Toxic C17-Sphinganine Analogue Mycotoxin, Contaminating Tunisian Mussels, Causes Flaccid Paralysis in Rodents

 and
and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
2.1. Results
2.1.1. Acute Mouse Toxicity
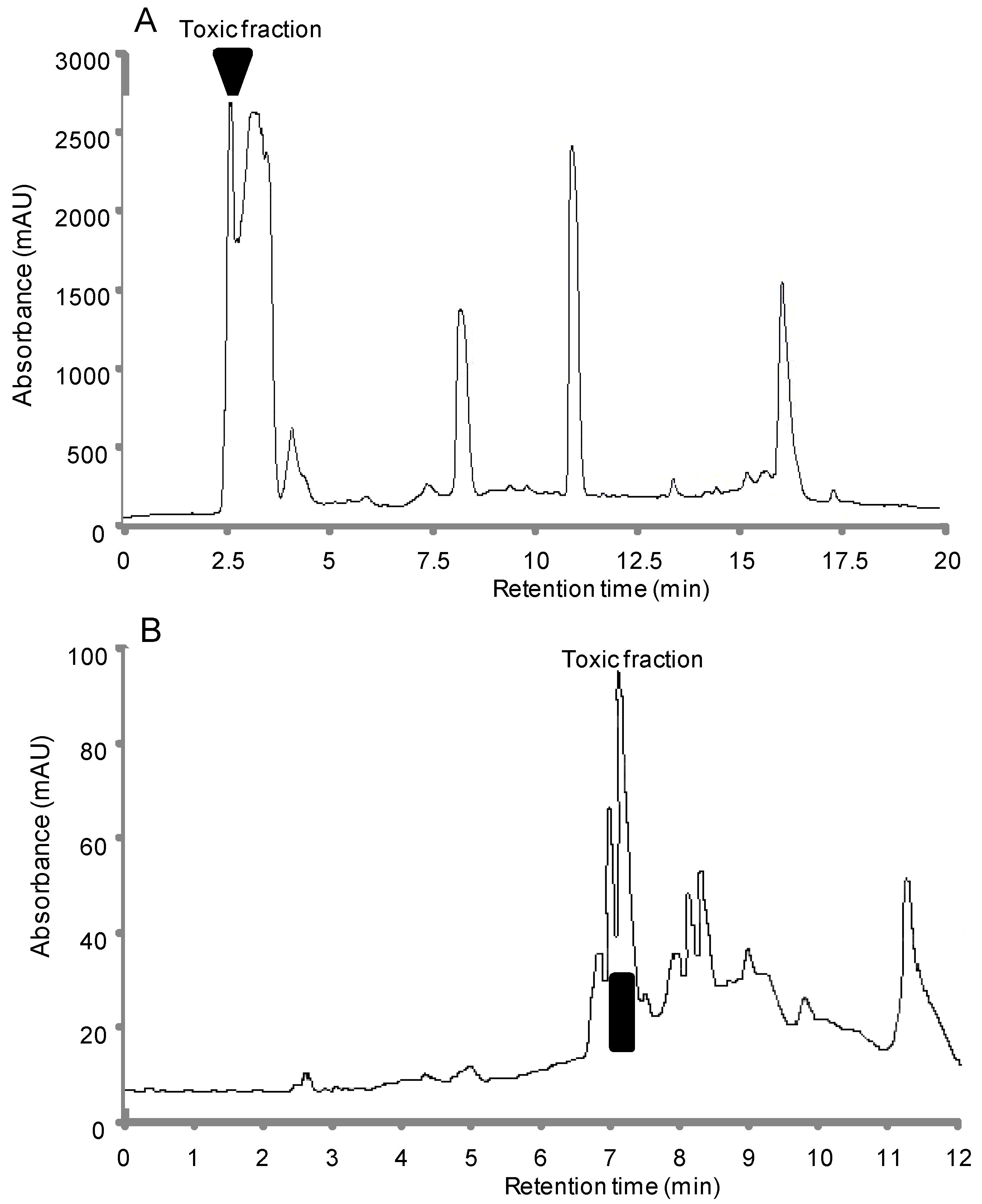
2.1.2. Bioassay-Guided Chromatographic Separation
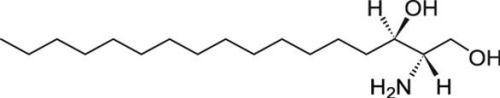
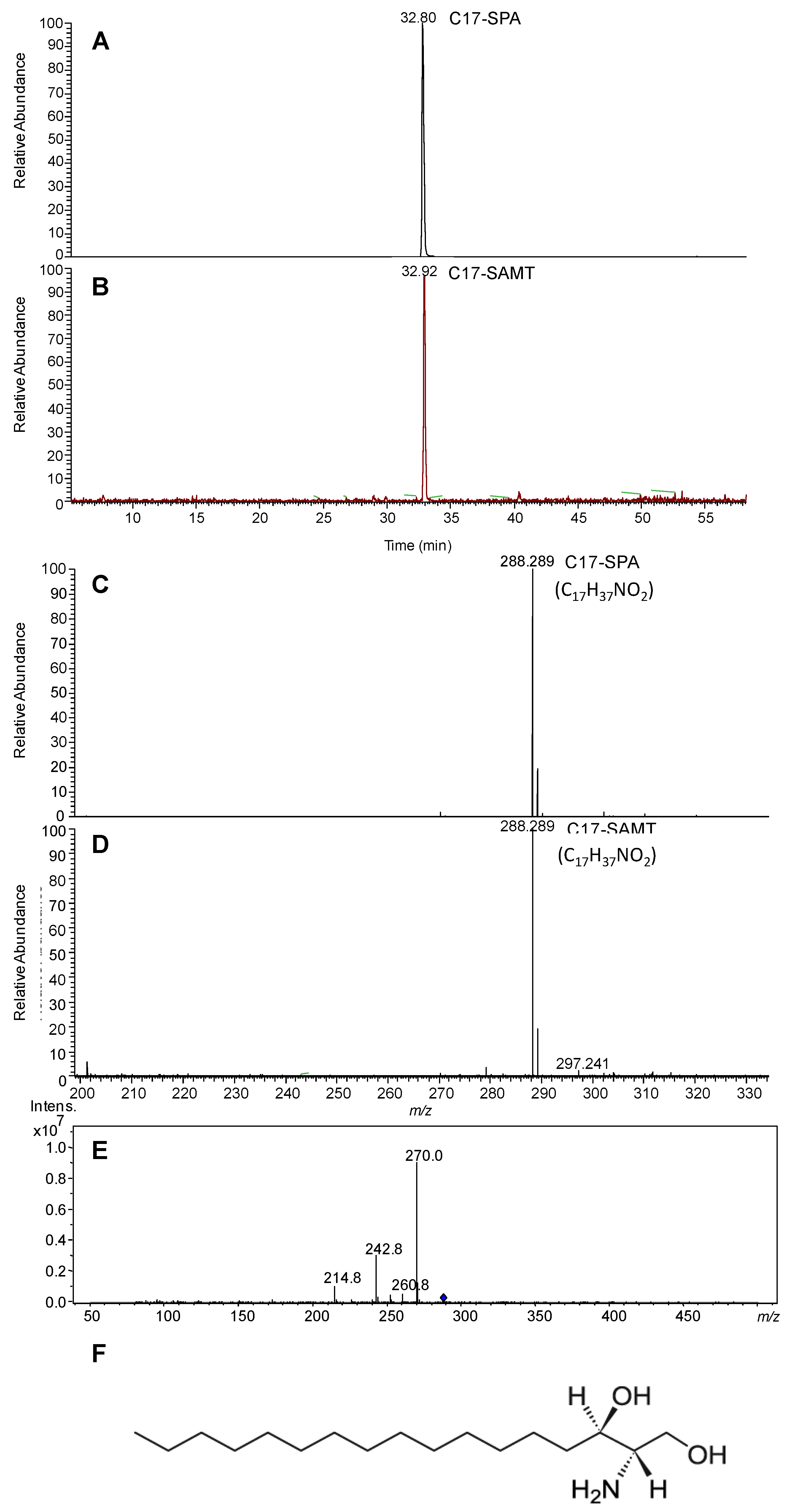
2.1.3. Mass Spectrometry Characterization


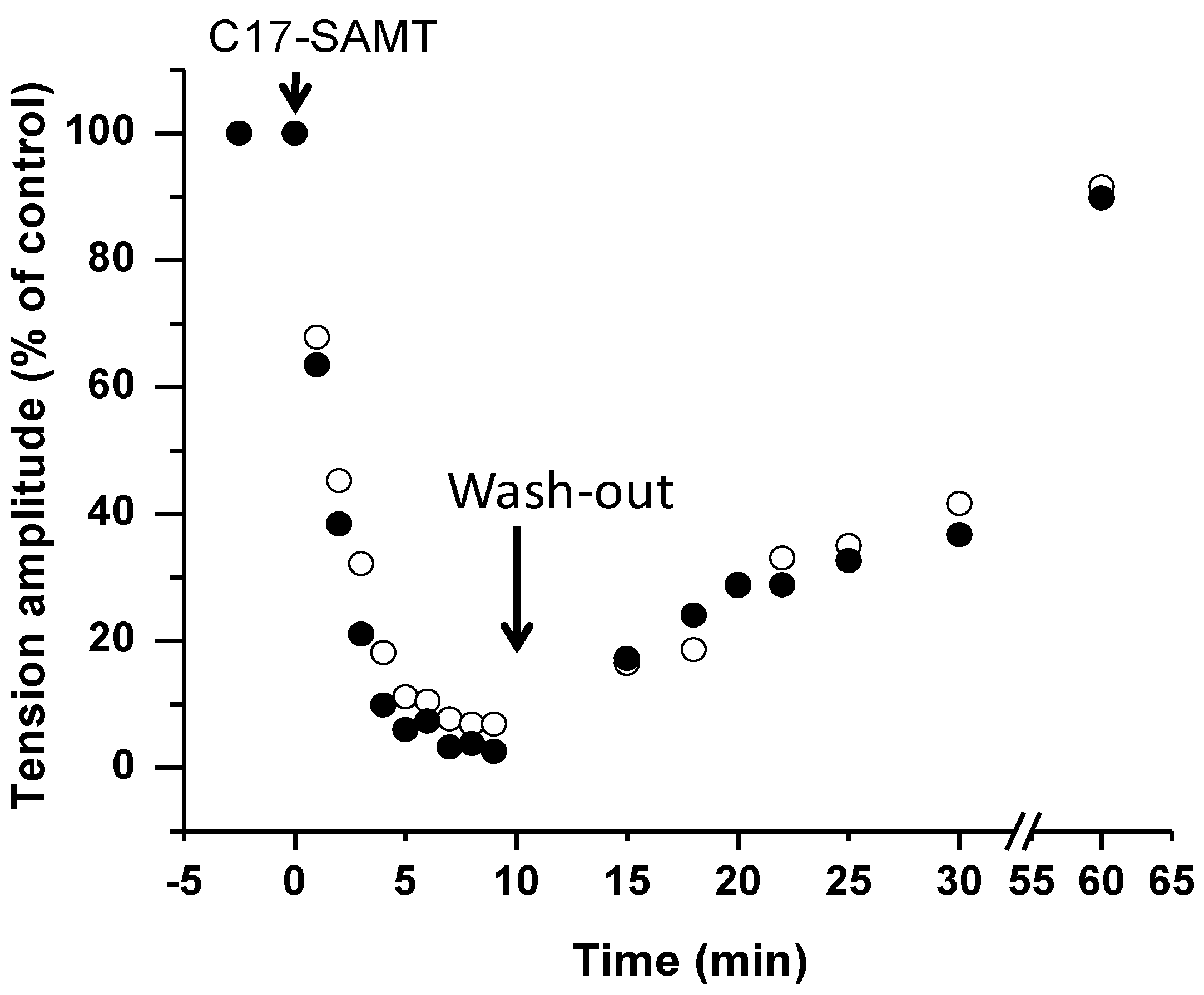
2.1.4. Toxicity of C17-SAMT
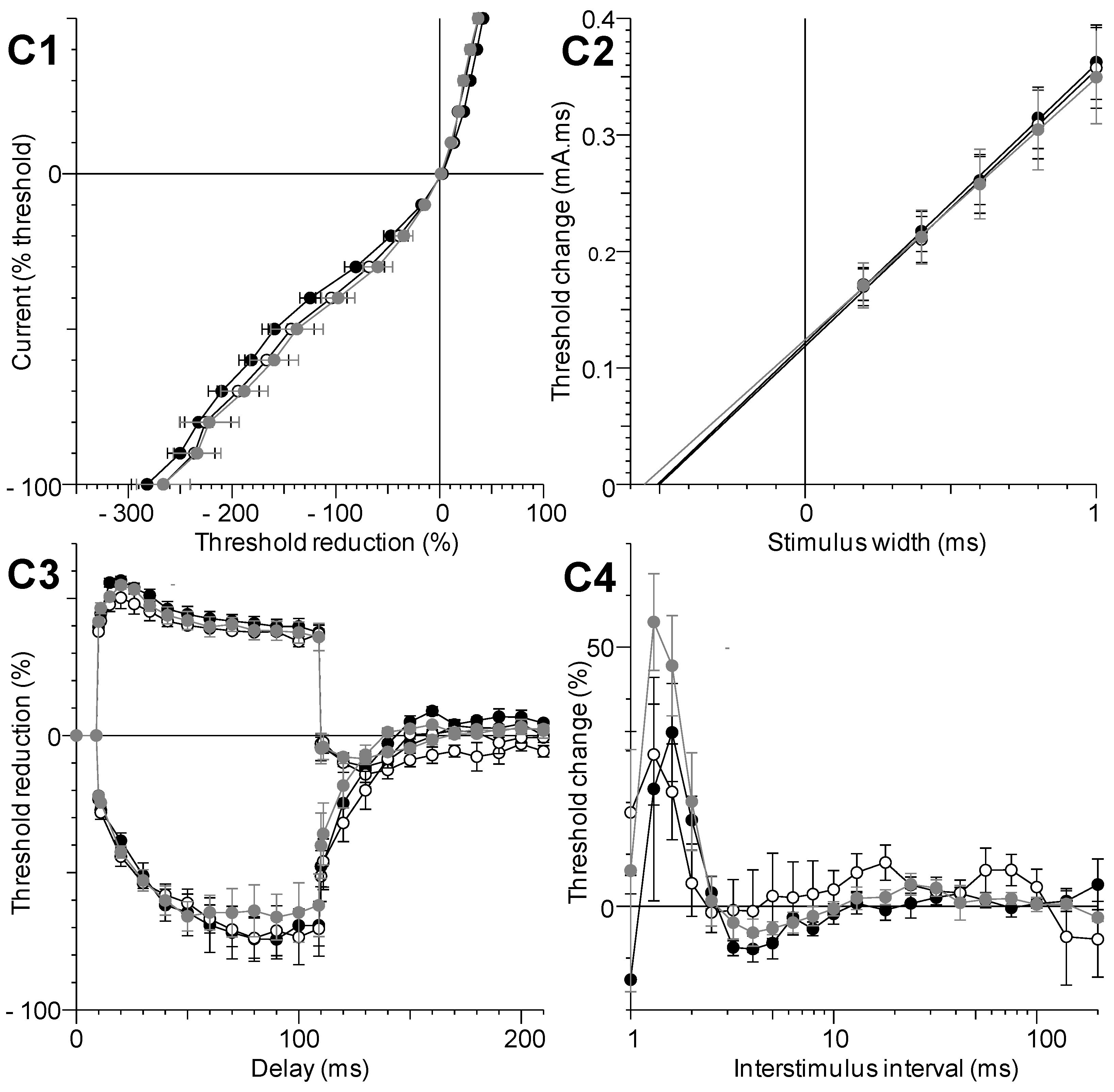
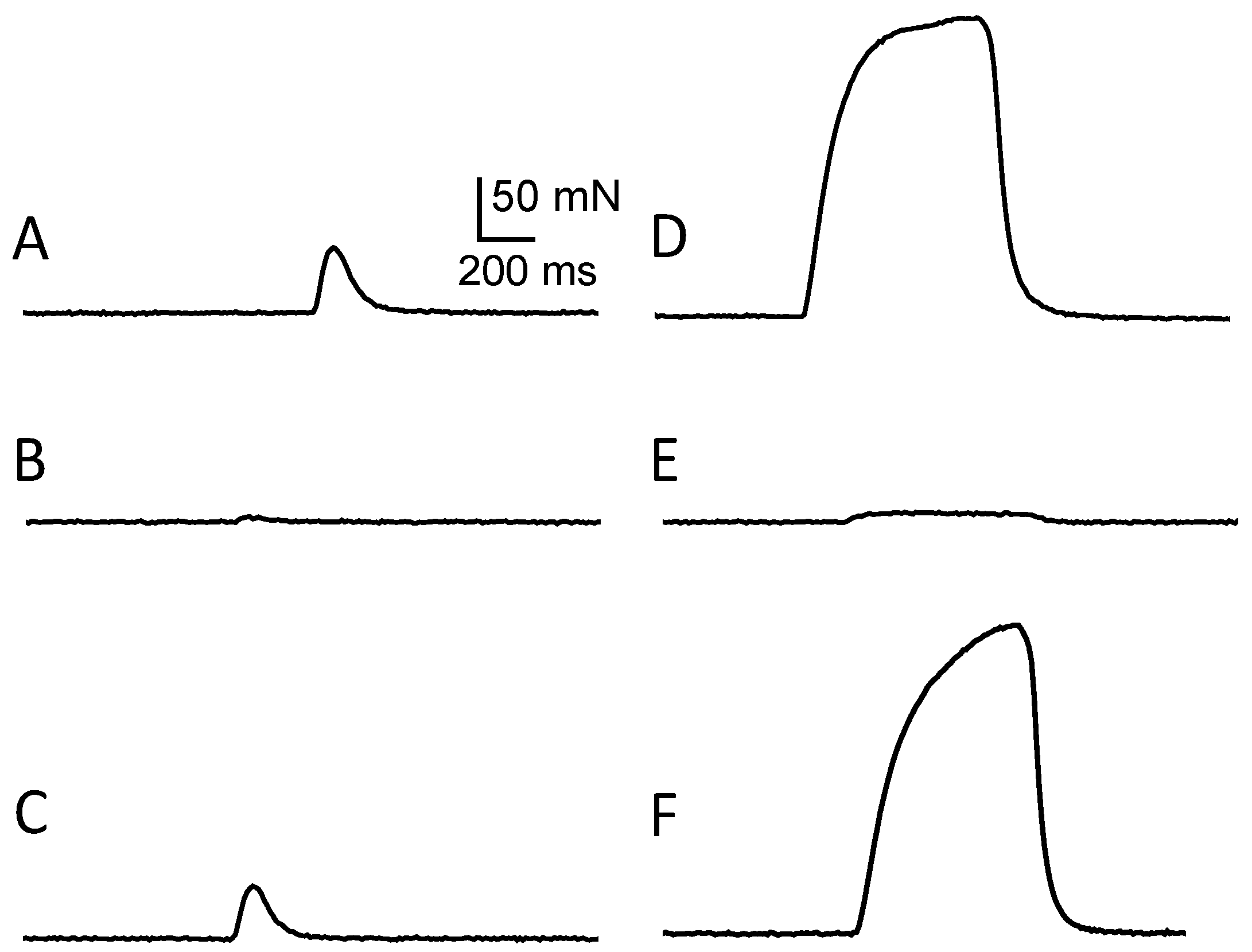
2.1.5. Local in Vivo Effects of C17-SAMT


2.1.6. In Vitro Effects of C17-SAMT


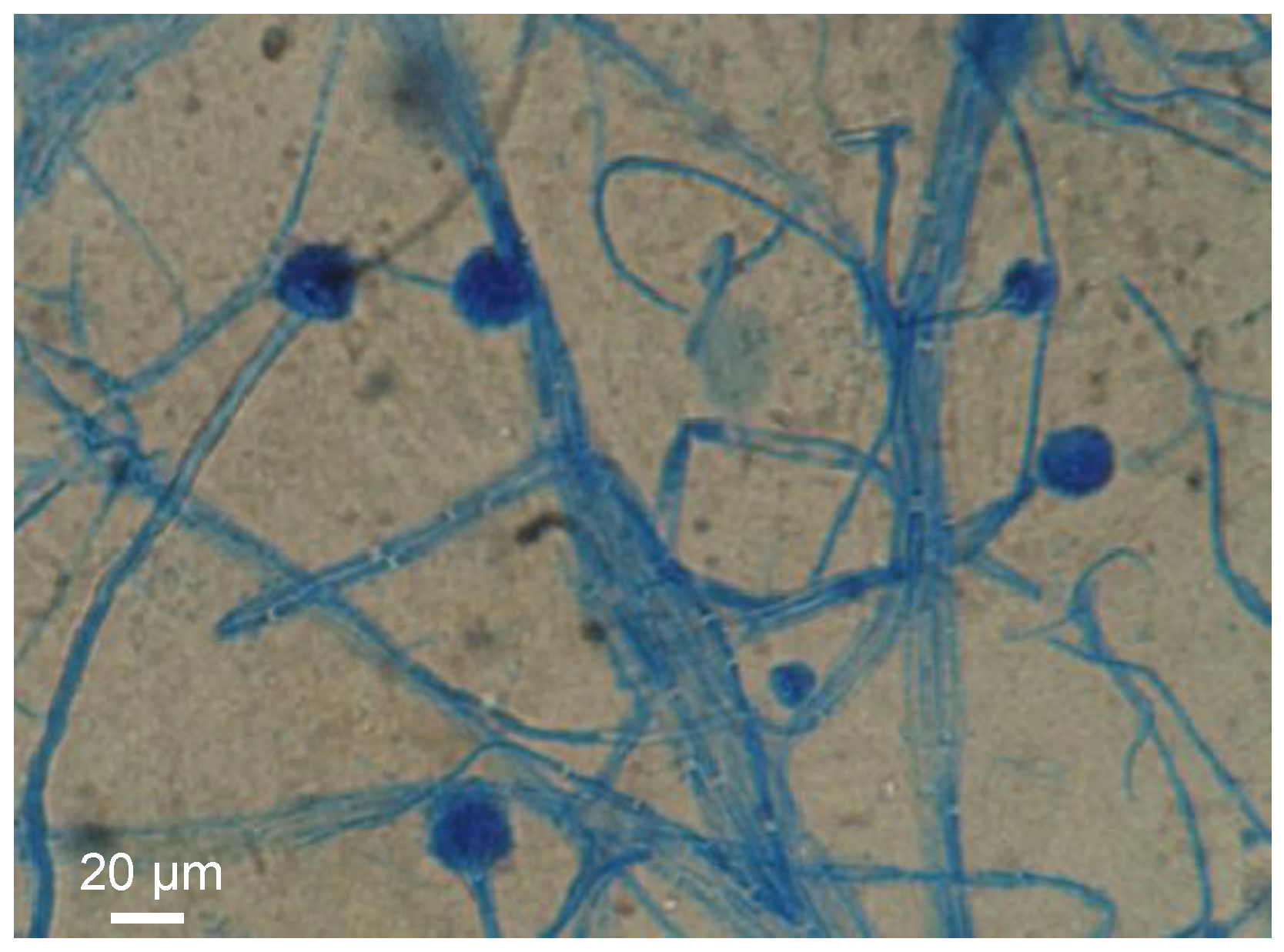
2.1.7. Causative Organism(s)

2.2. Discussion
3. Experimental Section
3.1. Materials
3.2. Methods
3.2.1. Sample Extraction and Solvent Partition
3.2.2. Toxicity Assays
3.2.3. Chromatographic Analysis
3.2.4. HPLC-ESI-LC Analysis
3.2.5. Determination of Toxin Concentration by LC-MSD-Trap-XCT
3.2.6. In Vivo Study on the Mouse Neuromuscular System
3.2.7. Twitch Tension Recordings on Isolated Mouse Neuromuscular Preparations
3.2.8. Fungal Culture and Strain Identification
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Smith, J.L.; Boyer, G.L.; Zimba, P.V. A review of cyanobacterial odorous and bioactive metabolites: Impacts and management alternatives in aquaculture. Aquaculture 2008, 280, 5–20. [Google Scholar] [CrossRef]
