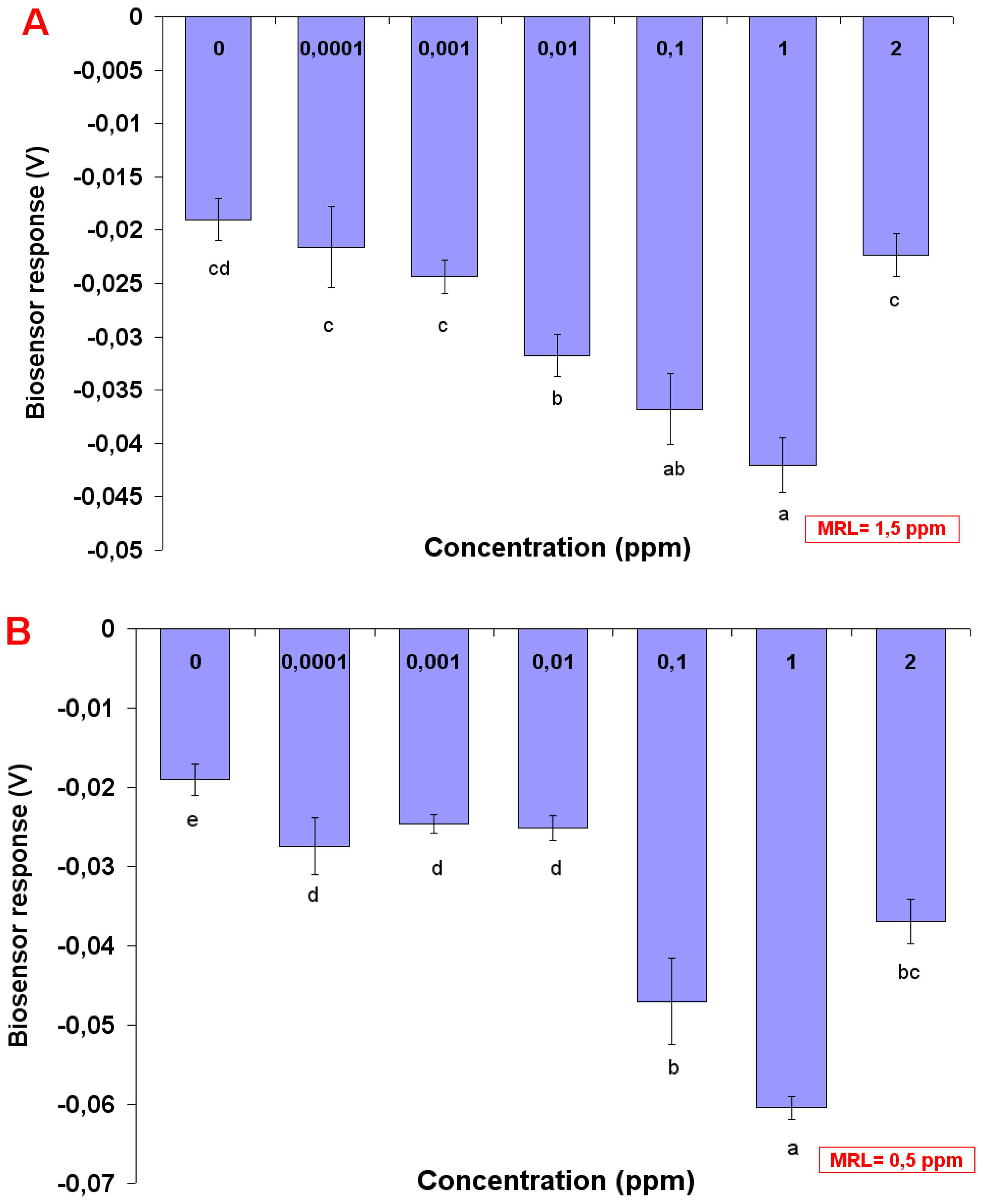
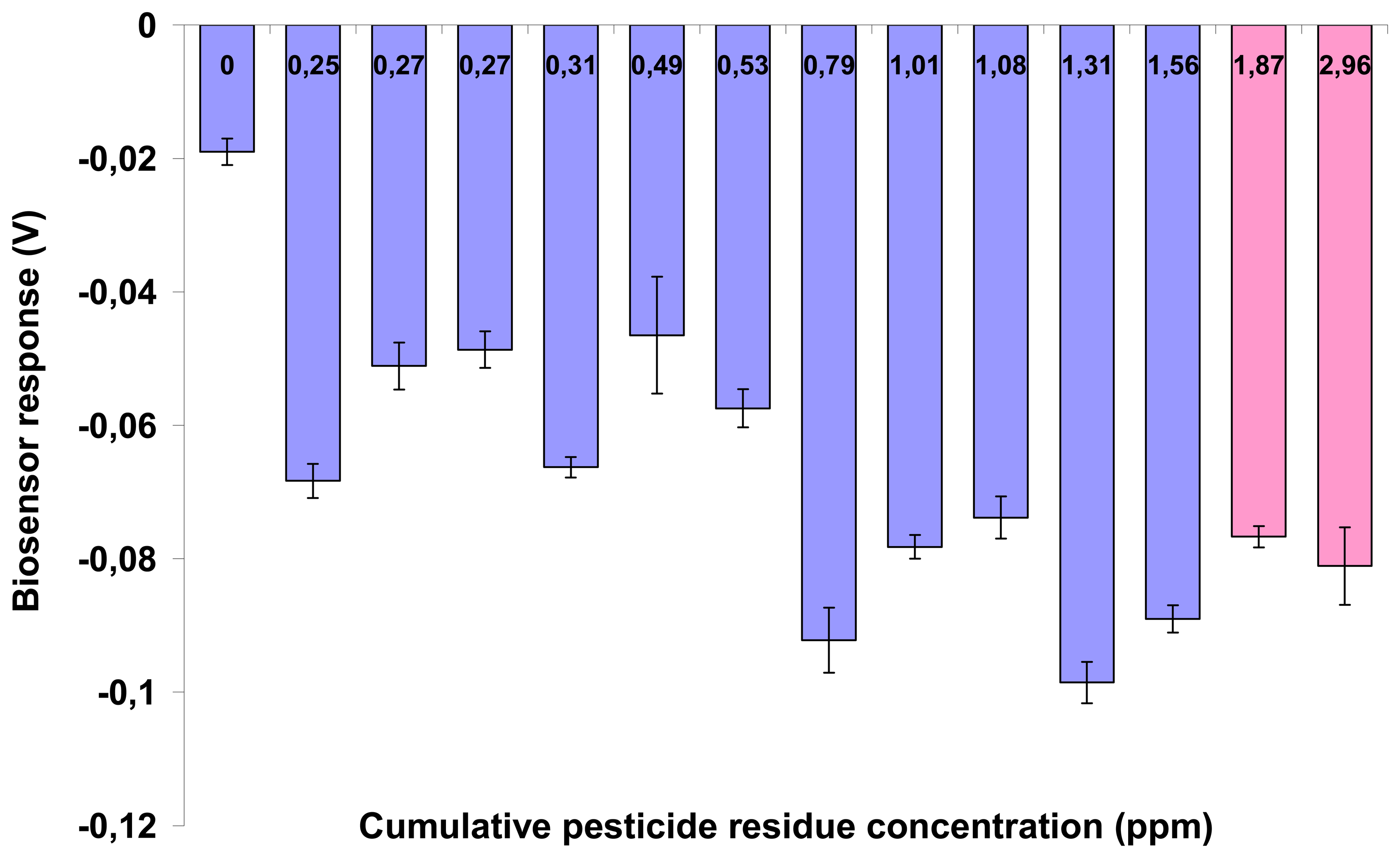
Standard solutions of either chlorpyriphos or carbaryl were prepared by spiking control (pesticide-free) tobacco samples with the respective pesticide at concentrations of 0.0001, 0.001, 0.01, 0.1, 1 and 2 ppm. This was done in order to avoid the so-called “matrix effect”, i.e. the interference of sample-specific components on the sensor performance. In this way, the baseline (control) response of the sensor would reflect the effect of plant constituents on the sensor.
In all samples the solvent used for extraction was water:acetone = 1:3 (v/v). A portion of ground tobacco (4 g) was placed in a 60 mL conical vial after adding 40 ml of the extraction solvent. The extract was placed under 37°C in a hot bath for 1 hour in order to allow the acetone evaporation. The supernatant was then collected and transferred to 1.5 mL eppendorfs.
Each sample solution consisted of 50 μl tobacco extract and 10μl acetylcholine iodide. Acetylcholine iodide (10 mM) was dissolved in a 50mM Tris-aminomethane buffer (Tris) of pH 8.
