PTP1B Inhibitory and Anti-Inflammatory Effects of Secondary Metabolites Isolated from the Marine-Derived Fungus Penicillium sp. JF-55

Abstract
:1. Introduction
2. Results and Discussion
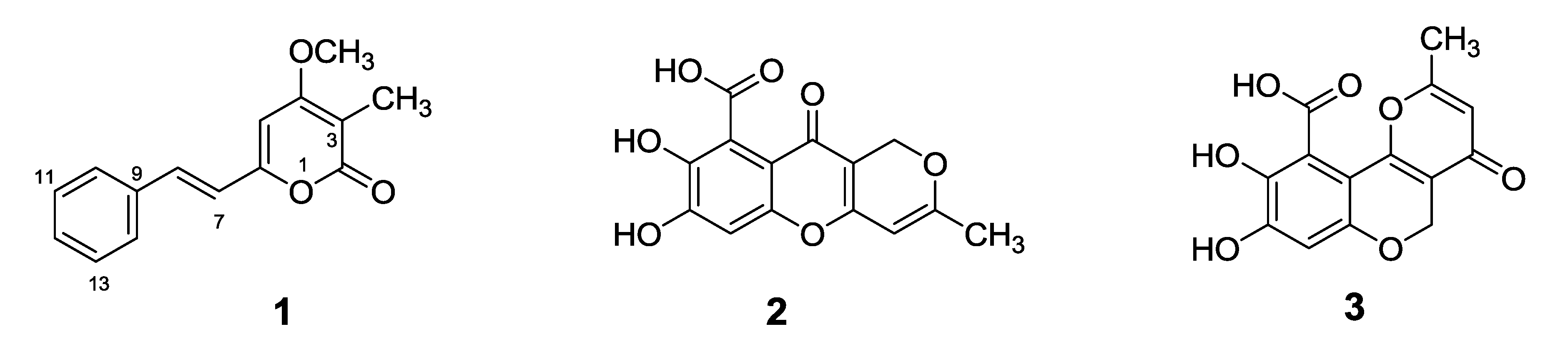
2.1. Chemical Structures of Compounds 1–3 Isolated from the Marine-Derived Fungus Penicillium sp. JF-55


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | δH (int, mult., J in Hz) a | δC b | HMBC (H → C) |
|---|---|---|---|
| 2 | -- | 163.4 | -- |
| 3 | -- | 101.0 | -- |
| 4 | -- | 165.8 | -- |
| 5 | 6.76 (1H, s) | 97.2 | 3, 4, 6, 7 |
| 6 | -- | 156.7 | -- |
| 7 | 7.04 (1H, d, 16.1) | 119.9 | 5, 6, 9 |
| 8 | 7.34 (1H, d, 16.1) | 133.4 | 6, 9, 14 |
| 9 | -- | 135.2 | -- |
| 10/14 | 7.65 (2H, d, 7.3) | 127.3 | 8, 12, 14 |
| 11/13 | 7.42 (2H, t, 7.3) | 128.9 | 9, 12, 14 |
| 12 | 7.36 (1H, t, 7.3) | 129.2 | 14 |
| 15 | 1.83 (3H, s) | 8.8 | 2, 3, 4 |
| 4-OCH3 | 3.93 (3H, s) | 56.7 | 4 |
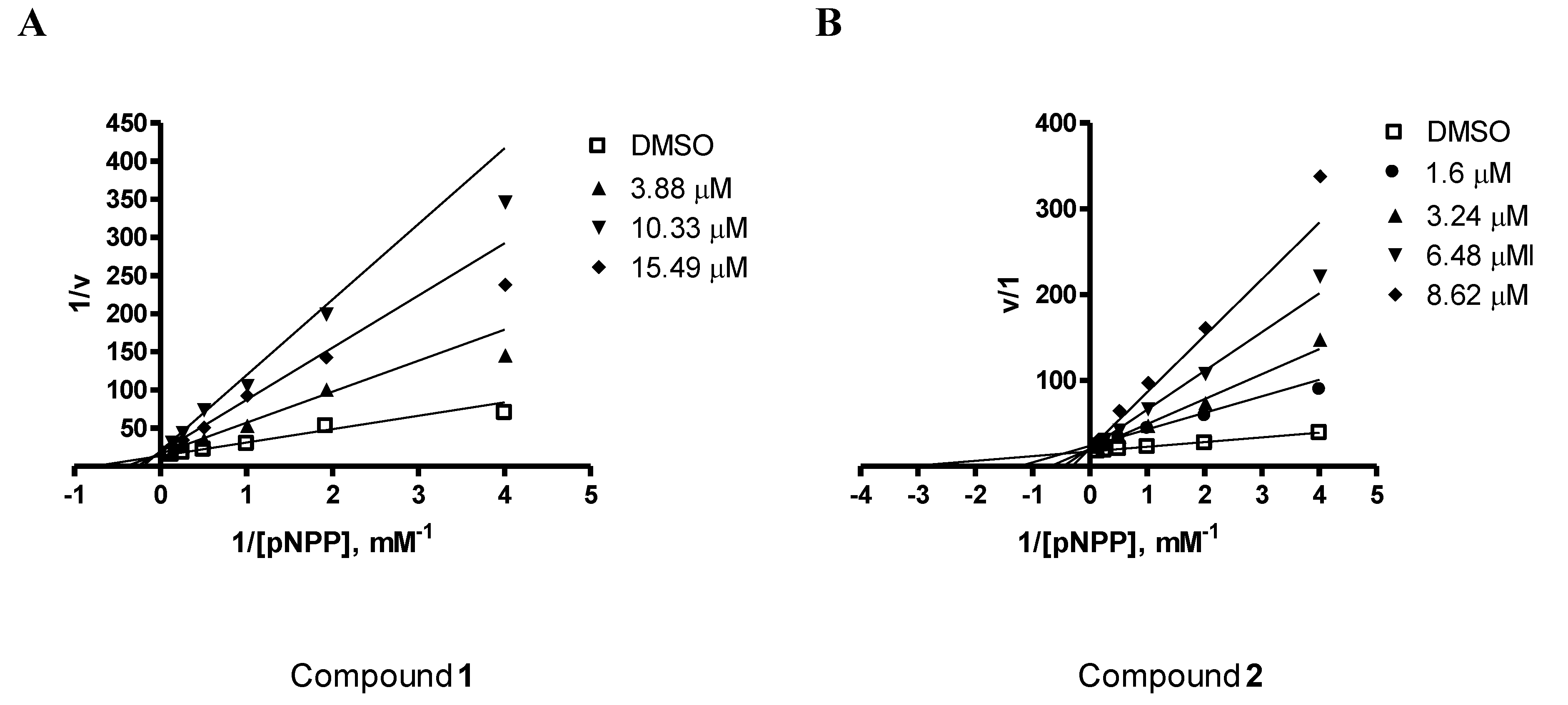
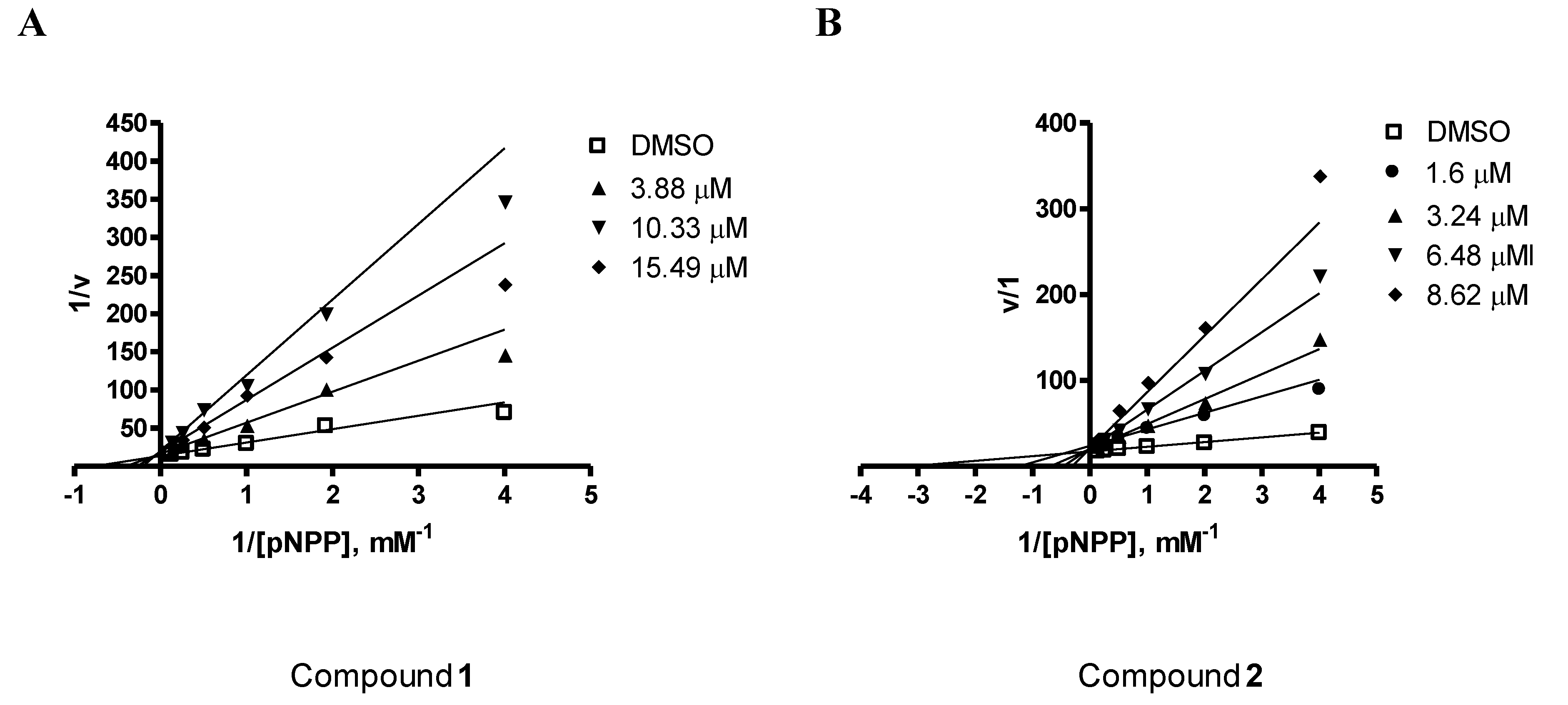
2.2. PTP1B Inhibitory Effects of Compounds 1–3
| Compounds | IC50 |
|---|---|
| Ursolic acid | 3.10 μM a |
| Penstyrylpyrone (1) | 5.28 μM |
| Anhydrofulvic acid (2) | 1.90 μM |
| Citromycetin (3) | >25.8 μM |
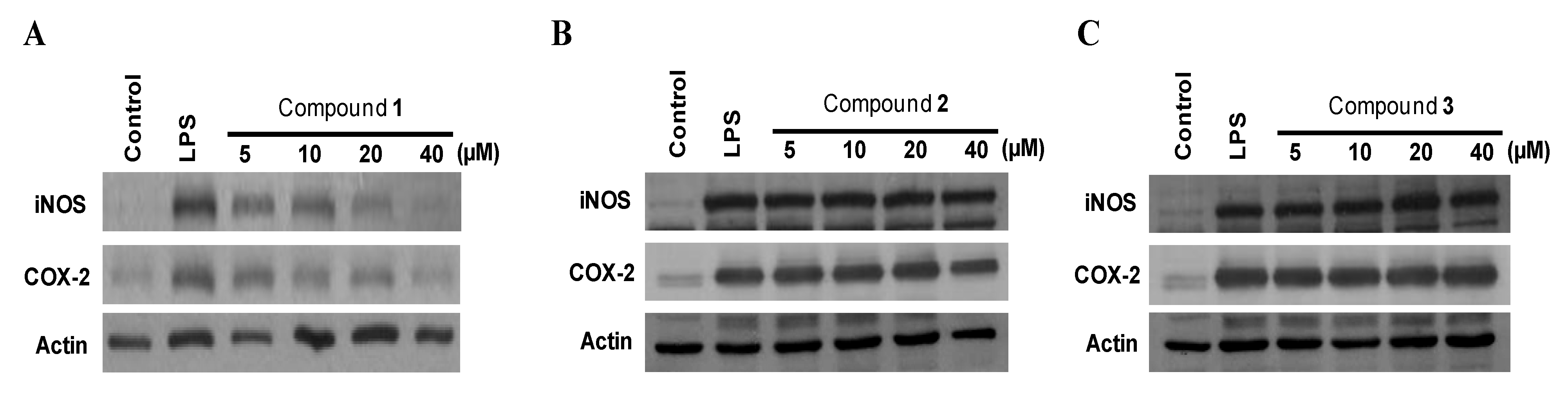
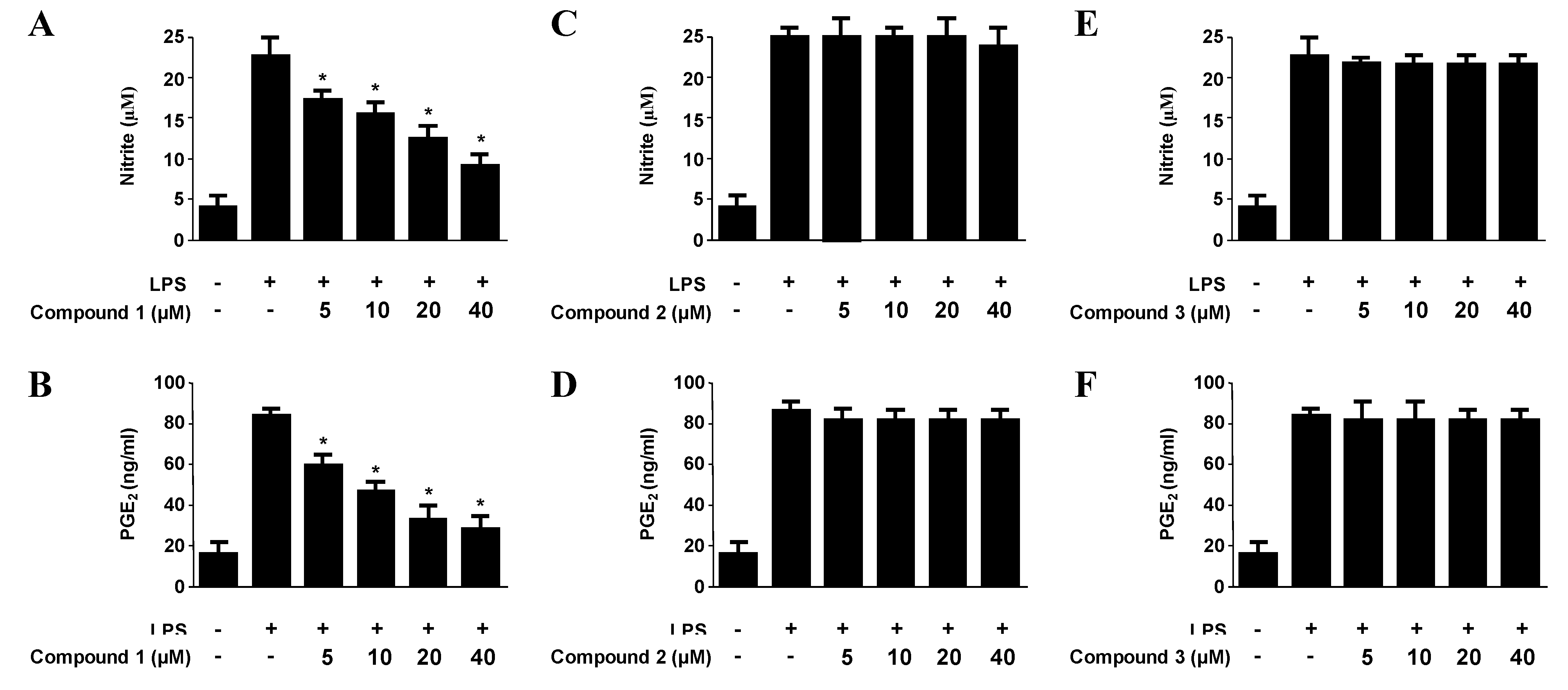
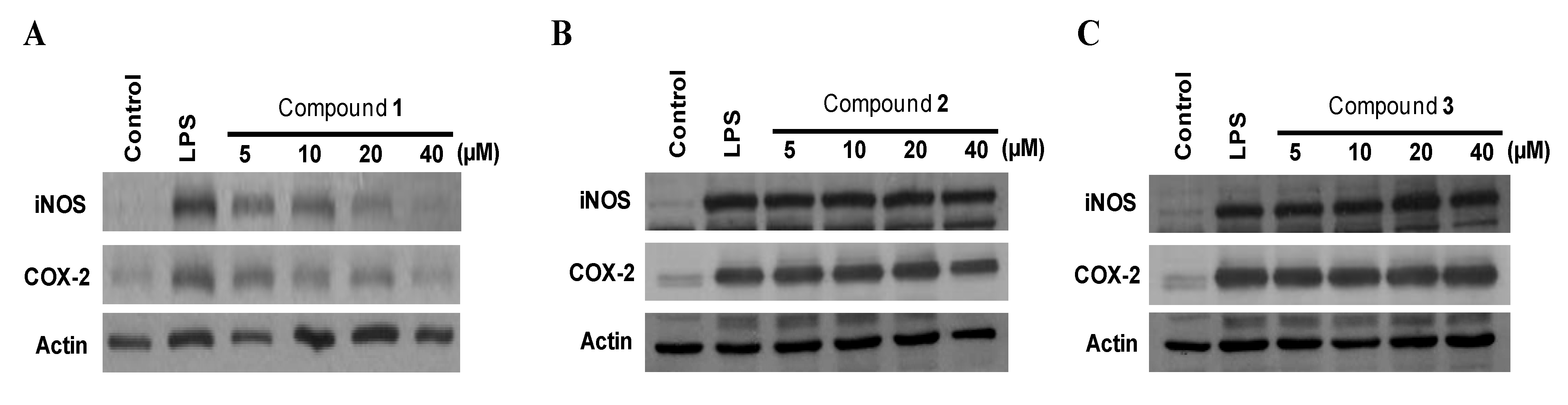
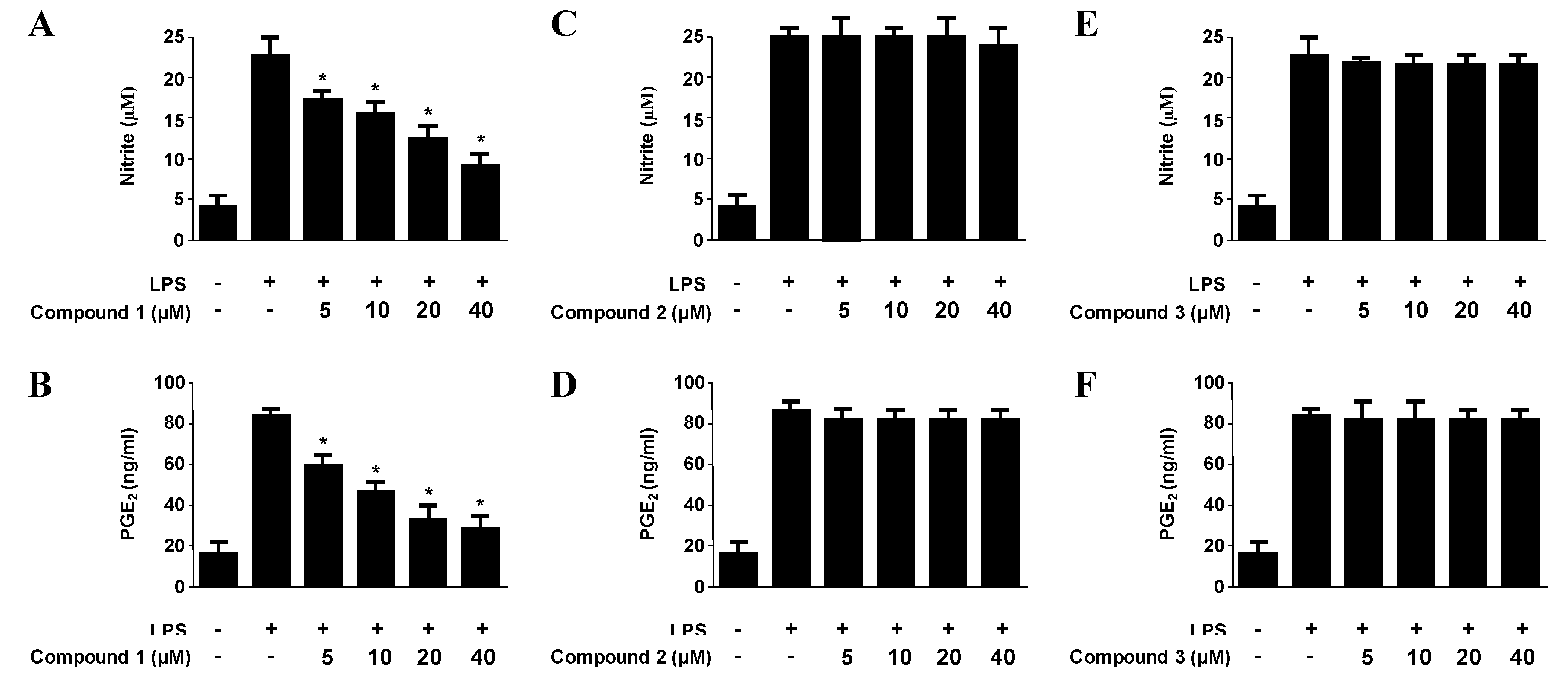
2.3. Effects of Compounds 1–3 on the Expression of Pro-Inflammatory Proteins and Production of Pro-Inflammatory Cytokines in Murine Peritoneal Macrophages Stimulated with Lipopolysaccharides (LPS)
| Compounds | NO production | PGE2 production | IL-1β production | TNF-α production |
|---|---|---|---|---|
| Penstyrylpyrone (1) | 12.32 μM | 9.35 μM | 13.54 μM | 18.32 μM |
| Anhydrofulvic acid (2) | >40 μM | >40 μM | >40 μM | >40 μM |
| Citromycetin (3) | >40 μM | >40 μM | >40 μM | >40 μM |