- Zaccaroni, A.; Scaravelli, D. Toxicity of Sea Algal Toxins to Human and Animal. In Algal Toxins-Nature Occurrence and Detection; Springer Netherlands: Heidelberg, The Netherlands, 2008; pp. 91–158. [Google Scholar]
- Krys, S.; Arnich, N.; Fremy, J.M. Emergence de nouvelles toxicités d’origine marine: Vers une évolution significative des dispositifs de sécurisation des coquillages? In Rencontres en Toxinologie, Toxines Émergentes: Nouveaux Risques; Lavoisier: Paris, France, 2007; pp. 117–122. [Google Scholar]
- Sallenave-Namont, C.; Pouchus, Y.F.; Robiou du Pont, T.; Lassus, P.; Verbist, J.F. Toxigenic saprophytic fungi in marine shellfish farming areas. Mycopathologia 2000, 149, 21–25. [Google Scholar] [CrossRef]
- Abbas, H.K.; Duke, S.O.; Shier, W.T.; Riley, R.T.; Kraus, G.A. The chemistry and biological activities of the natural products AAL toxin and the fumonisins. Adv. Exp. Med. Biol. 1996, 391, 293–308. [Google Scholar]
- Sweeney, M.J.; Dobson, A.D. Molecular biology of mycotoxin biosynthesis. FEMS Microbiol. Lett. 1999, 175, 149–163. [Google Scholar] [CrossRef]
- Desjardins, A.E.; Proctor, R.H. Molecular biology of Fusarium mycotoxins. Int. J. Food Microbiol. 2007, 119, 47–50. [Google Scholar] [CrossRef]