2.4. Assay principle
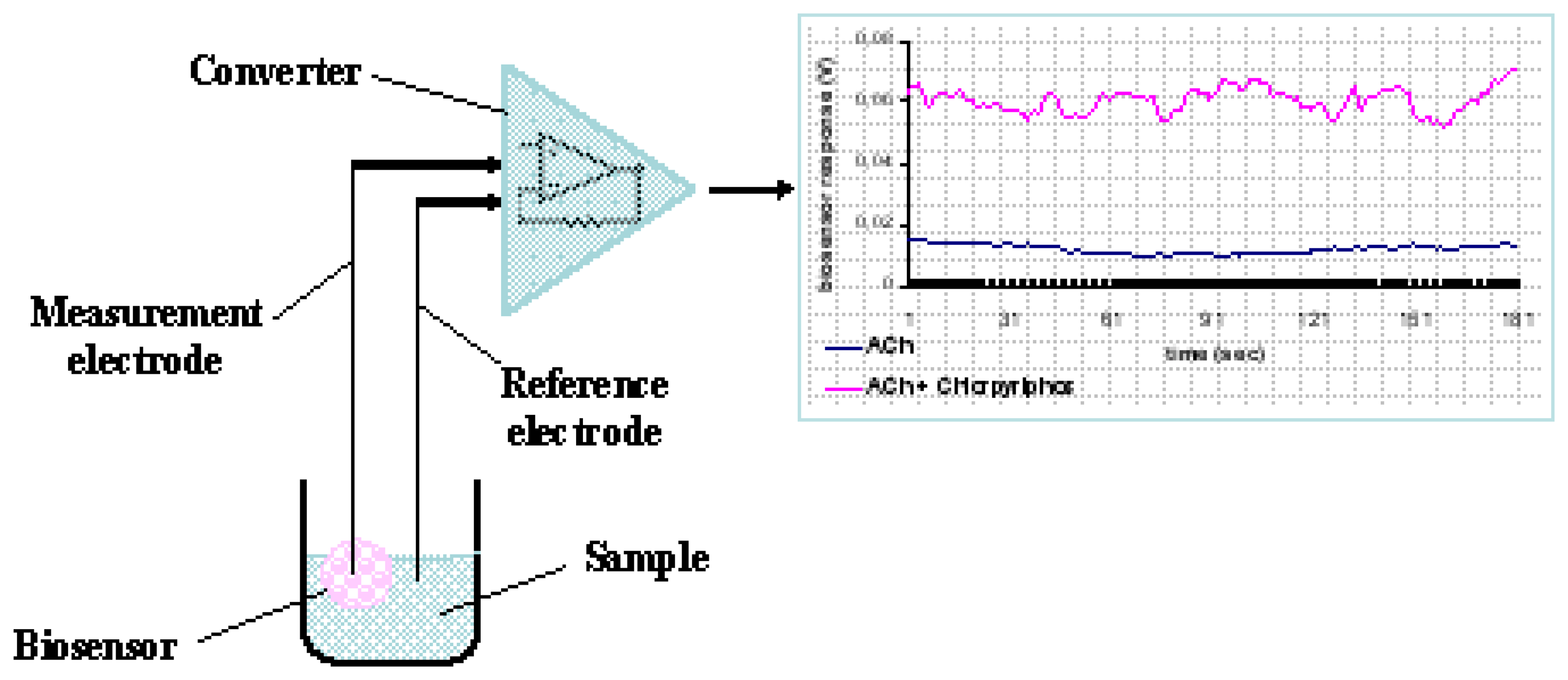
BERA sensors based on N2a cells were used for assaying the inhibition of cellular AChE to hydrolyze Ach to acetate and choline according to the following equation:
According to the working principle of the method, the presence of organophosphate or carbamate compounds is detected by the degree of inhibition of cellular AChE, which is inversely associated with ACh concentration. ACh is an excitatory neurotransmitter [
22], the activity of which is regulated by AChE. Therefore, inhibition of AChE can lead in increased excitatory ACh transmission, which can be measured by the depolarization of the cell membrane. In other words, inhibition of AChE by pesticide residues in the sample will result to excessive stimulation of N2a cells by Ach, which will further lead to membrane depolarization above a pre-determined threshold. Membrane depolarization