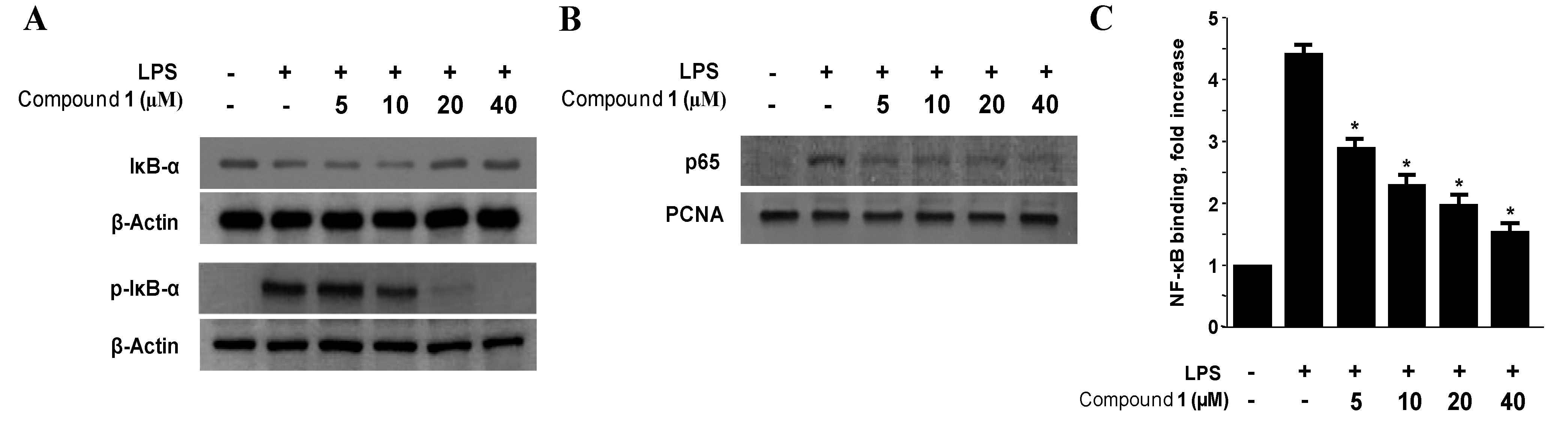
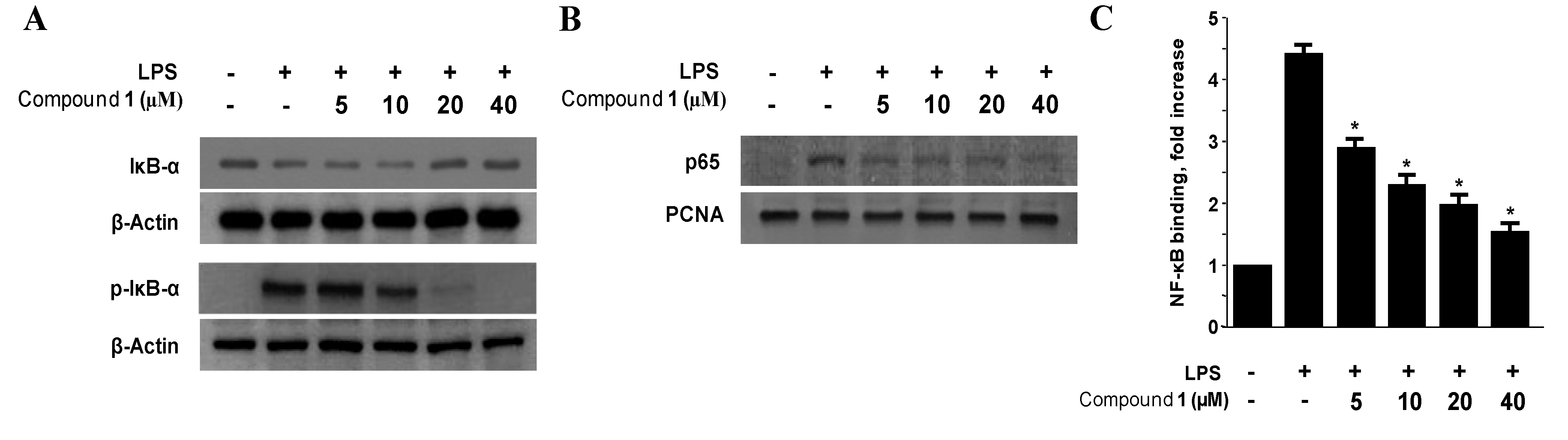
2.4. Effects of Penstyrylpyrone (1) on the Protein Expression Levels of IκB-α Phosphorylation and Degradation as well as NF-κB Translocation and DNA Binding Activity in Murine Peritoneal Macrophages
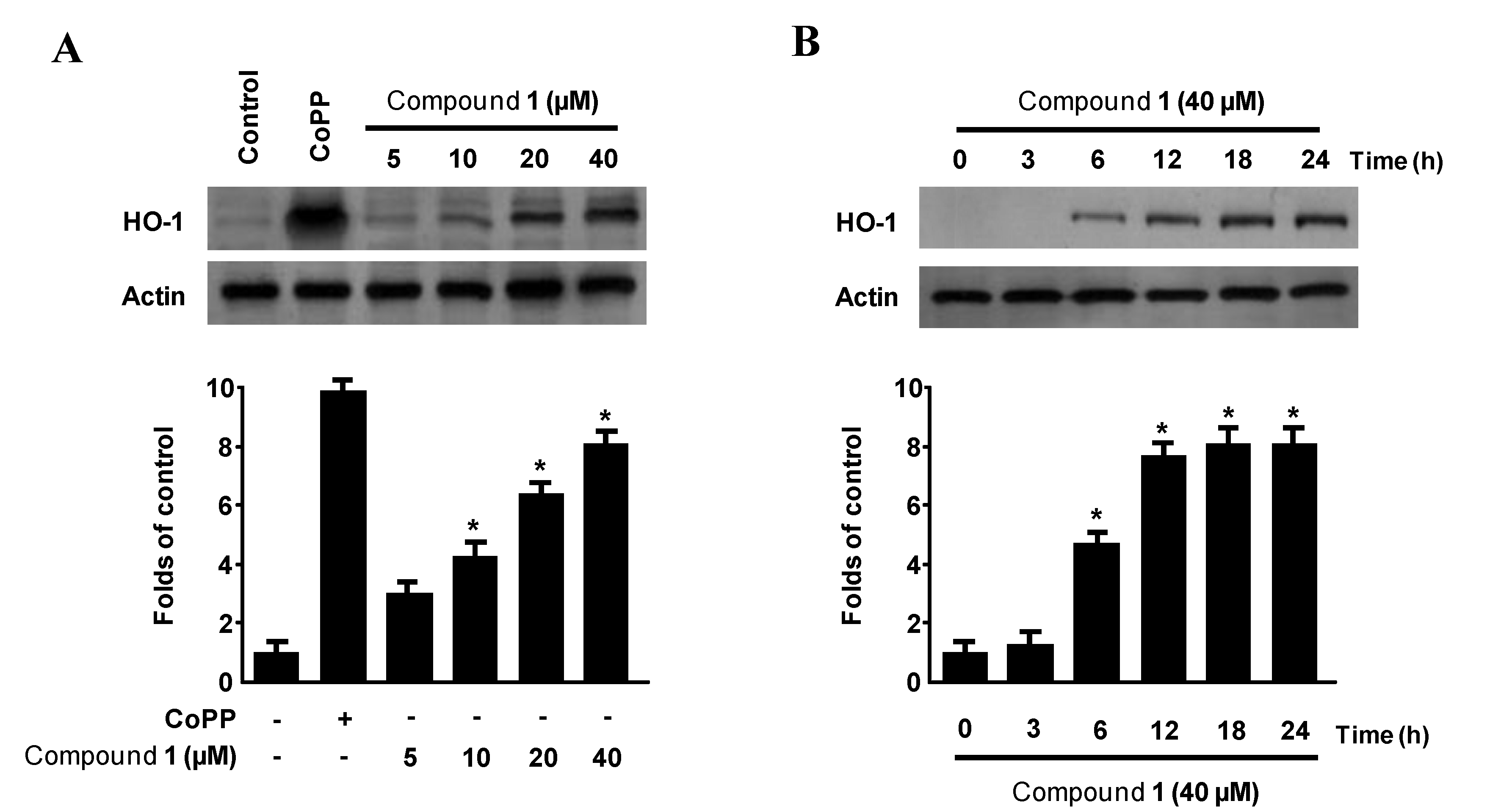
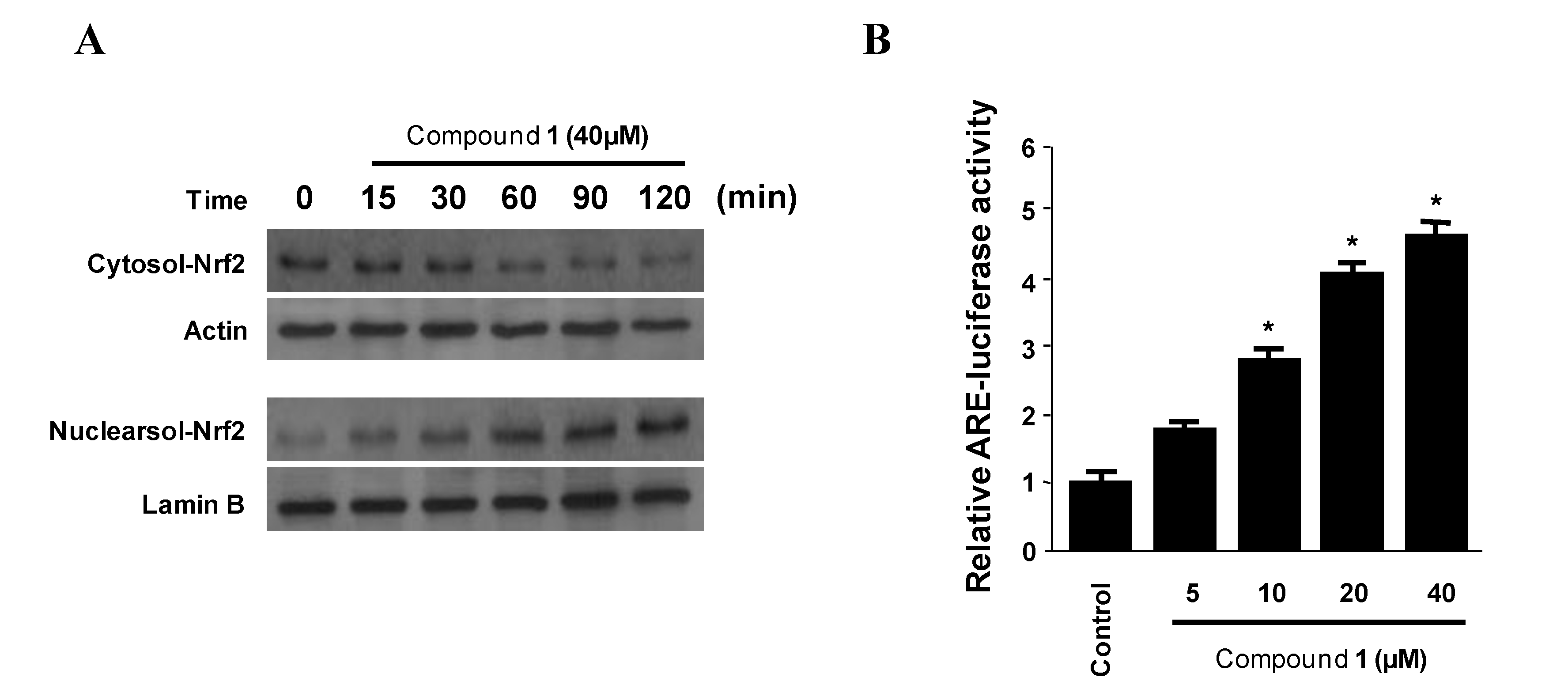
2.5. Effects of Penstyrylpyrone (1) on HO-1 Expression via Nuclear Translocation of Nrf2 in Murine Peritoneal Macrophages
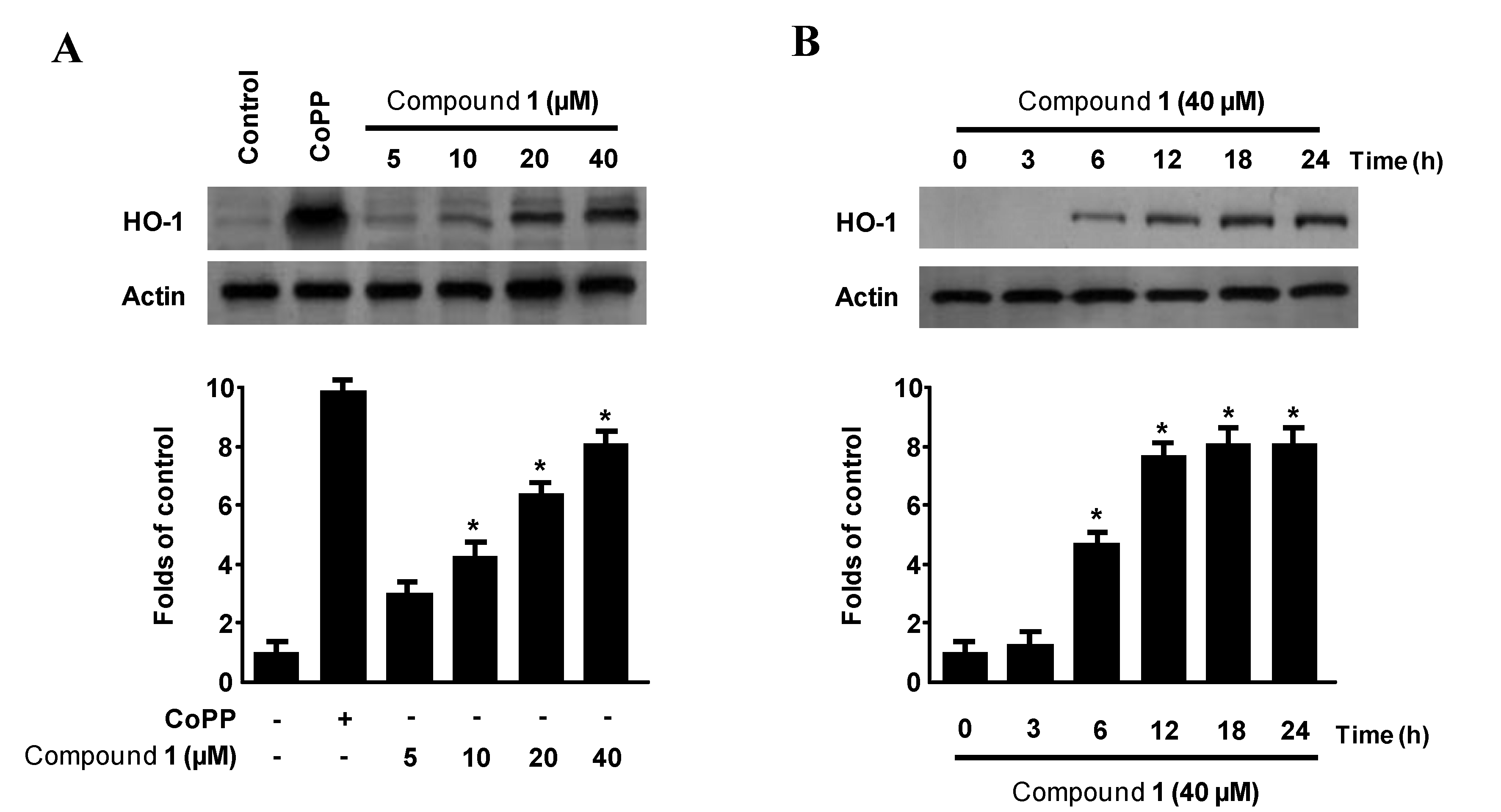
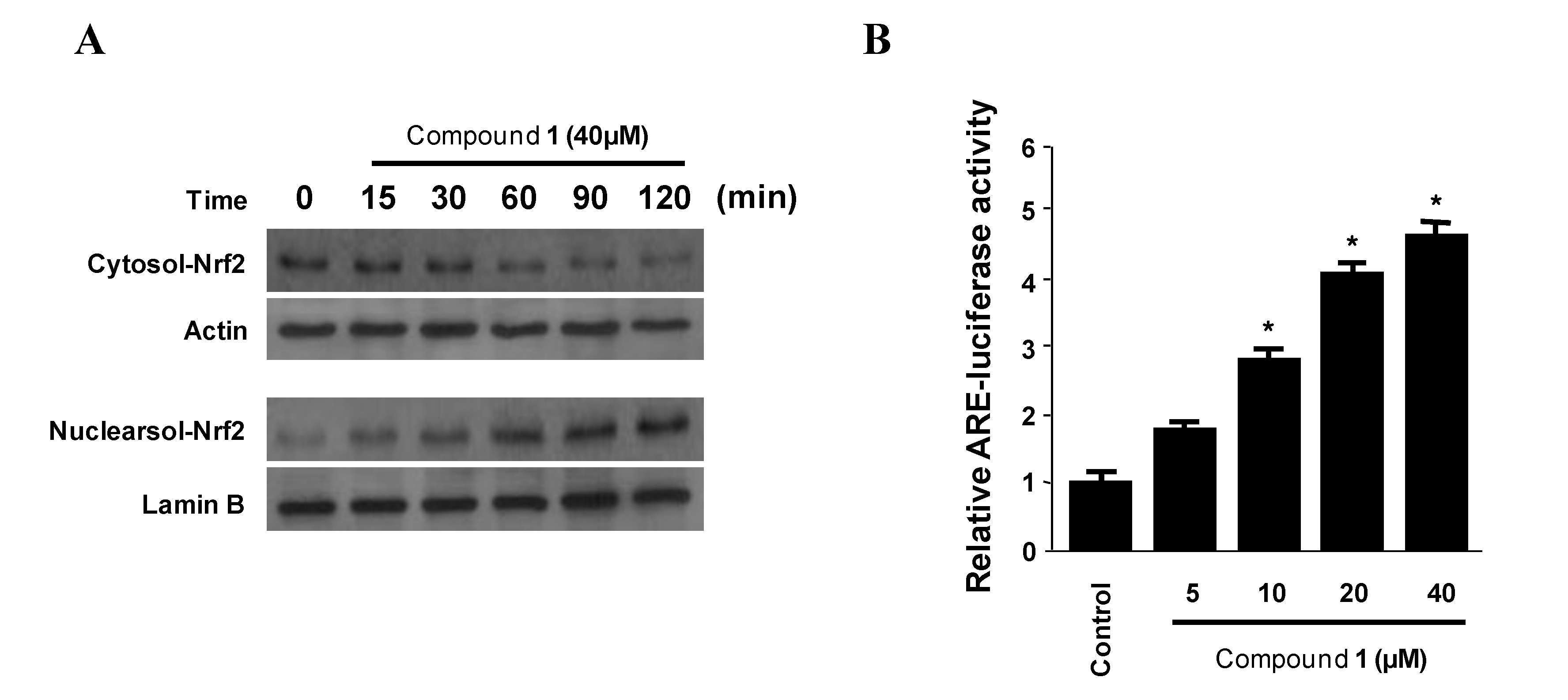
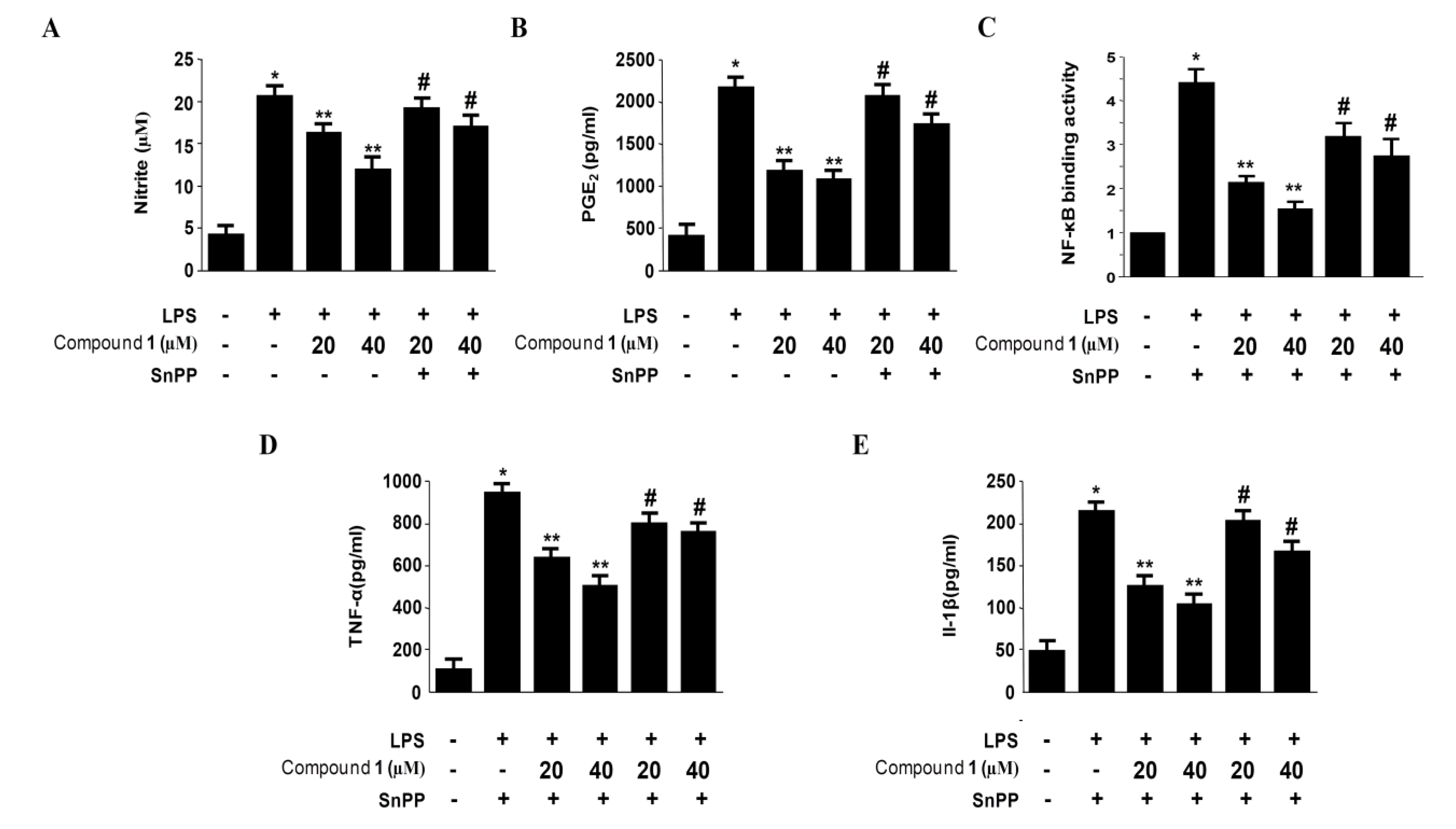
2.6. Effects of SnPP on the Inhibition of Pro-Inflammatory Mediator Production via the Pre-Treatment of Penstyrylpyrone (1) in LPS-Stimulated Murine Peritoneal Macrophages
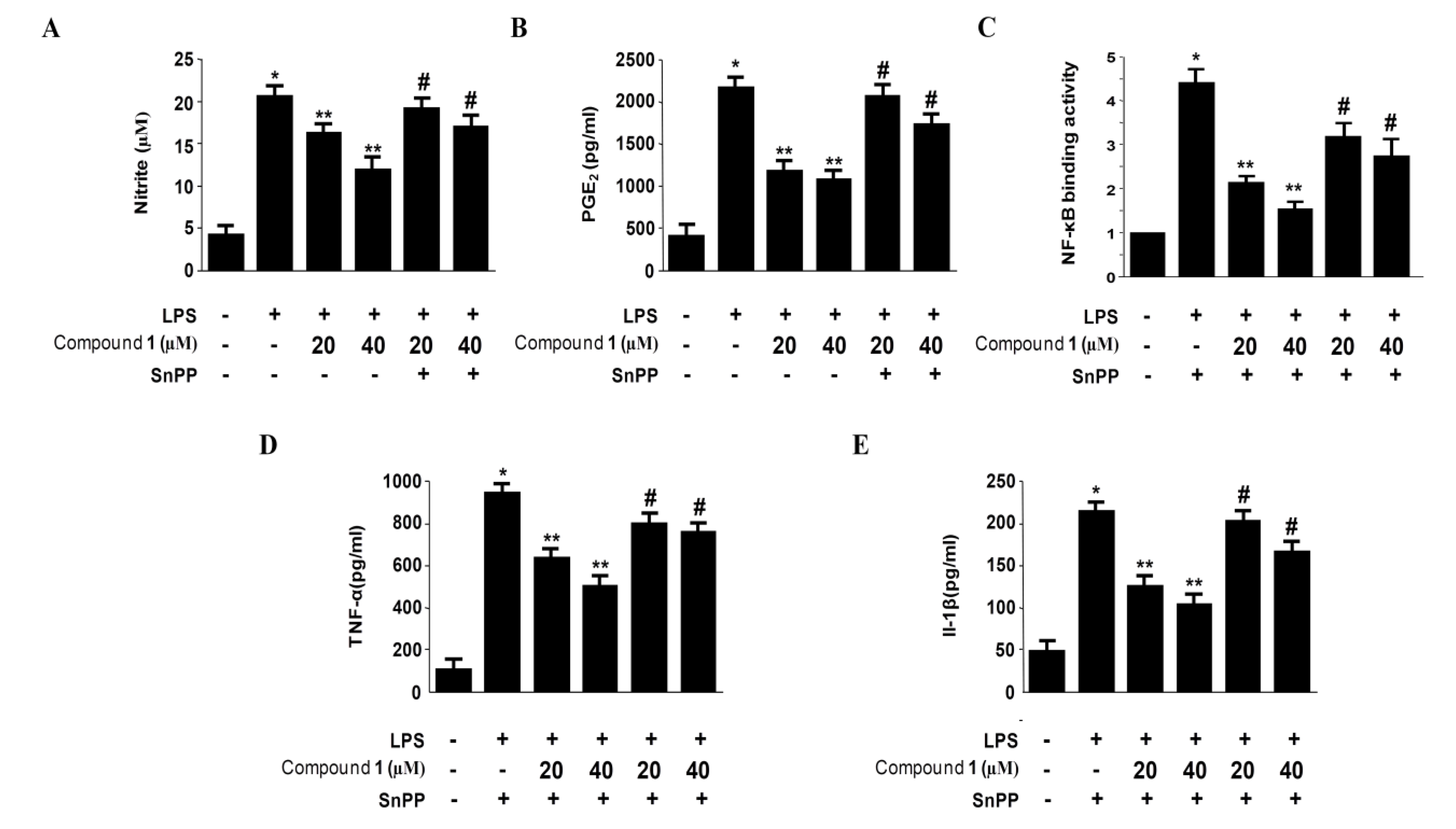
3. Experimental Section
3.1. General Experimental Procedures and Materials
3.2. Specimen Collection and Identification of the Marine-Derived Fungus Penicillium sp. JF-55
3.3. Isolation of Compounds 1–3 from the Marine-Derived Fungus Penicillium sp. JF-55
3.4. PTP1B Assay
3.5. Peritoneal Macrophage Cultures
3.6. Cell Viability Assay
3.7. Determination of Nitrite Production
3.8. PGE2, TNF-α and IL-1β Assay
3.9. Preparation of Cytosolic and Nuclear Fractions
3.10. Western Blotting Analysis
3.11. DNA-Binding Activity of NF-κB
3.12. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Olano, C.; Méndez, C.; Salas, J.A. Antitumor compounds from marine actinomycetes. Mar. Drugs 2009, 7, 210–248. [Google Scholar] [CrossRef]
- Rateb, M.E.; Ebel, R. Secondary metabolites of fungi from marine habitats. Nat. Prod. Rep. 2011, 28, 290–344. [Google Scholar] [CrossRef]
- Debbab, A.; Aly, A.H.; Lin, W.H.; Proksch, P. Bioactive compounds from marine bacteria and fungi. Microb. Biotechnol. 2010, 3, 544–563. [Google Scholar] [CrossRef]