- Biré, R.; Krys, S.; Fremy, J.M.; Dragacci, S.; Stirling, D.; Kharrat, R. First evidence on occurrence of gymnodimine in clams from Tunisia. J. Nat. Toxins 2002, 11, 269–275. [Google Scholar]
- Brandwagt, B.F.; Mesbah, L.A.; Takken, F.L.; Laurent, P.L.; Kneppers, T.J.; Hille, J.; Nijkamp, H.J. A longevity assurance gene homolog of tomato mediates resistance to Alternaria alternata f. sp. lycopersici toxins and fumonisin B1. Proc. Natl. Acad. Sci. USA 2000, 97, 4961–4966. [Google Scholar]
- Cestèle, S.; Catterall, W.A. Molecular mechanisms of neurotoxin action on voltage-gated sodium channels. Biochimie 2000, 82, 883–892. [Google Scholar]
- Schantz, E.J.; Ghazarossian, V.E.; Schnoes, H.K.; Strong, F.M.; Springer, J.P.; Pezzanite, J.O.; Clardy, J. The structure of saxitoxin. J. Am. Chem. Soc. 1975, 97, 1238–1239. [Google Scholar]
- Uhliga, S.; Petersen, D.; Flaøyen, A.; Wilkins, A. 2-Amino-14,16-dimethyloctadecan-3-ol, a new sphingosine analogue toxin in the fungal genus Fusarium. Toxicon 2005, 46, 513–522. [Google Scholar]
- Degenkolb, T.; Dieckmann, R.; Nielsen, K.F.; Gräfenhan, T.; Theis, C.; Zafari, D.; Chaverri, P.; Ismaiel, A.; Brückner, H.; Döhren, H.; et al. The Trichoderma brevicompactum clade: A separate lineage with new species, new peptaibiotics, and mycotoxins. Mycol. Prog. 2008, 7, 177–219. [Google Scholar] [CrossRef]
