Role of 5-HT3 Receptors in the Antidepressant Response

Abstract
: Serotonin (5-HT)3 receptors are the only ligand-gated ion channel of the 5-HT receptors family. They are present both in the peripheral and central nervous system and are localized in several areas involved in mood regulation (e.g., hippocampus or prefrontal cortex). Moreover, they are involved in regulation of neurotransmitter systems implicated in the pathophysiology of major depression (e.g., dopamine or GABA). Clinical and preclinical studies have suggested that 5-HT3 receptors may be a relevant target in the treatment of affective disorders. 5-HT3 receptor agonists seem to counteract the effects of antidepressants in non-clinical models, whereas 5-HT3 receptor antagonists, such as ondansetron, present antidepressant-like activities. In addition, several antidepressants, such as mirtazapine, also target 5-HT3 receptors. In this review, we will report major advances in the research of 5-HT3 receptor's roles in neuropsychiatric disorders, with special emphasis on mood and anxiety disorders.1. Introduction
Major depression is one of the most frequent psychiatric disorders. In the United States, the lifetime prevalence of this disease is about 16% [1]. It is an important public health problem since major depression induces disability, poor quality of life, economic burden or suicide. According to the monoaminergic theory of depression, deficiencies or imbalances in monoamine neurotransmitters, i.e., serotonin (5-HT), noradrenaline (NA) and dopamine (DA), are involved in the pathophysiology of this disease. Development of antidepressants in the last five decades has been mainly based on this hypothesis. In addition, it is now well established that both pathophysiology of depression and effect of antidepressant treatments involve neuroplasticity (e.g., hippocampal neurogenesis, expression of the brain-derived neurotrophic factors (BDNF)) and hypothalamic pituitary-adrenal (HPA) axis modulation [2,3]. Nevertheless, despite their large range, pharmacotherapy treatments of depression remain unsatisfactory [4]. Firstly, patients often receive several antidepressant agents before responding to a treatment and only about 65% experience some degree of therapeutic response [5]. Secondly, treatment for weeks or months is necessary before a therapeutic response is achieved; this therapeutic delay is critical since it can be associated with increased risk of suicide [6].
New pharmacological strategies have emerged to improve efficacy and reduce the time for antidepressants to act. Preclinical studies have suggested that targeting specific 5-HT receptors with selective agonist or antagonist drugs may enhance the antidepressant response and reduced its delay compared to currently used antidepressants. Both the 5-HT1A receptor antagonist pindolol and 5-HT1A receptor agonists, e.g., buspirone, have been largely investigated both in clinical and in preclinical studies in combination with antidepressants with some significant effects [7,8]. Moreover, the beneficial effect of atypical antipsychotics, such as quietapine, in combination with antidepressants in depression may be partly due to their 5-HT2A receptor targeting [9,10]. More recently, preclinical studies suggested that 5-HT4 receptor agonists are putative antidepressants with a fast onset of action [11,12]. Similarly, 5-HT7 receptor antagonists may improve efficacy and delay of action of classical antidepressants [13]. In this review, we will report major advances in the discovery of 5-HT3 receptor roles, with special emphasis on the potential role of 5-HT3 receptor antagonism in mood and anxiety disorders. Although their clinical use mainly concerns chemotherapy-induced emesis, several preclinical and clinical studies suggest their relevance in treatment of psychiatric disorders. After a brief description of the structure and the physiological function of 5-HT3 receptors, we will describe their brain distribution. Then, we will review the effects of 5-HT3 receptor agonists and antagonists in brain areas involved in the pathophysiology of depression. Finally we will summarize several behavioral non-clinical studies and clinical studies revealing its role in the antidepressant response.
2. Structure and Function of 5-HT3 Receptors in the Central Nervous System
2.1. Structure of 5-HT3 Receptors
The 5-HT3 receptors are the only ionotropic or ligand gated ion channel of the 5-HT receptor family [14]. They are members of the Cys-loop superfamily of ligand-gated ion channels [15] as are nicotinic acetylcholine (nAch) receptors and gamma-aminobutyric acid-type-A (GABA)A receptors. Thus, they are composed of five subunits forming a cylinder that can be crossed by cations [16]. In rats and mice, two subunits have been cloned: 5-HT3A [15] and 5-HT3B [17] receptor subunits which can be arranged in homomeric 5-HT3A receptors and heteromeric 5-HT3A/5-HT3B receptors [17]. Three others subunits have been also described in multiple mammalian species but not in rodents: 5-HT3C, 5-HT3D and 5-HT3E [18-20] which probably form only heteromeric receptors with 5-HT3A receptor subunits.
2.2. Location of the 5-HT3 Receptor in the CNS
Using different methods including autoradiography, immunohistochemistry and in situ hybridization, the 5-HT3 receptor distribution has been largely described with some differences between species. 5-HT3 receptors are expressed both in peripheral and central nervous system. In the periphery, 5-HT3 receptors are located on pre- and postganglionic neurons from autonomic nervous system and on neurons of the sensory and enteric nervous system [21]. In the central nervous system, 5-HT3 receptor density appeared low compared to others 5-HT receptors [14,22]. In all species, including humans, the most important densities of 5-HT3 receptors seems to be found in the hindbrain in particular in the dorsal motor nucleus of the vagus nerve, in the nucleus of the tractus solitaries and in the area postrema [22-32]. These brain areas are involved in the vomiting reflex explaining the relevance of 5-HT3 receptor antagonist in chemotherapy-induced emesis [14]. Significant densities of 5-HT3 receptors have also been described in the spinal cord [33,34].
Compared to the hindbrain, the density of 5-HT3 receptors in the forebrain is lower. Nevertheless, significant levels of these receptors have been found in brain areas involved in the pathophysiology of depression with densities varying across species.
In non primate mammals, 5-HT3 receptors are found in the limbic areas including the amygdala, hippocampus, nucleus accumbens and in the superficial layers of the cerebral cortex, i.e., the frontal parts (in particular cingulate, prelimbic and infralimbic areas and primary and secondary motor areas), entorhinal and temporal cortex [25,27-29,31,33,35-38]. In addition, some studies have demonstrated that, they are also present at low densities in the dorsal raphe nucleus, striatum, substantia nigria and nucleus accumbens [25,28-30,38].
In the marmoset forebrain, 5-HT3 receptors are found in medial habenula nucleus and the hippocampus [32]. In the human forebrain, 5-HT3 receptors are found essentially in limbic structures such as amygdala, hippocampus, nucleus accumbens and striatum whereas only low levels of 5-HT3 receptors have been found in cortex [23,28,39-42].
A significant amount of 5-HT3 receptors are localized on presynaptic nerve fibers and terminals [30,43-46]. Indeed, in cortical areas, amygdala [47] and in striatum [48,49], 5-HT3 receptors are essentially located presynaptically whereas in hippocampus, the postsynaptic receptors are predominant [47].
2.3. Pharmacology and Physiology of 5-HT3 Receptors
5-HT3 receptors are permeable to Na+, K+ and Ca2+ [50,51]. Stimulation leads to opening of the ion channel, inducing a rapid membrane depolarization mediated by cation flow [52,53]. The function of 5-HT3 receptors depend on their localization: nerve-terminal 5-HT3 receptors activation leads to release of various neurotransmitters such as 5-HT, DA or GABA [54], whereas the activation of postsynaptic 5-HT3 receptors is involved in fast synaptic transmission [55,56]. Pre- and postsynaptic receptors are associated with specific characteristics including a different hill coefficient, different single channel conductances, different kinetics, a different re-sensitization time-course [17,57,58]. In particular, presynaptic 5-HT3 receptor displays a high permeability to Ca2+ [45,46,48,59], whereas postsynaptic receptors display a lower permeability to Ca2+ compared to Na+ and K+ [50,60]. Similarly, homomeric 5-HT3A receptors are equally permeable to monovalent and divalent cations whereas heteromeric 5-HT3 receptors have a lower permeability to Ca2+ [17,61,62]. Moreover, heteromeric receptors display faster activation and deactivation kinetics than homomeric receptors [62]. In vitro studies are generally performed in cultured cells expressing only homomeric 5-HT3 receptors [61] which can explain several differences obtained by in vitro and in vivo studies.
5-HT3 receptor agonists and antagonists present different affinity and efficacy depending on the structure of 5-HT3 receptors, i.e., heteromeric or homomeric [17,58,61] (Table 1). 5-HT3 receptor agonists seem not have clinical interests. Frequently used preclinical tool agonists are 1-(m-chlorophenyl)-biguanide (mCPBG) and 2-methyl-5-HT (2-Me-5-HT) which do not cross the blood brain barrier. These compounds are not selective for 5-HT3 receptors since mCPBG has notable affinity for the DA transporter [63] while 2-Me-5-HT has notable affinity for other 5-HT receptor subtypes [64,65]. SR57227A is the mostly used 5-HT3 receptor agonist that crosses the blood brain barrier [66].
In comparison to 5-HT3 receptor agonists, many 5-HT3 receptor antagonists have been developed and they are widely used in the clinic. The main therapeutic use of 5-HT3 receptor antagonist is for chemotherapy-induced emesis [67]. However, other therapeutic uses of 5-HT3 receptor antagonist have been suggested e.g. pain, addiction and psychiatric disorders [68]. In regard with psychiatric disorders, 5-HT3 receptor antagonists present anxiolytic and antidepressants effect (see below) but they may also have antipsychotic effect even if data are yet controversial [21,69]. The 5-HT3 antagonists may be identified by the suffix setron. Different drugs belonging to the “setron class” are used in the clinic: MDL 73,147EF (dolasetron), GR38032F (ondansetron), BRL 43694 (granisetron), ICS 205-939 (tropisetron), palanosetron listed according to ascending receptor binding affinity (7.73 to 10.45 nM) [67]. Other “setrons” have been used in preclinical studies including DAU 6215 (itasetron), BRL-46470A (ricasetron), MDL 72222 and LY277359 (zatosetron).
Curiously, 5-HT3 receptor agonist or antagonist responses are frequently associated with a bell-shaped dose-response curve and this is the case for both clinical and preclinical studies. Generally, the maximum effect is typically observed at very low dose, in the microgram range, while higher doses are ineffective [21]. For example, such responses were observed in the rat learned helplessness [70], the forced swim test and the tail suspension test [71] as well as in the induction of theta rhythms [72]. The inverse dose-response relation of 5-HT3 receptor agonists may be explained by receptor desensitization. Receptor desensitization is involved in mediation of short-term plasticity of synapses. Such mechanisms seem to be involved in regulation of 5-HT3 receptors activity in cultured cells [52]. This desensitization can be explained by receptor internalization. In fact, when cells expressing 5-HT3 receptors in the plasma membrane were activated by the 5-HT3 receptor agonist, mCPBG, a decrease of 5-HT3 receptor density was observed after a few minutes [73]. Internalization can be prevented by the 5-HT3 receptor antagonist ondansetron [74]. Interestingly, it has been recently demonstrated that the 5-HT3 receptor antagonist palanosetron can also induce 5-HT3 receptor internalization and cause prolonged inhibition of receptor function whereas ondansetron and granisetron do not [75]. Moreover, MDL7222 fails to induce internalization of 5-HT3 receptors since a chronic treatment with this compound does not alter sensitivity to a 5-HT3 receptor agonist [76]. Thus, internalization of 5-HT3 receptors is not sufficient to explain the bell-shaped dose-response curve observed under various conditions [21] brought forward hypothesizes of steric hindrance at higher concentrations, different effect on hetero/homoreceptors inducing an effect on some receptors at a low concentration and another effect at higher concentration explaining a heterogeneous response depending on concentration.