- Greve, H.; Mohamed, I.E.; Pontius, A.; Kehraus, S.; Gross, H.; König, G.M. Fungal metabolites: Structural diversity as incentive for anticancer drug development. Phytochem. Rev. 2010, 9, 537–545. [Google Scholar] [CrossRef]
- Liu, D.; Li, X.M.; Meng, L.; Li, C.S.; Gao, S.S.; Shang, Z.; Proksch, P.; Huang, C.G.; Wang, B.G. Nigerapyrones A–H, α-pyrone derivatives from the marine mangrove-derived endophytic fungus Aspergillus niger MA-132. J. Nat. Prod. 2011, 74, 1787–1791. [Google Scholar]
- Yan, H.J.; Li, X.M.; Li, C.S.; Wang, B.G. Alkaloid and anthraquinone derivatives produced by the marine-derived endophytic fungus Eurotium rubrum. Helv. Chim. Acta 2012, 95, 163–168. [Google Scholar] [CrossRef]
- Seo, C.; Sohn, J.H.; Oh, H.; Kim, B.Y.; Ahn, J.S. Isolation of the protein tyrosine ohosphatase 1B inhibitory metabolite from the marine-derived fungus Cosmospora sp. SF-5060. Bioorg. Med. Chem. Lett. 2009, 19, 6095–6097. [Google Scholar] [CrossRef]
- Lee, D.S.; Jeong, G.S.; Li, B.; Lee, S.U.; Oh, H.; Kim, Y.C. Asperlin from the marine-derived fungus Aspergillus sp. SF-5044 exerts anti-inflammatory effects through heme oxygenase-1 expression in murine macrophages. J. Pharmacol. Sci. 2011, 116, 283–295. [Google Scholar] [CrossRef]
- Fisher, E.H.; Charbonneau, H.; Tonks, N.K. Protein tyrosine phosphatases: A diverse family of intracellular and transmembrane enzymes. Science 1991, 253, 401–406. [Google Scholar]
- Hunter, T. Protein kinases and phosphatases: The yin and yang of protein phosphorylation and signaling. Cell 1995, 80, 225–236. [Google Scholar] [CrossRef]
- Saltiel, A.R.; Kahn, C.R. Insulin signalling and the regulation of glucose and lipid metabolism. Nature 2001, 414, 799–806. [Google Scholar] [CrossRef]
- Elchebly, M.; Payette, P.; Michaliszyn, E.; Cromlish, W.; Collins, S.; Loy, A.L.; Normandin, D.; Cheng, A.; Himms-Hagen, J.; Chan, C.C.; et al. Increased insulin sensitivity and obesity resistance in mice lacking the protein tyrosine phosphatase-1B gene. Science 1999, 283, 1544–1548. [Google Scholar] [CrossRef]
