Reassembled Biosynthetic Pathway for a Large-scale Synthesis of CMP-Neu5Ac
by
Jing Song
1,
Hesheng Zhang
2,
Bingyuan Wu
2,
Yingxin Zhang
2,
Hanfen Li
2,
Min Xiao
1 and
Peng George Wang
1,2,* 
1
State Key Laboratory of Microbial Technology, College of Life Sciences, Shandong University, Jinan 250100, Shandong Province, P. R. China
2
Department of Biochemistry, College of Biological Sciences, The Ohio State University, Columbus, Ohio 43210, USA
*
Author to whom correspondence should be addressed.
Mar. Drugs 2003, 1(1), 34-45; https://doi.org/10.3390/md101034
Submission received: 10 October 2003
/
Accepted: 13 November 2003
/
Published: 26 November 2003
Abstract
:CMP-Neu5Ac is an important sugar nucleotide for biosynthesis of sialic acid and its conjugates. In this paper, a large-scale production system of CMP-Neu5Ac by a single strain is reported. The co-expression of Neu5Ac aldolase (EC4.1.3.3) and CMP-Neu5Ac synthetase (EC 2.7.7.43) was achieved by constructing individual genes into one plasmid and having a single culture that has both NeuAc aldolase and CMP-Neu5Ac synthetase activities. Overall this system only employed N-acetylmannosamine, excess of pyruvate and CTP to produce CMP-Neu5Ac. This work has demonstrated that a large-scale synthesis of sialic acid-derived oligosaccharides could be achieved economically and efficiently through a single, biosynthetic pathway engineered microorganism.
Introduction
It is well known that sialic acids (N-acetyl neuraminic acid) position within glycoconjugates, and it has become increasingly apparent that sialic acids are intimately involved in a number of important physiological phenomena and disease states, and they play essential roles in numerous biological processes such as cell recognition, viral infection, and toxin binding [1–3]. Generally, CMP-Neu5Ac (cytidine 5′-monophospho-N-acetyl neuraminic acid), the activated form of Neu5Ac, serves as the common activated sugar donor for all the sialyltransferases to transfer a single sialic acid to an acceptor or a growing oligosaccharide chain [4,5]. Thus, there is an increasing demand for its large scale production [6].
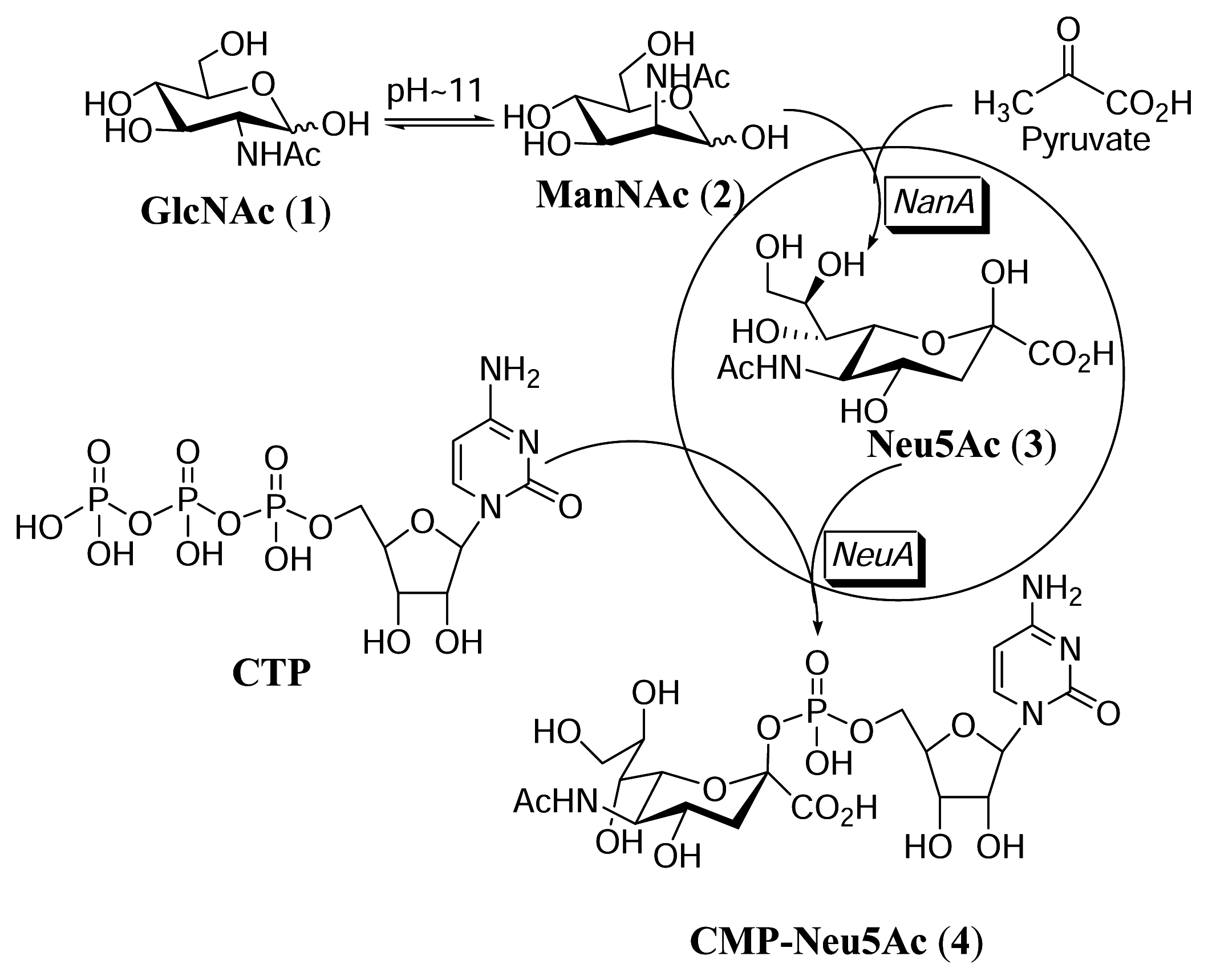
The synthesis of CMP-Neu5Ac is primarily catalyzed by the CMP-Neu5Ac synthetase, an enzyme that uses CTP (cytidine triphosphate) as energy source to condense with sialic acid and give rise to the activated form of Neu5Ac (Figure 1.). Neu5Ac aldolase (EC4.1.3.3) catalyzes the reversible cleavage of sialic acid to pyruvate and ManNAc [7]. CMP-Neu5Ac synthetase (EC 2.7.7.43) catalyzes the formation of CMP-Neu5Ac from Neu5Ac and CTP [8]. In the past, most of the enzymes used were from animal tissues, which contain only low levels of the synthetase activity and this made it difficult to provide CMP-Neu5Ac in large quantities with high quality. The first microbial enzyme used for the preparation of activated NeuAc was the CMP-Neu5Ac synthetase from Neisseria meningitides, but the specific activity of the synthetase in the cell extract was rather low [9,10]. More recently, the genes for CMP-Neu5Ac synthetase from E. coli strains were cloned and overexpressed in E. coli [11–13]. Synthesis of CMP-Neu5Ac through bacterial coupling has been previously reported [6]. Inspired by the efficient and cost effective approach in the production of sugar nucleotides in the whole cell reaction developed by Kyowa Hakko, our group has recently developed a “superbug” strategy to produce oligosaccharides in large quantities [14,15]. The “superbug” reaction can be highly productive and cost effective through conventional fermentation by using cheap starting materials such as monosaccharides.