3. Effect of 5-HT3 Receptor Ligands on Neuronal Firing and Neurotransmitter Release
3.1. Interaction with 5-HT Systems: Effect in Dorsal Raphe Nucleus
Dorsal raphe nucleus is the brain structure with the highest density of 5-HT cell bodies. Electrophysiological studies demonstrated that the 5-HT3 receptor agonist, phenylbiguanide, has no effect on the dorsal raphe nucleus 5-HT cell firing whereas the other 5-HT3 receptor agonist 2-Me-5-HT has an inhibitory action [65,77]. However, the interpretation of these findings are difficult as there are reports showing that a 5-HT1A receptor antagonist can prevent the suppressant effect of a 5-HT3 agonist [77], and others reporting a lack of effect of a 5-HT3 receptor antagonist and 5-HT1A antagonist to reverse the effect of the agonist [65]. Furthermore, both in vivo and in vitro, studies of various 5-HT3 receptor antagonists including ondansetron and zacopride report no significant effect on the dorsal raphe nucleus 5-HT cell firing [65,77]. Interestingly, it has been shown in vitro that 2-Me-5-HT induces a release of 5-HT in raphe nuclei slices both under basal conditions and also after stimulations [78]. This release of 5-HT explains the decrease of firing observed with 5-HT3 receptor agonists since dorsal raphe nucleus neurons mediate inhibitory effects at 5-HT1A autoreceptors. The 5-HT release induced by 5-HT3 receptor agonists is also not specific in basal conditions: it can be antagonized by fluoxetine and not by 5-HT3 receptor antagonists whereas it is reversed in stimulated conditions by ondansetron [78]. Thus, it seems that in dorsal raphe nucleus, there is no detectable 5-HT tone at 5-HT3 receptors and that contradictory results obtained in basal conditions can be explained by the poor selectivity of 5-HT3 receptors agonists.
3.2. Interaction with GABAergic Interneurons
The role of GABAergic neurotransmission in depression is a relative new area of research [79]. Changes in GABAergic function has been observed in animal model of depression [80] and GABA receptor agonists seems to present antidepressant-like properties [81]. Moreover, serotonergic neurons connect mainly GABAergic interneurons suggesting a strong interaction between the two systems [82,83]. GABAergic interneurons expressing 5-HT3 receptors have been essentially detected in the hippocampus and prefrontal cortex [84].
3.2.1. Hippocampus
Since antidepressants modulate serotoninergic and noradrenergic systems in the hippocampus, it has been suggested that the hippocampus is involved in the pathophysiology of depression [85]. The activity of hippocampus is strongly regulated by GABAergic interneurons. These inhibitory neurons are notably controlled by 5-HT inputs [86]. Hippocampus is one of the forebrain areas in which 5-HT3 receptors are localized both post- and pre-synaptically. In the hippocampus, 5-HT3 messenger ribonucleic acids (mRNAs) and proteins are essentially localized on interneurons [87]. Interestingly, more than 50% of the hippocampal GABAergic interneurons express 5-HT3 receptors [88]. 5-HT3A and 5-HT3B mRNA and protein subunits are present in the human hippocampus [17,89].
In vivo, infusion of the 5-HT3 receptor agonist 2-Me-5-HT induces 5-HT release in the hippocampus [78,90]. Interestingly, after an 10 µM infusion of 2-Me-5-HT the 5-HT level rapidly goes back to baseline while with after 1µM, the effect is maintained for 45 minutes [90]. Similarly, in vitro electrically-evoked 5-HT release was enhanced by a 5-HT3 receptor agonist [65]. These effects are prevented by 5-HT3 receptor antagonists while the antagonist has no effect on basal 5-HT release [65,90]. Thus, hippocampal 5-HT3 receptors seem not to be tonically activated. Nevertheless, even if 5-HT3 receptor antagonists have no basal effect on the hippocampal 5-HT release, they can increase 5-HT levels induced by the selective serotonin reuptake inhibitor (SSRI) paroxetine [91].
In vivo, 2-Me-5-HT reduces the hippocampal transmission by reducing the amplitude of evoked field potentials and this effect is blocked by the 5-HT3 receptor antagonist ricasetron [92] whereas in vitro 2-Me-5-HT has no significant electrophysiological effect on CA1 pyramidal cells at basal state [93,94]. Nevertheless, it reduces clearly hippocampal plasticity by decreasing long term potentiation (LTP) and long term depression (LTD), evoked by stimulation of Schaffer collaterals [93,94]. Conversely, 5-HT3 receptor antagonists induce an increase in LTP induction associated with behavioral increase of memory [72,94,95]. Granisetron and a GABA receptor antagonist prevent this effect whereas 5-HT1 or 5-HT2 receptor antagonists have no effect [93, 94]. As mentioned above, 5-HT3 receptor stimulation in hippocampus induces a release of 5-HT and it is well-known that 5-HT has an inhibitory effect on hippocampal LTP [96] which may explain the suppressant effect of a 5-HT3 receptor agonist on LTP. Interestingly, a 5-HT3 receptor antagonist prevented this 5-HT-mediated blockage of LTP [93,96,97]. Firing rate of in vivo CA1 pyramidal cells that are iontophoretically activated by glutamate is suppressed by SR57227A and 2-me-5-HT, whereas the 5HT3 receptor antagonist ricasetron prevents this suppressant effect [92].
In hippocampus slices, 5-HT increases the frequency and amplitude of spontaneous GABAergic inhibitory postsynaptic potentials (IPSPs), an effect that is inhibited by 5-HT3 receptor antagonists [43,59,98-100]. A 5-HT3 receptor agonist acts similar to the endogenous ligand on IPSPSs [59]. Thus, 5-HT3 receptor agonists depolarize hippocampal interneurons [101,102] and induce GABA release by opening of voltage-gated Ca2+ channels [59,100]. This effect has a rapid onset and desensitization [59]. These inhibitory GABAergic interneurons regulate function of both hippocampal CA1 pyramidal cells and dentate gyrus cells [101].
5-HT3 receptor agonists reduce both hippocampal transmission and plasticity and 5-HT3 receptor antagonists inhibit this effect. Stress reduces LTP and it has been suggested that alterations of hippocampal LTP may have a role in the etiology of depression. Yet, current antidepressants inhibit LTP [3]. Hence, addition of a 5-HT3 receptor antagonist may prevent the antidepressant-induced LTP decrease and thereby improve efficacy of current antidepressants and including memory deficits.
3.2.2. Prefrontal Cortex
Prefrontal cortex is frequently metabolically overactive in treatment-resistant depression and clinical improvement after pharmacotherapy, psychotherapy or limbic leucotomy is correlated with decreases in its metabolic activity [103]. The majority of 5-HT3 receptors in the prefrontal cortex cells are co-expressed with glutamic acid decarboxylase (GAD), a marker of GABAergic neurons [35]. In the neocortex, more than 90% of cells expressing 5-HT3 receptors are GABAergic neurons [84,88]. In prefrontal cortex, only 5-HT3A receptor subunit seems to be expressed [104]. GABAergic interneurons expressing 5-HT3 receptors are also co-localized with cholecystokinin (CCK) and vasoactive intestinal peptides but not somatostatin, and may express the Ca2+ binding proteins calbindin and calretinin but not parvalbumin [88,104]. Similarly in rhesus monkeys, the majorities of 5-HT3 receptor positive cortical cells are GABAergic neurons and generally co-expresses substance P receptors. These neurons can also express calbindin or calretinin [105].
In vitro 2-Me-5-HT enhances the 5-HT release in rat and guinea pig frontal cortex, an effect inhibited by 5-HT3 receptor antagonists [65,106]. Applied by microiontophoresis, they induce an inhibitory effect on the firing activity of medial prefrontal cortex cells [65,107-110]. Some investigators found that the effect is specific, i.e., it is blocked by selective 5-HT3 receptor antagonists and not by 5-HT1 and 5-HT2 receptor antagonist and GABA-A receptor antagonists [107-110], whereas for others report that the effect is not blocked by a 5-HT3 receptor antagonist but by a 5-HT1A receptor antagonist [65]. Similarly, 5-HT3 receptor agonists depressed firing of glutamate-activated quiescent medial prefrontal cortex cells [109]. Moreover, SR57227A depressed N-Methyl-d-aspartate (NMDA)-evoked membrane depolarization, action potentials, and inward current in the rat medial prefrontal pyramidal cells, an effect that was reversed by a 5-HT3 receptor antagonist [111]. Interestingly, acute administration of an SSRI, like citalopram, increases 5-HT levels in prefrontal cortex and ondansetron can enhance this increase [91].
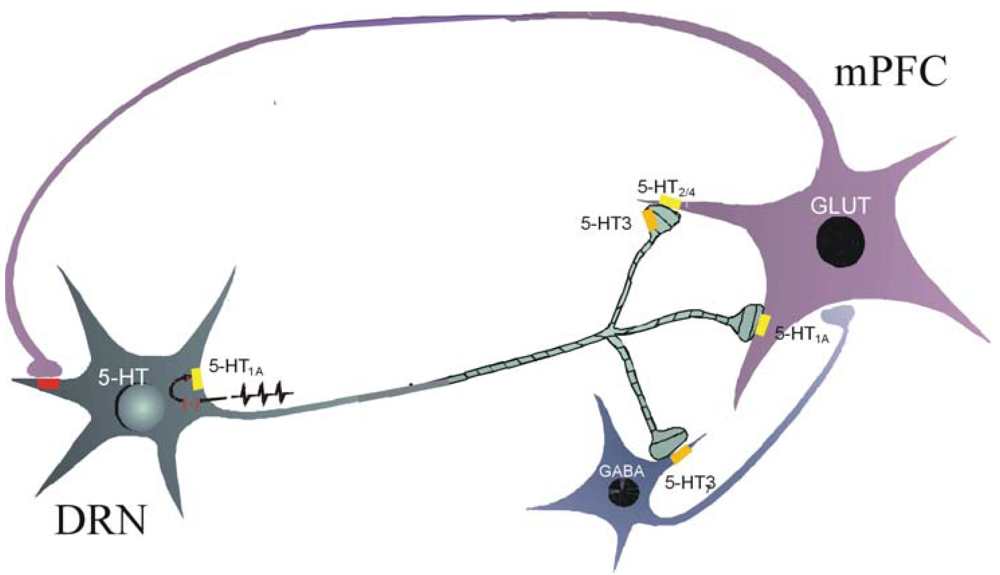
The suppressant effect on the firing activity of medial prefrontal cortex cells induced by a 5-HT3 receptor agonist is mediated by an indirect mechanism. A 5-HT3 receptor agonist induces a direct inward current with a rapid desensitization of interneurons but has no effect in cultured pyramidal cells [112]. Thus, it appears that 5-HT3 receptor agonists increase inhibitory postsynaptic currents (IPSCs) in cortical pyramidal neurons via stimulation of 5-HT3 receptors located on inhibitory GABAergic interneurons [104,112]. In contrast to 5-HT2A receptors which are localized both in inhibitory and excitatory neurons, 5-HT3 receptors have only an inhibitory effect in cortex [105,112]. As a consequence, a pulse of 5-HT is inhibitory while prolonged presence of 5-HT can induce enhancement of transmission because of 5-HT2A receptor activation and 5-HT3 receptor desensitization [112]. There is a regulatory loop between raphe dorsalis and prefrontal cortex since stimulation of dorsal raphe induces an excitation of slow-spiking GABAergic neurons in prefrontal cortex inducing a suppression of cortical cells, an effect blocked by a 5-HT3 receptor antagonist [35,108] (Figure 1).
3.3. Interaction with the Dopamine System
The dopamine system can be divided into three pathways. The nigrostriatal dopaminergic pathway consists of the substantia nigra pars compacta (SNc, A9) and their associated efferent targets in the dorsal striatum. The mesolimbic dopaminergic pathway contains A10 dopaminergic neurons located within the ventral tegmental area (VTA) and their associated efferent targets in the ventral striatum including nucleus accumbens and limbic structures (e.g., amygdala and hippocampus). The mesocortical dopaminergic pathway consists of the A10 dopaminergic neurons and their associated efferent targets in the prefrontal cortex. It has been suggested that abnormalities of dopaminergic neurotransmission have been implicated in the pathogenesis of affective disorders [113,114]. In particular, several studies have suggested that the mesolimbic pathway can be altered in mood disorders. For example, stress, in animal models of depression, activates VTA DA neurons and their limbic efferent targets [115]. Moreover, the DA system is now one of the targets of depression treatment, for example drugs initially used in schizophrenia like aripiprazole or quetiapine are now used in depressed patients [8].
3.3.1. Substantia nigra compacta (SNc)
Acute administration of ondansetron, zatosetron or itasetron fails to modify the number of DA neurons that are spontaneously active in SNc (Table 2). Similarly, 21-day-chronic treatment with 5-HT3 receptor antagonists has no effect, except for zatosetron at 0.1 mg/kg/day [116] and dolasetron at 5 mg/kg/day [117] which both decreased number of DA cells that are spontaneously active(Table 2). Moreover, 5-HT3 receptor antagonists fail to affect sensibility to the D2-like receptor agonist apomorphine in SNc [118].
