1. Introduction
Reconstitution of lipid membranes onto inorganic or metallic substrates in a biochip approach has been extensively studied during the last decade [
1;
2]. Especially in biosensor development, investigations on bio-molecular recognitions, interactions or captures in a biomimetic environment can be an advantage in comparison with others functionalization processes. This has been recently pointed out in many biochip approaches involving peptide or protein chips [
3-
5]. Moreover, a critical step in the making of these biochips was the development of appropriate surface immobilization protocols. In order to prevent steric hindrances and random organizations, the immobilization of ligands must be highly controlled. This major drawback was often occulted leading to not completely optimized sensors. Beyond the biomimetic point of view, lipid membranes have shown strong versatility in their composition leading to many lipid-based functionalization strategies [
6;
7]. The most common approach is based on the introduction of small fractions of modified lipids allowing chemical or biochemical immobilizations on surface. A very promising strategy, inspired by ion metallic affinity chromatography (IMAC), was developed by the Tampé and Arnold groups in 1997/1998 [
8;
9] and significantly developed since 2002 [
10-
12]. It consists of incorporating synthetic lipids bearing a nitrilotriacetic acid (NTA) or iminodiacetic acid (IDA) moieties that complex metallic divalent ions such as Cu
2+, Zn
2+ or Ni
2+. Such surfaces efficiently capture histidine-tagged proteins by coordination bonds but, contrary to covalent coupling, these bonds were characterized by reversibility properties. Thus, the regeneration was obtained completely by adding metal ion chelating agents such as EDTA or competitor compounds as free histidine or imidazole [
13].
We have previously immobilized on hybrid bilayers (HB) a unique supramolecular assembly of a redox protein with nucleic acids, called P-DNA blocks. The stability of this assembly was strengthened by reconstituting the complex below the transition phase of the lipid matrix. In this configuration, we have demonstrated that the P-DNA design offers the possibility to accurately control the density of immobilized probes on the bio-mimetic layer and optimize the DNA chip sensitivity and specificity [
10;
14]. These results have open the way for the development of a new generation of biosensors that allow analysis of the modulation of DNA–DNA and DNA–RNA interactions by a large range of chemicals or biological effectors.
In this paper, we present an original structure design based on assembly of P-DNA blocks driven by specific DNA hybridization process. The resulting supramolecular assembly is called (P-DNA)
2. (P-DNA)
2 blocks have been conceived to reconstitute a palindromic DNA response element called Estrogen Response Element (ERE) which is recognized by the estrogen receptor (ER), a member of the nuclear receptor super-family [
15]. (P-DNA)
2 blocks were extensively characterized by gel electrophoresis and spectrophotometric measurements. Mechanisms of reconstitution of (P-DNA)
2 complexes onto lipid matrices were intensely investigated by combining surface plasmon resonance (SPR) and atomic force microscopy (AFM). From our knowledge, this is the first study presenting in parallel a global analysis and nanoscale characterizations of bio-molecular building blocks on exactly the same gold substrate (commercial gold chip). These complementary investigations allowed to establish a biosensor devoted to the study of DNA-protein interactions which are illustrated herein in the case of estrogen receptors.