As reported herein, we assembled the genes of these two enzymes (NanA, NeuA) to form an artificial gene cluster onto a single plasmid (Figure 2.). Subsequent transformation of the plasmid into an E.coli strain affords a carbohydrate-producing bacterial strain. Such a strain can be fermented in large quantities and used for the synthesis of the sialylated oligosaccharides from inexpensive starting materials.
Results and Discussion
Co-expression and characterization of recombinant enzymes
The Neu5Ac aldolase gene (nanA) and CMP-Neu5Ac synthetase gene (neuA) were amplified from the chromosomal DNA of E. coli K12 and K1, respectively. The two DNA fragments obtained were cloned tandemly downstream of the T7 promoter, each with its own ribosome-binding site. The recombinant plasmid was used to transform E. coli BL21 (DE3) and the transformant was used for overexpressing NanA and NeuA. Sodium dodecylsulfate polyacrylamide gel electrophoresis (SDS-PAGE) indicated that recombinant enzymes were overexpressed in the host strain with the target protein representing more than 70% of the total soluble protein and the coexpression had no adverse effect on the levels of soluble protein (data not shown). Neu5Ac aldolase (EC4.1.3.3) from E. coli is stable between pH 6.0 to 9.0 with pH optimum around 7.7, thermostable up to 60°C. The gene of CMP-Neu5Ac synthetase (EC 2.7.7.43) has been cloned from E. coli K1 [10], Neisseria meningitides [11], Haemophilus ducreyi [12] as well as murine pituitary cell [8]. The enzyme purified from E. coli K1 has a requirement for Mg2+ or Mn2+, and exhibits optimal activity between pH 9.0 and 10 in the presence of 30 mM MgCl2, or is the most active near pH 7.5 in the presence of 5 mM MnCl2. The apparent Km for CTP and NeuAc are 0.31 mM and 4 mM, respectively. After induction, the enzyme activity of NeuA aldolase was obtained at a level of 4000 units/L and that of CMP-NeuAc synthetase was at a level of 200 units/L. The activities of the co-expression are comparable to those of the individual enzymes.
Isomerization of GlcNAc to ManNAc
The expensive ManNAc can be prepared practically from GlcNAc by epimerization of the N-acetylamino group at C-2. The epimerization can be achieved by using GlcNAc 2-epimerase (EC 5.1.3.8) and a continuous process for synthesis of NeuAc can be established by immobilizing enzymes in the membrane reactor [17]. Epimerase enzyme, however, is not easily available, which limits its application to large-scale production of NeuAc. Preparation of ManNAc from GlcNAc can also be accomplished by alkaline epimerization [18,19]. A number of processes have been described which provide isomerization of GlcNAc to ManNAc on a multigram scale using sodium hydroxide. Although GlcNAc dominates in the equilibrium mixture (the ratio of ManNAc to GlcNAc is approximately 1/4 ~ 1/3), ManNAc could be extracted into hot-ethanol to yield a ManNAc-enriched solution (about 80% to 88% pure). Such crude ManNAc can be used directly in the aldolase reaction, and the recovered GlcNAc could be recycled. In the first stage, epimerization of GlcNAc to ManNAc was achieved with an aqueous base at about pH~11. The equilibrium was reached after the mixture was allowed to stand for 2 days. To avoid using a large amount of hot ethanol due to the modest solubility of ManNAc, extraction with methanol was used to effectively enrich the ManNAc mixture (Table 1) [20].
After evaporation, the dry ManNAc/GlcNAc mixture was directly added to the enzyme reaction buffer. Typically ManNAc can be enriched in the range of about 75–80% in the mixture. The procedure was repeated using recovered GlcNAc to provide total 11.24 g of a crude solid mixture containing ~75% of ManNAc, and the filtrate was concentrated under reduced pressure to provide a ManNAc-enriched mixture (GlcNAc/ManNAc = 1:4), according to analysis by 1H-NMR spectroscopy (data not shown).