3.3.2. Striatum
Infusion of striatal slices with 5-HT3 receptor agonists, including 5-HT, produce an increase in DA release, an effect that is blocked by a 5-HT3 receptor antagonist [119-122]. Similarly, in vivo studies in rats demonstrated that endogenous 5-HT stimulates release of 5-HT in striatum and that this effect is partially blocked by a 5-HT3 receptor antagonist [123]. Notably, these data are controversial and in some studies the release induced by 5-HT3 receptor agonists is not found to be 5-HT3 receptor specific [124,125] wheras in other studies 5-HT3 receptor agonists fail to modify basal striatal DA release [126]. 5-HT3 receptor antagonists have failed to induce DA release in the striatum both in vitro and in vivo [127-132]. Dorsal raphe nucleus stimulation produces a DA decrease in the striatum which is not modified by 5-HT3 receptor antagonists [133]. In fact, 5-HT3 receptor-induced DA release may be effective only when both DA and 5-HT tone is increased [134].
3.3.3. Ventral tegmental area (VTA)
Local infusion of mCPBG induces an increase of extracellular levels of DA in VTA slices, an effect that is inhibited by a 5-HT3 receptor antagonist and depending on Ca2+ [76,135,136]. On the other hand, acute administration of 5-HT3 receptor antagonists ricasetron and itasetron has no effect on the firing of VTA DA neurons [118,137,138] and do not affect sensitivity to the D2-like receptor agonist apomorphine [118]. Nevertheless, clorgyline, a monoamine oxidase inhibitor (MAOI), inhibits VTA DA firing rate and ondansetron reverses this effect [138]. Effects of 5-HT3 receptor antagonists on the number of DA cells per track are rather variable. Both after acute and chronic treatment, 5-HT3 receptor antagonists have no effect or induced an increase or a decrease of the number of DA cells that are spontaneously active. These results depend on the 5-HT3 receptor antagonist used and the dose used (Table 2). Moreover, when a 5-HT3 receptor antagonist produces a decrease of the number of DA cells/track after a chronic treatment, apomorphine can or cannot reverse this effect [118].
3.3.4. Nucleus accumbens
In vivo and in vitro local administrations of a 5-HT3 receptor agonist induced an increase of extracellular DA release in nucleus accumbens, an effect which is blocked by a 5-HT3 receptor antagonist [76,139-141], suggesting a modulation of DA function by presynaptic 5-HT3 receptors in this area [142]. Moreover, in 5-HT3 receptors over-expressed mice, the DA release induced by a 5-HT3 receptor agonist was increased compared to wild type [143]. Similarly, dorsal raphe nucleus stimulation increases DA release in nucleus accumbens which is attenuated by a 5-HT3 receptor antagonist [133]. Interestingly, in an animal model of depression, namely the Flinder sensitive Line rats, 5-HT3 receptor agonists do not increase DA levels in nucleus accumbens while a chronic antidepressant treatment restore the DA increase in response to agonist [113]. Acute administration of a 5-HT3 receptor antagonist does not modify the rate of DA release in nucleus accumbens [130,144] while chronic treatment with the 5-HT3 receptor antagonist MDL72222 decreases the extracellular concentration of DA in the nucleus accumbens [76,131]. This effect is not due to an alteration of sensitivity of 5-HT3 receptors since the DA release induced by a 5-HT3 receptor agonist is not altered after MDL72222 treatment [76]. Finally, local administration of a 5-HT3 receptor antagonist in nucleus accumbens has an anxiolytic-like effect in different behavioral tests such as open-field, Vogel conflict test and light-dark exploration [145,146].
3.4. Interaction with Other Neurotransmitters
There is less data concerning other neurotransmitters release and this relevance of this data for depression is less clear data. Nevertheless, there is some evidence that 5-HT3 receptors may also induce a release of glutamate from nerve terminals [148-150]. Also, there might be a role for acetylcholine (Ach), since 5-HT3 receptor activation inhibits cortical release of Ach. It is not clear, however, whether this is a direct effect. Moreover 5-HT3 receptor antagonist induced an increase of cortical Ach release, an effect that was potentiated by GABA receptor antagonists. This has led to the suggestion that the augmented ACh release by 5-HT3 antagonists causes a blockade of GABA-mediated inhibition on cholinergic neurons [14,151]. Finally, some studies evoked an increase of noradrenaline induced by activation of 5-HT3 stimulation but some others data are controversial and this effect seems not specific [85].
4. 5-HT3 Receptors as Drug Target for Treatment of Anxiety and Depression
4.1. Effect of Current Antidepressants on 5-HT3 Receptors
Some of the currently used antidepressants show affinity for 5-HT3 receptors. Thus, the tricyclic antidepressant (TCA) Imipramine, the SSRI fluoxetine, the non-selective α2-adrenoceptor antagonist mirtazapine and the MAOI phenelzine dose-dependently block the inward current mediated by 5-HT3 receptors expressed in cultured cells [152-156]. Moreover, several antidepressants, including fluoxetine, can inhibit binding of a 5-HT3 receptor antagonist [157]. Interestingly, fluoxetine induced an increase of Polysialic Acid Neural Cell Adhesion Molecule (PSA-NCAM), a molecule implicated in neuroplasticity, an effect that was blocked by administration of 5-HT3 receptor antagonist, thus suggesting a direct effect of fluoxetine on 5-HT3 receptors [158]. Finally, fluoxetine inhibits 5-HT release induced by 5-HT3 receptor agonists in the dorsal raphe nucleus [78]. Except for mirtazapine, the latter effect of antidepressants on 5-HT3 receptors seems non-competitive[155,156,159]. Similarly, typical and atypical antipsychotics antagonize 5-HT3 receptor in a non competitive manner [160]. This has led to the suggestion of an allosteric recognition site different from the 5-HT3 binding site [155]. Interestingly, the non-competitive 5-HT3 antagonism of antidepressants seem not associated with an increase of internalization [161]. It has also been shown that 5-HT3 receptors and antidepressants are colocalized in specific domain of membrane cells, the raft-like domains. Nevertheless antidepressants may exert effect on 5-HT3 receptor despite disruption of lipid rafts [161,162].
Electrophysiological studies exploring recovery of firing after chronic (from 2 days to 3 weeks) antidepressant treatments in anesthetized rats suggested that mirtazapine may be more rapid than SSRIs in reversing the decrease of 5-HT dorsal raphe nucleus firing rate. Also, co-administration of paroxetine and mirtazapine may accelerate this index of antidepressant response compared to either drug alone [163]. In preclinical studies, cyamemazine, an atypical antipsychotic with D2-like, 5-HT2A, 5-HT2C and 5-HT3 receptor antagonism properties [164] presents anxiolytic-like activity. In clinical studies, cyamemazine improved the anxious syndrome [164]. Finally, it has been suggested that electroconvulsive therapy may potentiate 5-HT3 receptor function in hippocampal CA1 pyramidal cells [165].
4.2. Lu AA21004
Lu AA21004 is a 5-HT3 receptor antagonist, 5-HT1A receptor agonist (h5-HT3 and h5-HT1A receptors: Ki = 4.5 and 15 nM, respectively) and an inhibitor of the 5-HT transporter (5HTT) (h5HTT: IC50 = 5.4 nM) [166]. This in vitro profile translates into enhanced levels of 5-HT as well as other neurotransmitters (i.e., noradrenaline, dopamine, acetylcholine) in hippocampus and prefrontal cortex after in vivo administration [91,166]. It is different from others antidepressants since it has an effect on 5-HT levels even at a low transporter occupancy [91,166]. Moreover, it displays anxiolytic-like and antidepressant-like activity in various validated rodents models with a better efficacy than current antidepressant drugs [91,166]. Interestingly, in electrophysiological studies, Lu AA21004 has a preclinical profile of a fast-acting antidepressant, since it induced early 5-HT1A receptor desensitization compared to fluoxetine, an effect probably mediated by 5-HT3 receptors blockade [167]. Currently in phase III trials, LuAA21004 displays very good efficacy and well tolerance [168].
4.3. Preclinical Studies of 5-HT3 Receptor Ligands
Poncelet et al. [169] demonstrate that the selective 5-HT3 receptor agonist SR57227A produces antidepressant-like effect in different behavioral tests (forced swimming test, learned helplessness) in rodents. In contrast, other investigators report 5-HT3 receptor agonists alone or in combination with antidepressants to be ineffective in the forced swimming test [170] and others report that 5-HT3 receptor agonists attenuate the effects of antidepressants in this animal model [171]. These variable results may be explained by the different doses tested and may also ascribed to SR57227A being a partial 5HT3 agonist.
In the learned helplessness test, zacopride, ondansetron and tropisetron reverse the escape failure with a biphasic dose effect relationship [70]. In the forced swim test and tail suspension test, acute and chronic treatments with 5-HT3 receptor antagonists decrease the immobility time [71,172-175]. Interestingly, in the forced swim test, ondansetron increases the efficacy of fluoxetine, venlafaxine and citalopram [71,170]. In bulbectomized rats (another animal model of depression) 5-HT3 receptor antagonists reverse their depression-like phenotype [71,175]. The novel putative antidepressant (4-benzylpiperazin-1-yl) (quinoxalin-2-yl) methanone (QCF-3) which is a 5-HT3 receptor antagonist displayed also antidepressant properties both alone and in association with fluoxetine [176]. Finally, antidepressants induce a decrease in rapid eyes movements (REM) sleep. Microinjection of the 5-HT3 receptor agonist mCPBG in the dorsal raphe nucleus causes a reduction of rapid eye movement sleep (REMS) whereas ondansetron prevented this reduction. [150]. Thus, administration of a 5-HT3 receptor antagonist may have a beneficial effect on sleep disturbances induced by antidepressants.
Anxiolytic activity of 5-HT3 receptor antagonists have been extensively studied in animal models [177,178]. Griebel et al. [178] analyzed more than 75 5-HT3 receptor related experiments in which two-third of the studies demonstrated anxiolytic-like effect of 5-HT3 receptor antagonists and one-third of the studies failed to reveal an effect. For example, in some studies, 5-HT3 receptor antagonists disinhibited punished behavior in the Vogel test [145,179,180] and in other studies 5-HT3 receptor antagonists were inactive [181]. Interestingly, after chronic treatment with diazepam, 5-HT3 receptor agonists had no more anxiogenic effect, suggesting that diazepam may have induced a 5-HT3 receptor desensitization [182].
Another index used to evaluate anxiolytic effect of drugs is by examining its effect on cholecystokinin [183]. Interestingly, GABAergic interneurons expressing 5-HT3 receptors co-express cholecystokinin [88,104]. It has been shown that 5-HT or 5-HT3 receptor agonists induce a release of cholecystokinin in cortical or accumbal synaptosomes while 5-HT3 receptor antagonists decrease spontaneous- or induced-release of CCK [184,185]. However, in another study, 5-HT3 receptor antagonists fail to prevent the increase of CCK induced by stress in rat prefrontal cortex [186]. Finally, it is well known that amygdala is involved in physiology of anxiety [187]. As described previously, 5-HT3 receptors are present at significant levels in the amygdala. In the mouse amygdala, local administration of 5-HT3 receptor antagonists attenuates aversive response whereas 5-HT3 receptor agonists increase this [146]. Under the same conditions, in the social interaction test, 5-HT3 receptor antagonists have also an anxiolytic-like activity [146]. It has been suggested that the amygdala is involved in disinhibitory effects induced by various 5-HT3 receptor antagonists [188].
5-HT3A receptor knockout male mice present anxiety-like behavior in elevated plus maze, novelty interaction animal models and light/dark box models of anxiety [189,190]. In the forced swim test, male 5-HT3A knock-out (KO) did not differ from the wild type rats, whereas female 5-HT3 knockout mice showed increased immobility [191]. Mice overexpressing 5-HT3 receptors show decreased anxiety in the elevated plus-maze and in the exploration paradigm [192].
4.4. Clinical Studies with 5-HT3 Receptor Antagonists
5-HT3 receptor antagonists have been tested on anxiety and depressive syndromes associated to other diseases. For example, ondansetron reduced depressive symptoms in patients with chronic hepatitis C [193], with alcoholism [194] and in bulimic patients [195]. Similarly in fibromyalgic patients, tropisetron improved anxiety and depressive scores [196]. Moreover, in pathology with common symptoms of depression like chronic fatigue or fibromyalgia, 5-HT3 receptor antagonist have been reported efficacious [193,197-199], although negative results in chronic fatigue syndrome have been also reported [200]. Suspicion of 5-HT syndrome after use of setron alone or in combination with mirtazapine has reported in one study [201].