3. Discussion
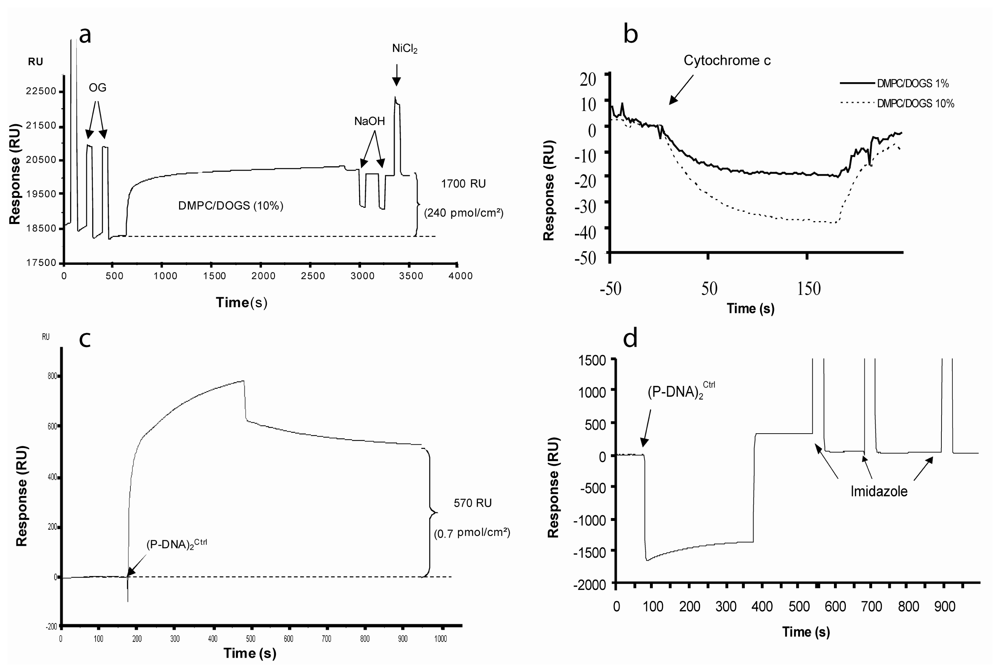
In the present study, a population of engineered protein derived from human microsomal cytochrome b5 specifically linked to an oligonucleotide with a hetero-bifunctional linker was synthesized to yield a building block called P-DNA. P-DNA was a versatile molecular block able to promote the formation of taller and highly controlled nano-objects. Many strategies can be used to obtain such supramolecular structures including (i) hybridization in solution followed by chromatographic steps or (ii) solid-phase synthesis strategy. Recently, the latter option was successfully used to construct large P-DNA blocks in a DNA network project [
16]. In our study, reconstitution in solution was convenient in regard to the final molecular assembly. Thus, P-DNA blocks were incubated in a optimized procedure with long overlapping complementary oligonucleotides to give an unique supramolecular block called (P-DNA)
2. At the end of hybridization process, excess P-DNA was easily separated by gel exclusion chromatography and complexes of interest were detected by non-denaturing electrophoresis. Specific stains revealed nucleic and protein parts of these blocks (see
supplementary result 1). Each (P-DNA)
2 block presented two tags with the –NGHHH-COOH sequence which allow the interaction with two lipid anchors (DOGS) through coordinated histidine/nickel binding. Thermodynamically, this structure is more strongly associated with the lipidic matrix than P-DNA and the mechanism of association/dissociation fits well with a kinetic model of “bivalent analyte” as shown in SPR experiments. Moreover, Stenberg's calibration, in SPR experiments, demonstrated that 1000 RU corresponds to 1 ng/mm
2 of protein [
17]. The amount of complexes covering the lipidic membrane as determined from SPR results was estimated taking into account molecular mass and area of 75 kDa and 100 nm
2 respectively for the (P-DNA)
2 blocks. Based on these theoretical dimensions, the maximum (P-DNA)
2 coverage achieved was 16.6 fmoles/mm
2 or 1250 RU. The amount of 1% of DOGS lipid represented the smaller amount of DOGS leading to complete (P-DNA)
2 immobilization. Upper limit was raised to 10% DOGS lipid content in order to magnify the amount of (P-DNA)
2 blocks immobilized. Experimental SPR data did not exceeded 800 RU following the injection of (P-DNA)
2 blocks, which corresponds to 10 fmoles/mm
2. Dissociation occurs at this level of loading and stabilization of the interactions was observed below 6 fmoles/mm
2. The phenomenon of dissociation of P-DNA has previously been demonstrated and resulted from both the rupture of the chelate-mediated link and the extraction of lipid anchors from the lipidic matrix [
14]. This major drawback was overcome by using two tags per (P-DNA)
2 block. However, (P-DNA)
2 blocks did not bind simultaneously with two DOGS. According to the availability of free DOGS in the lipid matrix, a part of them could link only one anchor domain. These “not fully bound” blocks were probably released from the surface during experiments. On the other hands, when ER sensors were established, regeneration processes using competitors of Ni
2+ coupling or Ni
2+ chelating agents were efficient as shown both in SPR and AFM investigations.