Synthesis of CMP-Neu5Ac in the whole cell reaction
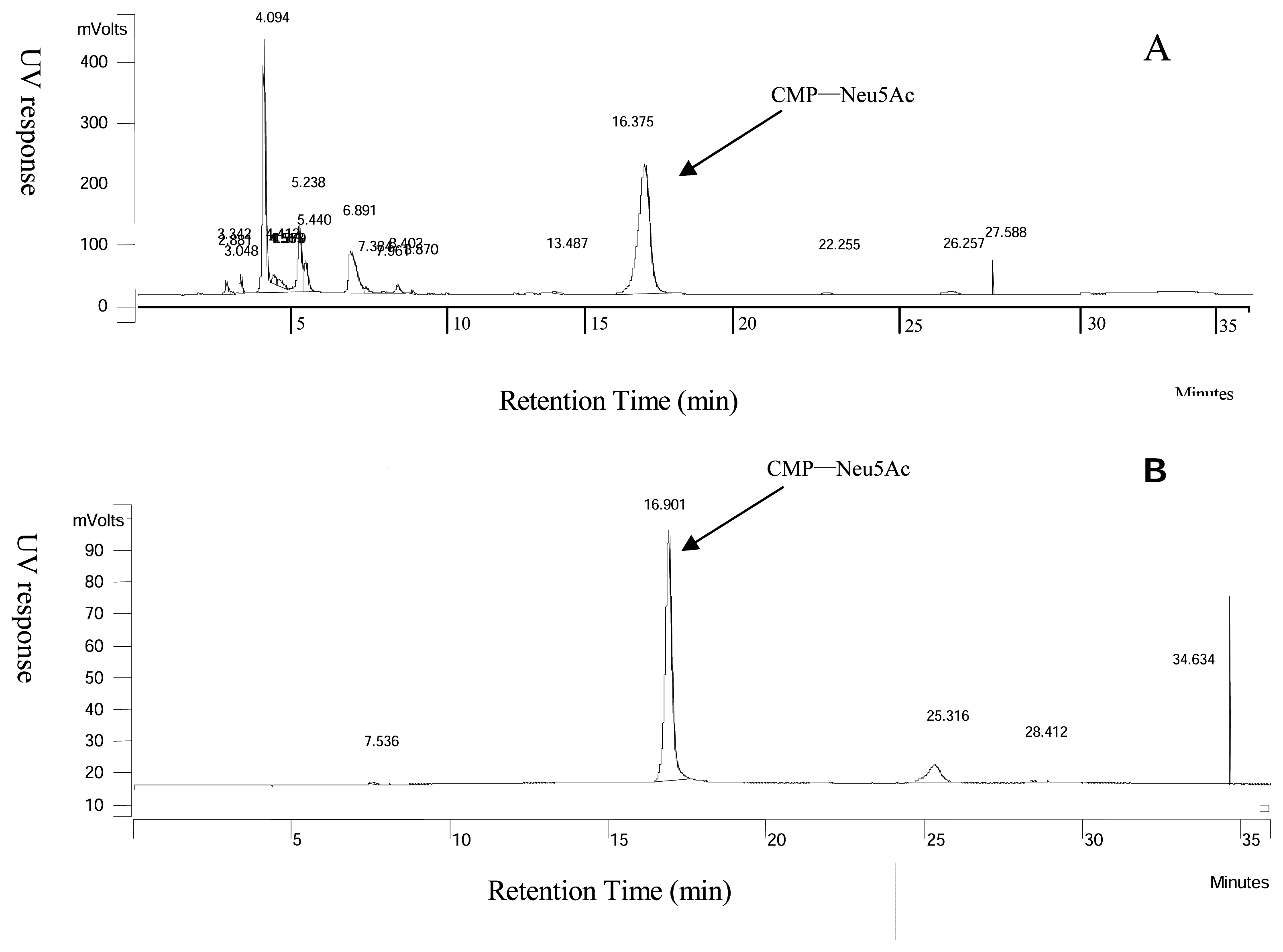
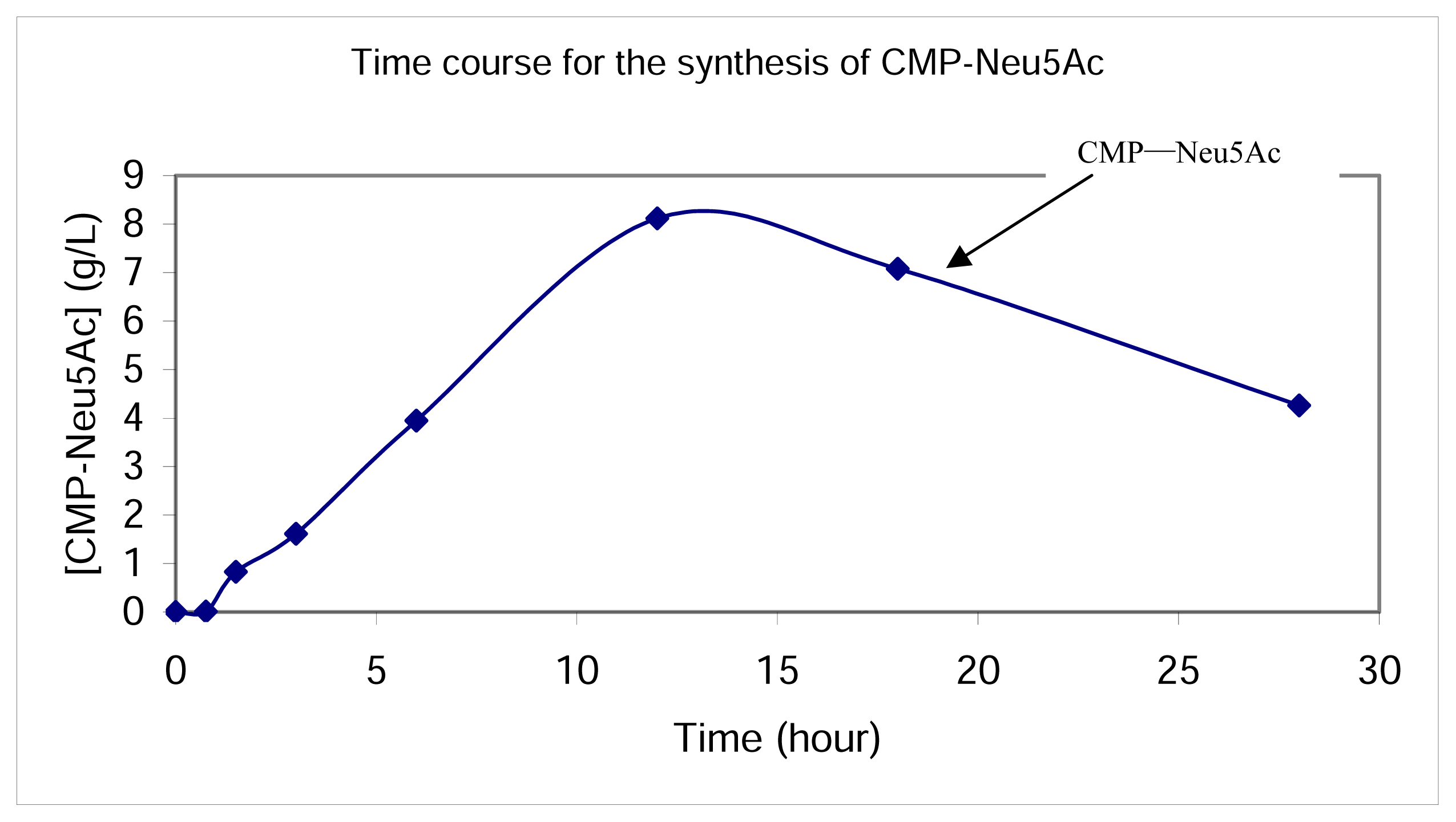
The synthesis of CMP-Neu5Ac from ManNAc, pyruvate and CTP with the fusion protein was performed in the whole cell reactions at pH 7.5, in the presence of 10 mM MnCl2. The formation of the sugar nucleotide was monitored by TLC (eluant: n-BuOH/AcOH/H2O = 5:3:2) as well as by HPLC with UV detection (Figure 3A). Time course studies, as shown in Figure 4, indicated that the accumulation of CMP-Neu5Ac reached its peak at around 12 h in the whole cell reaction catalyzed by the fusion protein that has both NeuAc aldolase and CMP-Neu5Ac synthetase activities, and it was accumulated to 13.22 mM (8.12 g/L) in the reaction mixture. When it was exposed in the reaction mixture after longer time, however, CMP-Neu5Ac seemed to be decomposed since the concentration decreased.