events and associated electrolyte influx/efflux will reflect themselves on the sensor's response as a change of the sensor's potential, due to changes in the concentration of electrolytes in the immediate vicinity of the working electrode.
2.5. Assay procedure
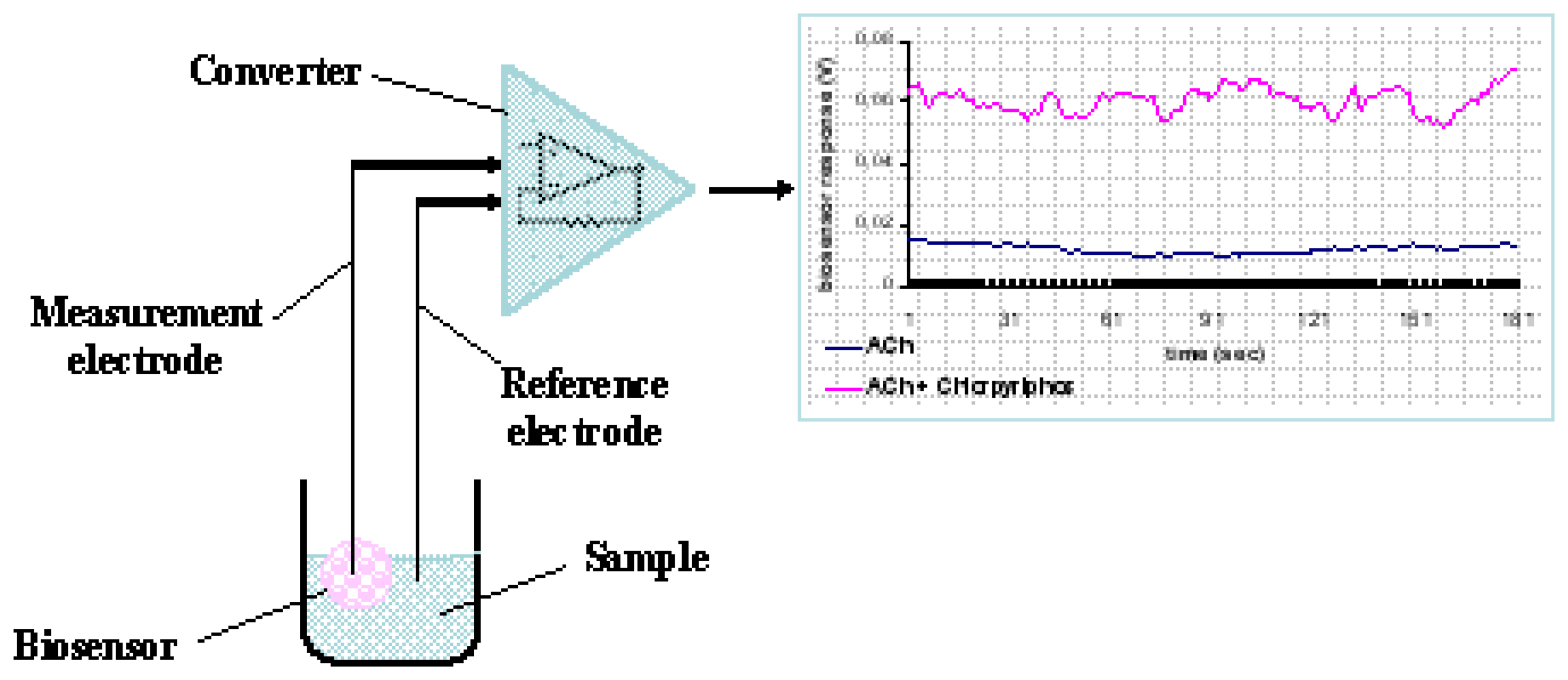
Each cell-bearing bead (cell sensor) was connected to a working electrode made from pure silver, electrochemically coated with an Ag/AgCl layer and having a diameter of 0.75 mm. Electrodes were connected to the recording device, which comprised the PMD-1608FS A/D card (Measurement Computing, Middleboro, MA). The software responsible for recording the signal and data processing was InstaCal (Measurement Computing) (
Fig. 1).
For each assay, the sensor system, comprising of the bead attached to the working electrode and a reference electrode, was immersed into each sample solution (200 μl). The response of each sensor was estimated by recording the average change of the sensor potential for a period of 180 sec after sample application.