- Klaman, L.D.; Boss, O.; Peroni, O.D.; Kim, J.K.; Martino, J.L.; Zabolotny, J.M.; Moghal, N.; Lubkin, M.; Kim, Y.B.; Sharpe, A.H.; et al. Increased energy expenditure, decreased adiposity, and tissue-specific insulin sensitivity in protein-tyrosine phosphatase 1B-deficient mice. Mol. Cell. Biol. 2000, 20, 5479–5489. [Google Scholar]
- Koren, S.; Fantus, I.G. Inhibition of the protein tyrosine phosphatase PTP1B: Potential therapy for obesity, insulin resistance and type-2 diabetes mellitus. Best Pract. Res. Clin. Endocrinol. Metab. 2007, 21, 621–640. [Google Scholar] [CrossRef]
- Dadke, S.; Chernoff, J. Protein-tyrosine phosphatase 1B as a potential drug target for obesity. Curr. Drug Targets Immune Endocr. Metabol. Disord. 2003, 3, 299–304. [Google Scholar] [CrossRef]
- Paine, A.; Eiz-Vesper, B.; Blasczyk, R.; Immenschuh, S. Signaling to heme oxygenase-1 and its anti-inflammatory therapeutic potential. Biochem. Pharmacol. 2010, 80, 1895–1903. [Google Scholar]
- Ryter, S.W.; Alam, J.; Choi, A.M. Heme oxygenase-1/carbon monoxide: From basic science to therapeutic applications. Physiol. Rev. 2006, 86, 583–650. [Google Scholar] [CrossRef]
- Otterbein, L.E.; Bach, F.H.; Alam, J.; Soares, M.; Lu, H.T.; Wysk, M.; Davis, R.J.; Flavell, R.A.; Choi, A.M. Carbon monoxide has anti-inflammatory effects involving the mitogen-activated protein kinase pathway. Nat. Med. 2000, 6, 422–428. [Google Scholar]
- Wiesel, P.; Foster, L.C.; Pellacani, A.; Layne, M.D.; Hsieh, C.M.; Huggins, G.S.; Strauss, P.; Yet, S.F.; Perrella, M.A. Thioredoxin facilitates the induction of heme oxygenase-1 in response to inflammatory mediators. J. Biol. Chem. 2000, 275, 24840–24846. [Google Scholar] [CrossRef]
