Coronatine Gene Expression In Vitro and In Planta, and Protein Accumulation During Temperature Downshift in Pseudomonas syringae

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial strains, plasmids, and growth conditions
2.2. Plant material and inoculation procedures
2.3. Confocal laser scanning microscopy
2.4. RNA isolation and spot blot analysis
2.5. Immunodetection of CmaB accumulation
3. Results and Discussion
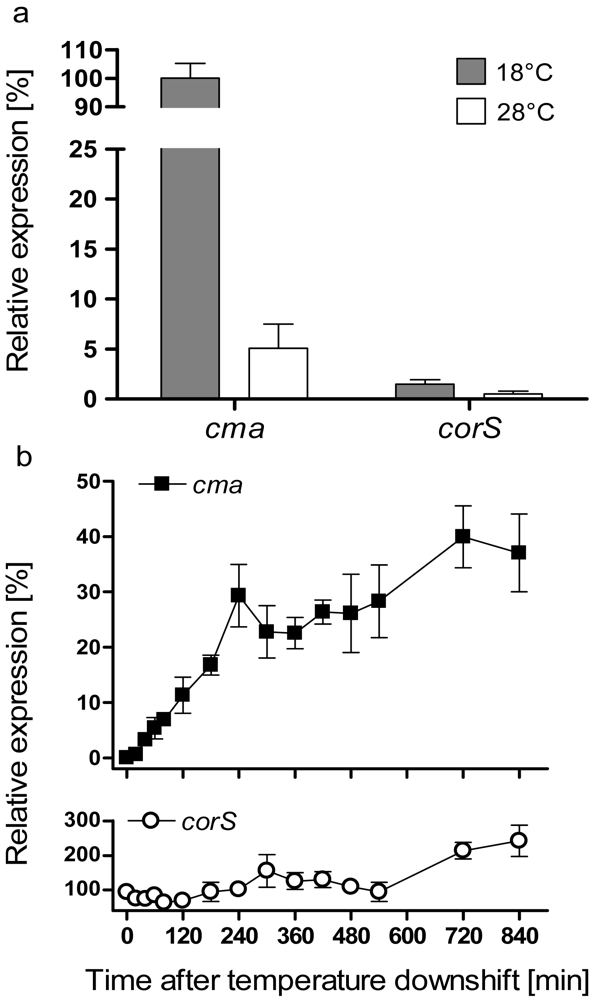
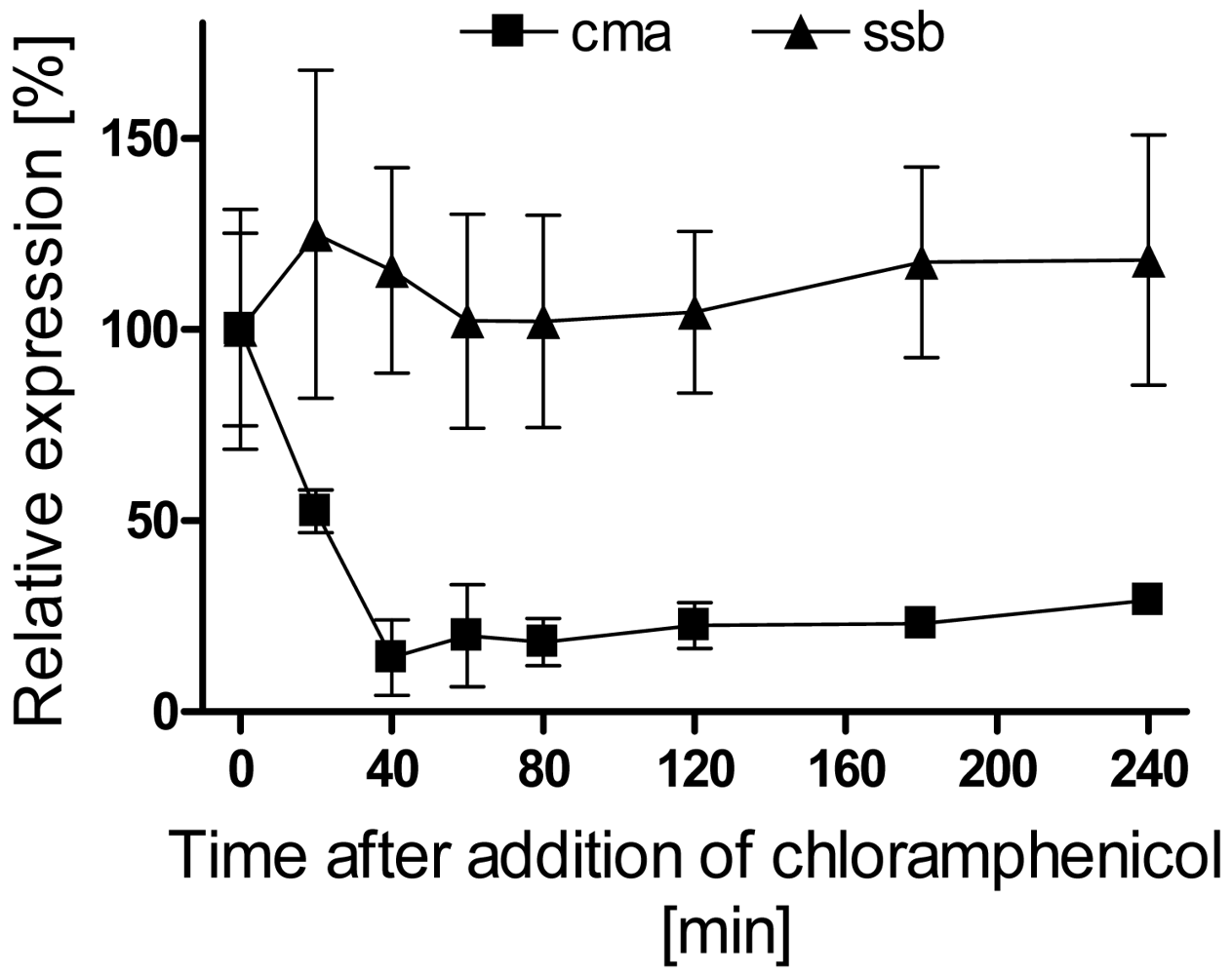
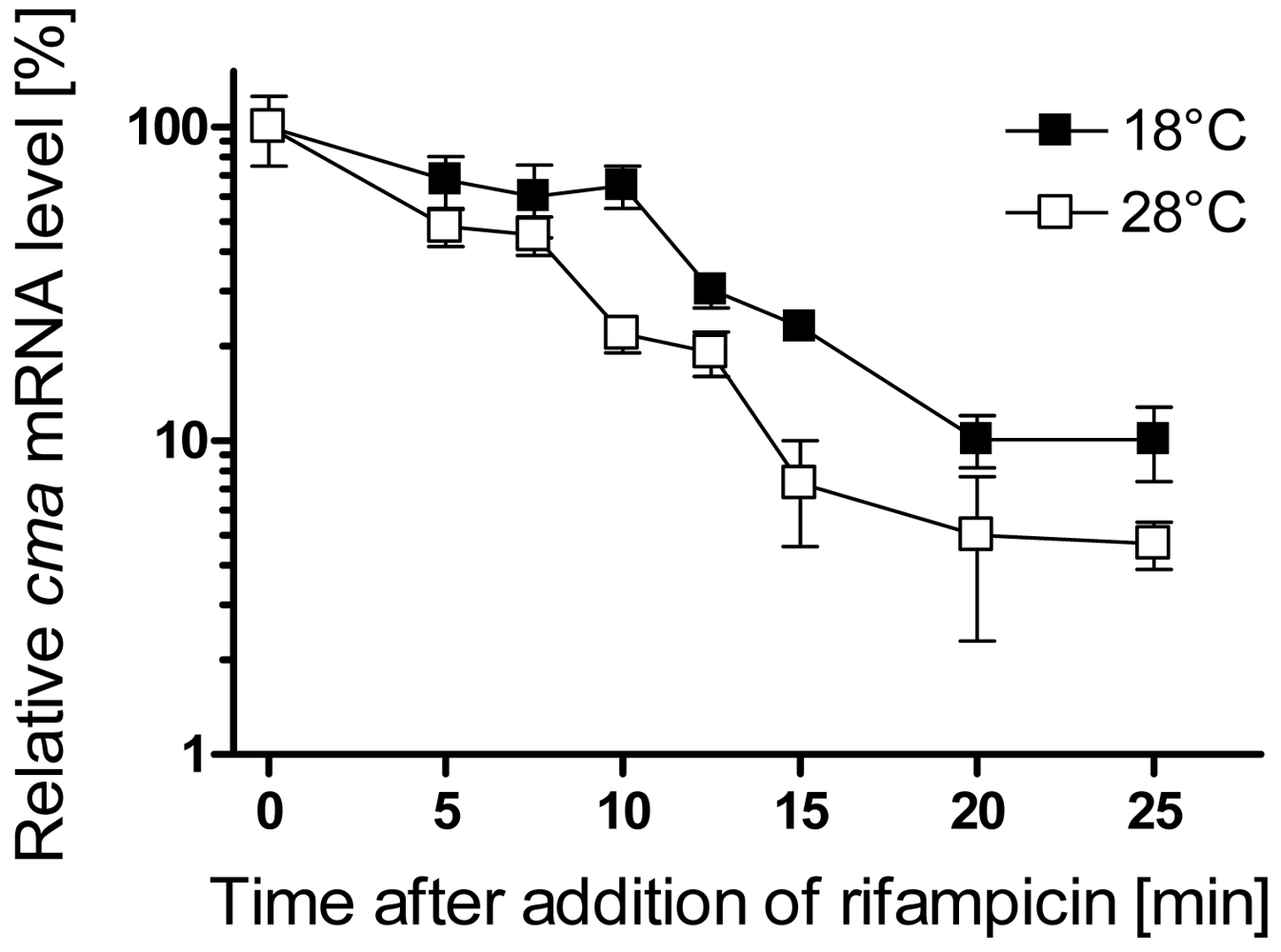
3.1. Analysis of cma and corS mRNA levels upon a temperature shift from 28 to 18 °C
3.2. Analysis of CmaB protein levels following a temperature shift from 28 to 18 °C
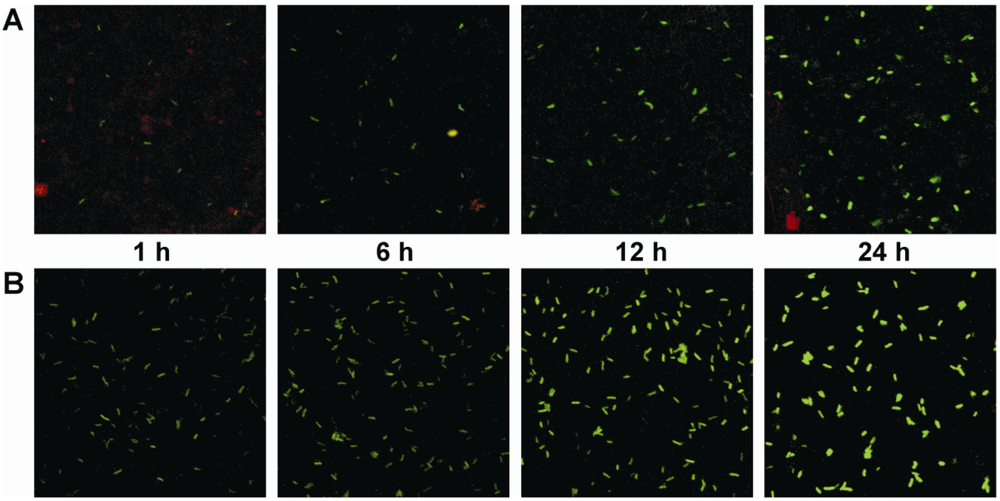
3.3. Analysis of cma promoter activity in planta following a temperature shift from 28 to 18 °C
4. Conclusions
Acknowledgments
References
- Eriksson, S.; Hurme, R.; Rhen, M. Low-temperature sensors in bacteria. Phil. Trans. R. Soc. Lond. B 2002, 357, 887–893. [Google Scholar]
- Hugouvieux-Cotte-Pattat, N.; Dominguez, H.; Robert-Baudouy, J. Environmental conditions affect transcription of the pectinase genes of Erwinia chrysanthemi 3937. J. Bacteriol. 1992, 174, 7807–7818. [Google Scholar]
- Jin, S.; Song, Y.N.; Deng, W.Y.; Gordon, M.P.; Nester, E.W. The regulatory VirA protein of Agrobacterium tumefaciens does not function at elevated temperatures. J. Bacteriol. 1993, 175, 6830–6835. [Google Scholar]
- Palmer, D.A.; Bender, C.L. Effects of environmental and nutritional factors on production of the polyketide phytotoxin coronatine by Pseudomonas syringae pv. glycinea. Appl. Environ. Microbiol 1993, 59, 1619–1626. [Google Scholar]
- Ullrich, M.; Peñaloza-Vázquez, A.; Bailey, A.M.; Bender, C.L. A modified two-component regulatory system is involved in temperature-dependent biosynthesis of the Pseudomonas syringae phytotoxin coronatine. J. Bacteriol. 1995, 177, 6160–6169. [Google Scholar]