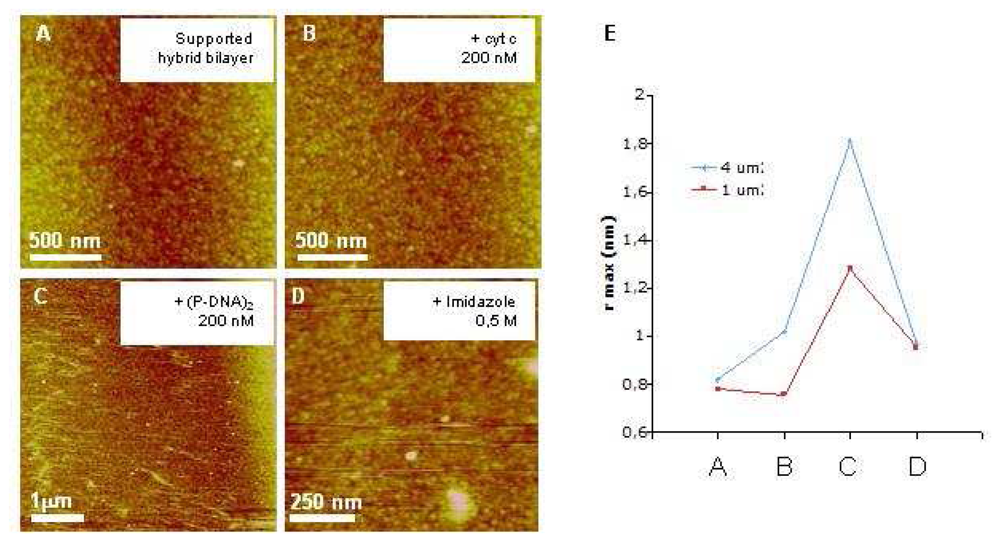
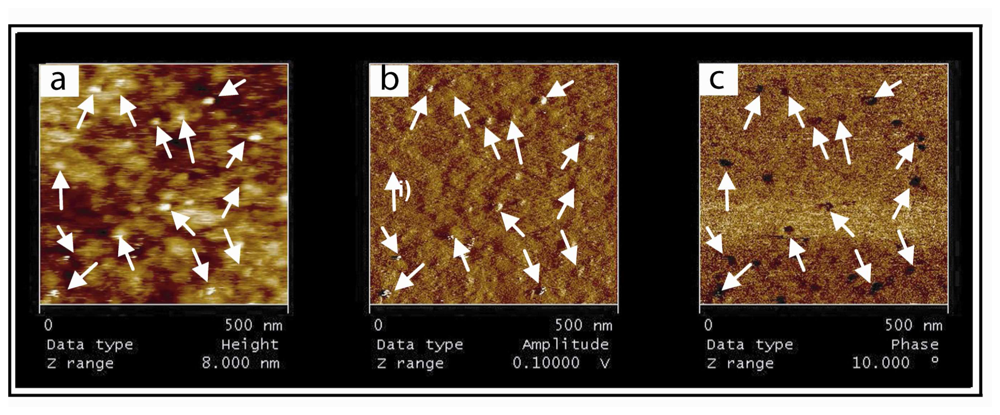
In order to better understand events that occur at the surface of the biochip, we performed AFM investigations of substrate employed for SPR experiments. The relevancy and originality of the present imaging study lies on the fact that visualization of specifically immobilized (P-DNA)
2 complexes was performed on the same commercial gold chips used for SPR characterizations. While this substrate presents a rough surface, far rougher than the flat mica usually used for AFM imaging, we managed to visualize (P-DNA)
2 complexes immobilized on a hybrid membrane and to demonstrate the specificity and reversibility of this grafting. SPR and AFM results support the model of (P-DNA)
2 complexes immobilization through histidine/nickel interactions. Indeed, in both methods, grafting of (P-DNA)
2 blocks is: i) DOGS membrane content dependent, ii) surface specific and iii) reversible. The combination of both techniques is still a challenge. Indeed, very few studies argue for the use of the AFM tool to visualize the organization of proteins on the same working surface. Indeed, in literature we can find AFM characterization study of sensors based on quartz crystal [
12] or gold electrode [
18]. But, in the first case [
12], AFM experiments were actually performed on atomically flat mica, which is a deeply different surface (flat mica versus rough gold substrates). In the other study [
18], while gold electrode presents a clear roughness (islands of 300 nm [
19]), their AFM observations were performed on atomically flat gold (Au (111)-(1×1)). Thus, it appears not evident at all to visualize by AFM molecules and their interactions “strictly on the sensor surface”. This increases the weight of our study, since we revealed (P-DNA)
2 complexes specifically and reversibly immobilized on similar lipid surface reconstituted on gold chip through both SPR and AFM methods.
Another challenge concerns the imaging of macromolecular complexes linked to a lipidic carriers in a lipid bilayer [
20]. The lateral mobility of chelating lipids allows (P-DNA)