2.6. Conventional sample analysis
Tobacco samples were also analyzed by GC and LC-MS/MS according to the International Standard ISO 4389 [
23] and by following the protocols of Djordjevic et al. [
24] and Haib et al. [
4].
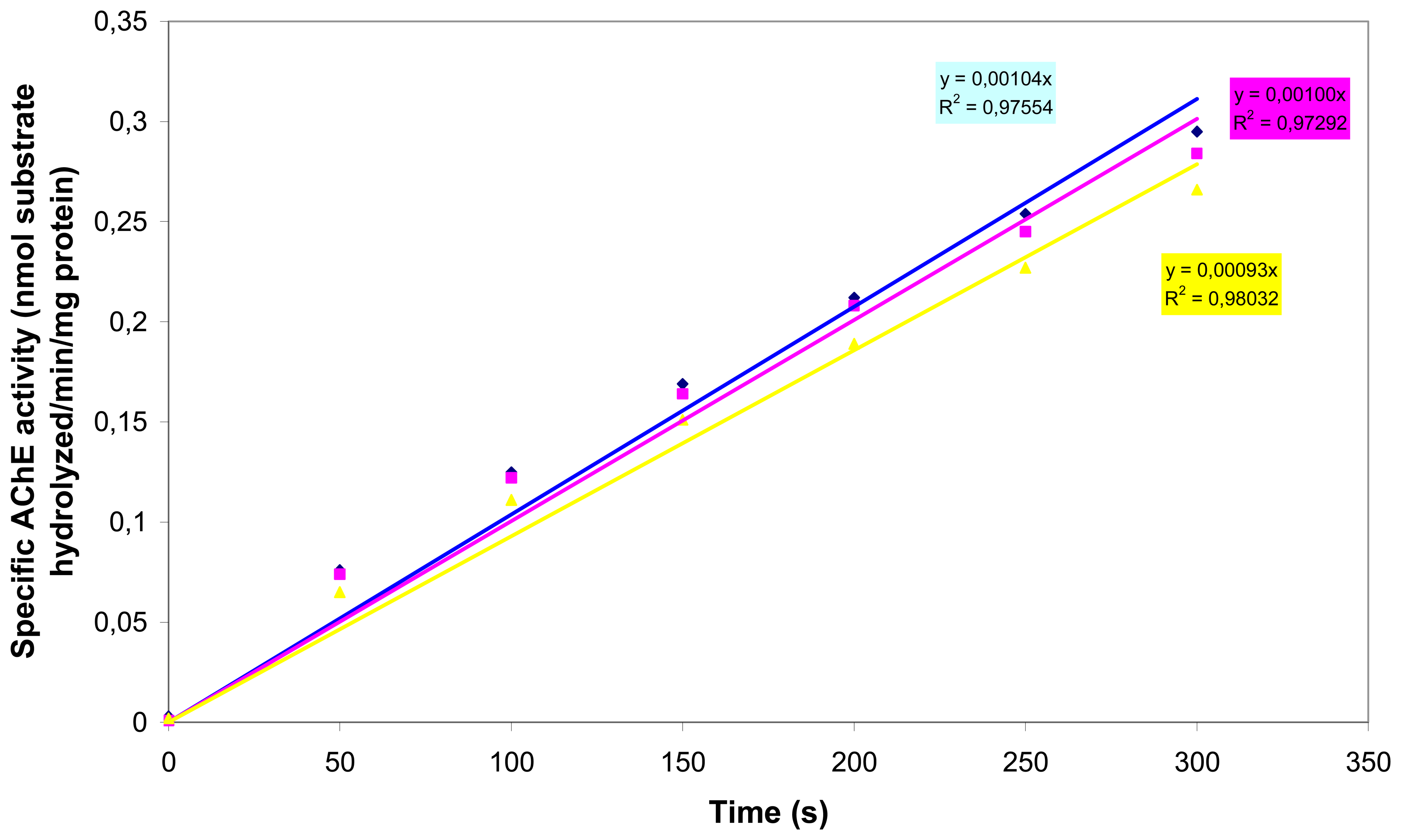
2.7. Acetylcholinesterase inhibition assay
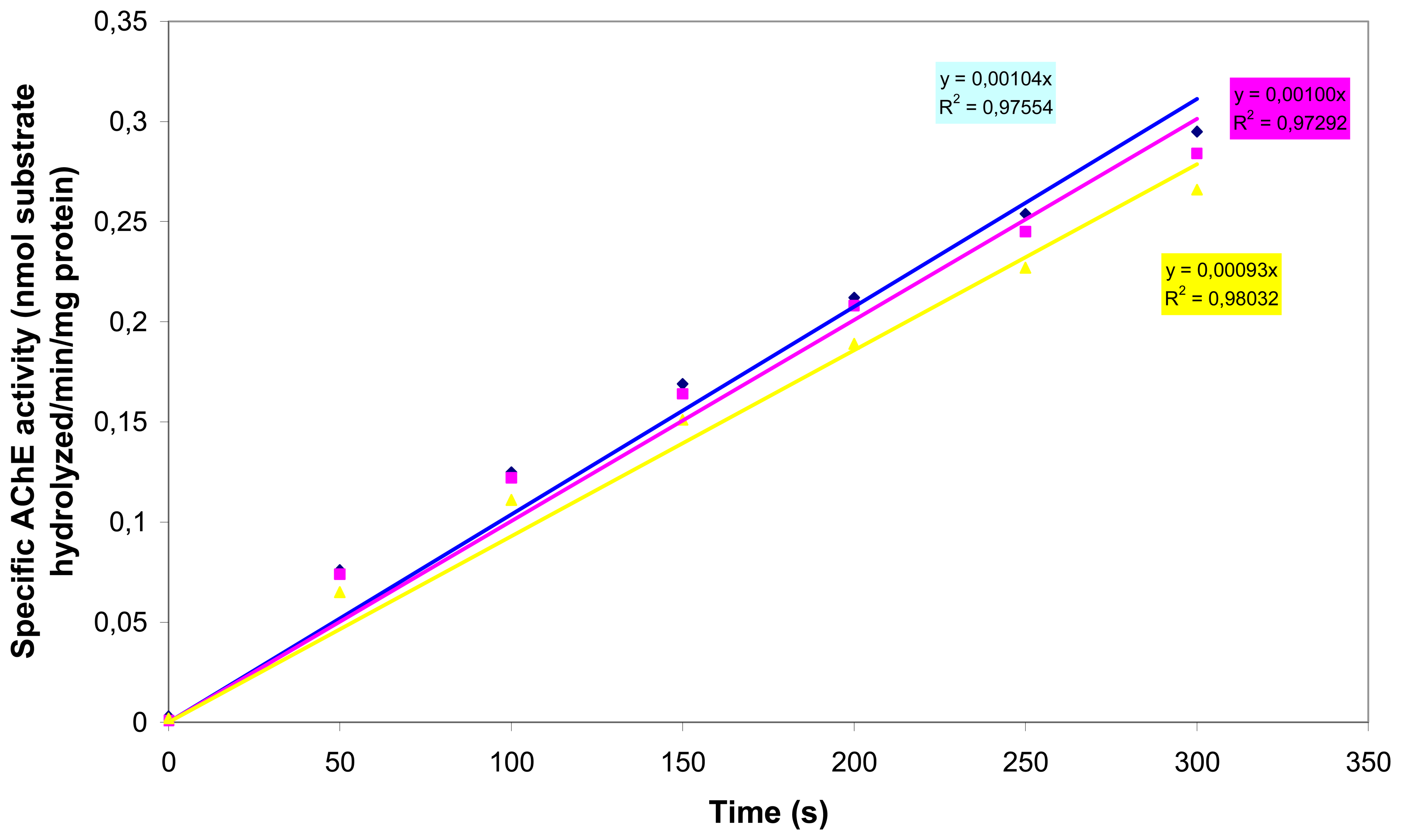
In order to demonstrate that (a) AChE is functionally expressed in N2a cells and (b) organophosphates and carbamates inhibit N2a-located AChE, we carried out AChE inhibition assays. AChE activity was determined spectrophotometrically with the Ellman AChE assay [
25], using acetylthiocholine as the substrate. To prepare the cells for the AChE assay, 1 ml of P-buffer (0.1M sodium phosphate, 1% Triton X-100, pH 7.4) was added to a 15 ml sterile Cellstar® tube containing the cell pellet (approximately 7,5 × 10