Analysis of CMP-Neu5Ac
CMP-Neu5Ac was analyzed by means of high-performance liquid chromatography with UV/Vis detection (Varian Prostar 320) using a Varian system equipped with a Microsorb 100-NH2 column, (250 × 4.6 mm). The concentration was measured at a flow rate of 0.75 mL/min, elution was performed with a linear gradient of 20 mM phosphate buffer (pH 6.2, composed of 16.16 mL of 1 M KH2PO4 and 3.84 mL of 1M K2HPO4) at increasing concentration from 6 mM to 20 mM, and of acetonitrile at decreasing concentration from 70% to 0% v/v, in a time period of 30 min, at injection volume of 20 μL, and UV detection at 272 nm.
Purification of CMP-Neu5Ac
Purification of the CMP-Neu5Ac from CTP, CMP, pyruvate, phosphate, inorganic salts and the mass of the bacteria cells was conducted by ion-exchange chromatography using the formate form of anion-exchange resin (Dowex 1×8) [21]. The column was eluted with a gradient of aqueous ammonium bicarbonate. The use of ammonium bicarbonate prevented hydrolysis of the extremely acid-labile CMP-Neu5Ac and provided it as the ammonium salt. Excess ammonium bicarbonate was easily removed by passing gel filtration column of the Bio-Gel P-2. Fractions containing CMP-Neu5Ac were identified by TLC analysis and collected. The purity of the pooled collections was analyzed by HPLC (Figure 3B) and the final product was dried by lyophilization.
Conclusions
Biocatalytic approaches in which isolated enzymes, especially Leloir glycosyltransferases, are used are powerful and complementary alternatives to chemical synthesis of carbohydrates and glycoconjugates [22,23]. So far, a large number of mammalian glycosyltransferases have been cloned and employed in oligosaccharide synthesis; these enzymes include bovine α1,3-galactosyltransferase [24]and human α 2,3-sialyltransferase [25] and α1,3-fucosyltransferase [26]. However, the utility of these enzymes in large-scale synthesis of glycoconjugates is partially limited by the high cost of eukaryotic cell culture and by the low level of protein expression. Most bacterial glycosyltransferases, on the other hand, are easily expressed at high levels in cheaper prokaryotic expression systems. Moreover, bacterial glycosyltransferases seem to have broader acceptor substrate specificities, thereby offering a tremendous advantage over mammalian enzymes in chemoenzymatic synthesis of oligosaccharides and their derivatives. Nowdays availability of more and more new glycosyltransferases has increased the demand for sugar nucleotides in the production of oligosaccharides and glycoconjugates. Enzymatic synthesis of sugar nucleotides is more attractive than chemical methods because the yields are high and no organic solvents are used. CMP-Neu5Ac is synthesized in vivo by reaction of CTP and sialic acid in the presence of CMP-Neu5Ac synthetase. Endo et al has reported a large-scale production system of CMP-Neu5Ac by two recombinant E.Coli strains that overexpressed the genes of CMP-Neu5Ac synthetase and CTP synthetase and corynebacterium .ammoniagenes using Neu5Ac, orotic acid and fructose as starting materials [6]. Our approach here is the coexpression of two enzymes involved in the CMP-Neu5Ac biosynthetic pathway in one microbial strain to modify the host cells towards the accumulation of the desired product. Compared to the Kyowa Hakko technology which involves several plasmids and multiple bacterial strains, our approach achieves the same goal with the use of only one plasmid and a single strain. All the enzymes essential for CMP-Neu5Ac are in one E.Coli strain. CMP-Neu5Ac can be produced on a large-scale through an artificial bacterial strain which has both Neu5Ac aldolase and CMP-Neu5Ac synthetase activities by using CTP, sodium pyruvate and ManNAc as starting materials. This artificial cluster, consisting of only two genes, together with inexpensive starting material GlcNAc make this approach more attractive. The simplicity of the synthetic cycle, and thus that of the plasmid, allows for further modification, such as the addition or replacement of genes. Work is in progress to incorporate a α 2,3-sialyltransferase gene into pET-NII to form plasmids consisting of three genes for the synthesis of sialylated oligosaccharides starting from N-acetylglucosamine.
In summary, this paper describes the integrated, efficient and economical system for the large-scale production of CMP-Neu5Ac through introducing an artificial gene cluster into a single E.coli strain. It can be envisioned that these will become a new generation of bio-reagents that can be coupled with a variety of sugar nucleotides and applied to a large scale synthesis of other sialylated oligosaccharides.