- Matallah-Boutiba, A.; Ruiz, N.; Sallenave-Namont, C.; Grovel, O.; Amiard, J.C.; Pouchus, Y.F.; Boutiba, Z. Screening for toxigenic marine-derived fungi in Algerian mussels and their immediate environment. Aquaculture 2012, 342–343, 75–79. [Google Scholar]
- Arvanitidou, M.; Kanellou, K.; Katsouyannopoulos, V.; Tsakris, A. Occurrence and densities of fungi from northern Greek coastal bathing waters and their relation with faecal pollution indicators. Water Res. 2002, 36, 5127–5131. [Google Scholar] [CrossRef]
- Salvo, V.S.; Fabiano, M. Mycological assessment of sediments in Ligurian beaches in the Northwestern Mediterranean: Pathogens and opportunistic pathogens. J. Environ. Manag. 2007, 83, 365–369. [Google Scholar] [CrossRef]
- Sallenave, C.; Pouchus, Y.F.; Bardouil, M.; Lassus, P.; Roquebert, M.F.; Verbist, J.F. Bioaccumulation of mycotoxins by shellfish: Contamination of mussels by metabolites of a Trichoderma koningii strain isolated in the marine environment. Toxicon 1999, 37, 77–83. [Google Scholar]
- Zvereva, L.V.; Vysotskaya, M.A. Filamentous fungi associated with bivalve mollusks from polluted biotopes of Usuriiskii Bay, Sea of Japan. Russ. J. Mar. Biol. 2005, 31, 382–385. [Google Scholar] [CrossRef]