- Liyanage, H.; Palmer, D.A.; Ullrich, M.; Bender, C.L. Characterization and transcriptional analysis of the gene cluster for coronafacic acid, the polyketide component of the phytotoxin coronatine. Appl. Environ. Microbiol. 1995, 61, 3843–3848. [Google Scholar]
- Ullrich, M.; Bender, C.L. The biosynthetic gene cluster for coronamic acid, an ethylcyclopropyl amino acid, contains genes homologous to amino acid-activating enzymes and thioesterases. J. Bacteriol. 1994, 176, 7574–7586. [Google Scholar]
- Budde, I.P.; Rohde, B.H.; Bender, C.L.; Ullrich, M.S. Growth phase and temperature influence promoter activity, transcript abundance, and protein stability during biosynthesis of the Pseudomonas syringae phytotoxin coronatine. J. Bacteriol. 1998, 180, 1360–1367. [Google Scholar]
- Couch, R.; O'Connor, S.E.; Seidle, H.; Walsh, C.T.; Parry, R. Characterization of CmaA, an adenylation-thiolation didomain enzyme involved in the biosynthesis of coronatine. J. Bacteriol. 2004, 186, 35–42. [Google Scholar]
- Kelly, W.L.; Boyne, M.T., II; Yeh, E.; Vosburg, D.A.; Galonic, D.P.; Kelleher, N.L.; Walsh, C.T. Characterization of the aminocarboxycyclopropane-forming enzyme CmaC. Biochemistry 2007, 46, 359–368. [Google Scholar]
- Rangaswamy, V.; Jiralerspong, S.; Parry, R.; Bender, C.L. Biosynthesis of the Pseudomonas polyketide coronafacic acid requires monofunctional and multifunctional polyketide synthase proteins. Proc. Natl. Acad. Sci. USA 1998, 95, 15469–15474. [Google Scholar]
- Seidle, H.; Rangaswamy, V.; Couch, R.; Bender, C.L.; Parry, R.J. Characterization of Cfa1, a monofunctional acyl carrier protein involved in the biosynthesis of the phytotoxin coronatine. J. Bacteriol. 2004, 186, 2499–2503. [Google Scholar]
- Rangaswamy, V. ; Bender C.L. Phosphorylation of CorS and CorR, regulatory proteins that modulate production of the phytotoxin coronatine in Pseudomonas syringae. FEMS Microbiol. Lett. 2000, 193, 13–18. [Google Scholar]
