Antiproliferative Activity of Cyanophora paradoxa Pigments in Melanoma, Breast and Lung Cancer Cells


Abstract
:1. Introduction
2. Results and Discussion
2.1. Antiproliferative Activity of Cp Extracts

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell line | |||
|---|---|---|---|
| A-549 Lung adenocarcinoma | MCF-7 Mammary carcinoma | A-2058 Malignant melanoma | |
| DCM extract | −15.4 ± 2.7 | −14.6 ± 1.8 | 53.3 ± 4.8 * |
| EtOH extract 1 | 26.2 ± 2.8 | 22.2 ± 2.2 | 61.8 ± 4.1 * |
| EtOH extract 2 | 23.3 ± 0.3 | 23.8 ± 0.2 | 26.6 ± 0.3 * |
| Water extract 1 | ND | ND | ND |
| Water extract 2 | 8.6 ± 0.3 | 25.5 ± 0.4 * | 19.4 ± 0.3 |
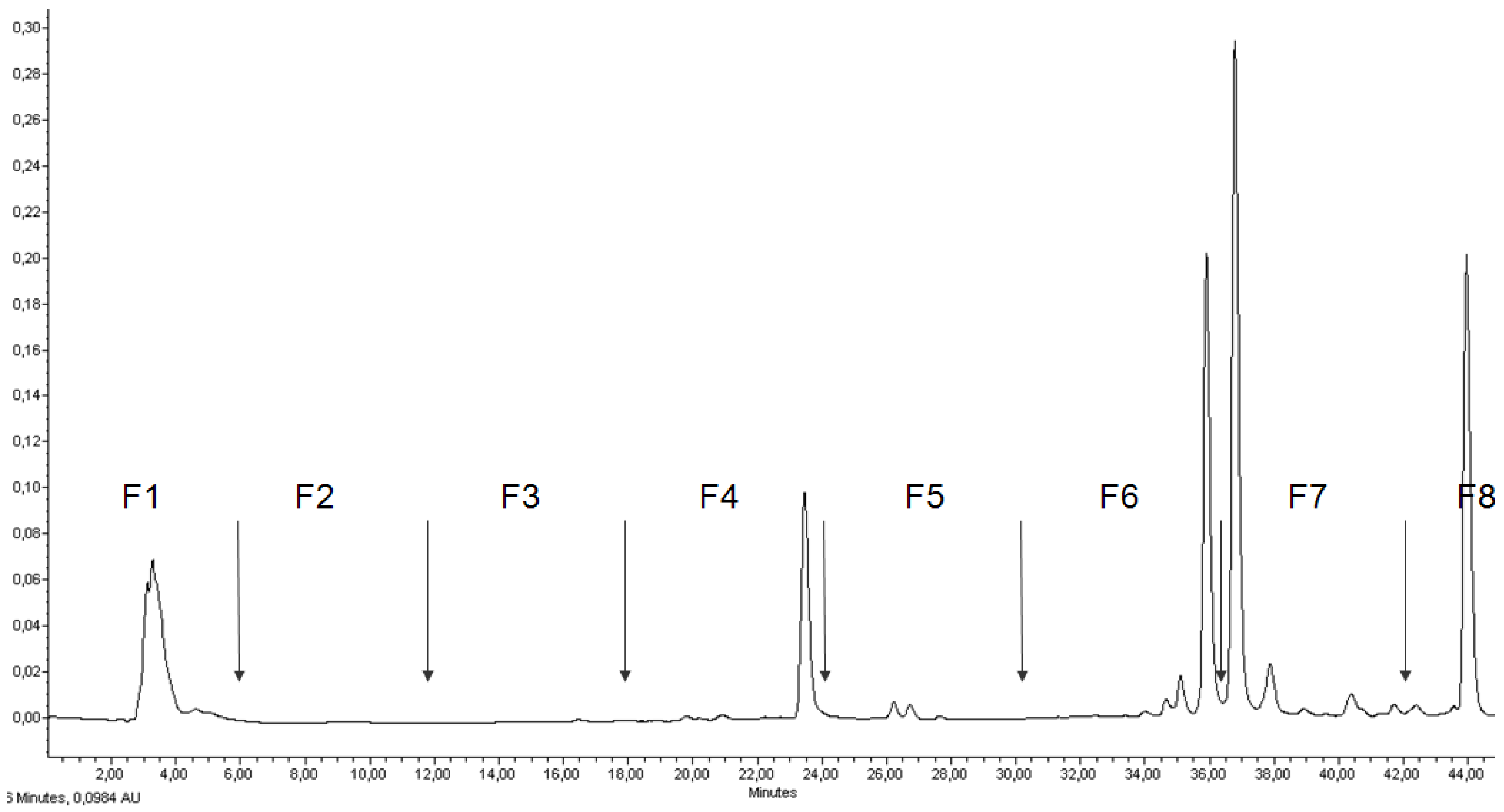
2.2. RP-HPLC Analysis, Fractionation of the Ethanol Extract, and Antiproliferative Activity of Flash Chromatography Fractions
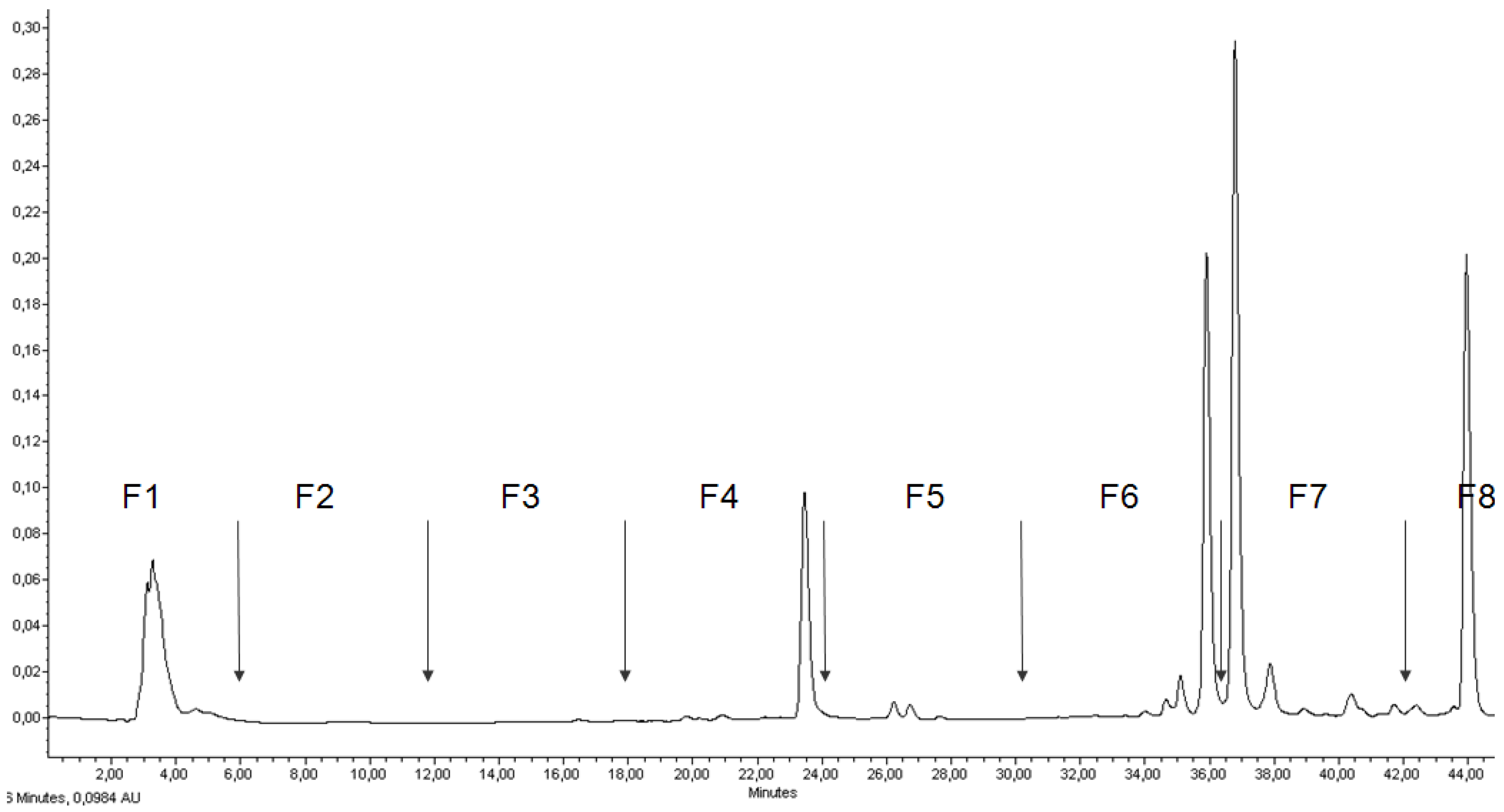
| Cell line | |||
|---|---|---|---|
| A-2058 Malignant melanoma | A-549 Lung adenocarcinoma | MCF-7 Mammary carcinoma | |
| Flash chromatography F1 | −2.9 ± 0.3 | 1.6 ± 0.2 | 7.9 ± 0.1 |
| Flash chromatography F2 | 19.2 ± 0.2 | 5.2 ± 0.2 | 12.4 ± 0.1 |
| Flash chromatography F3 | 91.2 ± 0.1 * | 54.4 ± 0.2 ** | 36.8 ± 0.1 * |
| Flash chromatography F4 | 34.4 ± 0.2 ** | 45.7 ± 0.2 ** | 29.0 ± 0.1 * |
| Flash chromatography F5 | 93.0 ± 0.1 * | 19.2 ± 0.2 ** | 36.1 ± 0.1 |
| Flash chromatography F6 | 21.3 ± 0.2 * | 17.6 ± 0.2 ** | 57.3 ± 0.2 * |
| Flash chromatography F7 | 5.0 ± 0.2 | −4.7 ± 0.2 | −2.6 ± 0.2 |
| Flash chromatography F8 | 0.5 ± 0.3 | −26.9 ± 0.2 | −10.5 ± 0.2 |
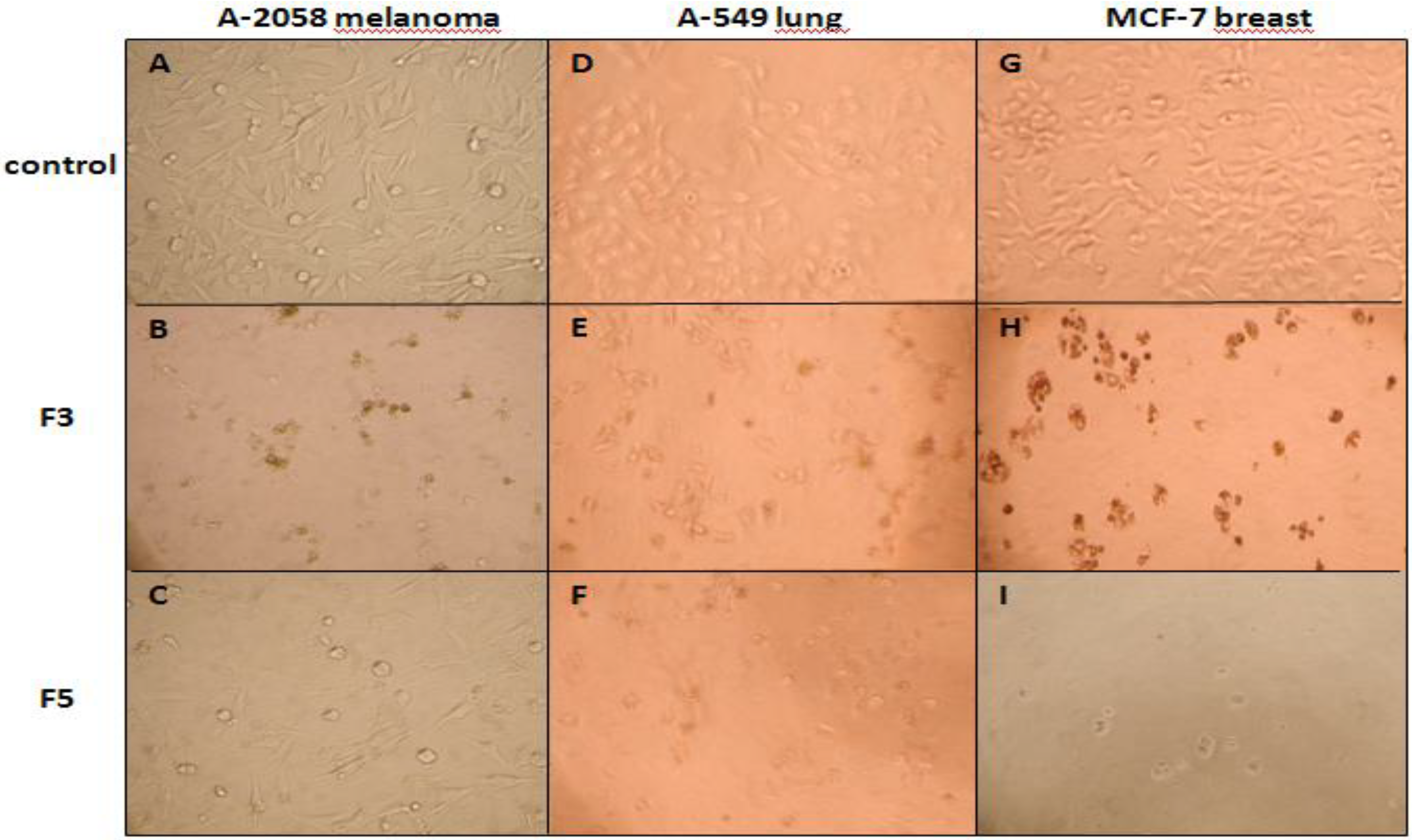
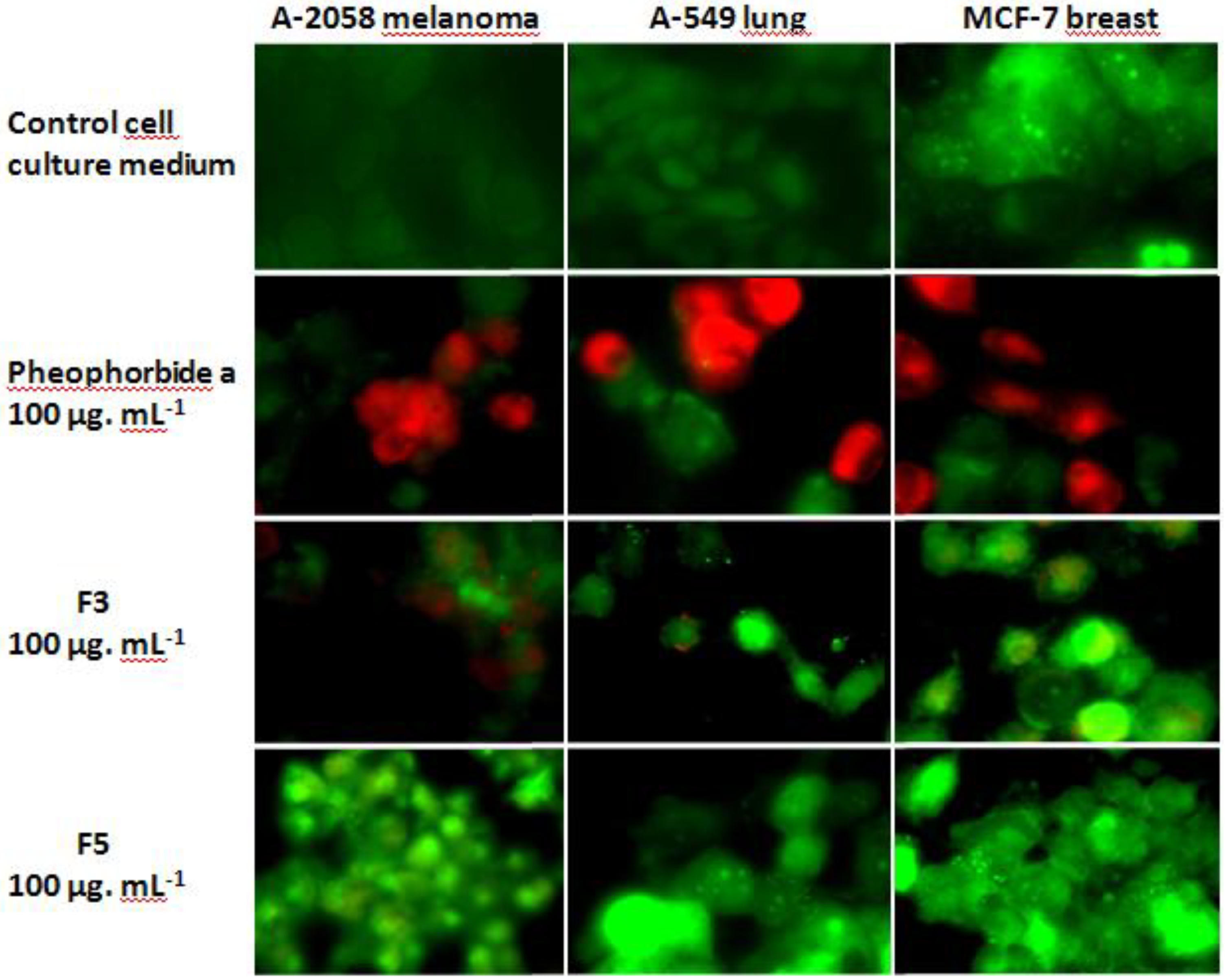
2.3. Cytotoxic and Pro-Apoptotic Activity of F3 and F5

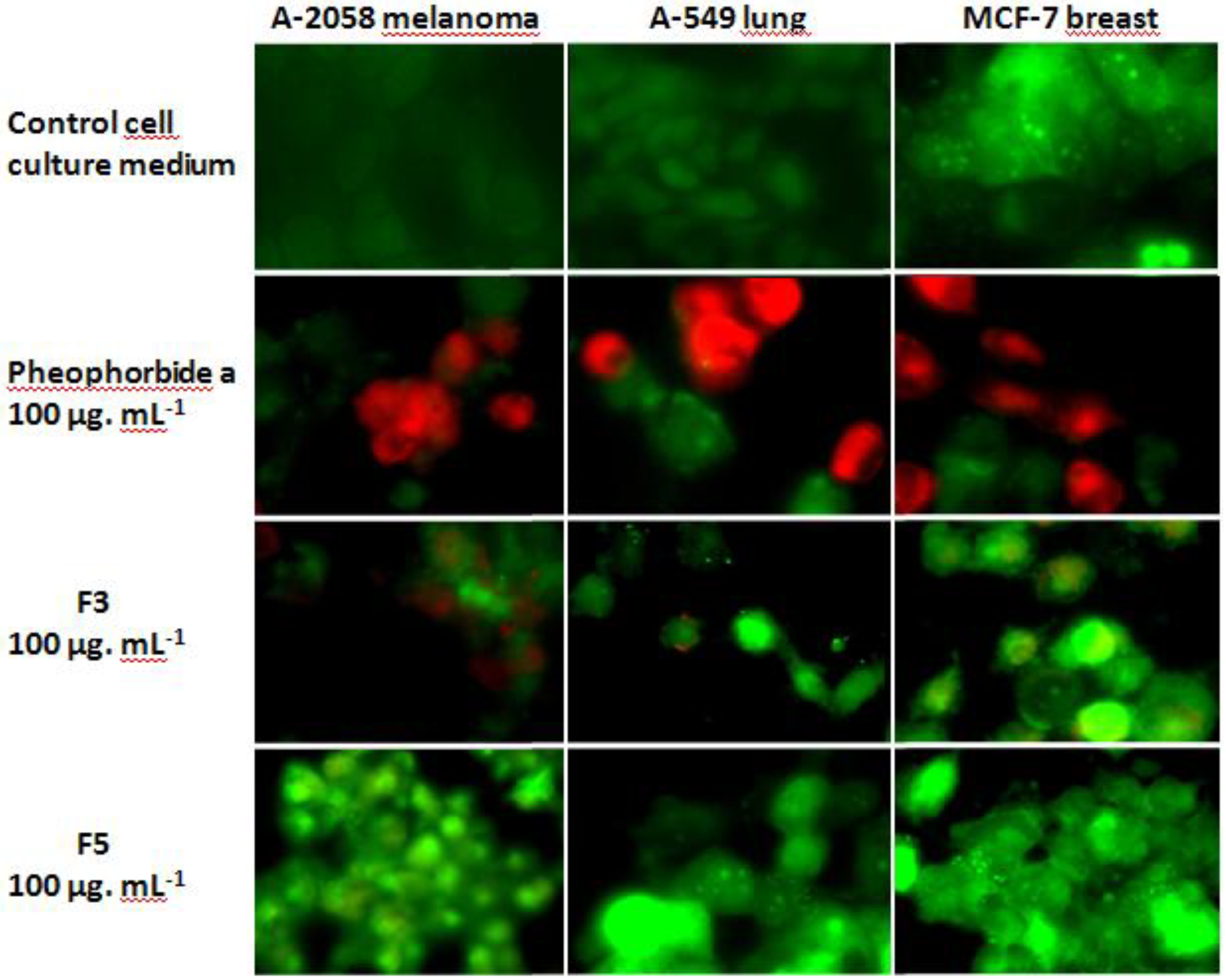
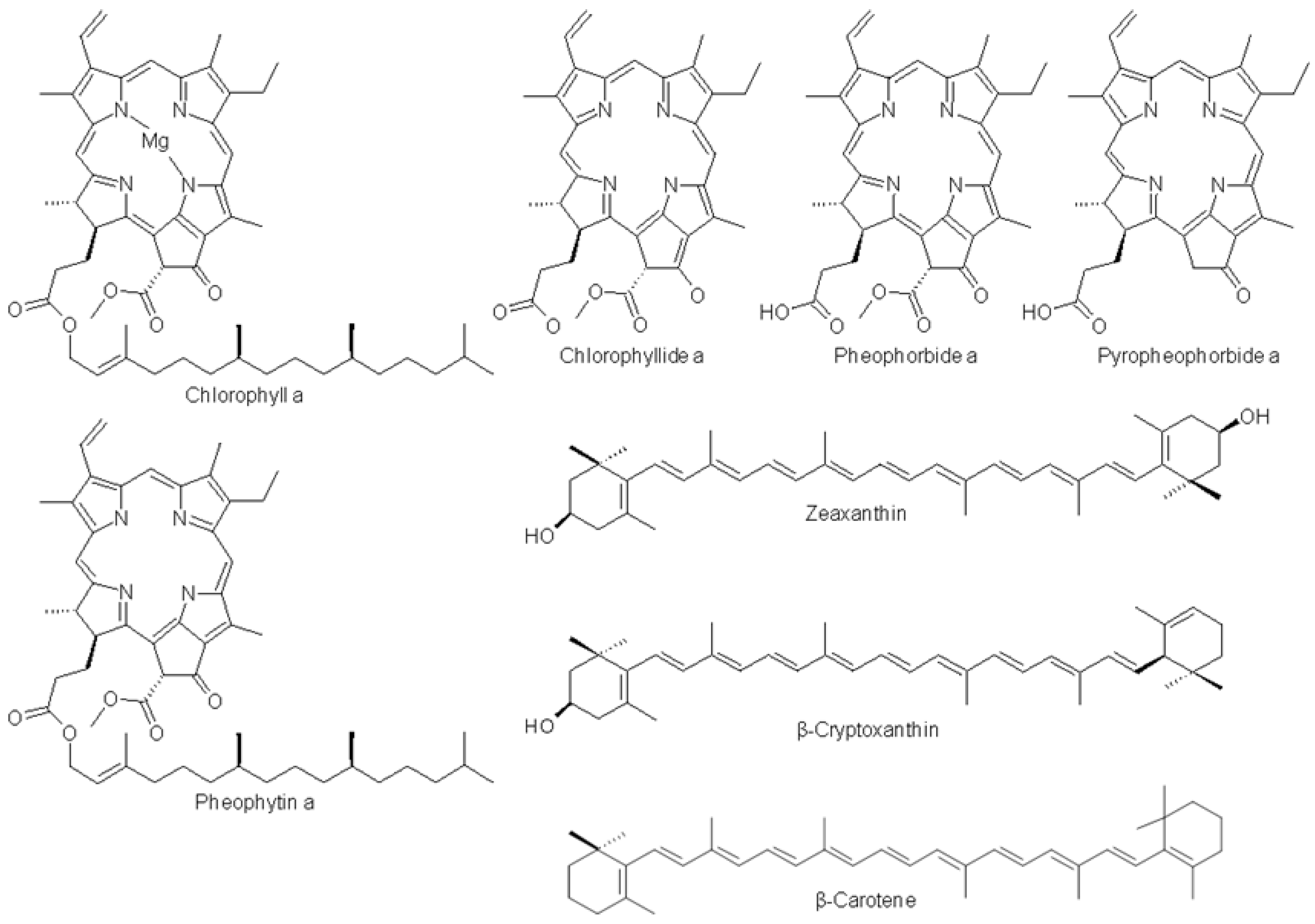
2.4. Characterization of the Cp Fractions
| Standard Pigment | λmax (nm) | Ratio | ||