In healthy volunteers, ondansetron has been explored in emotional processing tasks. It abolishes the emotion potentiated startle effect, revealing an anxiolytic-like activity [202]. In anxious patients, zatosetron tends to reduce anxiety however the results from this study was not significant [203]. Similarly, tropisteron improves anxiety scores in patients suffering from generalized anxiety disorders [204]. In clinical studies, results concerning CCK release were contradictory as in preclinical studies. Indeed, in patients with panic disorder and social phobia, ondansetron fails to prevent the anxiety induced by pentagastrin, a CCK agonist [205]. Whereas, in healthy volunteers, ondansetron prevents panic symptoms induced by CCK tetrapeptide [206].
5. Conclusions
5-HT3 receptors are in numerous ways intimately involved in the regulation of neurotransmitter systems that are known to be of relevance for several psychiatric disorders including mood disorders, schizophrenia, eating disorders and addiction [21,207]. As becomes evident from the present review, the involvement of 5-HT3 receptors is complex and also context dependent. Their molecular structure, function and regulation are only partly elucidated. It will be important to understand why several responses associated with 5-HT3 receptor ligands present a bell-shaped dose-response curve. In conclusion, we feel that additional knowledge about 5-HT3 receptor function and their role in several diseases may offer new therapeutic opportunities in the future.


{kind=link}
| 5-HT3 receptor agonist | 5-HT3 receptor antagonists |
|---|---|
| 1-(m-chlorophenyl)-biguanide (mCPBG) | MDL 73,147EF (dolasetron) |
| 2-methyl-5-HT (2-me-5-HT) SR57227A | GR38032F (ondansetron) |
| BRL 43694 (granisetron) | |
| ICS 205-939 (tropisetron) | |
| DAU 6215 (itasetron) | |
| BRL-46470A (ricasetron) | |
| LY277359 (zatosetron) | |
| MDL 72222 | |
| Palanosetron |
| Cell per track | ||||
|---|---|---|---|---|
| 5-HT3 receptor antagonist | Duration | Dose | A9 (SNc) | A10 (VTA) |
| Dolasetron [117] | Acute treatment | 500 µg/kg, i.v. | 0 | 0 |
| Zatosetron [116] | 0.1 mg/kg, i.v. | 0 | + | |
| 1 mg/kg, i.v. | 0 | + | ||
| Zatosetron [147] | 10 mg/kg, i.v. | 0 | 0 | |
| 0.01 mg/kg, i.p. | 0 | 0 | ||
| 0.1 mg/kg, i.p. | 0 | − | ||
| 1 mg/kg, i.p. | 0 | 0 | ||
| 10 mg/kg, i.p. | 0 | 0 | ||
| Itasetron [118] | 15 µg/kg, s.c. | 0 | + | |
| Dolasetron [117] | Chronic treatment (21 days) | 5 mg/kg/day, i.p. | − | − |
| Granisetron [137] | 5 mg/kg/day, i.p. | 0 | 0 | |
| 10 mg/kg/day, i.p. | 0 | 0 | ||
| Itasetron [118] | 30 µg/kg/day, s.c. | 0 | − | |
| Zatosetron [116] | 0.1 mg/kg/day, i.p. | − | − | |
| 1 mg/kg/day, i.p. | 0 | + | ||
| 10 mg/kg/day, i.p. | 0 | − | ||
| Zatosetron [147] | 0.01 mg/kg, i.p. | 0 | 0 | |
| 0.1 mg/kg, i.p. | 0 | − | ||
| 1 mg/kg, i.p. | 0 | 0 | ||
| 10 mg/kg, i.p. | 0 | 0 | ||
References
- Kessler, R.C.; Kessler, R.C.; Berglund, P.; Demler, O.; Jin, R.; Koretz, D.; Merikangas, K.R.; Rush, A.J.; Walters, E.E.; Wang, P.S. The epidemiology of major depressive disorder: Results from the National Comorbidity Survey Replication (NCS-R). JAMA 2003, 289, 3095–3105. [Google Scholar]
- Kunugi, H.; Hori, H.; Adachi, N.; Numakawa, T. Interface between hypothalamic-pituitary-adrenal axis and brain-derived neurotrophic factor in depression. Psychiat. Clin. Neurosci. 2010, 64, 447–459. [Google Scholar]
- Faure, C.; Mnie-Filali, O.; Haddjeri, N. Long-term adaptive changes induced by serotonergic antidepressant drugs. Expert Rev. Neurother. 2006, 6, 235–245. [Google Scholar]
- Berton, O.; Nestler, E.J. New approaches to antidepressant drug discovery: Beyond monoamines. Nat. Rev. Neurosci. 2006, 7, 137–151. [Google Scholar]
- Duman, R.S. Depression: A case of neuronal life and death? Biol Psychiat. 2004, 56, 140–145. [Google Scholar]
- Blier, P. The pharmacology of putative early-onset antidepressant strategies. Eur. Neuropsychopharmacol. 2003, 13, 57–66. [Google Scholar]
- Artigas, F.; Adell, A.; Celada, P. Pindolol augmentation of antidepressant response. Curr.Drug Targets 2006, 7, 139–147. [Google Scholar]
- Etievant, A.; Betry, C.; Haddjeri, N. Partial dopamine D2/serotonin 5-HT1A receptor agonists as new therapeutic agents. Open Neuropsychopharmacol. J. 2010, 3, 1–12. [Google Scholar]
- Celada, P.; Puig, M.; Amargos-Bosch, M.; Adell, A.; Artigas, F. The therapeutic role of 5-HT1A and 5-HT2A receptors in depression. J. Psychiat. Neurosci. 2004, 29, 252–265. [Google Scholar]
- Marek, G.J.; Carpenter, L.L.; McDougle, C.J.; Price, L.H. Synergistic action of 5-HT2A antagonists and selective serotonin reuptake inhibitors in neuropsychiatric disorders. Neuropsychopharmacology 2003, 28, 402–412. [Google Scholar]
- Lucas, G.; Rymar, V.V.; Du, J.; Mnie-Filali, O.; Bisgaard, C.; Manta, S.; Lambas-Senas, L.; Wiborg, O.; Haddjeri, N.; Piñeyro, G.; et al. Serotonin(4) (5-HT(4)) receptor agonists are putative antidepressants with a rapid onset of action. Neuron 2007, 55, 712–725. [Google Scholar]
- Lucas, G. Serotonin receptors, type 4: A new hope? Curr.Drug Targets 2009, 10, 1085–1095. [Google Scholar]
- Mnie-Filali, O.; Lambas-Senas, L.; Zimmer, L.; Haddjeri, N. 5-HT7 receptor antagonists as a new class of antidepressants. Drug News Perspect. 2007, 20, 613–618. [Google Scholar]
- Barnes, N.M.; Sharp, T. A review of central 5-HT receptors and their function. Neuropharmacology 1999, 38, 1083–1152. [Google Scholar]
- Maricq, A.V.; Peterson, A.S.; Brake, A.J.; Myers, R.M.; Julius, D. Primary structure and functional expression of the 5HT3 receptor, a serotonin-gated ion channel. Science 1991, 254, 432–437. [Google Scholar]
- Cederholm, J.M.; Schofield, P.R.; Lewis, T.M. Gating mechanisms in Cys-loop receptors. Eur. Biophys. J. 2009, 39, 37–49. [Google Scholar]
- Davies, P.A.; Pistis, M.; Hanna, M.C.; Peters, J.A.; Lambert, J.J.; Hales, T.G.; Kirkness, E.F. The 5-HT3B subunit is a major determinant of serotonin-receptor function. Nature 1999, 397, 359–363. [Google Scholar]
- Niesler, B.; Frank, B.; Kapeller, J.; Rappold, G.A. Cloning, physical mapping and expression analysis of the human 5-HT3 serotonin receptor-like genes HTR3C, HTR3D and HTR3E. Gene 2003, 310, 101–111. [Google Scholar]
- Karnovsky, A.M.; Gotow, L.F.; McKinley, D.D.; Piechan, J.L.; Ruble, C.L.; Mills, C.J.; Schellin, K.A.; Slightom, J.L.; Fitzgerald, L.R.; Benjamin, C.W.; et al. A cluster of novel serotonin receptor 3-like genes on human chromosome 3. Gene 2003, 319, 137–148. [Google Scholar]
- Holbrook, J.D.; Gill, C.H.; Zebda, N.; Spencer, J.P.; Leyland, R.; Rance, K.H.; Trinh, H.; Balmer, G.; Kelly, F.M.; Yusaf, S.P.; et al. Gunthorpe, Characterisation of 5-HT3C, 5-HT3D and 5-HT3E receptor subunits: evolution, distribution and function. J. Neurochem. 2009, 108, 384–396. [Google Scholar]
- Faerber, L.; Drechsler, S.; Ladenburger, S.; Gschaidmeier, H.; Fischer, W. The neuronal 5-HT3 receptor network after 20 years of research—Evolving concepts in management of pain and inflammation. Eur. J. Pharmacol. 2007, 560, 1–8. [Google Scholar]
- Pratt, G.D.; Bowery, N.G.; Kilpatrick, G.J.; Leslie, R.A.; Barnes, N.M.; Naylor, R.J.; Jones, B.J.; Nelson, D.R.; Palacids, J.M.; Slater, P.; et al. Consensus meeting agrees distribution of 5-HT3 receptors in mammalian hindbrain. Trends Pharmacol. Sci. 1990, 11, 135–137. [Google Scholar]
- Waeber, C.; Hoyer, D.; Palacios, J.M. 5-Hydroxytryptamine3 receptors in the human brain: autoradiographic visualization using [3H]ICS 205-930. Neuroscience 1989, 31, 393–400. [Google Scholar]
- Barnes, J.M.; Barnes, N.M.; Costall, B.; Deakin, J.F.; Ironside, J.W.; Kilpatrick, G.J.; Naylor, R.J.; Rudd, J.A.; Simpson, M.D.; Slater, P.; et al. Identification and distribution of 5-HT3 recognition sites within the human brainstem. Neurosci. Lett. 1990, 111, 80–86. [Google Scholar]
- Laporte, A.M.; Koscielniak, T.; Ponchant, M.; Verge, D.; Hamon, M.; Gozlan, H. Quantitative autoradiographic mapping of 5-HT3 receptors in the rat CNS using [125I]iodo-zacopride and [3H]zacopride as radioligands. Synapse 1992, 10, 271–281. [Google Scholar]
- Chameau, P.; van Hooft, J.A. Serotonin 5-HT(3) receptors in the central nervous system. Cell Tissue Res. 2006, 326, 573–581. [Google Scholar]
- Kilpatrick, G.J.; Jones, B.J.; Tyers, M.B. The distribution of specific binding of the 5-HT3 receptor ligand [3H]GR65630 in rat brain using quantitative autoradiography. Neurosci. Lett. 1988, 94, 156–160. [Google Scholar]
- Kilpatrick, G.J.; Jones, B.J.; Tyers, M.B. Binding of the 5-HT3 ligand, [3H]GR65630, to rat area postrema, vagus nerve and the brains of several species. Eur. J. Pharmacol. 1989, 159, 157–164. [Google Scholar]
- Gehlert, D.R.; Gackenheimer, S.L.; Wong, D.T.; Robertson, D.W. Localization of 5-HT3 receptors in the rat brain using [3H]LY278584. Brain Res. 1991, 553, 149–154. [Google Scholar]
- Waeber, C.; Dixon, K.; Hoyer, D.; Palacios, J.M. Localisation by autoradiography of neuronal 5-HT3 receptors in the mouse CNS. Eur. J. Pharmacol. 1988, 151, 351–352. [Google Scholar]
- Waeber, C.; Pinkus, L.M.; Palacios, J.M. The (S)-isomer of [3H]zacopride labels 5-HT3 receptors with high affinity in rat brain. Eur. J. Pharmacol. 1990, 181, 283–287. [Google Scholar]
- Jones, B.J.; Costall, B.; Domeney, A.M.; Kelly, M.E.; Naylor, R.J.; Oakley, N.R.; Tyers, M.B. The distribution of 5-HT3 recognition sites in the marmoset brain. Eur. J. Pharmacol. 1992, 215, 63–67. [Google Scholar]
- Morales, M.; Battenberg, E.; de Lecea, L.; Sanna, P.P.; Bloom, F.E. Cellular and subcellular immunolocalization of the type 3 serotonin receptor in the rat central nervous system. Brain Res. Mol. Brain Res. 1996, 36, 251–260. [Google Scholar]