- Peñaloza-Vázquez, A.; Bender, C.L. Characterization of CorR, a transcriptional activator which is required for biosynthesis of the phytotoxin coronatine. J. Bacteriol. 1998, 180, 6252–6259. [Google Scholar]
- Wang, L.; Bender, C.L.; Ullrich, M.S. The transcriptional activator CorR is involved in biosynthesis of the phytotoxin coronatine and binds to the cmaABT promoter region in a temperature-dependent manner. Mol. Gen. Genet. 1999, 262, 250–260. [Google Scholar]
- Alarcón-Chaidez, F.J.; Keith, L.; Zhao, Y.; Bender, C.L. RpoN (σ54) is required for plasmid-encoded coronatine biosynthesis in Pseudomonas syringae. Plasmid 2003, 49, 106–117. [Google Scholar]
- Smirnova, A.V.; Ullrich, M.S. Topological and deletion analysis of CorS, a Pseudomonas syringae sensor kinase. Microbiology 2004, 150, 2715–2726. [Google Scholar]
- Shu, C.J.; Ulrich, L.E.; Zhulin, I.B. The NIT domain: a predicted nitrate-responsive module in bacterial sensory receptors. Trends Biochem. Sci. 2003, 28, 121–124. [Google Scholar]
- Gerharz, T.; Reinelt, S.; Kaspar, S.; Scapozza, L.; Bott, M. Identification of basic amino acid residues important for citrate binding by the periplasmic receptor domain of the sensor kinase CitA. Biochemistry 2003, 42, 5917–5924. [Google Scholar]
- Bender, C.L.; Liyanage, H.; Palmer, D.; Ullrich, M.; Young, S.; Mitchell, R. Characterization of the genes controlling the biosynthesis of the polyketide phytotoxin coronatine including conjugation between coronafacic and coronamic acid. Gene 1993, 133, 31–38. [Google Scholar]
- Keane, P.J.; Kerr, A.; New, P.B. Crown gall of stone fruit. 2. Identification and nomenclature of Agrobacterium isolates. Aust. J. Biol. Sci. 1970, 23, 585–595. [Google Scholar]
- Weingart, H.; Stubner, S.; Schenk, A.; Ullrich, M.S. Impact of temperature on in planta expression of genes involved in synthesis of the Pseudomonas syringae phytotoxin coronatine. Mol. Plant-Microbe Interact. 2004, 17, 1095–1102. [Google Scholar]