- Suh, G.Y.; Jin, Y.; Yi, A.K.; Wang, X.M.; Choi, A.M. CCAAT/enhancer-binding protein mediates carbon monoxide-induced suppression of cyclooxygenase-2. Am. J. Respir. Cell Mol. Biol. 2006, 35, 220–226. [Google Scholar] [CrossRef]
- Oh, G.S.; Pae, H.O.; Lee, B.S.; Kim, B.N.; Kim, J.M.; Kim, H.R.; Jeon, S.B.; Jeon, W.K.; Chae, H.J.; Chung, H.T. Hydrogen sulfide inhibits nitric oxide production and nuclear factor-kappaB via heme oxygenase-1 expression in RAW264.7 macrophages stimulated with lipopolysaccharide. Free Radic. Biol. Med. 2006, 41, 106–119. [Google Scholar] [CrossRef]
- Itoh, K.; Chiba, T.; Takahashi, S.; Ishii, T.; Igarashi, K.; Katoh, Y.; Oyake, T.; Hayashi, N.; Satoh, K.; Hatayama, I.; et al. An Nrf2/small Maf heterodimer mediates the induction of phase II detoxifying enzyme genes through antioxidant response elements. Biochem. Biophys. Res. Commun. 1997, 236, 313–322. [Google Scholar]
- Kobayashi, M.; Yamamoto, M. Molecular mechanisms activating the Nrf2-Keap1 pathway of antioxidant gene regulation. Antioxid. Redox Signal. 2005, 7, 385–394. [Google Scholar] [CrossRef]
- Schmidt, L.E.; Gloer, J.B.; Wicklow, D.T. Solanapyrone analogues from a Hawaiian fungicolous fungus. J. Nat. Prod. 2007, 70, 1317–1320. [Google Scholar] [CrossRef]
- Kemami, H.V.K.; Hertweck, C. Squarrosidine and pinillidine: 3,3′-fused bis (styrylpyrones) from Pholiota squarrosa and Phellinus pini. Eur. J. Org. Chem. 2007, 20, 3292–3295. [Google Scholar]
- Mo, S.Y.; Yang, Y.C.; He, W.Y.; Shi, J.G. Two pyrone derivatives from fungus Phellinus igniarius. Chin. Chem. Lett. 2003, 14, 704–706. [Google Scholar]
- Ali, N.A.A.; Jansen, R.; Pilgrim, H.; Liberra, K.; Lindequist, U. Hispolon, a yellow pigment from Inonotus hispidus. Phytochemistry 1996, 41, 927–929. [Google Scholar]
- Fujita, K.I.; Nagamine, Y.; Ping, X.; Taniguchi, M. Mode of action of anhydrofulvic acid against Candida utilis ATCC 42402 under acidic condition. J. Antibiot. 1999, 52, 628–634. [Google Scholar]