- Kia, H.K.; Miquel, M.C.; McKernan, R.M.; Laporte, A.M.; Lombard, M.C.; Bourgoin, S.; Hamon, M.; Verge, D. Localization of 5-HT3 receptors in the rat spinal cord: immunohistochemistry and in situ hybridization. Neuroreport 1995, 6, 257–261. [Google Scholar]
- Puig, M.V.; Santana, N.; Celada, P.; Mengod, G.; Artigas, F. In vivo excitation of GABA interneurons in the medial prefrontal cortex through 5-HT3 receptors. Cereb. Cortex 2004, 14, 1365–1375. [Google Scholar]
- Barnes, J.M.; Barnes, N.M.; Champaneria, S.; Costall, B.; Naylor, R.J. Characterisation and autoradiographic localisation of 5-HT3 receptor recognition sites identified with [3H]-(S)-zacopride in the forebrain of the rat. Neuropharmacology 1990, 29, 1037–1045. [Google Scholar]
- Steward, L.J.; West, K.E.; Kilpatrick, G.J.; Barnes, N.M. Labelling of 5-HT3 receptor recognition sites in the rat brain using the agonist radioligand [3H]meta-chlorophenylbiguanide. Eur. J. Pharmacol. 1993, 243, 13–18. [Google Scholar]
- Kilpatrick, G.J.; Jones, B.J.; Tyers, M.B. Identification and distribution of 5-HT3 receptors in rat brain using radioligand binding. Nature 1987, 330, 746–748. [Google Scholar]
- Barnes, J.M.; Barnes, N.M.; Costall, B.; Ironside, J.W.; Naylor, R.J. Identification and characterisation of 5-hydroxytryptamine 3 recognition sites in human brain tissue. J. Neurochem. 1989, 53, 1787–1793. [Google Scholar]
- Bufton, K.E.; Steward, L.J.; Barber, P.C.; Barnes, N.M. Distribution and characterization of the [3H]granisetron-labelled 5-HT3 receptor in the human forebrain. Neuropharmacology 1993, 32, 1325–1331. [Google Scholar]
- Parker, R.M.; Barnes, J.M.; Ge, J.; Barber, P.C.; Barnes, N.M. Autoradiographic distribution of [3H]-(S)-zacopride-labelled 5-HT3 receptors in human brain. J. Neurol. Sci. 1996, 144, 119–127. [Google Scholar]
- Abi-Dargham, A.; Laruelle, M.; Lipska, B.; Jaskiw, G.E.; Wong, D.T.; Robertson, D.W.; Weinberger, D.R.; Kleinman, J.E. Serotonin 5-HT3 receptors in schizophrenia: a postmortem study of the amygdala. Brain Res. 1993, 616, 53–57. [Google Scholar]
- Katsurabayashi, S.; Kubota, H.; Tokutomi, N.; Akaike, N. A distinct distribution of functional presynaptic 5-HT receptor subtypes on GABAergic nerve terminals projecting to single hippocampal CA1 pyramidal neurons. Neuropharmacology 2003, 44, 1022–1030. [Google Scholar]
- Kidd, E.J.; Laporte, A.M.; Langlois, X.; Fattaccini, C.M.; Doyen, C.; Lombard, M.C.; Gozlan, H.; Hamon, M. 5-HT3 receptors in the rat central nervous system are mainly located on nerve fibres and terminals. Brain Res. 1993, 612, 289–298. [Google Scholar]
- Nichols, R.A.; Mollard, P. Direct observation of serotonin 5-HT3 receptor-induced increases in calcium levels in individual brain nerve terminals. J. Neurochem. 1996, 67, 581–592. [Google Scholar]
- Nayak, S.V.; Ronde, P.; Spier, A.D.; Lummis, S.C.; Nichols, R.A. Calcium changes induced by presynaptic 5-hydroxytryptamine-3 serotonin receptors on isolated terminals from various regions of the rat brain. Neuroscience 1999, 91, 107–117. [Google Scholar]
- Miquel, M.C.; Emerit, M.B.; Nosjean, A.; Simon, A.; Rumajogee, P.; Brisorgueil, M.J.; Doucet, E.; Hamon, M.; Verge, D. Differential subcellular localization of the 5-HT3-As receptor subunit in the rat central nervous system. Eur. J. Neurosci. 2002, 15, 449–457. [Google Scholar]
- Ronde, P.; Nichols, R.A. High calcium permeability of serotonin 5-HT3 receptors on presynaptic nerve terminals from rat striatum. J. Neurochem. 1998, 70, 1094–1103. [Google Scholar]
- Nayak, S.V.; Ronde, P.; Spier, A.D.; Lummis, S.C.; Nichols, R.A. Nicotinic receptors co-localize with 5-HT(3) serotonin receptors on striatal nerve terminals. Neuropharmacology 2000, 39, 2681–2690. [Google Scholar]
- Yang, J. Ion permeation through 5-hydroxytryptamine-gated channels in neuroblastoma N18 cells. J. Gen. Physiol. 1990, 96, 1177–1198. [Google Scholar]
- Hargreaves, A.C.; Lummis, S C.; Taylor, C.W. Ca2+ permeability of cloned and native 5-hydroxytryptamine type 3 receptors. Mol. Pharmacol. 1994, 46, 1120–1128. [Google Scholar]
- Yakel, J.L.; Jackson, M.B. 5-HT3 receptors mediate rapid responses in cultured hippocampus and a clonal cell line. Neuron 1988, 1, 615–621. [Google Scholar]
- Peters, J.A.; Lambert, J.J. Electrophysiology of 5-HT3 receptors in neuronal cell lines. Trends Pharmacol. Sci. 1989, 10, 172–175. [Google Scholar]
- van Hooft, J.A.; Vijverberg, H.P. 5-HT(3) receptors and neurotransmitter release in the CNS: A nerve ending story? Trends Neurosci. 2000, 23, 605–610. [Google Scholar]
- Sugita, S.; Shen, K.Z.; North, R.A. 5-hydroxytryptamine is a fast excitatory transmitter at 5-HT3 receptors in rat amygdala. Neuron 1992, 8, 199–203. [Google Scholar]
- Roerig, B.; Katz, L.C. Modulation of intrinsic circuits by serotonin 5-HT3 receptors in developing ferret visual cortex. J. Neurosci. 1997, 17, 8324–9338. [Google Scholar]
- Barrera, N.P.; Herbert, P.; Henderson, R.M.; Martin, I.L.; Edwardson, J.M. Atomic force microscopy reveals the stoichiometry and subunit arrangement of 5-HT3 receptors. Proc. Natl. Acad. Sci. USA 2005, 102, 12595–12600. [Google Scholar]
- Hapfelmeier, G.; Tredt, C.; Haseneder, R.; Zieglgansberger, W.; Eisensamer, B.; Rupprecht, R.; Rammes, G. Co-expression of the 5-HT3B serotonin receptor subunit alters the biophysics of the 5-HT3 receptor. Biophys. J. 2003, 84, 1720–1733. [Google Scholar]
- Turner, T.J.; Mokler, D.J.; Luebke, J.I. Calcium influx through presynaptic 5-HT3 receptors facilitates GABA release in the hippocampus: In vitro slice and synaptosome studies. Neuroscience 2004, 129, 703–718. [Google Scholar]
- Yakel, J.L.; Shao, X.M.; Jackson, M.B. The selectivity of the channel coupled to the 5-HT3 receptor. Brain Res. 1990, 533, 46–52. [Google Scholar]
- Stewart, A.; Davies, P.A.; Kirkness, E.F.; Safa, P.; Hales, T.G. Introduction of the 5-HT3B subunit alters the functional properties of 5-HT3 receptors native to neuroblastoma cells. Neuropharmacology 2003, 44, 214–223. [Google Scholar]
- Walstab, J.; Rappold, G.; Niesler, B. 5-HT(3) receptors: Role in disease and target of drugs. Pharmacol. Ther. 2010, 128, 146–169. [Google Scholar]
- Campbell, A.D.; Womer, D.E.; Simon, J.R. The 5-HT3 receptor agonist 1-(m-chlorophenyl)-biguanide interacts with the dopamine transporter in rat brain synaptosomes. Eur. J. Pharmacol. 1995, 290, 157–162. [Google Scholar]
- Ismaiel, A.M.; Titeler, M.; Miller, K.J.; Smith, T.S.; Glennon, R.A. 5-HT1 and 5-HT2 binding profiles of the serotonergic agents alpha-methylserotonin and 2-methylserotonin. J. Med. Chem. 1990, 33, 755–758. [Google Scholar]
- Haddjeri, N.; Blier, P. Pre- and post-synaptic effects of the 5-HT3 agonist 2-methyl-5-HT on the 5-HT system in the rat brain. Synapse 1995, 20, 54–67. [Google Scholar]
- Bachy, A.; Heaulme, M.; Giudice, A.; Michaud, J.C.; Lefevre, I.A.; Souilhac, J.; Manara, L.; Emerit, M.B.; Gozlan, H.; Hamon, M.; et al. SR 57227A: a potent and selective agonist at central and peripheral 5-HT3 receptors in vitro and in vivo. Eur. J. Pharmacol. 1993, 237, 299–309. [Google Scholar]
- Herrstedt, J.; Dombernowsky, P. Anti-emetic therapy in cancer chemotherapy: Current status. Basic Clin. Pharmacol. Toxicol. 2007, 101, 143–150. [Google Scholar]
- Greenshaw, A.J.; Silverstone, P.H. The non-antiemetic uses of serotonin 5-HT3 receptor antagonists. Clin. Pharmacol. Ther. Appl. Drugs 1997, 53, 20–39. [Google Scholar]
- Greenshaw, A.J. Behavioural pharmacology of 5-HT3 receptor antagonists: a critical update on therapeutic potential. Trends Pharmacol. Sci. 1993, 14, 265–270. [Google Scholar]
- Martin, P.; Gozlan, H.; Puech, A.J. 5-HT3 receptor antagonists reverse helpless behaviour in rats. Eur. J. Pharmacol. 1992, 212, 73–78. [Google Scholar]
- Ramamoorthy, R.; Radhakrishnan, M.; Borah, M. Antidepressant-like effects of serotonin type-3 antagonist, ondansetron: an investigation in behaviour-based rodent models. Behav. Pharmacol. 2008, 19, 29–40. [Google Scholar]
- Staubli, U.; Xu, F.B. Effects of 5-HT3 receptor antagonism on hippocampal theta rhythm, memory, and LTP induction in the freely moving rat. J. Neurosci. 1995, 15, 2445–2452. [Google Scholar]
- Ilegems, E.; Pick, H.M.; Deluz, C.; Kellenberger, S.; Vogel, H. Noninvasive imaging of 5-HT3 receptor trafficking in live cells: from biosynthesis to endocytosis. J. Biol. Chem. 2004, 279, 53346–53352. [Google Scholar]
- Freeman, S.L.; Glatzle, J.; Robin, C.S.; Valdellon, M.; Sternini, C.; Sharp, J.W.; Raybould, H.E. Ligand-induced 5-HT3 receptor internalization in enteric neurons in rat ileum. Gastroenterology 2006, 131, 97–107. [Google Scholar]
- Rojas, C.; Thomas, A.G.; Alt, J.; Stathis, M.; Zhang, J.; Rubenstein, E.B.; Sebastiani, S.; Cantoreggi, S.; Slusher, B.S. Palonosetron triggers 5-HT(3) receptor internalization and causes prolonged inhibition of receptor function. Eur. J. Pharmacol. 2010, 626, 193–199. [Google Scholar]
- Liu, W.; Thielen, R.J.; McBride, W.J. Effects of repeated daily treatments with a 5-HT3 receptor antagonist on dopamine neurotransmission and functional activity of 5-HT3 receptors within the nucleus accumbens of Wistar rats. Pharmacol. Biochem. Behav. 2006, 84, 370–377. [Google Scholar]
- Adrien, J.; Tissier, M.H.; Lanfumey, L.; Haj-Dahmane, S.; Jolas, T.; Franc, B.; Hamon, M. Central action of 5-HT3 receptor ligands in the regulation of sleep-wakefulness and raphe neuronal activity in the rat. Neuropharmacology 1992, 31, 519–529. [Google Scholar]
- Bagdy, E.; Solyom, S.; Harsing, L.G., Jr. Feedback stimulation of somatodendritic serotonin release: a 5-HT3 receptor-mediated effect in the raphe nuclei of the rat. Brain Res. Bull. 1998, 45, 203–208. [Google Scholar]
- Sanacora, G.; Saricicek, A. GABAergic contributions to the pathophysiology of depression and the mechanism of antidepressant action. CNS Neurol. Disord. Drug Targets 2007, 6, 127–140. [Google Scholar]