- Kovach, M.E.; Phillips, R.W.; Elzer, P.H.; Roop, R.M.; Peterson, K.M. pBBR1MCS: a broad-host-range cloning vector. BioTechniques 1994, 16, 800–802. [Google Scholar]
- Schenk, A.; Weingart, H.; Ullrich, M.S. Extraction of high-quality bacterial RNA from infected leaf tissue for bacterial in planta gene expression analysis by multiplexed fluorescent Northern hybridization. Mol. Plant Pathol. 2008, 9, 227–235. [Google Scholar]
- Herrin, D.L.; Schmidt, G.W. Rapid, reversible staining of northern blots prior to hybridization. BioTechniques 1988, 6, 196–200. [Google Scholar]
- Bradford, M.M. Rapid and sensitive method for quantitation of microgram quantities of protein utilizing principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar]
- Dunleavy, J.M. Bacterial, fungal, and viral diseases affecting soybean leaves. In Soybean Diseases of the North Central Region; Wylie, T.D., Scott, D.H., Eds.; American Phytopathological Society: St. Paul, MN, USA, 1988; pp. 40–46. [Google Scholar]
- Smirnova, A.; Li, H.; Weingart, H.; Aufhammer, S.; Burse, A.; Finis, K.; Schenk, A.; Ullrich, M. S. Thermoregulated expression of virulence factors in plant-associated bacteria. Arch. Microbiol. 2001, 176, 393–399. [Google Scholar]
- Graumann, P.; Marahiel, M.A. Some like it cold: response of microorganisms to cold shock. Arch. Microbiol. 1996, 166, 293–300. [Google Scholar]
- Aguilar, P.S.; Lopez, P.; de Mendoza, D. Transcriptional control of the low-temperature-inducible des gene, encoding the Δ5 desaturase of Bacillus subtilis. J. Bacteriol. 1999, 181, 7028–7033. [Google Scholar]
- Los, D.A.; Ray, M.K.; Murata, N. Differences in the control of the temperature-dependent expression of four genes for desaturases in Synechocystis sp. PCC 6803. Mol. Microbiol. 1997, 25, 1167–1175. [Google Scholar]