- Capon, R.J.; Stewart, M.; Ratnayake, R.; Lacey, E.; Gill, J.H. Citromycetins and bilains A-C: New aromatic polyketides and diketopiperazines from Australian marine-derived and terrestrial Penicillium spp. J. Nat. Prod. 2007, 70, 1746–1752. [Google Scholar] [CrossRef]
- Na, M.; Tang, S.; He, L.; Oh, H.; Kim, B.S.; Oh, W.K.; Kim, B.Y.; Ahn, J.S. Inhibition of protein tyrosine phosphatase 1B by ursane-type triterpenes isolated from Symplocos paniculata. Planta Med. 2006, 72, 261–263. [Google Scholar]
- Zhang, W.; Hong, D.; Zhou, Y.; Zhang, Y.; Shen, Q.; Li, J.; Hu, L.; Li, J. Ursolic acid and its derivative inhibit protein tyrosine phosphatase 1B, enhancing insulin receptor phosphorylation and stimulating glucose uptake. Biochem. Biophy. Acta 2006, 1760, 1505–1512. [Google Scholar]
- Surh, Y.J.; Chun, K.S.; Cha, H.H.; Han, S.S.; Keum, Y.S.; Park, K.K.; Lee, S.S. Molecular mechanisms underlying chemopreventive activities of anti-inflammatory phytochemicals: Down-regulation of COX-2 and iNOS through suppression of NF-kappaB activation. Mutat. Res. 2001, 480–481, 243–268. [Google Scholar] [CrossRef]
- Verma, I.M.; Stevenson, J.K.; Schwarz, E.M.; van Antwerp, D.; Miyamoto, S. Rel/NF-kappaB/IkappaB family: Intimate tales of association and dissociation. Genes Dev. 1995, 9, 2723–2735. [Google Scholar] [CrossRef]
- Sato, K.; Balla, J.; Otterbein, L.; Smith, R.N.; Brouard, S.; Lin, Y.; Csizmadia, E.; Sevigny, J.; Robson, S.C.; Vercellotti, G.; et al. Garbon monoxide generated by heme oxygenase-1 suppresses the rejection of mouse-to-rat cardiac transplants. J. Immunol. 2001, 166, 4185–4194. [Google Scholar]
- Hamaguchi, T.; Sudo, T.; Osada, H. RK-682, a potent inhibitor of tyrosine phosphatase, arrested the mammalian cell cycle progression at G1phase. FEBS Lett. 1995, 372, 54–58. [Google Scholar] [CrossRef]