- Holm, M.M.; Nieto-Gonzalez, J.L.; Vardya, I.; Henningsen, K.; Jayatissa, M.N.; Wiborg, O.; Jensen, K. Hippocampal GABAergic dysfunction in a rat chronic mild stress model of depression. Hippocampus 2011, 21, 422–423. [Google Scholar]
- Methippara, M.; Bashir, T.; Suntsova, N.; Szymusiak, R.; McGinty, D. Hippocampal adult neurogenesis is enhanced by chronic eszopiclone treatment in rats. J. Sleep Res. 2010, 19, 384–393. [Google Scholar]
- Smiley, J.F.; Goldman-Rakic, P.S. Serotonergic axons in monkey prefrontal cerebral cortex synapse predominantly on interneurons as demonstrated by serial section electron microscopy. J. Comp. Neurol. 1996, 367, 431–443. [Google Scholar]
- Hornung, J.P.; Celio, M.R. The selective innervation by serotoninergic axons of calbindin-containing interneurons in the neocortex and hippocampus of the marmoset. J. Comp. Neurol. 1992, 320, 457–467. [Google Scholar]
- Morales, M.; Battenberg, E.; de Lecea, L.; Bloom, F.E. The type 3 serotonin receptor is expressed in a subpopulation of GABAergic neurons in the rat neocortex and hippocampus. Brain Res. 1996, 731, 199–202. [Google Scholar]
- Mongeau, R.; Blier, P.; de Montigny, C. The serotonergic and noradrenergic systems of the hippocampus: their interactions and the effects of antidepressant treatments. Brain Res. Brain Res. Rev. 1997, 23, 145–195. [Google Scholar]
- Gulyas, A.I.; Acsady, L.; Freund, T.F. Structural basis of the cholinergic and serotonergic modulation of GABAergic neurons in the hippocampus. Neurochem. Int. 1999, 34, 359–372. [Google Scholar]
- Tecott, L.H.; Maricq, A.V.; Julius, D. Nervous system distribution of the serotonin 5-HT3 receptor mRNA. Proc. Natl. Acad. Sci. USA 1993, 90, 1430–1434. [Google Scholar]
- Morales, M.; Bloom, F.E. The 5-HT3 receptor is present in different subpopulations of GABAergic neurons in the rat telencephalon. J. Neurosci. 1997, 17, 3157–3167. [Google Scholar]
- Brady, C.A.; Dover, T.J.; Massoura, A.N.; Princivalle, A.P.; Hope, A.G.; Barnes, N.M. Identification of 5-HT3A and 5-HT3B receptor subunits in human hippocampus. Neuropharmacology 2007, 52, 1284–1290. [Google Scholar]
- Martin, K.F.; Hannon, S.; Phillips, I.; Heal, D.J. Opposing roles for 5-HT1B and 5-HT3 receptors in the control of 5-HT release in rat hippocampus in vivo. Br. J. Pharmacol. 1992, 106, 139–142. [Google Scholar]
- Mørk, A.; Brennum, L.T.; Fallón, S.; Bisulco, S.; Frederiksen, K.; Bang-Andersen, B.; Lassen, A.B.; Zhong, H.; Sanchez, C.; Stensbøl, T.B. P.2.d.017 Pharmacological profile of Lu AA21004, a novel multi-target drug for the treatment of mood disorders. Eur. Neuropsychopharmacol. 2009, 19 Suppl. 3, S439. [Google Scholar]
- Zhang, J.Y.; Zeise, M.L.; Wang, R.Y. Serotonin3 receptor agonists attenuate glutamate-induced firing in rat hippocampal CA1 pyramidal cells. Neuropharmacology 1994, 33, 483–491. [Google Scholar]
- Zeise, M.L.; Batsche, K.; Wang, R.Y. The 5-HT3 receptor agonist 2-methyl-5-HT reduces postsynaptic potentials in rat CA1 pyramidal neurons of the hippocampus in vitro. Brain Res. 1994, 651, 337–341. [Google Scholar]
- Maeda, T.; Kaneko, S.; Satoh, M. Inhibitory influence via 5-HT3 receptors on the induction of LTP in mossy fiber-CA3 system of guinea-pig hippocampal slices. Neurosci. Res. 1994, 18, 277–282. [Google Scholar]
- Reznic, J.; Staubli, U. Effects of 5-HT3 receptor antagonism on hippocampal cellular activity in the freely moving rat. J. Neurophysiol. 1997, 77, 517–521. [Google Scholar]
- Corradetti, R.; Ballerini, L.; Pugliese, A.M.; Pepeu, G. Serotonin blocks the long-term potentiation induced by primed burst stimulation in the CA1 region of rat hippocampal slices. Neuroscience 1992, 46, 511–518. [Google Scholar]
- Passani, M.B.; Pugliese, A.M.; Azzurrini, M.; Corradetti, R. Effects of DAU 6215, a novel 5-hydroxytryptamine3 (5-HT3) antagonist on electrophysiological properties of the rat hippocampus. Br. J. Pharmacol. 1994, 112, 695–703. [Google Scholar]
- Ropert, N.; Guy, N. Serotonin facilitates GABAergic transmission in the CA1 region of rat hippocampus in vitro. J. Physiol. 1991, 441, 121–136. [Google Scholar]
- Piguet, P.; Galvan, M. Transient and long-lasting actions of 5-HT on rat dentate gyrus neurones in vitro. J. Physiol. 1994, 481, 629–639. [Google Scholar]
- Dorostkar, M.M.; Boehm, S. Opposite effects of presynaptic 5-HT3 receptor activation on spontaneous and action potential-evoked GABA release at hippocampal synapses. J. Neurochem. 2007, 100, 395–405. [Google Scholar]
- McMahon, L.L.; Kauer, J.A. Hippocampal interneurons are excited via serotonin-gated ion channels. J. Neurophysiol. 1997, 78, 2493–2502. [Google Scholar]
- Kawa, K. Distribution and functional properties of 5-HT3 receptors in the rat hippocampal dentate gyrus: a patch-clamp study. J. Neurophysiol. 1994, 71, 1935–1947. [Google Scholar]
- Mayberg, H.S.; Brannan, S.K.; Tekell, J.L.; Silva, J.A.; Mahurin, R.K.; McGinnis, S.; Jerabek, P.A. Regional metabolic effects of fluoxetine in major depression: serial changes and relationship to clinical response. Biol. Psychiat. 2000, 48, 830–843. [Google Scholar]
- Ferezou, I.; Cauli, B.; Hill, E.L.; Rossier, J.; Hamel, E.; Lambolez, B. 5-HT3 receptors mediate serotonergic fast synaptic excitation of neocortical vasoactive intestinal peptide/cholecystokinin interneurons. J. Neurosci. 2002, 22, 7389–7397. [Google Scholar]
- Jakab, R.L.; Goldman-Rakic, P.S. Segregation of serotonin 5-HT2A and 5-HT3 receptors in inhibitory circuits of the primate cerebral cortex. J. Comp. Neurol. 2000, 417, 337–348. [Google Scholar]
- Blier, P.; Bouchard, C. Functional characterization of a 5-HT3 receptor which modulates the release of 5-HT in the guinea-pig brain. Br. J. Pharmacol. 1993, 108, 13–22. [Google Scholar]
- Ashby, C.R., Jr.; Edwards, E.; Harkins, K.; Wang, R.Y. Characterization of 5-hydroxytryptamine3 receptors in the medial prefrontal cortex: a microiontophoretic study. Eur. J. Pharmacol. 1989, 173, 193–196. [Google Scholar]
- Ashby, C.R., Jr.; Edwards, E.; Wang, R.Y. Action of serotonin in the medial prefrontal cortex: mediation by serotonin3-like receptors. Synapse 1992, 10, 7–15. [Google Scholar]
- Edwards, E.; Hampton, E.; Ashby, C.R.; Zhang, J.; Wang, R.Y. 5-HT3-like receptors in the rat medial prefrontal cortex: further pharmacological characterization. Brain Res. 1996, 733, 21–30. [Google Scholar]
- Ashby, C.R., Jr.; Minabe, Y.; Edwards, E.; Wang, R.Y. 5-HT3-like receptors in the rat medial prefrontal cortex: an electrophysiological study. Brain Res. 1991, 550, 181–191. [Google Scholar]
- Liang, X.; Arvanov, V.L.; Wang, R.Y. Inhibition of NMDA-receptor mediated response in the rat medial prefrontal cortical pyramidal cells by the 5-HT3 receptor agonist SR 57227A and 5-HT: Intracellular studies. Synapse 1998, 29, 257–268. [Google Scholar]
- Zhou, F.M.; Hablitz, J.J. Activation of serotonin receptors modulates synaptic transmission in rat cerebral cortex. J. Neurophysiol. 1999, 82, 2989–2999. [Google Scholar]
- Dremencov, E.; Weizmann, Y.; Kinor, N.; Gispan-Herman, I.; Yadid, G. Modulation of dopamine transmission by 5HT2C and 5HT3 receptors: a role in the antidepressant response. Curr. Drug Targets 2006, 7, 165–175. [Google Scholar]
- Alex, K.D.; Pehek, E.A. Pharmacologic mechanisms of serotonergic regulation of dopamine neurotransmission. Pharmacol. Ther. 2007, 113, 296–320. [Google Scholar]
- Nestler, E.J.; Carlezon, W.A., Jr. The mesolimbic dopamine reward circuit in depression. Biol. Psychiat. 2006, 59, 1151–1159. [Google Scholar]
- Minabe, Y.; Ashby, C.R., Jr.; Wang, R.Y. The effect of acute and chronic LY 277359, a selective 5-HT3 receptor antagonist, on the number of spontaneously active midbrain dopamine neurons. Eur. J. Pharmacol. 1991, 209, 151–156. [Google Scholar]
- Sorensen, S.M.; Humphreys, T.M.; Palfreyman, M.G. Effect of acute and chronic MDL 73,147EF, a 5-HT3 receptor antagonist, on A9 and A10 dopamine neurons. Eur. J. Pharmacol. 1989, 163, 115–118. [Google Scholar]
- Prisco, S.; Pessia, M.; Ceci, A.; Borsini, F.; Esposito, E. Chronic treatment with DAU 6215, a new 5-HT3 receptor antagonist, causes a selective decrease in the number of spontaneously active dopaminergic neurons in the rat ventral tegmental area. Eur. J. Pharmacol. 1992, 214, 13–19. [Google Scholar]
- King, G.R.; Xue, Z.; Calvi, C.; Ellinwood, E.H., Jr. 5-HT3 agonist-induced dopamine overflow during withdrawal from continuous or intermittent cocaine administration. Psychopharmacology (Berl) 1995, 117, 458–465. [Google Scholar]
- Blandina, P.; Goldfarb, J.; Green, J.P. Activation of a 5-HT3 receptor releases dopamine from rat striatal slice. Eur. J. Pharmacol. 1988, 155, 349–350. [Google Scholar]
- Blandina, P.; Goldfarb, J.; Craddock-Royal, B.; Green, J.P. Release of endogenous dopamine by stimulation of 5-hydroxytryptamine3 receptors in rat striatum. J. Pharmacol. Exp. Ther. 1989, 251, 803–809. [Google Scholar]
- Richter, J.A.; Bare, D.J.; Yu, H.; Ghetti, B.; Simon, J.R. Dopamine transporter-dependent and -independent endogenous dopamine release from weaver mouse striatum in vitro. J. Neurochem. 1995, 64, 191–198. [Google Scholar]
- Benloucif, S.; Keegan, M.J.; Galloway, M.P. Serotonin-facilitated dopamine release in vivo: Pharmacological characterization. J. Pharmacol. Exp. Ther. 1993, 265, 373–377. [Google Scholar]
- Schmidt, C.J.; Black, C.K. The putative 5-HT3 agonist phenylbiguanide induces carrier-mediated release of [3H]dopamine. Eur. J. Pharmacol. 1989, 167, 309–310. [Google Scholar]
- Zazpe, A.; Artaiz, I.; Del Rio, J. Role of 5-HT3 receptors in basal and K(+)-evoked dopamine release from rat olfactory tubercle and striatal slices. Br. J. Pharmacol. 1994, 113, 968–972. [Google Scholar]