- Mohamed-Benkada, M. Evaluation du Risque Fongique en Zone Conchylicoles: Substances Toxiques de Souche Marines du Genre Trichoderma. In Thèse de Doctorat; Université de Nantes: Nantes, France, 2006; p. 9. [Google Scholar]
- Zielinski, O.; Bush, J.A.; Cembella, A.D.; Daly, K.L.; Engel Bbrektsson, J.; Hannides, A.K.; Schmidt, H. Detecting marine hazardous substances and organisms: Sensors for pollutants, toxins and pathogens. Ocean Sci. 2009, 6, 953–1005. [Google Scholar]
- Petit, K.E.; Mondeguer, F.; Roquebert, M.F.; Biard, J.F.; Pouchus, Y.F. Detection of griseofulvin in a marine strain of Penicillium waksmanii by ion trap mass spectrometry. J. Microbiol. Methods 2004, 58, 59–65. [Google Scholar]
- Kerzaon, I.; Pouchus, Y.F.; Monteau, F.; Le Bizec, B.; Nourrisson, M.R.; Biard, J.F.; Grovel, O. Structural investigation and elucidation of new communesins from a marine-derived Penicillium expansum link by liquid chromatography/electrospray ionization mass spectrometry. Rapid Commun. Mass Spectrom. 2009, 23, 3928–3938. [Google Scholar]
- Grovel, O.; Pouchus, Y.F.; Verbist, J.F. Accumulation of gliotoxin, a cytotoxic mycotoxin from Aspergillus fumigatus, in blue mussel (Mytilus edulis). Toxicon 2003, 42, 297–300. [Google Scholar] [CrossRef]
- De Respinis, S.; Vogel, G.; Benagli, C.; Tonolla, M.; Petrini, O.; Samuels, J.G. MALDI-TOF MS of Trichoderma: A model system for the identification of microfungi. Mycol. Prog. 2010, 9, 79–100. [Google Scholar] [CrossRef]