|---|---|---|---|---|
| I | II | III | III/II | |
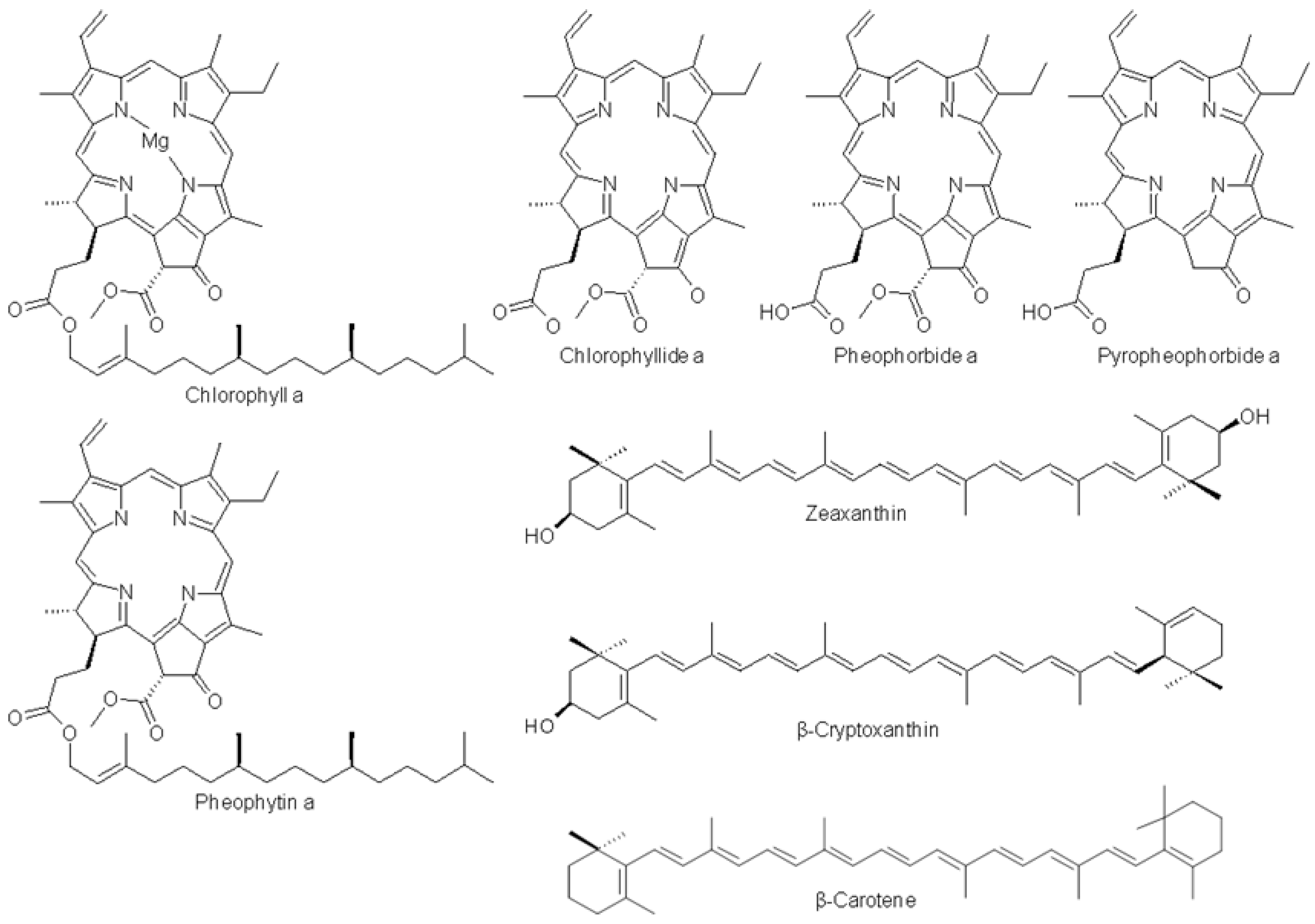
| Chlorophyllide a | - | 434.0 | 668.0 | - |
| Pheophorbide a | - | 410.0 | 665.4 | - |
| Pyropheophorbide a | - | 407.6 | 660.4 | - |
| Zeaxanthin | - | 454.8 | 481.6 | 32% |
| β-cryptoxanthin | - | 447.3 | 474.9 | 23% |
| Chlorophyll a | - | 429.3 | 660.5 | - |
| Pheophytin a | - | 409.0 | 666.0 | - |
| β-carotene | - | 453.6 | 481.6 | 25% |
| Fraction | Peak | HPLC Rt (min) | λmax (nm) | Ratio | Tentative Identification | ||
|---|---|---|---|---|---|---|---|
| I | II | III | III/II | ||||
| F1 | 1 | 3.5 | 430.0 | 666.7 | - | Chlorophyllide a | |
| F2 | 2 | 9.3 | 408.8 | 666.7 | - | Chlorophyllone a | |
| F3 | 3 | 12.5 | 410.0 | 666.7 | - | Pheophorbide a | |
| F4 | 4 | 21.0 | 410.0 | 660.0 | - | Pyropheophorbide a | |
| 5 | 24.0 | 454.9 | 481.7 | 28% | Zeaxanthin | ||
| F5 | 6 | 26.4 | 447.6 | 474.4 | 46% | 5,6-epoxy β-cryptoxanthin | |
| 7 | 26.9 | 447.9 | 474.4 | 20% | β-cryptoxanthin | ||
| F6 | 8 | 34.7 | 429.4 | 666.7 | - | Chlorophyll a allomer | |
| 9 | 35.0 | 433.0 | 451.2 | 485.3 | 50% | Unidentified carotenoid | |
| 10 | 36.0 | 429.4 | 661.7 | - | Chlorophyll a | ||
| F7 | 11 | 36.8 | 430.6 | 663.0 | - | Chlorophyll a epimer | |
| 12 | 37.7 | 417.3 | 661.7 | - | DV-pheophytin a | ||
| 13 | 39.0 | 429.4 | 454.9 | 40% | Mutatochrome | ||
| 14 | 40.5 | 408.8 | 669.1 | - | MV-pheophytin a | ||
| 15 | 41.4 | 401.5 | 669.1 | - | Purpurin-7 phytyl ester | ||
| 16 | 42.3 | 410.0 | 666.7 | - | Pheophytin a | ||
| F8 | 17 | 43.6 | 454.9 | 480.4 | 23% | β-carotene | |
2.4.1. F1
2.4.2. F2
| Standardpigment | Formula | UPLC Rt (min) | Theoricalm/z | Experimentalm/z | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M. | [M + H]+ | [M + Na]+ | [M − Mg + 3H]+ | M. | [M + H]+ | [M + Na]+ | [M − Mg + 3H]+ | Fragments | |||||