Experimental
General
E. coli K-12 (ATCC 47076) and E. coli K1 (ATCC13027) were purchased from American Type Culture Collection (ATCC), Manassas, Va. E. coli DH5α (lacZΔM15 hsdR recA) was from Gibco-BRL Life Technology, Rockville, Md. E. coli BL21(DE3) [F− ompT hsdSB(rB−mB−) gal dcm (DE3)], plasmid DNA pET15b and pET-NII were bought from Novagen, Madison, Wis. All kits for DNA isolation and purification were from Qiagen, Santa Clarita, Calif. All restriction enzymes, 1 kb DNA ladder, and T4 DNA ligase were obtained from Promega, Madison, Wis. Vent DNA polymerase was purchased from New England Biolabs. Polyphosphate (Pn) and other chemical reagents were from Sigma. Silica gel F254 plates (Merck) and Silica Gel 60 (70–230 mesh, Merck) were used in analytical thin-layer chromatography (TLC) and flash column chromatography, respectively. 1H-NMR was recorded on a Varian Gemini-300, a Mercury-400 or a Varian Unity-500 spectrometer.
PCR and Molecular Cloning
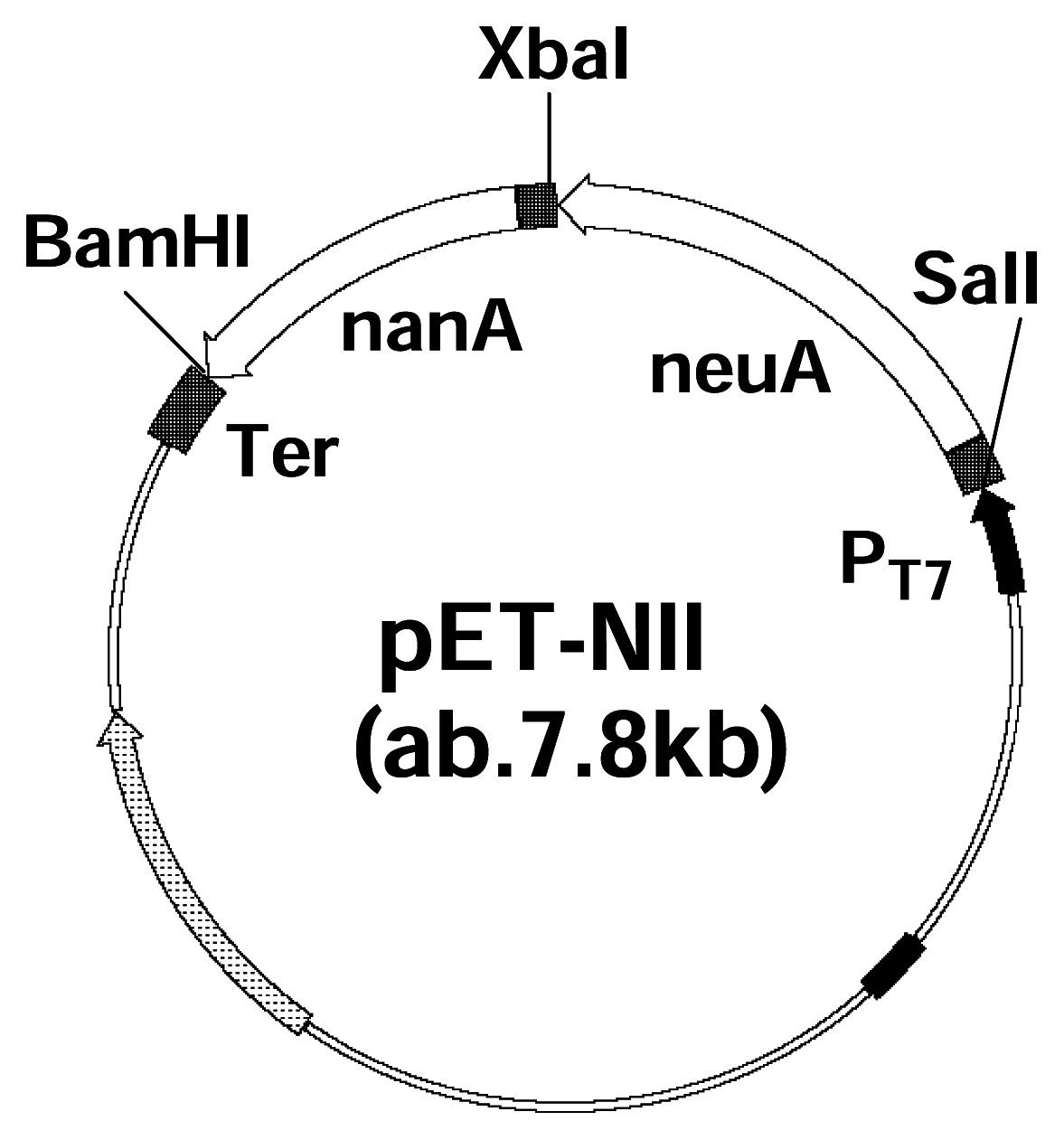
DNA manipulations was performed according to the procedures described by Sambrook et al. [16]. Primers with appropriate restriction sites (underlined) were synthesized by Invitrogen. The Neu5Ac aldolase (EC4.1.3.3) gene (nanA) of E. coli K-12 was amplified from its chromosomal DNA by PCR with primers 5′GCATCGTCGACAAGAAGGAGATATACCATGGCAACGAATTTACGTG3′/5′GGTACGGATCCTCAGTGATGATGATGATGATGCCCGCGCTCTTGCATC3′ (SalI/BamHI are shown underlined). The E. coli K1 CMP-Neu5Ac synthetase (EC 2.7.7.43) gene (neuA) was obtained from its chromosomal DNA by PCR with primers 5′GCATCTCTAGAAAGAAGGAGATATACCATGAGAACAAAAATTATTG3′/5′GCATCTGTCGACTCAGTGATGATGATGATGATGTTTAACA ATCTCCGC3′ (XbaI/SalI, underlined). A His6-coding sequence was incorporated into the reverse primers to attach a poly-His tag at the C-terminus of each protein. PCR was performed in a total volume of 50 μL consisting of Tris-HCl 20 mM (pH 8.8 at 25°C), KCl 10 mM, MgSO4 2 mM, (NH4)2SO4 10 mM, Triton X-100 0.1%, 50 ng template DNA, primers 200 nM each, dNTPs 0.2 mM each, and 0.5 U Vent DNA polymerase. After heating at 96 °C for 2 min, 25 cycles were carried out as denaturation at 96°C for 20 sec, annealing at 55°C for 30 sec and elongation at 72°C for 1 min followed by a final elongation of 7 min. The amplified neuA and nanA genes were digested with XbaI/SalI and SalI/BamHI, respectively, and ligated to pET15b linearized with XbaI/BamHI, resulting in pET-NII (Figure 2.).