2 complexes to move in the plane of membrane. Such a behavior makes high resolution imaging of complexes more difficult. Our AFM images, while highly demonstrative, are probably limited in terms of resolution due to this diffusion. At this stage, it is important to note that our biosensor offers through its structure (hybrid bilayer) the possibility to overcome this drawback. Decreasing the working temperature would indeed rigidify the supported membrane (phase transition between liquid and gel states still exist with few % of DOGS in DMPC hybrid bilayers as previously demonstrated [
14]), thus limiting mobility of (P-DNA)
2 blocks).
In order to realize an estrogen receptor biosensor, we have built two populations of (P-DNA)
2 blocks presenting respectively the specific palindromic DNA estrogen response element (ERE) and a control DNA (ctrl). In preliminary experiments, these populations were reconstituted on the HB at a level of 2 fmoles/mm
2 (around 150 RU). On the other side, the binding of ligand (such as E2) to the estrogen receptors induces their dimerization. This conformational change induced activation of ER and their interaction with the ERE sequence. Moreover ERα protein is known to be really difficult to store. This nuclear receptor is particularly sensitive to the denaturizing and aggregating processes [
21]. Every authors working on SPR based ER/ERE interactions studies used the same ERα origin which was commercialized at 80% of purity and could be denaturized or aggregated with time [
22-
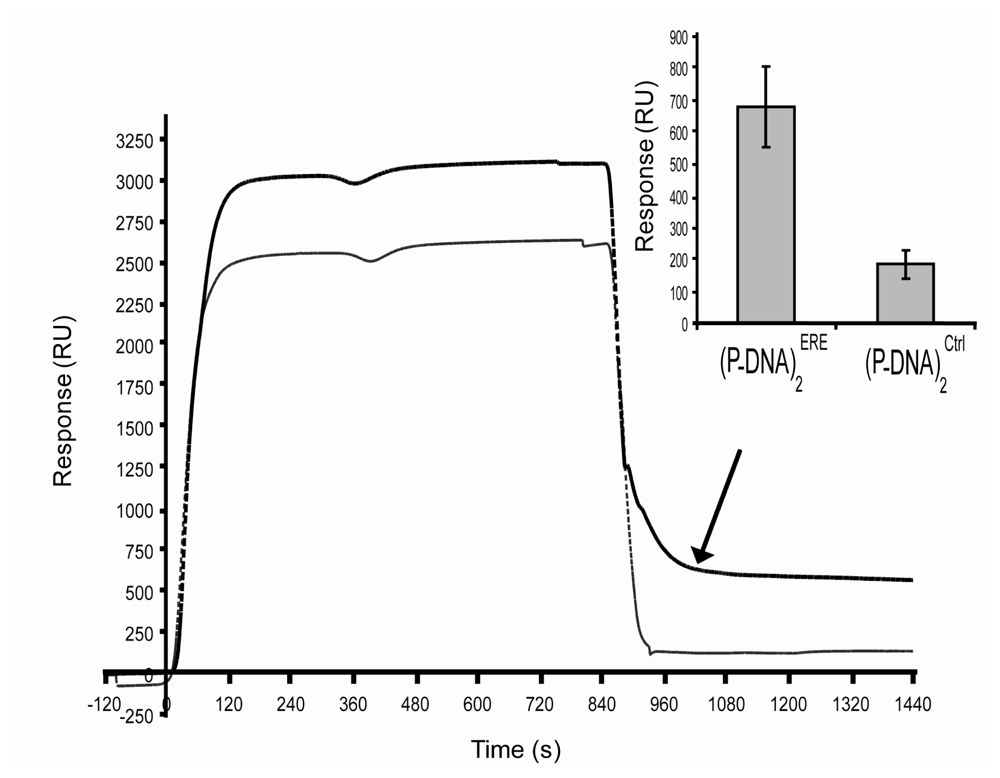
27]. In our study, the activated ER interacts with the DNA biosensor during injection steps and especially with the ERE sequence. This was illustrated in
figure 4, where few minutes after the end of injection, 10.2 fmol/mm
2 of activated ERα have strongly interacted with specific ERE target whereas unspecific linkage to control sequence reached 2.8 fmol/mm
2. Whereas, due to the large variability of the control sequences in others publications, it was difficult to compare ERα/control DNA interaction responses. The level of unspecific signal (27%) obtained with our biosensor was in the range of previous publications presented results with control sequences (between 10 to 50%) [