| Chlorophyllide a | C35H34N4O5Mg | 7.08 | - | 615.2458 | - | 593.2764 | - | 615.2580 | - | 593.2761 | 533.2545 | - | - |
| Pheophorbide a | C35H36N4O5 | 7.05 | - | 593.2764 | - | - | - | 593.2762 | - | - | 533.2544 | 539.2417 | - |
| Pyropheophorbide a | C33H34N4O3 | 7.45 | - | 535.2709 | 557.2529 | - | - | 535.2699 | 557.2520 | - | 435.2537 | 256.3005 | - |
| Zeaxanthin | C40H56O2 | 8.14 | 568.4280 | - | - | - | 568.4268 | - | - | - | 549.4098 | - | - |
| β-Cryptoxanthin | C40H56O | 10.04 | - | 553.4409 | 575.4137 | - | - | 553.4346 | - | - | 533.4146 | 460.3685 | - |
| Chlorophyll a | C55H72N4O5Mg | 10.03 | - | 893.5432 | 915.5251 | 871.5737 | 893.5403 | 915.5237 | 871.5727 | 614.2376 | 555.2232 | 637.2258 | |
| β-Carotene | C40H56 | 10.83 | 536.4382 | - | - | - | 536.4370 | - | - | - | 444.3751 | - | - |
| Fractions | UPLC Rt (min) | Experimental m/z | Confirmation of pigment identification | ||||||
|---|---|---|---|---|---|---|---|---|---|
| - | M. | [M + H]+ | [M + Na]+ | [M − Mg + 3H]+ | Fragments | ||||
| F1 | 7.08 | - | - | - | 593.2743 | 533.2540 | - | - | Chlorophyllide a |
| F2 | 7.08 | - | - | - | 593.2743 | 533.2540 | - | - | Lack of chlorophyllone a standard to confirm that the detected ions derive from it |
| F3 | 7.05 | - | 593.2756 | - | - | 533.2542 | - | - | Pheophorbide a |
| F4 | 7.45 | - | 535.2702 | 557.2519 | - | 435.2534 | - | - | Pyropheophorbide a Zeaxanthin |
| 8.15 | 568.4260 | - | - | - | 549.4086 | - | - | ||
| F5 | 10.05 | - | - | 575.4137 | - | 371.2270 | - | - | β-cryptoxanthin Lack of 5,6-epoxy-β-cryptoxanthin standard to confirm that the detected ions derive from it |
| F6 | 10.04 | - | 893.5405 | 915.5229 | 871.5722 | 614.2380 | 555.2234 | 637.2262 | Chlorophyll a Pheophytin a (pheophytination of chlorophyll a) No confirmation of the presence of the unidentified carotenoid |