- Li, D.; Sun, L.; Chen, Z.; He, X.; Lin, B. Survey of the distribution of red tide toxins okadaic acid and dinophysistoxin-1 in the Dalian Bay sea area of China by micellar electrokinetic capillary chromatography. Electrophoresis 2001, 22, 3583–3588. [Google Scholar]
- Van Buynder, P.G.; Oughtred, T.; Birkby, B.; Phillips, S.; Eaglesham, G.; Thomas, K.; Burch, M. Nodularin uptake by seafood during a cyanobacterial bloom. Environ. Toxicol. 2001, 16, 468–471. [Google Scholar] [CrossRef]
- Kharrat, R.; Servent, D.; Girard, E.; Ouanounou, G.; Amar, M.; Marrouchi, R.; Benoit, E.; Molgó, J. The marine phycotoxin gymnodimine targets muscular and neuronal nicotinic acetylcholine receptor subtypes with high affinity. J. Neurochem. 2008, 21, 952–963. [Google Scholar]
- Kodama, M. Paralytic shellfish poisoning toxins: Biochemistry and origin. Aqua-Biosci. Monogr. 2010, 3, 1–38. [Google Scholar] [CrossRef]
- Marrouchi, R.; Dziri, F.; Belayouni, N.; Hamza, A.; Benoit, E.; Molgó, J.; Kharrat, R. Quantitative determination of gymnodimine-A by high performance liquid chromatography in contaminated clams from Tunisia coastline. Mar. Biotechnol. 2009, 12, 579–585. [Google Scholar]