6 cells) and the mixture was slowly triturated until the pellet was resuspended. All experiments were performed in P-buffer. S-acetylthiocholine-iodide (ATCi) was used as a substrate to determine specific cholinesterase activity. ATCi hydrolysis was measured as an increase in OD
405 (50 s time intervals) caused by the reaction of thiocholine with DTNB to produce the yellow 5-thio-2-nitro-benzoic acid anion [
25]. Specific activity is reported as nmol substrate hydrolyzed/min/mg protein. The protein content was determined by the Bradford method using bovine serum albumin as the standard [
26]. The spectrophotometric assays were carried out with a spectrophotometer (UV-160A SHIMADZU) at a wavelength of 595nm.
2.8. Assay of pesticide-cell interaction
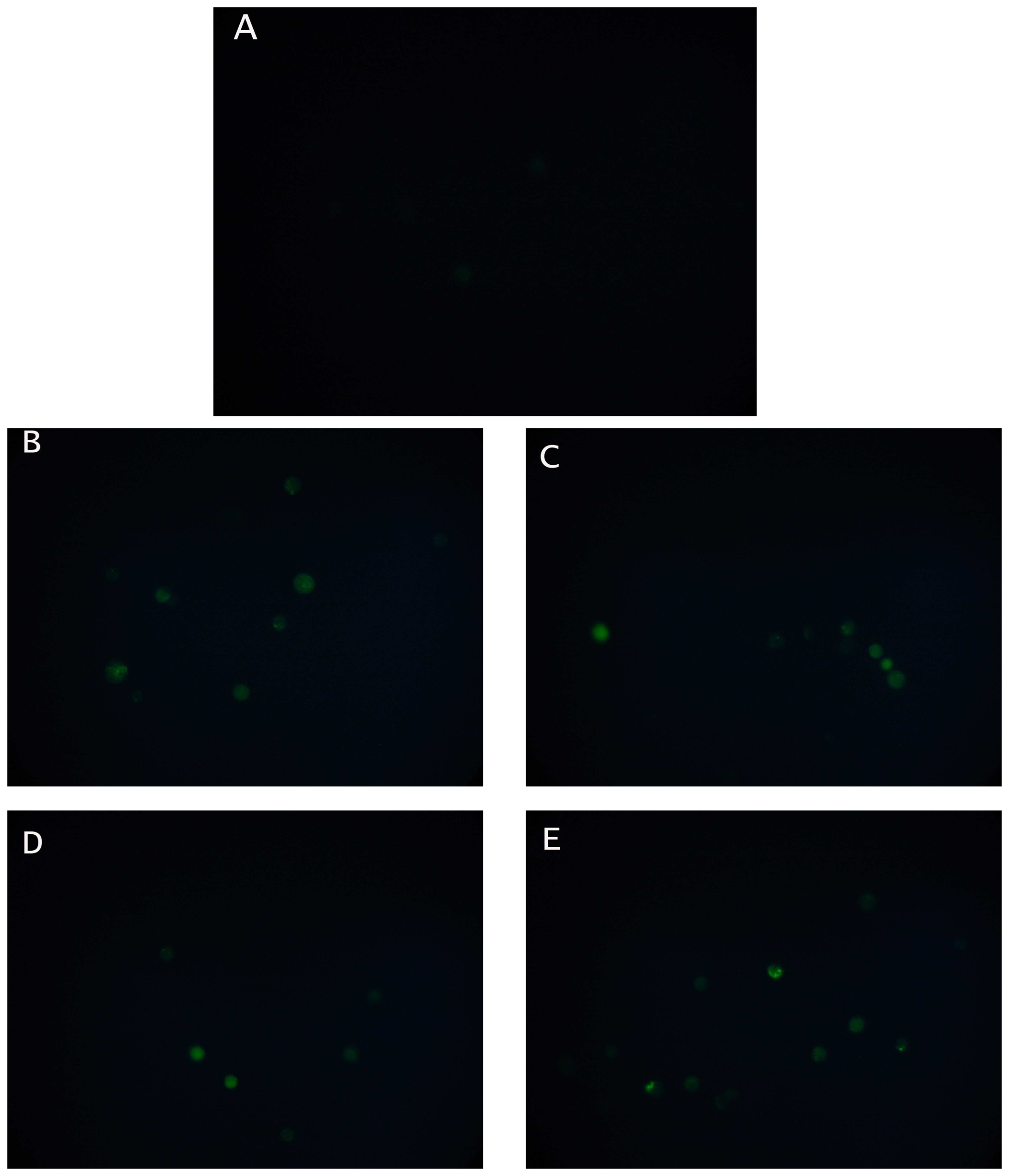
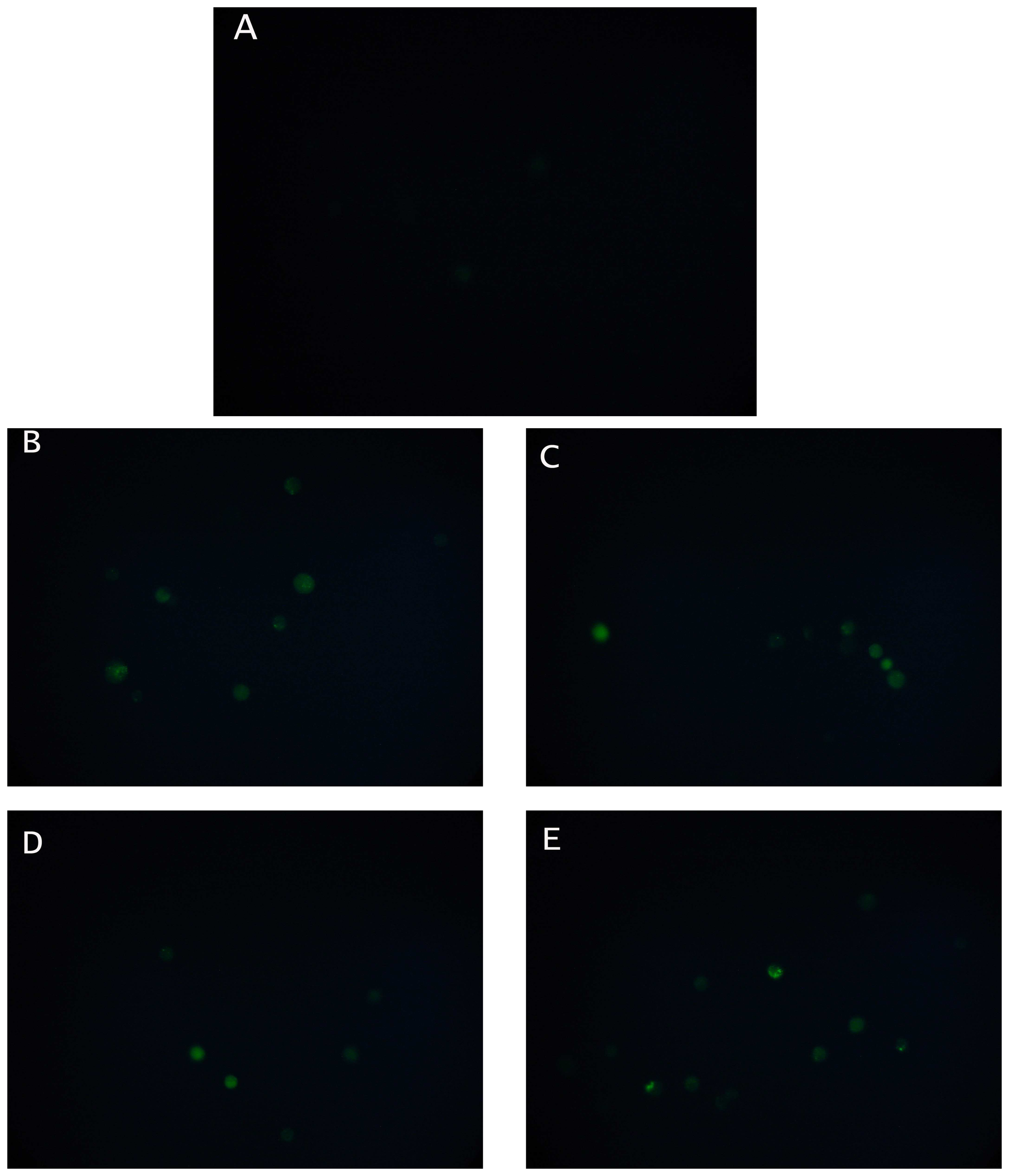
Changes in cytoplasmic Ca
2+ concentration in N2a cells before and after the addition of an organophosphate (chlorpyrifos) or a carbamate (carbaryl), each at two different concentrations (0.03 or 0.3 μM) were monitored by the uptake of the acetomethyl ester of Fluo3 [
27]. After application of five μl of the dye, the fluorescence of the specimens was recorded for five minutes at 10 s intervals. Slides with stained cells were mounted on a Zeiss Axiolab fluorescent microscope equipped with a BP-546 excitation filter and an FT-580 chromatic beam splitter. A digital camera (SONY S75 digital still camera) was attached to the microscope with adjustable BP-546/FT-580 excitation filter/chromatic beam splitter combinations. In order to control photobleaching, we kept specimen exposure times at a minimum level. No significant alteration of the intensity of the fluorescence was observed during the observation of the specimens.