| 10.61 | 871.5724 | - | - | 533.2548 | - | - | |||
| F7 | 10.04 | - | 893.5405 | 915.5229 | 871.5722 | 614.2380 | 555.2234 | 637.2262 | Chlorophyll a epimer Pheophytin a No confirmation of the presence of DV- and MV-pheophytin a, mutatochrome and purpuryl-7-phytyl ester |
| 10.62 | - | 871.5723 | 893.5538 | - | 533.2539 | - | - | ||
| F8 | 10.80 | 536.4372 | - | - | - | - | - | - | β-carotene |
2.4.3. F3
2.4.4. F4
2.4.5. F5
2.4.6. F6
2.4.7. F7
2.4.8. Fraction 8
3. Experimental Section
3.1. Chemicals, Reagents and Chromatography Columns
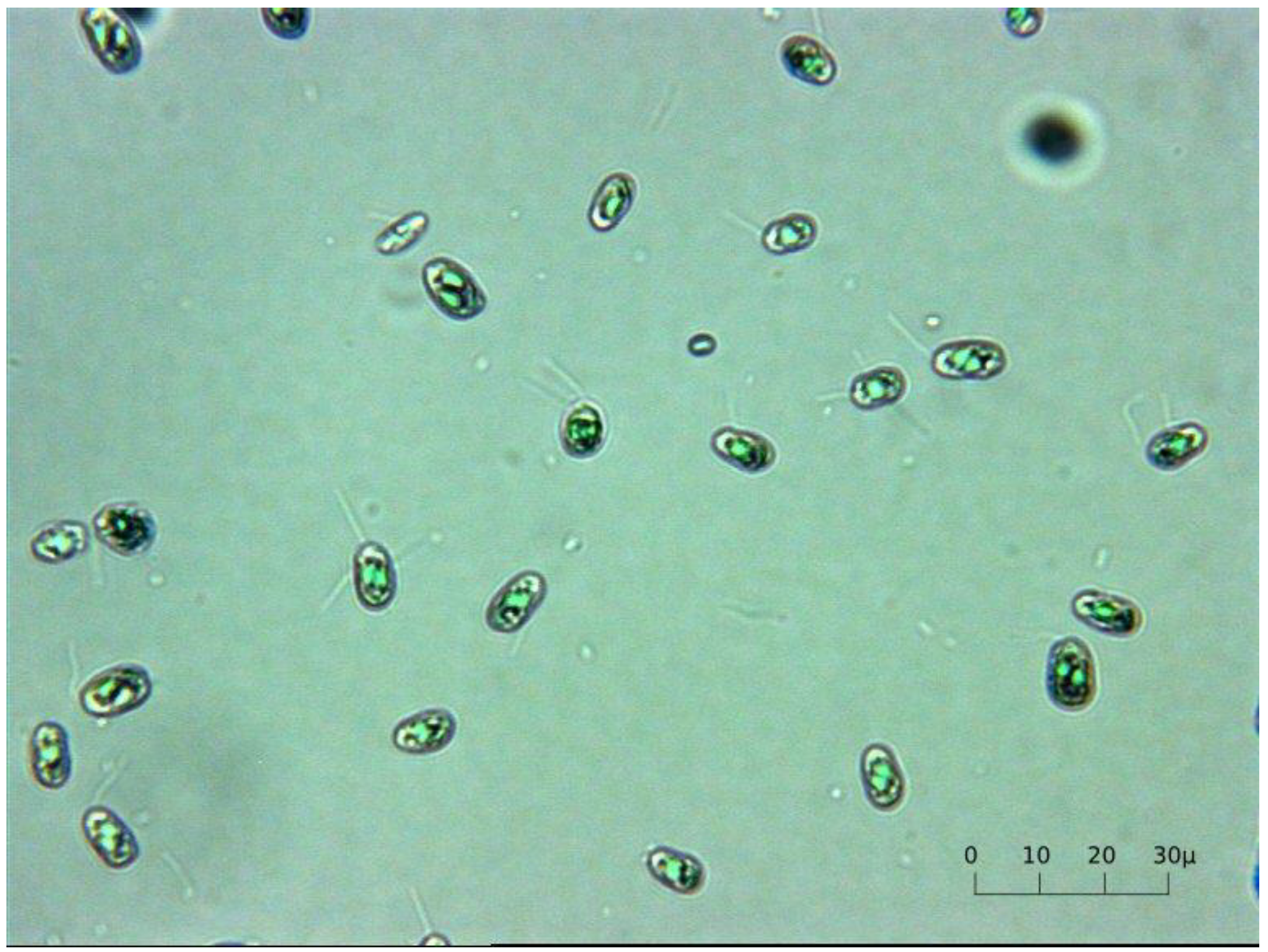
3.2. Microalgae
3.3. Microalgae Culture, Collection and Storage
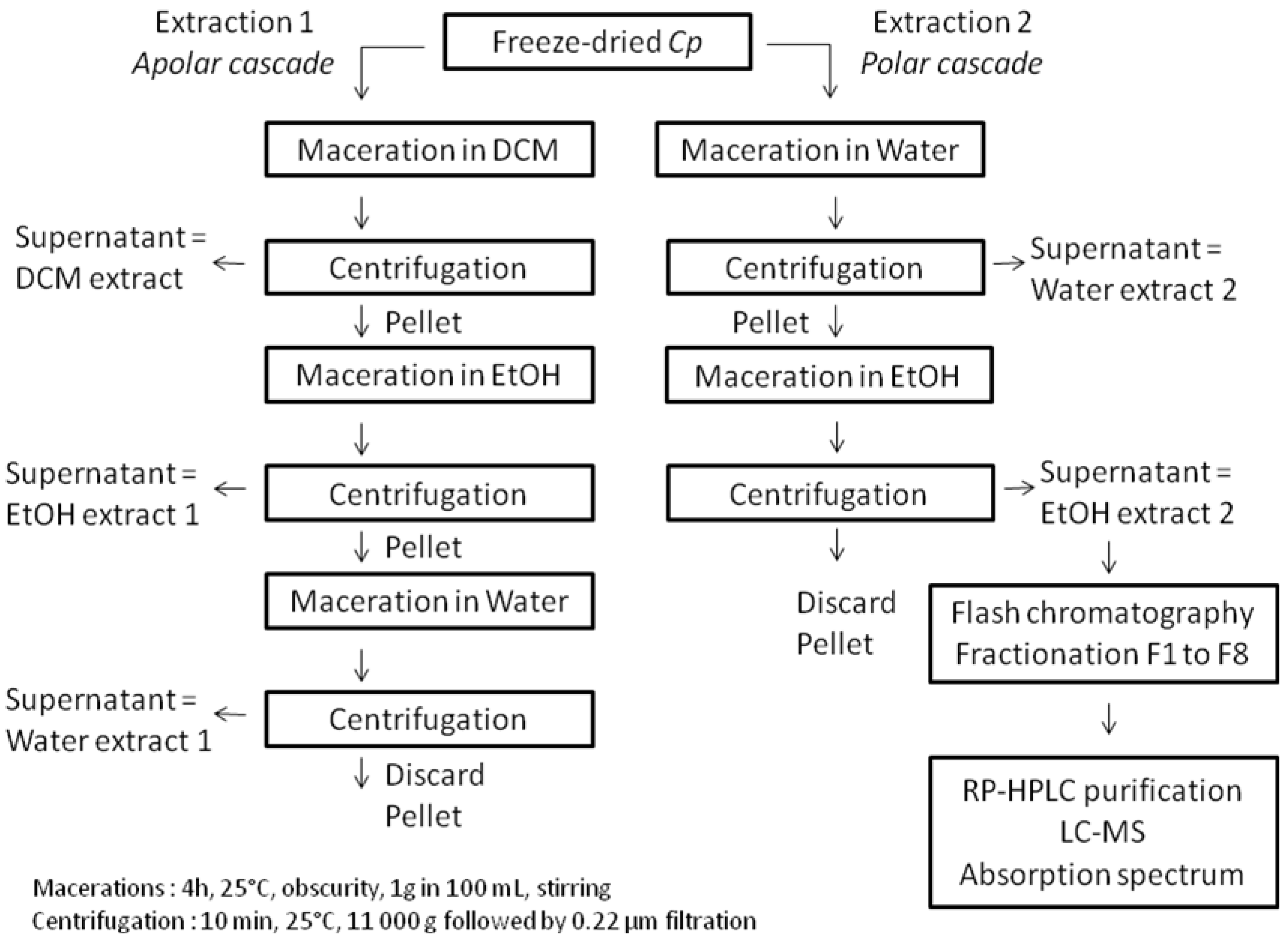
3.4. Pigments Extraction, Fractionation and Purification
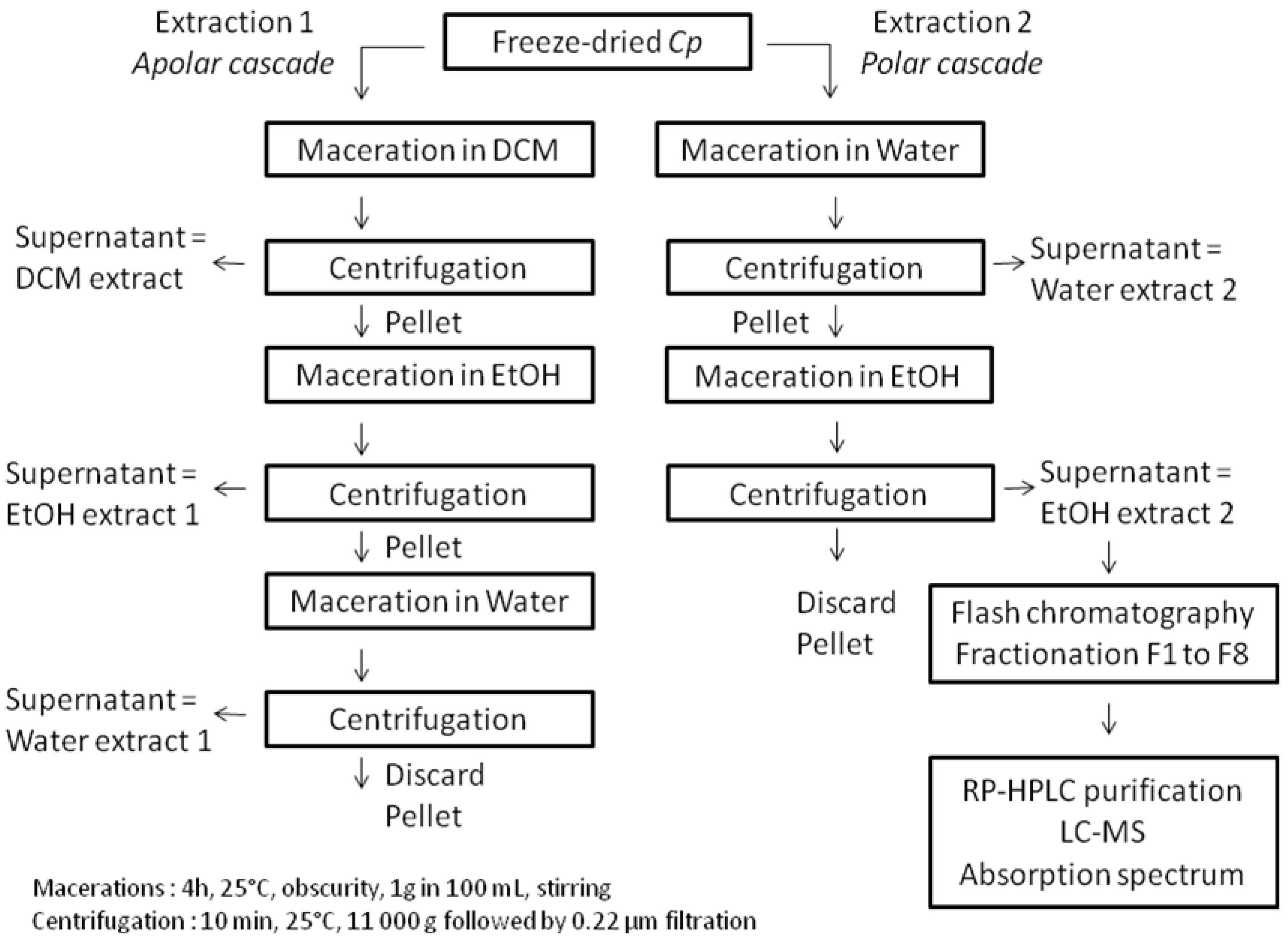
3.5. RP-HPLC Pigments Analysis
3.6. Mass Spectrometry
3.7. Cell Culture and Viability Assay
3.8. Apoptosis Assay
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Talero, E.; Avila-Roman, J.; Motilva, V. Chemoprevention with phytonutrients and microalgae products in chronic inflammation and colon cancer. Curr. Pharm. Des. 2012, 18, 3939–3965. [Google Scholar] [CrossRef]
- Nappo, M.; Berkov, S.; Massucco, C.; di Maria, V.; Bastida, J.; Codina, C.; Avila, C.; Messina, P.; Zupo, V.; Zupo, S. Apoptotic activity of the marine diatom Cocconeis scutellum and eicosapentaenoic acid in BT20 cells. Pharm. Biol. 2012, 50, 529–535. [Google Scholar] [CrossRef]
- Guedes, A.C.; Amaro, H.M.; Malcata, F.X. Microalgae as sources of carotenoids. Mar. Drugs 2011, 9, 625–644. [Google Scholar] [CrossRef]
- Bhatnagar, I.; Kim, S.-K. Immense essence of excellence: marine microbial bioactive compounds. Mar. Drugs 2010, 8, 2673–2701. [Google Scholar] [CrossRef]