- Yasumoto, T.; Oshima, Y.; Yamaguchi, M. Occurrence of a new type of toxic shellfish poisoning in the Tohoku district. Bull. Jpn. Soc. Sci. Fish 1978, 44, 1249–1255. [Google Scholar] [CrossRef]
- Boërio, D.; Greensmith, L.; Bostock, H. Excitability properties of motor axons in the maturing mouse. J. Peripher. Nerv. Syst. 2009, 14, 45–53. [Google Scholar]
- Boërio-Guéguen, D.; Lefaucheur, J.P.; Créange, A.; Benoit, E. A Non-Invasive Method to Appraise Time-Dependent Effects of Toxins on the Mouse Neuromuscular Excitability in Vivo, and Its Clinical Applications. Toxins and Signalling; Benoit, E., Goudey-Perrière, F., Marchot, P., Servent, D., Eds.; SFET Publications: Châtenay-Malabry, France, 2009; pp. 123–130. Available online: http://www.sfet.asso.fr (ISSN 1760-6004) (accessed on 5 January 2010).
- Kiernan, M.C.; Burke, D.; Andersen, K.V.; Bostock, H. Multiple measures of axonal excitability: A new approach in clinical testing. Muscle Nerve 2000, 23, 399–409. [Google Scholar]
- Lefaucheur, J.P.; Boërio, D.; Hogrel, J.Y.; Créange, A. Nerve excitability studies in the assessment of dysimmune neuropathies. Rev. Neurol. (Paris) 2006, 162, 17–26. [Google Scholar]
- Krishnan, A.; Park, S.; Lin, C.S.; Kiernan, M.C. Assessment of nerve excitability in toxic and metabolic neuropathies. J. Peripher. Nerv. Syst. 2008, 13, 7–26. [Google Scholar] [CrossRef]