Co-expression and purification of Neu5Ac aldolase gene and CMP-Neu5Ac synthetase
The recombinant plasmid pET-NII containing both nanA gene and neuA gene was transformed into E. coli BL21 (DE3) and the transformant was grown in LB (50 mL) with ampicillin (150 μg mL−1) overnight (30°C, 225 rpm), then transferred into freshly prepared LB (1 L) with ampicillin (150μg mL−1) for another 3 h at 37°C. When OD600 reached 0.8–1.0, the culture was induced with 400 μM isopropyl-1-thio-β-D-galactopyranoside (IPTG) for 4 h at 37°C. The cells were harvested by centrifugation (4500 rpm, 10 min, 4°C), disrupted by sonication on ice in 25 mL binding buffer (20 mM Tris-HCl, pH 7.9, 0.5 M NaCl, 5 mM imidazole), clarified by centrifugation and loaded onto a Ni-NTA column (Qiagen). The column was first washed with binding buffer (20 mM Tris-HCl, pH 7.9, 0.5 M NaCl, 5 mM imidazole), then with washing buffer (20 mM Tris-HCl, pH 7.9, 0.5 M NaCl, 20 mM imidazole), followed by elution with elution buffer (20 mM Tris-HCl, pH 7.9, 0.5 M NaCl, 200 mM imidazole). The eluants were concentrated by ultrafiltration, dialyzed against dialysis buffer (20% glycerol, 20 mM, Tris-HCl, pH 7.9).
Isomerization of N-Acetylglucosamine to N-Acetylmannosamine
The isomerization of N-acetylglucosamine was carried out using 0.055 M of NaOH at room temperature. Typically, 20 g of GlcNAc was dissolved in 200 mL of NaOH aqueous solution (0.055M) and allowed to stand at room temperature for 48 h. The resulting pale yellow solution was neutralized by addition of glacial acetic acid and evaporated to dryness under high vacuum. The residue was extracted by refluxing it in 100 ml methanol for 1 h and then standing at room temperature for 4 h, and filtered to remove GlcNAc. The ratio of ManNAc to GlcNAc was determined by 1H-NMR analysis of the dry mixture, particularly by looking at the integrals of both anomeric protons in α- and β- isomers.
Synthesis of CMP-NeuAc in whole cell reaction
The synthesis of CMP-Neu5Ac was carried out in a 200 mL flask containing 50 mL of reaction mixture. Cell pellets (2.5 g, wet weight) suspended in a Tris-HCl (50 mM, pH 7.5) were added to a reaction mixture of ManNAc-enriched solid mixture (GlcNAc/ManNAc, 3/1) (370 mg, 1.25 mM), sodium pyruvate (330 mg, 3.0 mM), CTP (480mg, 1.0 mM), MnCl2 (30 mM), DTT (10 mM) in Tris-HCl buffer (50 mM, pH 7.5). The total volume was brought up to 50 mL by addition of water. The reaction was incubated in an isotherm incubator with agitation in 400 rpm at 37°C for 12 h, when TLC analysis [n-BuOH-AcOH-H2O = 5:3:2 (vol/vol/vol)] indicated that reaction was complete. The reaction was quenched by addition of 50 mL 95% ethanol. The cells were separated from the reaction mixture by centrifugation and washed with deionized water. The combined supernatants were centrifuged again to further remove the pellets. After concentration, the reaction mixture was purified by a Bio-Rad P-2 gel filtration column with distilled water as the mobile phase. The isolation process was monitored by UV (at 272 nm) and TLC analysis analysis [n-BuOH-AcOH-H2O = 5:3:2 (vol/vol/vol)].
Purification of CMP-Neu5Ac
The reaction mixture was centrifuged at 15,000 rpm for 30 min at 4°C to remove the proteins. The pellet was resuspended in 50% aqueous alcohol solution and centrifuged again. The wash step was repeated three times. The combined supernatants were concentrated to a small volume and purified by ion-exchange chromatography. The anion resin (Dowex 1×8) was pretreated with 50 mM of aqueous sodium formate solution to load the column (25 × 500 mm) with formate form. The column was first washed with two volumes of distilled water to remove the monosaccharides and metal ions, and then one volume of 50 mM ammonium bicarbonate solution was used as eluant followed by a 50 mM gradient of ammonium bicarbonate. The fractions containing CMP-Neu5Ac, as evidenced by TLC analysis (eluant: n-BuOH-AcOH-H2O = 5:3:2), were pooled and concentrated to give a white residue. Excess ammonium bicarbonate was removed by dissolving the residue in small amount of water and passing through a column (2.5 × 125 cm) of Sephadex G-15 (Sigma), which has been equilibrated and eluted with distilled water to remove the contamination salts before use. The fractions containing the CMP-Neu5Ac, as identified from TLC analysis were pooled and lyophilized to give rise to the final product. Components other than CMP-Neu5Ac were further removed by means of gel-filtration (size exclusion chromatography) with a column of Bio-Gel P-2 (1.5 × 80 cm) eluting with distilled water. Fractions containing CMP-Neu5Ac were collected, freeze-dried to afford 365 mg of CMP-Neu5Ac (57% based on CTP). The 1H-NMR (D2O) of CMP-Neu5Ac was recorded and it was in good accordance with the literature [21]: δ 8.15 (s, 1H, -NHAc), 7.87 (d, 1H, J = 7.3 Hz, H-6 of cytidine), 6.25 (d, 1H, J = 8.1 Hz, H-5 of cytidine), 5.91 (d, 1H, J = 4.0 Hz, H-1 of ribose), 4.26 (t, 1H, J = 4.0 Hz, H-2 of ribose), 4.23 (t, 1H, J = 4.9 Hz, H-3 of ribose), 4.17 (m, 3H, H-5a, 5b, H-4 of ribose), 4.09~3.98 (m, 2H, H-9a and H-8), 3.90~3.80 (m, 3H, H-5, H-6, H-4), 3.59~3.53 (dd, 1H, J = 11.4, 6.5 Hz, H-9b), 3.40 (d, 1H, J = 9.7 Hz, H-7), 2.42 (dd, 1H, J = 12.9, 4.9 Hz, H-3eq.), 1.98 (s, 3H, -CH3), 1.64~1.54 (ddd, 1H, J = 12.2, 11.2, 4.7 Hz, H-3ax.); 13C-NMR (D2O): δ 174.89 (-C=O), 174.60 (C-1 of NeuAc), 166.31 (C-2 of cytidine), 157.97 (C-4 of cytidine), 141.73 (C-5 of cytidine), 100.24 (C-6 of cytidine), 96.81 (C-2 of NeuAc), 89.25 (C-1 of ribose), 83.09 (C-4 of ribose), 74.43 (C-2 of ribose), 71.95 (C-3 of ribose), 69.75 (C-5 of ribose), 69.24 (C-6 of NeuAc), 68.97 (C-8 of NeuAc), 67.00 (C-4 of NeuAc), 65.10 (C-7 of NeuAc), 63.09 (C-9 of NeuAc), 51.97 (C-5 of NeuAc), 41.27 (C-3 of NeuAc), 22.29 (-CH3). MS (ESI): 634.91 (M + Na+); 656.90 (M + 2Na+).