- Leya, T.; Rahn, A.; Lütz, C.; Remias, D. Response of arctic snow and permafrost algae to high light and nitrogen stress by changes in pigment composition and applied aspects for biotechnology. FEMS Microbiol. Ecol. 2009, 67, 432–443. [Google Scholar] [CrossRef]
- Gagez, A.-L.; Thiéry, V.; Pasquet, V.; Cadoret, J.-P.; Picot, L. Epoxycarotenoids and cancer. Review. Curr. Bioact. Compd. 2012, 8, 109–141. [Google Scholar] [CrossRef]
- Pasquet, V.; Morisset, P.; Ihammouine, S.; Chepied, A.; Aumailley, L.; Berard, J.-B.; Serive, B.; Kaas, R.; Lanneluc, I.; Thiery, V.; et al. Antiproliferative activity of violaxanthin isolated from bioguided fractionation of dunaliella tertiolecta extracts. Mar. Drugs 2011, 9, 819–831. [Google Scholar] [CrossRef]
- Moreau, D.; Tomasoni, C.; Jacquot, C.; Kaas, R.; Le Guedes, R.; Cadoret, J.-P.; Muller-Feuga, A.; Kontiza, I.; Vagias, C.; Roussis, V.; et al. Cultivated microalgae and the carotenoid fucoxanthin from Odontella aurita as potent anti-proliferative agents in bronchopulmonary and epithelial cell lines. Environ. Toxicol. Pharmacol. 2006, 22, 97–103. [Google Scholar] [CrossRef]
- Mathieu, V.; de Lassalle, E.M.; Toelen, J.; Mohr, T.; Bellahcène, A.; van Goietsenoven, G.; Verschuere, T.; Bouzin, C.; Debyser, Z.; de Vleeschouwer, S.; et al. Galectin-1 in melanoma biology and related neo-angiogenesis processes. J. Investig. Dermatol. 2012, 132, 2245–2254. [Google Scholar] [CrossRef]
- Jeffrey, S.; Mantoura, R.; Wright, S. Iternational council of scientific unions. In SCOR UNESCO Phytoplankton Pigments in Oceanography: Guidelines to Modern Methods; Jeffrey, S., Mantoura, R., Wright, S., Eds.; UNESCO Publishing: Paris, France, 1997. [Google Scholar]
- Pickering, M. Low Temperature Sequestration of Photosynthetic Pigments: Model Studies and Natural Aquatic Environments. Ph.D. Thesis, University of York, York, UK, August 2009. [Google Scholar]