- Schlumberger, S.; Ouanounou, G.; Girard, E.; Sasaki, M.; Fuwa, H.; Louzao, M.C.; Botana, L.M.; Benoit, E.; Molgó, J. The marine polyether gambierol enhances muscle contraction and blocks a transient K(+) current in skeletal muscle cells. Toxicon 2010, 56, 785–791. [Google Scholar] [CrossRef]
- Pitt, J.I. The Genus Penicillium and Its Teleomorphic States Eupenicillium and Talaromyces; Academic Press: London, UK, New York, NY, USA, 1979; p. 634. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Marrouchi, R.; Benoit, E.; Le Caer, J.-P.; Belayouni, N.; Belghith, H.; Molgó, J.; Kharrat, R. Toxic C17-Sphinganine Analogue Mycotoxin, Contaminating Tunisian Mussels, Causes Flaccid Paralysis in Rodents. Mar. Drugs 2013, 11, 4724-4740. https://doi.org/10.3390/md11124724
Marrouchi R, Benoit E, Le Caer J-P, Belayouni N, Belghith H, Molgó J, Kharrat R. Toxic C17-Sphinganine Analogue Mycotoxin, Contaminating Tunisian Mussels, Causes Flaccid Paralysis in Rodents. Marine Drugs. 2013; 11(12):4724-4740. https://doi.org/10.3390/md11124724
Chicago/Turabian StyleMarrouchi, Riadh, Evelyne Benoit, Jean-Pierre Le Caer, Nawel Belayouni, Hafedh Belghith, Jordi Molgó, and Riadh Kharrat. 2013. "Toxic C17-Sphinganine Analogue Mycotoxin, Contaminating Tunisian Mussels, Causes Flaccid Paralysis in Rodents" Marine Drugs 11, no. 12: 4724-4740. https://doi.org/10.3390/md11124724