22;
24;
26;
28].
Moreover, a particular result of our biosensor must be pointed out. (P-DNA)
2 surface coverage has been fixed to 2 fmole/mm
2 which corresponded to 4 fmole/mm
2 of ERE immobilized on the lipidic matrix. When activated ER was injected at 50 nM during 15 min, the specific signal of ER/ERE interactions reached a plateau at 490 RU, i.e. around 0.49 ng/mm
2 or 7.4 fmoles/mm
2. In these conditions the efficiency of RE/ERE biorecognitions was up to 90%. These first results on protein/DNA interactions based on this biomolecular architecture seem to be promising and will be further investigated. Finally regeneration of RE/ERE biosensors were usually based on dsDNA stripping or denaturizing protein processes [
23;
25]. Our biochip offers an alternative to these methods which allow overcoming the unspecific adsorption on DNA. (P-DNA)
2 blocks linked to ER by affinity anchorage were simply eliminated by imidazole pulses, offering a lipidic matrix available for a new DNA grafting. These preliminary results allowed us to conclude that ERα interaction is mainly ERE specific, which confers biological relevancy to our biosensor.
4. Experimental Section
4.1. Materials
LC-SPDP (Pierce Biotechnology, Rockford, USA) was used to link oligonucleotides to modified cytochrome b5 reduced by 1.4-DiThioTreitol (DTT) (Sigma Saint Louis, USA). Oligonucleotides (Eurogentec, Liege, Belgium) were able to create ERE or double strand control sequence (Ctrl) by complementary hybridizations (A1/A3 and A4/A6). P-DNA structures were purified by chromatography using DiEthylAminoEthyl (DEAE), IminoDiacetic Acid (IDA) and Sephadex G75 gels purchased from Sigma. Lipid surfaces were constituted by a mixture of DiMyristoyl-Phosphatidyl-Choline (DMPC) and 1.2-DiOleoyl-sn-Glycero-3[(N(5-amino-1carboxypenty) iminodiacetic acid)] Succinyl (DOGS) (Aventi Polar Lipids, Albaster, USA). Human recombinant Estrogen Receptor-α (ERα) (PanVera, Invitrogen Corporation, Carlsbad, USA) was conserved at -80°C into 10 μl aliquots to limit the number of freeze-thaw cycles. Estradiol-17-β (E2) (Sigma, St Louis, USA) was prepared at 1 μM in ethanol and stored at -20°C.