- Chan, J.Y.-W.; Tang, P.M.-K.; Hon, P.-M.; Au, S.W.-N.; Tsui, S.K.-W.; Waye, M.M.-Y.; Kong, S.-K.; Mak, T.C.-W.; Fung, K.-P. Pheophorbide a, a major antitumor component purified from Scutellaria barbata, induces apoptosis in human hepatocellular carcinoma cells. Planta Med. 2006, 72, 28–33. [Google Scholar] [CrossRef]
- Hibasami, H.; Kyohkon, M.; Ohwaki, S.; Katsuzaki, H.; Imai, K.; Nakagawa, M.; Ishi, Y.; Komiya, T. Pheophorbide a, a moiety of chlorophyll a, induces apoptosis in human lymphoid leukemia molt 4B cells. Int. J. Mol. Med. 2000, 6, 277–279. [Google Scholar]
- Magyar, A.; Bowman, M.; Molnar, P.; Kispert, L. Neutral carotenoid radicals in photoprotection of wild-type Arabidopsis thaliana. J. Phys. Chem. B 2013, 117, 2239–2246. [Google Scholar] [CrossRef]
- Wu, N.-L.; Chiang, Y.-C.; Huang, C.-C.; Fang, J.-Y.; Chen, D.-F.; Hung, C.-F. Zeaxanthin inhibits PDGF-BB-induced migration in human dermal fibroblasts. Exp. Dermatol. 2010, 19, e173–e181. [Google Scholar]
- Bellnier, D.A.; Greco, W.R.; Loewen, G.M.; Nava, H.; Oseroff, A.R.; Pandey, R.K.; Tsuchida, T.; Dougherty, T.J. Population pharmacokinetics of the photodynamic therapy agent 2-[1-hexyloxyethyl]-2-devinyl pyropheophorbide-a in cancer patients. Cancer Res. 2003, 63, 1806–1813. [Google Scholar]
- Rissler, H.M.; Durnford, D.G. Isolation of a novel carotenoid-rich protein in Cyanophora paradoxa that is immunologically related to the light-harvesting complexes of photosynthetic eukaryotes. Plant Cell Physiol. 2005, 46, 416–424. [Google Scholar] [CrossRef]
- Schmidt, V.; Kies, L.; Weber, A. Die Pigmente von Cyanophora paradoxa, Gloechaete wittrockiana und Glaucocystis nostochinearum. Arch. Protistenk 1979, 122, 164–170. [Google Scholar] [CrossRef]