2.9. Experimental design
Both biosensor and conventional sample analysis were conducted according to a double-blind protocol. Experiments were set up in a completely randomized design and each experiment was repeated three times. In each application, a set of five biosensors was tested against each individual sample. Data means among different days were compared using Duncan's multiple range test (with significance at p<0.05).
4. Discussion
The operating principle of a cellular biosensor is entirely different from conventional methods for pesticide residue detection: cell sensors respond to different molecules according to their bioactivity rather than their structure or physical chemical properties (as assayed, for instance, by mass spectrometry or chromatography) [
6,
8]. Additionally, biosensors rarely provide a full spectrum of the compounds present in a sample [
1,
5,
24]. One of the traits that make cell biosensors attractive as a clinical analytical tool is their considerable sensitivity, which is assumed to enable, in some cases, the possibility to detect just a single target molecule [
13,
14]. This theoretical assumption is based on the intricate interaction, which has emerged as a result of relentless trial-and-error testing through countless millennia and allows cells to respond to individual molecules and organisms in rather precise and reproducible ways. This ability, however, comes at a cost, or, rather, a compromise: because cells can react in roughly the same manner against an amazingly large number of different molecules, cell sensors can exhibit a very poor selectivity. This is a very common problem in toxicity assays, where cell sensors successfully detect the presence of a toxic (or genotoxic) compound, but generally fail to determine the exact nature of the toxic analyte [
10,
11,
28].
Despite this, the present biosensor system offers a number of distinct performance characteristics for pesticide residue detection, including:
Ultra-high speed: an assay can be completed in less than 3 min.
High sensitivity, with the ability to detect pesticides at concentrations as low as 1 ppb (for carbaryl).
High reproducibility.
Low cost, not exceeding 30-50 € per assay batch.
The ability to monitor at real-time conditions.
Finally, the ability to detect toxicity with complex, “real world” mixtures of toxicants and interfering compounds.
From a practical point of view, these traits could turn the novel biosensor into an attractive future technology for routine, field-based pesticide assays. Providing novel solutions for food quality assurance can be a very critical issue, especially in view of the new EU [
29] and international regulations for minimal residue concentration in marketed food and agricultural products. The development of the novel sensor concept is still at a preliminary stage: this means that further experiments are required in order to define the linear range of the response and the possible interference of different pesticides on the selectivity of the sensor. In the same context, it may be worth mentioning that, within the framework of the present study, standard solutions of pesticides for calibrating the novel biosensor were prepared by spiking residue-free tobacco, in order to avoid the so-called “matrix effect”, i.e. the interference of sample-specific components on the sensor performance. Different samples, different types of extract, different commodities and different “concentrations” of matrix may exhibit matrix effects of different magnitudes. Matrix effects are variable in occurrence and intensity and calibration should be matrix-matched routinely [
30]. For example, when we prepared pesticide standard solutions by directly dissolving carbaryl or chlorpyriphos in a solvent, rather than by spiking them in tobacco, an entirely different pattern of sensor response to either compound was observed (analytical results not shown).
According to the working hypothesis of our method, inhibition of AChE by either pesticide may have caused excessive stimulation of N2a cells by Ach, thus resulting to membrane depolarization [
22]. The considerably increased concentration of intracellular Ca
2+ stores after treating neuroblastoma cells with the pesticides provides an indication that this hypothesis may be true, although it is yet uncertain whether calcium ions are the sole contributors to the observed depolarization effect. In various cell types (including neural and cancer cells) membrane polarization events are associated with other stress-induced changes, such as an increase in cytoplasmic free calcium [
31]. Whelan and Zare [
27] have previously shown that receptor-like interactions between molecules on the cell surface and target analytes resulted in a detectable change in the concentration of cytosolic Ca
2+, while an association between conformational changes of electroinserted SOD molecules, calcium ion efflux and cell membrane potential has been demonstrated by Moschopoulou and Kintzios [
32]. In this way, cells with a particular type of surface-bound receptor serve as recognition platforms for homologous target molecules, whereas the translation of the recognition reaction into potential changes makes the assay much more rapid (and, in some cases, more sensitive) than any conventional methods.
Further increase in the sensor sensitivity could arise by increasing the concentration of immobilized cells per sensor, whereas the selectivity of the sensor's response against different pesticides could be increased by developing genetically engineered cell lines expressing different AChE isoforms on their surface. In addition, the utilization of artificial neural network technology [
33] for rapid, automated pattern recognition of the sensor's response could transform the method into a powerful, high-throughput analytical tool. These approaches are being currently tested by our research group.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}