- Ng, N.K.; Lee, H.S.; Wong, P.T. Regulation of striatal dopamine release through 5-HT1 and 5-HT2 receptors. J. Neurosci. Res. 1999, 55, 600–607. [Google Scholar]
- Porras, G.; De Deurwaerdere, P.; Moison, D.; Spampinato, U. Conditional involvement of striatal serotonin3 receptors in the control of in vivo dopamine outflow in the rat striatum. Eur. J. Neurosci. 2003, 17, 771–781. [Google Scholar]
- Yi, S.J.; Gifford, A.N.; Johnson, K.M. Effect of cocaine and 5-HT3 receptor antagonists on 5-HT-induced [3H]dopamine release from rat striatal synaptosomes. Eur. J. Pharmacol. 1991, 199, 185–189. [Google Scholar]
- Sarhan, H.; Cloez-Tayarani, I.; Massot, O.; Fillion, M.P.; Fillion, G. 5-HT1B receptors modulate release of [3H]dopamine from rat striatal synaptosomes. Naunyn-Schmied. Arch. Pharmacol. 1999, 359, 40–47. [Google Scholar]
- Koulu, M.; Sjoholm, B.; Lappalainen, J.; Virtanen, R. Effects of acute GR38032F (odansetron), a 5-HT3 receptor antagonist, on dopamine and serotonin metabolism in mesolimbic and nigrostriatal dopaminergic neurons. Eur. J. Pharmacol. 1989, 169, 321–324. [Google Scholar]
- Invernizzi, R.; Pozzi, L.; Samanin, R. Selective reduction of extracellular dopamine in the rat nucleus accumbens following chronic treatment with DAU 6215, a 5-HT3 receptor antagonist. Neuropharmacology 1995, 34, 211–215. [Google Scholar]
- Jacocks, H.M., 3rd; Cox, B.M. Serotonin-stimulated release of [3H]dopamine via reversal of the dopamine transporter in rat striatum and nucleus accumbens: a comparison with release elicited by potassium, N-methyl-D-aspartic acid, glutamic acid and D-amphetamine. J. Pharmacol. Exp. Ther. 1992, 262, 356–364. [Google Scholar]
- De Deurwaerdere, P.; Stinus, L.; Spampinato, U. Opposite change of in vivo dopamine release in the rat nucleus accumbens and striatum that follows electrical stimulation of dorsal raphe nucleus: Role of 5-HT3 receptors. J. Neurosci. 1998, 18, 6528–6538. [Google Scholar]
- Chameau, P.; Inta, D.; Vitalis, T.; Monyer, H.; Wadman, W.J.; van Hooft, J.A. The N-terminal region of reelin regulates postnatal dendritic maturation of cortical pyramidal neurons. Proc. Natl. Acad. Sci. USA 2009, 106, 7227–7232. [Google Scholar]
- Liu, W.; Thielen, R.J.; Rodd, Z.A.; McBride, W.J. Activation of serotonin-3 receptors increases dopamine release within the ventral tegmental area of Wistar and alcohol-preferring (P) rats. Alcohol 2006, 40, 167–176. [Google Scholar]
- Campbell, A.D.; Kohl, R.R.; McBride, W.J. Serotonin-3 receptor and ethanol-stimulated somatodendritic dopamine release. Alcohol 1996, 13, 569–574. [Google Scholar]
- Ashby, C.R., Jr.; Jiang, L.H.; Wang, R.Y. Chronic BRL 43694, a selective 5-HT3 receptor antagonist, fails to alter the number of spontaneously active midbrain dopamine neurons. Eur. J. Pharmacol. 1990, 175, 347–350. [Google Scholar]
- Chenu, F.; El Mansari, M.; Blier, P. Long-term administration of monoamine oxidase inhibitors alters the firing rate and pattern of dopamine neurons in the ventral tegmental area. Int. J. Neuropsychopharmacol. 2009, 12, 475–485. [Google Scholar]
- Campbell, A.D.; McBride, W.J. Serotonin-3 receptor and ethanol-stimulated dopamine release in the nucleus accumbens. Pharmacol. Biochem. Behav. 1995, 51, 835–842. [Google Scholar]
- Jiang, L.H.; Ashby, C.R., Jr.; Kasser, R.J.; Wang, R.Y. The effect of intraventricular administration of the 5-HT3 receptor agonist 2-methylserotonin on the release of dopamine in the nucleus accumbens: an in vivo chronocoulometric study. Brain Res. 1990, 513, 156–160. [Google Scholar]
- Matell, M.S.; King, G.R. 5-HT3 receptor mediated dopamine release in the nucleus accumbens during withdrawal from continuous cocaine. Psychopharmacology (Berl) 1997, 130, 242–248. [Google Scholar]
- Chen, J.P.; van Praag, H.M.; Gardner, E.L. Activation of 5-HT3 receptor by 1-phenylbiguanide increases dopamine release in the rat nucleus accumbens. Brain Res. 1991, 543, 354–357. [Google Scholar]
- Allan, A.M.; Galindo, R.; Chynoweth, J.; Engel, S.R.; Savage, D.D. Conditioned place preference for cocaine is attenuated in mice over-expressing the 5-HT(3) receptor. Psychopharmacology (Berl) 2001, 158, 18–27. [Google Scholar]
- Carboni, E.; Acquas, E.; Frau, R.; Di Chiara, G. Differential inhibitory effects of a 5-HT3 antagonist on drug-induced stimulation of dopamine release. Eur. J. Pharmacol. 1989, 164, 515–519. [Google Scholar]
- Stefanski, R.; Palejko, W.; Bidzinski, A.; Kostowski, W.; Plaznik, A. Serotonergic innervation of the hippocampus and nucleus accumbens septi and the anxiolytic-like action of the 5-HT3 receptor antagonists. Neuropharmacology 1993, 32, 987–993. [Google Scholar]
- Costall, B.; Kelly, M.E.; Naylor, R.J.; Onaivi, E.S.; Tyers, M.B. Neuroanatomical sites of action of 5-HT3 receptor agonist and antagonists for alteration of aversive behaviour in the mouse. Br. J. Pharmacol. 1989, 96, 325–332. [Google Scholar]
- Rasmussen, K.; Stockton, M.E.; Czachura, J.F. The 5-HT3 receptor antagonist zatosetron decreases the number of spontaneously active A10 dopamine neurons. Eur. J. Pharmacol. 1991, 205, 113–116. [Google Scholar]
- Ashworth-Preece, M.A.; Jarrott, B.; Lawrence, A.J. 5-Hydroxytryptamine3 receptor modulation of excitatory amino acid release in the rat nucleus tractus solitarius. Neurosci. Lett. 1995, 191, 75–78. [Google Scholar]
- Funahashi, M.; Mitoh, Y.; Matsuo, R. Activation of presynaptic 5-HT3 receptors facilitates glutamatergic synaptic inputs to area postrema neurons in rat brain slices. Methods Find. Exp. Clin. Pharmacol. 2004, 26, 615–622. [Google Scholar]
- Monti, J.M.; Jantos, H. Activation of the serotonin 5-HT3 receptor in the dorsal raphe nucleus suppresses REM sleep in the rat. Prog. Neuropsychopharmacol. Biol. Psychiat. 2008, 32, 940–947. [Google Scholar]
- Diez-Ariza, M.; Garcia-Alloza, M.; Lasheras, B.; Del Rio, J.; Ramirez, M.J. GABA(A) receptor antagonists enhance cortical acetylcholine release induced by 5-HT(3) receptor blockade in freely moving rats. Brain Res. 2002, 956, 81–85. [Google Scholar]
- Fan, P. Effects of antidepressants on the inward current mediated by 5-HT3 receptors in rat nodose ganglion neurones. Br. J. Pharmacol. 1994, 112, 741–744. [Google Scholar]
- Fan, P. Inhibition of a 5-HT3 receptor-mediated current by the selective serotonin uptake inhibitor, fluoxetine. Neurosci. Lett. 1994, 173, 210–212. [Google Scholar]
- Choi, J.S.; Choi, B.H.; Ahn, H.S.; Kim, M.J.; Rhie, D.J.; Yoon, S.H.; Min, D.S.; Jo, Y.H.; Kim, M.S.; Sungm, K.W.; et al. Mechanism of block by fluoxetine of 5-hydroxytryptamine3 (5-HT3)-mediated currents in NCB-20 neuroblastoma cells. Biochem. Pharmacol. 2003, 66, 2125–2132. [Google Scholar]
- Eisensamer, B.; Rammes, G.; Gimpl, G.; Shapa, M.; Ferrari, U.; Hapfelmeier, G.; Bondy, B.; Parsons, C.; Gilling, K.; Zieglgansberger, W.; et al. Antidepressants are functional antagonists at the serotonin type 3 (5-HT3) receptor. Mol. Psychiat. 2003, 8, 994–1007. [Google Scholar]
- Breitinger, H.G.; Geetha, N.; Hess, G.P. Inhibition of the serotonin 5-HT3 receptor by nicotine, cocaine, and fluoxetine investigated by rapid chemical kinetic techniques. Biochemistry 2001, 40, 8419–8429. [Google Scholar]
- Lucchelli, A.; Santagostino-Barbone, M.G.; Barbieri, A.; Candura, S.M.; Tonini, M. The interaction of antidepressant drugs with central and peripheral (enteric) 5-HT3 and 5-HT4 receptors. Br. J. Pharmacol. 1995, 114, 1017–1025. [Google Scholar]
- Varea, E.; Blasco-Ibáñez, J.M.; Gómez-Climent, M.Á.; Castillo-Gómez, E.; Crespo, C.; Martínez-Guijarro, F.J.; Nácher, J. Chronic fluoxetine treatment increases the expression of PSA-NCAM in the medial prefrontal cortex. Neuropsychopharmacology 2007, 32, 803–812. [Google Scholar]
- Kent, J.M. SNaRIs, NaSSAs, and NaRIs: New agents for the treatment of depression. Lancet 2000, 355, 911–918. [Google Scholar]
- Rammes, G.; Eisensamer, B.; Ferrari, U.; Shapa, M.; Gimpl, G.; Gilling, K.; Parsons, C.; Riering, K.; Hapfelmeier, G.; Bondy, B.; et al. Antipsychotic drugs antagonize human serotonin type 3 receptor currents in a noncompetitive manner. Mol. Psychiat. 2004, 9, 846–858. [Google Scholar]
- Eisensamer, B.; Uhr, M.; Meyr, S.; Gimpl, G.; Deiml, T.; Rammes, G.; Lambert, J.J.; Zieglgansberger, W.; Holsboer, F.; Rupprecht, R. Antidepressants and antipsychotic drugs colocalize with 5-HT3 receptors in raft-like domains. J. Neurosci. 2005, 25, 10198–10206. [Google Scholar]
- Nothdurfter, C.; Tanasic, S.; Di Benedetto, B.; Rammes, G.; Wagner, E.M.; Kirmeier, T.; Ganal, V.; Kessler, J.S.; Rein, T.; Holsboer, F.; et al. Impact of lipid raft integrity on 5-HT3 receptor function and its modulation by antidepressants. Neuropsychopharmacology 2010, 35, 1510–1519. [Google Scholar]
- Besson, A.; Haddjeri, N.; Blier, P.; de Montigny, C. Effects of the co-administration of mirtazapine and paroxetine on serotonergic neurotransmission in the rat brain. Eur. Neuropsychopharmacol. 2000, 10, 177–188. [Google Scholar]
- Bourin, M.; Dailly, E.; Hascoet, M. Preclinical and clinical pharmacology of cyamemazine: anxiolytic effects and prevention of alcohol and benzodiazepine withdrawal syndrome. CNS Drug Rev. 2004, 10, 219–229. [Google Scholar]
- Ishihara, K.; Sasa, M. Potentiation of 5-HT(3) receptor functions in the hippocampal CA1 region of rats following repeated electroconvulsive shock treatments. Neurosci. Lett. 2001, 307, 37–40. [Google Scholar]
- Moore, N.; Bang-Andersen, B.; Brennum, L.; Fredriksen, K.; Hogg, S.; Mork, A.; Stensbol, T.; Zhong, H.; Sanchez, C.; Smith, D. Lu AA21004: a novel potential treatment for mood disorders. Eur. Neuropsychopharmacol. 2008, 18, S321. [Google Scholar]
- Haddjeri, N.; Etievant, A.; Moore, N.; Miller, S.; Sánchez, C.; Betry, C. P.2.d.013 Electrophysiological study of the effects of the novel antidepressant Lu AA21004 on the rat 5-HT neuronal activity. Eur. Neuropsychopharmacol. 2009, 19 Suppl.3, S437. [Google Scholar]