- De Rosso, V.V.; Mercadante, A.Z. Carotenoid composition of two Brazilian genotypes of acerola (Malpighia punicifolia L.) from two harvests. Food Res. Int. 2005, 38, 1073–1077. [Google Scholar] [CrossRef]
- Yamauchi, N.; Harada, K.; Watada, A. In vitro chlorophyll degradation in stored broccoli (Brassica oleracea L. 6ar. italica Plen.) florets. Postharvest Biol. Technol. 1997, 12, 239–245. [Google Scholar] [CrossRef]
- Zapata, M.; Rodriguez, F.; Garrido, J. Separation of chlorophylls and carotenoids from marine phytoplankton: A new HPLC method using a reversed phase C8 column and pyridine-containing mobile phases. Mar. Ecol. Progr. Ser. 2000, 195, 29–45. [Google Scholar] [CrossRef]
- Smith, A.; Witty, M. Heme, Chlorophyll, and Bilins: Methods and Protocols; Springer: Cambridge, UK, 2002. [Google Scholar]
- Hegazi, M.; Perez-Ruzafa, A.; Almela, L.; Candela, M. Separation and identification of chlorophylls and carotenoids from Caulerpa prolifera, Jania rubens and Padina pavonica by reversed-phase high-performance liquid chromatography. J. Chromatogr. A 1998, 829, 153–159. [Google Scholar] [CrossRef]
- Chapman, D.J. The pigments of the symbiotic algae (cyanomes) of Cyanophora paradoxa and Glaucocystis nostochinearum and two Rhodophyceae, Porphyridium aerugineum and Asterocytis ramosa. Arch. Mikrobiol. 1966, 55, 17–25. [Google Scholar] [CrossRef]
- Kies, L.; Kremer, B. Phylum Gaucocystophyta. In Handbook of Protocista; Margulis, L., Corliss, J., Melkonian, M., Chapman, D., Eds.; Jones and Bartlett Publishers: Boston, MA, USA, 1990; pp. 152–166. [Google Scholar]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Testard, A.; Picot, L.; Lozach, O.; Blairvacq, M.; Meijer, L.; Murillo, L.; Piot, J.-M.; Thiéry, V.; Besson, T. Synthesis and evaluation of the antiproliferative activity of novel thiazoloquinazolinone kinases inhibitors. J. Enzyme Inhib. Med. Chem. 2005, 20, 557–568. [Google Scholar] [CrossRef]