4.2. Supramolecular building
A1 (5′-AGTTCTTTGATCAGGTCACTGTGACCTGAACTTGCT-3′) (ε
260nm = 334.400 M
-1 .cm
-1) or A4 (5′-AGTTCTTTGATACGTCCCATCAAGTCAGACTTGCT-3′) (ε
260nm = 335.900 M
-1 .cm
-1) oligonucleotides were coupled to LC-SPDP by incubation for 17 hours in 50 mM phosphate buffer pH 7.5 (called PB) at room temperature with 1/25 molecular ratio. Excess LC-SPDP was eliminated by ion exchange chromatography (DEAE) in PB buffer. Oligonucleotides linked to the column were eluted with 1 M NaCl in PB buffer. Coupling efficiency was evaluated in reducing conditions (20 mM DTT). This reducing agent cleaves LC-SPDP, releasing thiopyridine which was quantified by spectrophotometry at 343 nm. The genetically engineered cytochrome b5 was previously described [
29]. Briefly, a S24C mutation has been introduced, by directed engineering, to enable protein / linker coupling. Then, a tag of six amino acids (NGHHH) was added at the C-terminus to allow IMAC chromatography processes and grafting on DOGS.
DNA/LC-SPDP complexes were coupled to modified cytochrome b5 (ε412nm = 117.000 M-1 .cm-1) through sulfhydril residue bearing by the unique cystein at position 24. This cystein was reduced by a 10-minute incubation at room temperature in DTT excess (1/10 mole/mole), which was eliminated by exclusion chromatography (Sephadex G25). DNA/LC-SPDP complexes were incubated with modified cytochrome b5 (molecular ratio 1/1) at room temperature overnight, after which DNA/LC-SPDP/b5 complexes (called P-DNA) were purified in several steps. First, complexes without cytochrome b5 were eliminated by affinity chromatography with an iminodiacetic acid column loaded using a NiCl2 solution (0.1 M acetate buffer pH 7.8). All cytochrome b5, bearing DNA or not, were eluted by 1 mg/ml histidine solution (PB buffer). Then P-DNA complexes were purified by ions exchange chromatography. Free cytochrome b5 were eliminated by 0.25 M NaCl (PB buffer) and P-DNA were eluted in 1 M NaCl (PB buffer). Complexes were quantified by spectrophotometric measurements at 260 and 412 nm.
A dimerization process was based on the complementary hybridization properties of DNA that lead to a structure called the (P-DNA)2 block. A3 (5′-AGCAAGTTCAGGTCACAGTGACCTGATCAAAGAATATATAGCAAGTTCAGGTCACAGTGACCTGATCAAAGA-3′) (ε260nm = 735.700 M-1 .cm-1) or A6 (5′-AGCAAGTCGTGACTTGATGGGACGTATCAAAGAATATATAGCAAGTCGTGACTTGATGGGACGTATCAAAGA-3′) (ε260nm = 739.900 M-1 .cm-1) oligonucleotides, which presented two complementary domains with respectively A1 and A4 sequences, were able to link two P-DNA complexes. Two molecular ratios between A3 or A6 and corresponding complexes were used to evaluate the most efficient dimerization process. One mole ssDNA with 3 moles of P-DNA at 4°C in PB buffer overnight. Theoretically hybridization process can lead to the building of three species: i) complexes (A1/LC-SPDP/b5)2-A3 or (A4/LC-SPDP/b5)2-A6 called respectively (P-DNA)2ERE and (P-DNA)2Ctrl, ii) (A1/LC-SPDP/b5)-A3 or (A4/LC-SPDP/b5)-A6 (with only one b5) called respectively P-(DNA)2ERE and P-(DNA)2Ctrl and iii) A1/LC-SPDP/b5 or A4/LC-SPDP/b5 called respectively P-DNAERE and P-DNACtrl. These different species were separated by gel filtration (Sephadex G75, 0.1 M Phosphate Buffer Saline (PBS)). Spectrophotometric study was used to identify these different species by determining the characteristic A260/A412 ratio.