- Artigas, F.; Dragheim, M.; Loft, H.; Perez, V.; Álvarez, E. P.2.c.040 A randomised, double-blind, placebo-controlled, active-referenced study of Lu AA21004 in patients with major depression. Eur. Neuropsychopharmacol. 2009, 19 Suppl.3, S426. [Google Scholar]
- Poncelet, M.; Perio, A.; Simiand, J.; Gout, G.; Soubrie, P.; Le Fur, G. Antidepressant-like effects of SR 57227A, a 5-HT3 receptor agonist, in rodents. J. Neural. Transm. Gen. Sect. 1995, 102, 83–90. [Google Scholar]
- Redrobe, J.P.; Bourin, M. Partial role of 5-HT2 and 5-HT3 receptors in the activity of antidepressants in the mouse forced swimming test. Eur. J. Pharmacol. 1997, 325, 129–135. [Google Scholar]
- Nakagawa, Y.; Ishima, T.; Takashima, T. The 5-HT3 receptor agonist attenuates the action of antidepressants in the forced swim test in rats. Brain Res. 1998, 786, 189–193. [Google Scholar]
- Bravo, G.; Maswood, S. Acute treatment with 5-HT3 receptor antagonist, tropisetron, reduces immobility in intact female rats exposed to the forced swim test. Pharmacol. Biochem. Behav. 2006, 85, 362–368. [Google Scholar]
- Kos, T.; Popik, P.; Pietraszek, M.; Schafer, D.; Danysz, W.; Dravolina, O.; Blokhina, E.; Galankin, T.; Bespalov, A.Y. Effect of 5-HT3 receptor antagonist MDL 72222 on behaviors induced by ketamine in rats and mice. Eur. Neuropsychopharmacol. 2006, 16, 297–310. [Google Scholar]
- Bourin, M.; Hascoet, M.; Colombel, M.C.; Redrobe, J.P.; Baker, G.B. Differential effects of clonidine, lithium and quinine in the forced swimming test in mice for antidepressants: possible roles of serotoninergic systems. Eur. Neuropsychopharmacol. 1996, 6, 231–236. [Google Scholar]
- Mahesh, R.; Rajkumar, R.; Minasri, B.; Venkatesha Perumal, R. Potential antidepressants: pharmacology of 2-(4-methyl piperazin-1-yl)-1,8-naphthyridine-3-carbonitrile in rodent behavioural models. Pharmazie 2007, 62, 919–924. [Google Scholar]
- Devadoss, T.; Pandey, D.K.; Mahesh, R.; Yadav, S.K. Effect of acute and chronic treatment with QCF-3 (4-benzylpiperazin-1-yl) (quinoxalin-2-yl) methanone, a novel 5-HT(3) receptor antagonist, in animal models of depression. Pharmacol. Rep. 2010, 62, 245–257. [Google Scholar]
- Olivier, B.; van Wijngaarden, I.; Soudijn, W. 5-HT(3) receptor antagonists and anxiety; a preclinical and clinical review. Eur. Neuropsychopharmacol. 2000, 10, 77–95. [Google Scholar]
- Griebel, G. 5-Hydroxytryptamine-interacting drugs in animal models of anxiety disorders: More than 30 years of research. Pharmacol. Ther. 1995, 65, 319–395. [Google Scholar]
- Stefanski, R.; Palejko, W.; Kostowski, W.; Plaznik, A. The comparison of benzodiazepine derivatives and serotonergic agonists and antagonists in two animal models of anxiety. Neuropharmacology 1992, 31, 1251–1258. [Google Scholar]
- Filip, M.; Baran, L.; Siwanowicz, J.; Chojnacka-Wojcik, E.; Przegalinski, E. The anxiolytic-like effects of 5-hydroxytryptamine3 (5-HT3) receptor antagonists. Pol. J. Pharmacol. Pharm. 1992, 44, 261–269. [Google Scholar]
- Jones, B.J.; Costall, B.; Domeney, A.M.; Kelly, M.E.; Naylor, R.J.; Oakley, N.R.; Tyers, M.B. The potential anxiolytic activity of GR38032F, a 5-HT3-receptor antagonist. Br. J. Pharmacol. 1988, 93, 985–993. [Google Scholar]
- Andrews, N.; File, S.E. Are there changes in sensitivity to 5-HT3 receptor ligands following chronic diazepam treatment? Psychopharmacology (Berl) 1992, 108, 333–337. [Google Scholar]
- Rotzinger, S.; Lovejoy, D.A.; Tan, L.A. Behavioral effects of neuropeptides in rodent models of depression and anxiety. Peptides 2010, 31, 736–756. [Google Scholar]
- Raiteri, M.; Paudice, P.; Vallebuona, F. Inhibition by 5-HT3 receptor antagonists of release of cholecystokinin-like immunoreactivity from the frontal cortex of freely moving rats. Naunyn-Schmied. Arch. Pharmacol. 1993, 347, 111–114. [Google Scholar]
- Paudice, P.; Raiteri, M. Cholecystokinin release mediated by 5-HT3 receptors in rat cerebral cortex and nucleus accumbens. Br. J. Pharmacol. 1991, 103, 1790–1794. [Google Scholar]
- Nevo, I.; Becker, C.; Hamon, M.; Benoliel, J.J. Stress- and yohimbine-induced release of cholecystokinin in the frontal cortex of the freely moving rat: prevention by diazepam but not ondansetron. J. Neurochem. 1996, 66, 2041–2049. [Google Scholar]
- Ninan, P.T. The functional anatomy, neurochemistry, and pharmacology of anxiety. J. Clin. Psychiat. 1999, 60 Suppl. 22, 12–17. [Google Scholar]
- Higgins, G.A.; Jones, B.J.; Oakley, N.R.; Tyers, M.B. Evidence that the amygdala is involved in the disinhibitory effects of 5-HT3 receptor antagonists. Psychopharmacology (Berl) 1991, 104, 545–551. [Google Scholar]
- Bhatnagar, S.; Sun, L.M.; Raber, J.; Maren, S.; Julius, D.; Dallman, M.F. Changes in anxiety-related behaviors and hypothalamic-pituitary-adrenal activity in mice lacking the 5-HT-3A receptor. Physiol. Behav. 2004, 81, 545–555. [Google Scholar]
- Kelley, S.P.; Bratt, A.M.; Hodge, C.W. Targeted gene deletion of the 5-HT3A receptor subunit produces an anxiolytic phenotype in mice. Eur. J. Pharmacol. 2003, 461, 19–25. [Google Scholar]
- Bhatnagar, S.; Nowak, N.; Babich, L.; Bok, L. Deletion of the 5-HT3 receptor differentially affects behavior of males and females in the Porsolt forced swim and defensive withdrawal tests. Behav. Brain Res. 2004, 153, 527–535. [Google Scholar]
- Harrell, A.V.; Allan, A.M. Improvements in hippocampal-dependent learning and decremental attention in 5-HT(3) receptor overexpressing mice. Learn. Mem. 2003, 10, 410–419. [Google Scholar]
- Piche, T.; Vanbiervliet, G.; Cherikh, F.; Antoun, Z.; Huet, P.M.; Gelsi, E.; Demarquay, J.F.; Caroli-Bosc, F.X.; Benzaken, S.; Rigault, M.C.; et al. Effect of ondansetron, a 5-HT3 receptor antagonist, on fatigue in chronic hepatitis C: A randomised, double blind, placebo controlled study. Gut 2005, 54, 1169–1173. [Google Scholar]
- Johnson, B.A.; Ait-Daoud, N.; Ma, J.Z.; Wang, Y. Ondansetron reduces mood disturbance among biologically predisposed, alcohol-dependent individuals. Alcohol. Clin. Exp. Res. 2003, 27, 1773–1779. [Google Scholar]
- Faris, P.L.; Eckert, E.D.; Kim, S.W.; Meller, W.H.; Pardo, J.V.; Goodale, R.L.; Hartman, B.K. Evidence for a vagal pathophysiology for bulimia nervosa and the accompanying depressive symptoms. J. Affect. Disord. 2006, 92, 79–90. [Google Scholar]
- Haus, U.; Varga, B.; Stratz, T.; Spath, M.; Muller, W. Oral treatment of fibromyalgia with tropisetron given over 28 days: influence on functional and vegetative symptoms, psychometric parameters and pain. Scand. J. Rheumatol. Suppl. 2000, 113, 55–58. [Google Scholar]
- Spath, M.; Welzel, D.; Farber, L. Treatment of chronic fatigue syndrome with 5-HT3 receptor antagonists—preliminary results. Scand. J. Rheumatol. Suppl. 2000, 113, 72–77. [Google Scholar]
- The, G.K.; Prins, J.; Bleijenberg, G.; van der Meer, J.W. The effect of granisetron, a 5-HT3 receptor antagonist, in the treatment of chronic fatigue syndrome patients—A pilot study. Neth. J. Med. 2003, 61, 285–289. [Google Scholar]
- Spath, M. Current experience with 5-HT3 receptor antagonists in fibromyalgia. Rheum. Dis. Clin. North Am. 2002, 28, 319–328. [Google Scholar]
- The, G.K.; Bleijenberg, G.; Buitelaar, J.K.; van der Meer, J.W. The effect of ondansetron, a 5-HT3 receptor antagonist, in chronic fatigue syndrome: A randomized controlled trial. J. Clin. Psychiat. 2010, 71, 528–533. [Google Scholar]
- Turkel, S.B.; Nadala, J.G.; Wincor, M.Z. Possible serotonin syndrome in association with 5-HT(3) antagonist agents. Psychosomatics 2001, 42, 258–260. [Google Scholar]
- Harmer, C.J.; Reid, C.B.; Ray, M.K.; Goodwin, G.M.; Cowen, P.J. 5HT(3) antagonism abolishes the emotion potentiated startle effect in humans. Psychopharmacology (Berl) 2006, 186, 18–24. [Google Scholar]
- Smith, W.T.; Londborg, P.D.; Blomgren, S.L.; Tollefson, G.D.; Sayler, M.E. Pilot study of zatosetron (LY277359) maleate, a 5-hydroxytryptamine-3 antagonist, in the treatment of anxiety. J. Clin. Psychopharmacol. 1999, 19, 125–131. [Google Scholar]
- Lecrubier, Y.; Puech, A.J.; Azcona, A.; Bailey, P.E.; Lataste, X. A randomized double-blind placebo-controlled study of tropisetron in the treatment of outpatients with generalized anxiety disorder. Psychopharmacology (Berl) 1993, 112, 129–133. [Google Scholar]
- McCann, U.D.; Morgan, C.M.; Geraci, M.; Slate, S.O.; Murphy, D.L.; Post, R.M. Effects of the 5-HT3 antagonist, ondansetron, on the behavioral and physiological effects of pentagastrin in patients with panic disorder and social phobia. Neuropsychopharmacology 1997, 17, 360–369. [Google Scholar]
- Depot, M.; Caille, G.; Mukherjee, J.; Katzman, M.A.; Cadieux, A.; Bradwejn, J. Acute and chronic role of 5-HT3 neuronal system on behavioral and neuroendocrine changes induced by intravenous cholecystokinin tetrapeptide administration in humans. Neuropsychopharmacology 1999, 20, 177–187. [Google Scholar]
- Walstab, J.; Rappold, G.; Niesler, B. 5-HT(3) receptors: Role in disease and target of drugs. Pharmacol. Ther. 2010, 128, 146–169. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bétry, C.; Etiévant, A.; Oosterhof, C.; Ebert, B.; Sanchez, C.; Haddjeri, N. Role of 5-HT3 Receptors in the Antidepressant Response. Pharmaceuticals 2011, 4, 603-629. https://doi.org/10.3390/ph4040603
Bétry C, Etiévant A, Oosterhof C, Ebert B, Sanchez C, Haddjeri N. Role of 5-HT3 Receptors in the Antidepressant Response. Pharmaceuticals. 2011; 4(4):603-629. https://doi.org/10.3390/ph4040603
Chicago/Turabian StyleBétry, Cécile, Adeline Etiévant, Chris Oosterhof, Bjarke Ebert, Connie Sanchez, and Nasser Haddjeri. 2011. "Role of 5-HT3 Receptors in the Antidepressant Response" Pharmaceuticals 4, no. 4: 603-629. https://doi.org/10.3390/ph4040603