- Oh, H.; Kim, B.S.; Bae, E.Y.; Kim, M.S.; Kim, B.Y.; Lee, H.B.; Kim, C.J.; Ahn, J.S. Inhibition of PTP1B by metabolites from Micromucor ramannianus var. angulisporus CRM000232. J. Antibiot. 2004, 57, 528–531. [Google Scholar] [CrossRef]
- Narumi, S.; Finke, J.H.; Hamilton, T.A. Interferon gamma and interleukin 2 synergize to induce selective monokine expression in murine peritoneal macrophages. J. Biol. Chem. 1990, 265, 7036–7041. [Google Scholar]
- Berridge, M.V.; Tan, A.S. Characterization of the cellular reduction of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT): Subcellular localization, substrate dependence, and involvement of mitochondrial electron transport in MTT reduction. Arch. Biochem. Biophys. 1993, 303, 474–482. [Google Scholar] [CrossRef]
- Titheradge, M.A. The enzymatic measurement of nitrate and nitrite. Meth. Mol. Biol. 1998, 303, 474–482. [Google Scholar]
- Sawle, P.; Foresti, R.; Mann, B.E.; Johnson, T.R.; Green, C.J.; Motterlini, R. Carbon monoxide-releasing molecules (CO-RMs) attenuate the inflammatory response elicited by lipopolysaccharide in RAW264.7 murine macrophages. Br. J. Pharmacol. 2005, 145, 800–810. [Google Scholar] [CrossRef]
- Liljebris, C.; Martinsson, J.; Tedenborg, L.; Williams, M.; Barker, E.; Duffy, J.E.S.; Nygren, A.; James, S. Pigments of fungi (macromycetes). Nat. Prod. Rep. 2010, 27, 1531–1570. [Google Scholar] [CrossRef]
- Lee, I.K.; Yun, B.S. Styrylpyrone-class compounds from medicinal fungi Phellinus and Inonotus spp., and their medicinal importance. J. Antiobiot. 2011, 64, 349–359. [Google Scholar] [CrossRef]
- Wrigley, S.K.; Latif, M.A.; Gibson, T.M.; Chicarelli-Robinson, M.I.; Williams, D.H. Structure elucidation of xanthone derivatives with CD4-binding activity from Penicillium glabrum (Wehmer) Westling. Pure Appl. Chem. 1994, 66, 2383–2386. [Google Scholar] [CrossRef]