4.3. Assembling of complexes onto the chip
First, commercial gold chips SIA (GE Healthcare Life Sciences, Pittsburgh, USA) were chemically functionalized in 1 mM OM (Sigma, Saint Louis, USA) at room temperature overnight as previously published [
9]. Then, 1 mM (DMPC)/(DOGS) SUVs were prepared by extrusion using a 50 nm polycarbonate membrane in PB buffer. Several DOGS/DMPC ratios were used in this study, from 0.01 to 0.1 range mole/mole, in order to modulate (P-DNA)
2 surface density. After wetting the hydrophobic SAM with 50% ethanol, the surface was washed with 40 mM OG. Lipid vesicles spread spontaneously onto the hydrophobic surface at 25°C. Excess lipid was removed by treatment with 20 mM NaOH leading to a stable baseline. Injections of (P-DNA)
2 onto this lipidic surface was performed at 5 μl/min.
4.4. REα/ DNA interaction
Interaction study was performed after 50 nM ERα dimerization in the presence of 1 nM E2 (4 hours incubation in PBS at 4°C). Then, receptor solution was simultaneously injected onto both (P-DNA)2Ctrl and (P-DNA)2ERE channels at 2 μl/min for 15 minutes.
4.5. SPR experiments
SPR experiments were run on BIAcore 2000 (GE Healthcare Life Sciences, Pittsburgh, USA) at 25 °C, with a flow rate of 5–50 ul/min, in PB buffer or in PBS for (P-DNA)2 graftings and ERα interaction experiments.
4.6. Spectrophotometric characterizations
Spectrophotometric study with a λ900 spectrophotometer (PerkinElmer Instrument, Waltham, USA) was performed to identify P-DNA species. For calculations, absorbance at 260 nm had to be corrected to take into account the contribution of cytochrome b5 at this wavelength (about 17% of the 413 nm absorbance value). Molar extinction coefficient at 260 nm of DNA component in (P-DNA)
2ERE complexes, for example, was
, so global coefficient at 260 nm for (P- DNA)
2ERE was
ε (P - DNA)2ERE260nm =
ε(A1)2 - A3 260nm + 2×
εb5260nm. For P-(DNA)
2ERE molar extinction coefficient at 260 nm of DNA part was
, so global coefficient of P-(DNA)
2ERE was
εP − (DNA)2ERE260nm =
εA1 − A3260nm +
εb5260nm. Molar extinction coefficient of P-DNA
ERE was
εP − DNAERE260nm =
εA1260nm +
εb5260nm. The coefficient at 412 nm was only affected by number of cytochrome b5 molecules. Each species present a characteristic molar extinction coefficient ratio. Thus, all complexes could be discriminated by determining the A
260 nm/A
412 nm ratio (
Table 1).
4.7. AFM characterizations
The AFM used was a Nanoscope III (Veeco, Santa Barbara, CA). Imaging was performed in contact and oscillating contact mode (tapping™ mode) using NPS-oxide sharpened silicon nitride probes (Veeco) exhibiting spring constants of 0.32 N/m or 0.58 N/m at resonance frequencies of 8.5 to 9.5 kHz. For the feedback controls, typical values of set-point for imaging were between 0.5 to 1.5 V, depending on scan size and drive amplitude in oscillating contact mode. The oscillation amplitude was generally maintained at 5-10 nm away from the surface.






{kind=link}
{kind=link}
{kind=link}
{kind=link}