- Firdous, A.P.; Sindhu, E.R.; Ramnath, V.; Kuttan, R. Anti-mutagenic and anti-carcinogenic potential of the carotenoid meso-zeaxanthin. Asian Pac. J. Cancer Prev. APJCP 2010, 11, 1795–1800. [Google Scholar]
- Cha, K.H.; Koo, S.Y.; Lee, D.-U. Antiproliferative effects of carotenoids extracted from Chlorella ellipsoidea and Chlorella vulgaris on human colon cancer cells. J. Agric. Food Chem. 2008, 56, 10521–10526. [Google Scholar] [CrossRef]
- Ferreres, F.; Pereira, D.M.; Gil-Izquierdo, A.; Valentão, P.; Botelho, J.; Mouga, T.; Andrade, P.B. HPLC-PAD-atmospheric pressure chemical ionization-MS metabolite profiling of cytotoxic carotenoids from the echinoderm Marthasterias glacialis (spiny sea-star). J. Sep. Sci. 2010, 33, 2250–2257. [Google Scholar] [CrossRef]
Supplementary Files
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Baudelet, P.-H.; Gagez, A.-L.; Bérard, J.-B.; Juin, C.; Bridiau, N.; Kaas, R.; Thiéry, V.; Cadoret, J.-P.; Picot, L. Antiproliferative Activity of Cyanophora paradoxa Pigments in Melanoma, Breast and Lung Cancer Cells. Mar. Drugs 2013, 11, 4390-4406. https://doi.org/10.3390/md11114390
Baudelet P-H, Gagez A-L, Bérard J-B, Juin C, Bridiau N, Kaas R, Thiéry V, Cadoret J-P, Picot L. Antiproliferative Activity of Cyanophora paradoxa Pigments in Melanoma, Breast and Lung Cancer Cells. Marine Drugs. 2013; 11(11):4390-4406. https://doi.org/10.3390/md11114390
Chicago/Turabian StyleBaudelet, Paul-Hubert, Anne-Laure Gagez, Jean-Baptiste Bérard, Camille Juin, Nicolas Bridiau, Raymond Kaas, Valérie Thiéry, Jean-Paul Cadoret, and Laurent Picot. 2013. "Antiproliferative Activity of Cyanophora paradoxa Pigments in Melanoma, Breast and Lung Cancer Cells" Marine Drugs 11, no. 11: 4390-4406. https://doi.org/10.3390/md11114390