- Zabolotny, J.M.; Kim, Y.B.; Welsh, L.A.; Kershaw, E.E.; Neel, B.G.; Kahn, B.B. Proteintyrosine phosphatase 1B expression is induced by inflammation in vivo. J. Biol. Chem. 2008, 283, 14230–14241. [Google Scholar]
- González-Rodríguez, A.; Más-Gutierrez, J.A.; Mirasierra, M.; Fernandez-Pérez, A.; Lee, Y.J.; Ko, H.J.; Kim, J.K.; Romanos, E.; Carrascosa, J.M.; Ros, M.; et al. Essential role of protein tyrosine phosphatase 1B in obesity-induced inflammation and peripheral insulin resistance during aging. Aging Cell 2012, 11, 284–296. [Google Scholar] [CrossRef] [Green Version]
- García-San Frutos, M.; Fernández-Agulló, T.; Carrascosa, J.M.; Horrillo, D.; Barrús, M.T.; Oliveros, E.; Sierra, J.; Ros, M. Involvement of protein tyrosine phosphatases and inflammation in hypothalamic insulin resistance associated with ageing: Effect of caloric restriction. Mech. Aging Dev. 2012, 133, 489–497. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lee, D.-S.; Jang, J.-H.; Ko, W.; Kim, K.-S.; Sohn, J.H.; Kang, M.-S.; Ahn, J.S.; Kim, Y.-C.; Oh, H. PTP1B Inhibitory and Anti-Inflammatory Effects of Secondary Metabolites Isolated from the Marine-Derived Fungus Penicillium sp. JF-55. Mar. Drugs 2013, 11, 1409-1426. https://doi.org/10.3390/md11041409
Lee D-S, Jang J-H, Ko W, Kim K-S, Sohn JH, Kang M-S, Ahn JS, Kim Y-C, Oh H. PTP1B Inhibitory and Anti-Inflammatory Effects of Secondary Metabolites Isolated from the Marine-Derived Fungus Penicillium sp. JF-55. Marine Drugs. 2013; 11(4):1409-1426. https://doi.org/10.3390/md11041409
Chicago/Turabian StyleLee, Dong-Sung, Jae-Hyuk Jang, Wonmin Ko, Kyoung-Su Kim, Jae Hak Sohn, Myeong-Suk Kang, Jong Seog Ahn, Youn-Chul Kim, and Hyuncheol Oh. 2013. "PTP1B Inhibitory and Anti-Inflammatory Effects of Secondary Metabolites Isolated from the Marine-Derived Fungus Penicillium sp. JF-55" Marine Drugs 11, no. 4: 1409-1426. https://doi.org/10.3390/md11041409