Figure 1.
Synthesis of CMP-Neu5Ac in the whole cell reaction.

Figure 2.
Plasmid map of CMP-Neu5Ac producing superbug. The map of superbug harboring two genes encoding enzymes involved in the biosynthetic pathway. Introduced restriction sites: SalI, XbaI, and BamHI.
Figure 2.
Plasmid map of CMP-Neu5Ac producing superbug. The map of superbug harboring two genes encoding enzymes involved in the biosynthetic pathway. Introduced restriction sites: SalI, XbaI, and BamHI.

Figure 3.
CMP-Neu5Ac identification by HPLC. The reaction mixture of CMP-Neu5Ac synthesized in whole cell reaction was collected and analyzed by HPLC (A), and the purified CMP-Neu5Ac was collected and analyzed by HPLC (B).
Figure 3.
CMP-Neu5Ac identification by HPLC. The reaction mixture of CMP-Neu5Ac synthesized in whole cell reaction was collected and analyzed by HPLC (A), and the purified CMP-Neu5Ac was collected and analyzed by HPLC (B).

Figure 4.
Time course for CMP-Neu5Ac production at a 2-mL scale. The accumulation of CMP-Neu5Ac reached its peak at around 12 h in the whole cell reaction catalyzed by the superbug containing fusion protein of both Neu5Ac aldolase and CMP-Neu5Ac synthetase. It was accumulated to 13.22 mM (8.12 g/L) in the reaction mixture. When it was exposed in the reaction mixture after longer time, however, CMP-Neu5Ac might be decomposed since the concentration decreased.
Figure 4.
Time course for CMP-Neu5Ac production at a 2-mL scale. The accumulation of CMP-Neu5Ac reached its peak at around 12 h in the whole cell reaction catalyzed by the superbug containing fusion protein of both Neu5Ac aldolase and CMP-Neu5Ac synthetase. It was accumulated to 13.22 mM (8.12 g/L) in the reaction mixture. When it was exposed in the reaction mixture after longer time, however, CMP-Neu5Ac might be decomposed since the concentration decreased.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Alcohols | Solubility (g/L) | ManNAc/GlcNAc | |
|---|---|---|---|
| ManNAc | GlcNAc | ||
| Methanol | 90.6 | 4.2 | 21.6 |
| Ethanol | 8.0 | 0.7 | 11.4 |
| n-Propanol | 1.1 | 0.2 | 5.5 |
| i-Propanol | 1.0 | 0.2 | 5.0 |
- Sample Availability: Available from the authors.
References and Notes
- Schauer, R. Chemistry, metabolism, and biological functions of sialic acids. Adv. Carbohydr. Chem. Biochem 1982, 40, 131–234. [Google Scholar]
- Angata, T.; Varki, A. Chemical Diversity in the Sialic Acids and Related a-Keto Acids: An Evolutionary Perspective. Chem. Rev. (Washington, D. C.) 2002, 102, 439–469. [Google Scholar]
- Sialic acids. In Essentials of Glycobiology; Varki, A.; Cummings, R.; Esko, J.; Freeze, H.; Harth, G.; Marth, J. (Eds.) Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, 1999; pp. 195–209.