- Thieringer, H.A.; Jones, P.G.; Inouye, M. Cold shock and adaptation. BioEssays 1998, 20, 49–57. [Google Scholar]
- Ullrich, M.S.; Schergaut, M.; Boch, J.; Ullrich, B. Temperature-responsive genetic loci in the plant pathogen Pseudomonas syringae pv. glycinea. Microbiology 2000, 146, 2457–2468. [Google Scholar]
- Andersen, J.B.; Sternberg, C.; Poulsen, L.K.; Bjørn, S.P.; Givskov, M.; Molin, S. New unstable variants of green fluorescent protein for studies of transient gene expression in bacteria. Appl. Environ. Microbiol. 1998, 64, 2240–2246. [Google Scholar]
- Smirnova, A.V.; Wang, L.; Rohde, B.; Budde, I.; Weingart, H.; Ullrich, M.S. Control of temperature-responsive synthesis of the phytotoxin coronatine in Pseudomonas syringae by the unconventional two-component system CorRPS. J. Mol. Microbiol. Biotechnol. 2002, 4, 191–196. [Google Scholar]
- Vaillancourt, F.H.; Yeh, E.; Vosburg, D.A.; O'Connor, S.E.; Walsh, C.T. Cryptic chlorination by a non-haem iron enzyme during cyclopropyl amino acid biosynthesis. Nature 2005, 436, 1191–1194. [Google Scholar]





© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Braun, Y.; Smirnova, A.V.; Weingart, H.; Schenk, A.; Ullrich, M.S. Coronatine Gene Expression In Vitro and In Planta, and Protein Accumulation During Temperature Downshift in Pseudomonas syringae . Sensors 2009, 9, 4272-4285. https://doi.org/10.3390/s90604272
Braun Y, Smirnova AV, Weingart H, Schenk A, Ullrich MS. Coronatine Gene Expression In Vitro and In Planta, and Protein Accumulation During Temperature Downshift in Pseudomonas syringae . Sensors. 2009; 9(6):4272-4285. https://doi.org/10.3390/s90604272
Chicago/Turabian StyleBraun, Yvonne, Angela V. Smirnova, Helge Weingart, Alexander Schenk, and Matthias S. Ullrich. 2009. "Coronatine Gene Expression In Vitro and In Planta, and Protein Accumulation During Temperature Downshift in Pseudomonas syringae " Sensors 9, no. 6: 4272-4285. https://doi.org/10.3390/s90604272