- Martin, T. J.; Schmidt, R. R. Convenient chemical synthesis of CMP-N-acetylneuraminate (CMP-Neu-5-Ac). Tetrahedron Lett 1993, 34, 1765–8. [Google Scholar]
- Kittelmann, M.; Klein, T.; Kragl, U.; Wandrey, C.; Ghisalba, O. CMP-N-acetylneuraminic acid synthetase from Escherichia coli. Fermentation production and application for the preparative synthesis of CMP-neuraminic acid. Appl. Microbiol. Biotechnol 1995, 44, 59–67. [Google Scholar]
- Endo, T.; Koizumi, S.; Tabata, K.; Ozaki, A. Large-scale production of CMP-NeuAc and sialylated oligosaccharides through bacterial coupling. Appl. Microbiol. Biotechnol 2000, 53, 257–261. [Google Scholar]
- Vimr, E. R.; Troy, F. A. Regulation of sialic acid metabolism in Escherichia coli: role of Nacylneuraminate pyruvate-lyase. J. Bacteriol 1985, 164, 854–60. [Google Scholar]
- Munster, A. K.; Eckhardt, M.; Potvin, B.; Muhlenhoff, M.; Stanley, P.; Gerardy-Schahn, R. Mammalian cytidine 5′-monophosphate N-acetylneuraminic acid synthetase: a nuclear protein with evolutionarily conserved structural motifs. Proc. Nat. Acad. Sci. U.S.A 1998, 95, 9140–5. [Google Scholar]
- Vann, W. F.; Silver, R. P.; Abeijon, C.; Chang, K.; Aaronson, W.; Sutton, A.; Finn, C. W.; Lindner, W.; Kotsatos, M. Purification, properties, and genetic location of Escherichia coli cytidine 5′-monophosphate N-acetylneuraminic acid synthetase. J. Biol. Chem 1987, 262, 17556–62. [Google Scholar]
- Zapata, G.; Vann, W. F.; Aaronson, W.; Lewis, M. S.; Moos, M. Sequence of the cloned Escherichia coli K1 CMP-N-acetylneuraminic acid synthetase gene. J. Biol. Chem 1989, 264, 14769–74. [Google Scholar]
- Edwards, U.; Frosch, M. Sequence and functional analysis of the cloned Neisseria meningitidis CMP-NeuNAc synthetase. FEMS Microbiol. Lett 1992, 75, 161–6. [Google Scholar]
- Tullius, M. V.; Munson, R. S., Jr; Wang, J.; Gibson, B. W. Purification, cloning, and expression of a cytidine 5′-monophosphate N-acetylneuraminic acid synthetase from Haemophilus ducreyi. J. Biol. Chem 1996, 271, 15373–80. [Google Scholar]
- Stoughton, D. M.; Zapata, G.; Picone, R.; Vann, W. F. Identification of Arg-12 in the active site of Escherichia coli K1 CMP-sialic acid synthetase. Biochem. J 1999, 343 Pt 2, 397–402. [Google Scholar]
- Chen, X.; Fang, J.; Zhang, J.; Liu, Z.; Shao, J.; Kowal, P.; Andreana, P.; Wang, P. G. Sugar nucleotide regeneration beads (superbeads): a versatile tool for the practical synthesis of oligosaccharides. J. Am. Chem. Soc 2001, 123, 2081–2082. [Google Scholar]
- Chen, X.; Liu, Z.; Zhang, J.; Zhang, W.; Kowal, P.; Wang, P. G. Reassembled biosynthetic pathway for large-scale carbohydrate synthesis: a-gal epitope producing “superbug”. Chem BioChem 2002, 3, 47–53. [Google Scholar]
- Maniatis, T.; Fritsch, E. F.; Sambrook, J. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, 1982; pp. 1–35. [Google Scholar]
- Keppler, O. T.; Hinderlich, S.; Langner, J.; Schwartz-Albiez, R.; Reutter, W.; Pawlita, M. UDP-GlcNAc 2-epimerase: a regulator of cell surface sialylation. Science 1999, 284, 1372–6. [Google Scholar]
- Maru, I.; Ohnishi, J.; Ohta, Y.; Tsukada, Y. Simple and large-scale production of N-acetylneuraminic acid from N-acetyl-D-glucosamine and pyruvate using N-acyl-D-glucosamine 2-epimerase and N-acetylneuraminate lyase. Carbohydr. Res 1998, 306, 575–8. [Google Scholar]
- Muhmoudian, M.; Noble, D.; Drake, C. S.; Middleton, R. F.; Montgomery, D. S.; Piercey, J. E.; Ramlakhan, D.; Todd, M.; Dawson, M. J. An efficient process for production of N-acetylneuraminic acid using N-acetylneuraminic acid aldolase. Enzyme Microb. Technol 1997, 20, 393–400. [Google Scholar]
- Uchida, Y.; Tsukada, Y.; Sugimori, T. Purification and properties of N-acetylneuraminate lyase from Escherichia coli. J. Biochem 1984, 96, 507–22. [Google Scholar]
- Simon, E. S.; Bednarski, M. D.; Whitesides, G. M. Synthesis of CMP-NeuAc from N-acetylglucosamine: generation of CTP from CMP using adenylate kinase. J. Am. Chem. Soc 1988, 110, 7159–63. [Google Scholar]
- Koeller, K. M.; Wong, C.-H. Synthesis of Complex Carbohydrates and Glycoconjugates: Enzyme-Based and Programmable One-Pot Strategies. Chem. Rev. (Washington, D. C.) 2000, 100, 4465–4493. [Google Scholar]
- Palcic, M. M. Biocatalytic synthesis of oligosaccharides. Curr. Opin. Biotechnol 1999, 10, 616–624. [Google Scholar]
- Fang, J.; Li, J.; Chen, X.; Zhang, Y.; Wang, J.; Guo, Z.; Zhang, W.; Yu, L.; Brew, K.; Wang, P. G. Highly Efficient Chemoenzymic Synthesis of a-Galactosyl Epitopes with a Recombinant a(1- >3)-Galactosyltransferase. J. Am. Chem. Soc 1998, 120, 6635–6638. [Google Scholar]
- Seitz, O.; Wong, C.-H. Chemoenzymic solution- and solid-phase synthesis of O-glycopeptides of the Mucin domain of MAdCAM-1. A general route to O-LacNAc, O-Sialyl-LacNAc, and O-Sialyl- Lewis-X peptides. J. Am. Chem. Soc 1997, 119, 8766–8776. [Google Scholar]
- Baisch, G.; Ohrlein, R.; Streiff, M.; Kolbinger, F. Enzymatic synthesis of sialyl-Lewis(a)- libraries with two non-natural monosaccharide units. Bioorg. Med. Chem. Lett 1998, 8, 755–8. [Google Scholar]
© 2003 by MDPI Reproduction is permitted for noncommercial purposes.
Share and Cite
MDPI and ACS Style
Song, J.; Zhang, H.; Wu, B.; Zhang, Y.; Li, H.; Xiao, M.; Wang, P.G. Reassembled Biosynthetic Pathway for a Large-scale Synthesis of CMP-Neu5Ac. Mar. Drugs 2003, 1, 34-45. https://doi.org/10.3390/md101034
AMA Style
Song J, Zhang H, Wu B, Zhang Y, Li H, Xiao M, Wang PG. Reassembled Biosynthetic Pathway for a Large-scale Synthesis of CMP-Neu5Ac. Marine Drugs. 2003; 1(1):34-45. https://doi.org/10.3390/md101034
Chicago/Turabian StyleSong, Jing, Hesheng Zhang, Bingyuan Wu, Yingxin Zhang, Hanfen Li, Min Xiao, and Peng George Wang. 2003. "Reassembled Biosynthetic Pathway for a Large-scale Synthesis of CMP-Neu5Ac" Marine Drugs 1, no. 1: 34-45. https://doi.org/10.3390/md101034