Diarrhetic Shellfish Toxins and Other Lipophilic Toxins of Human Health Concern in Washington State

Abstract
:1. Introduction

2. Results





{kind=link}
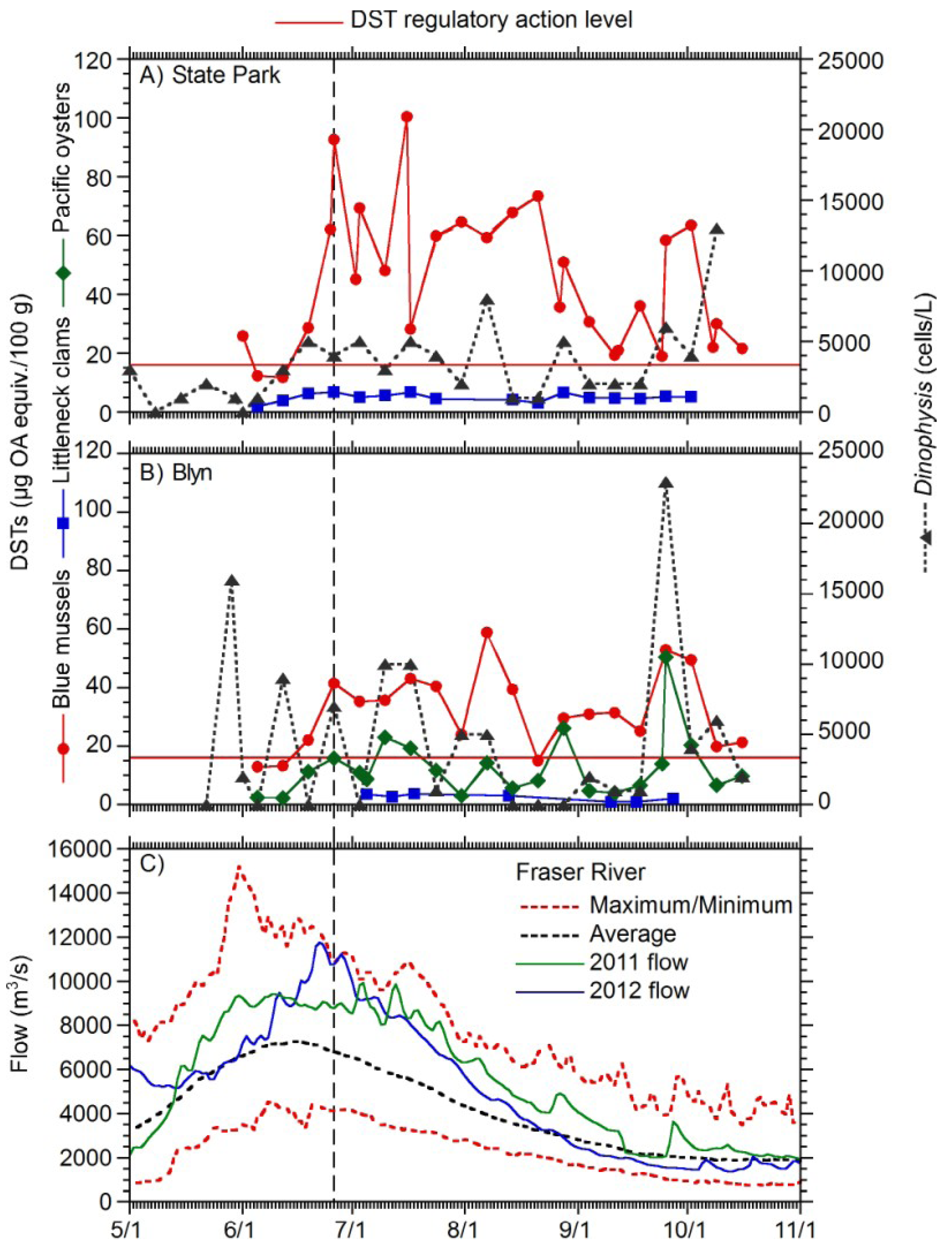{kind=link}
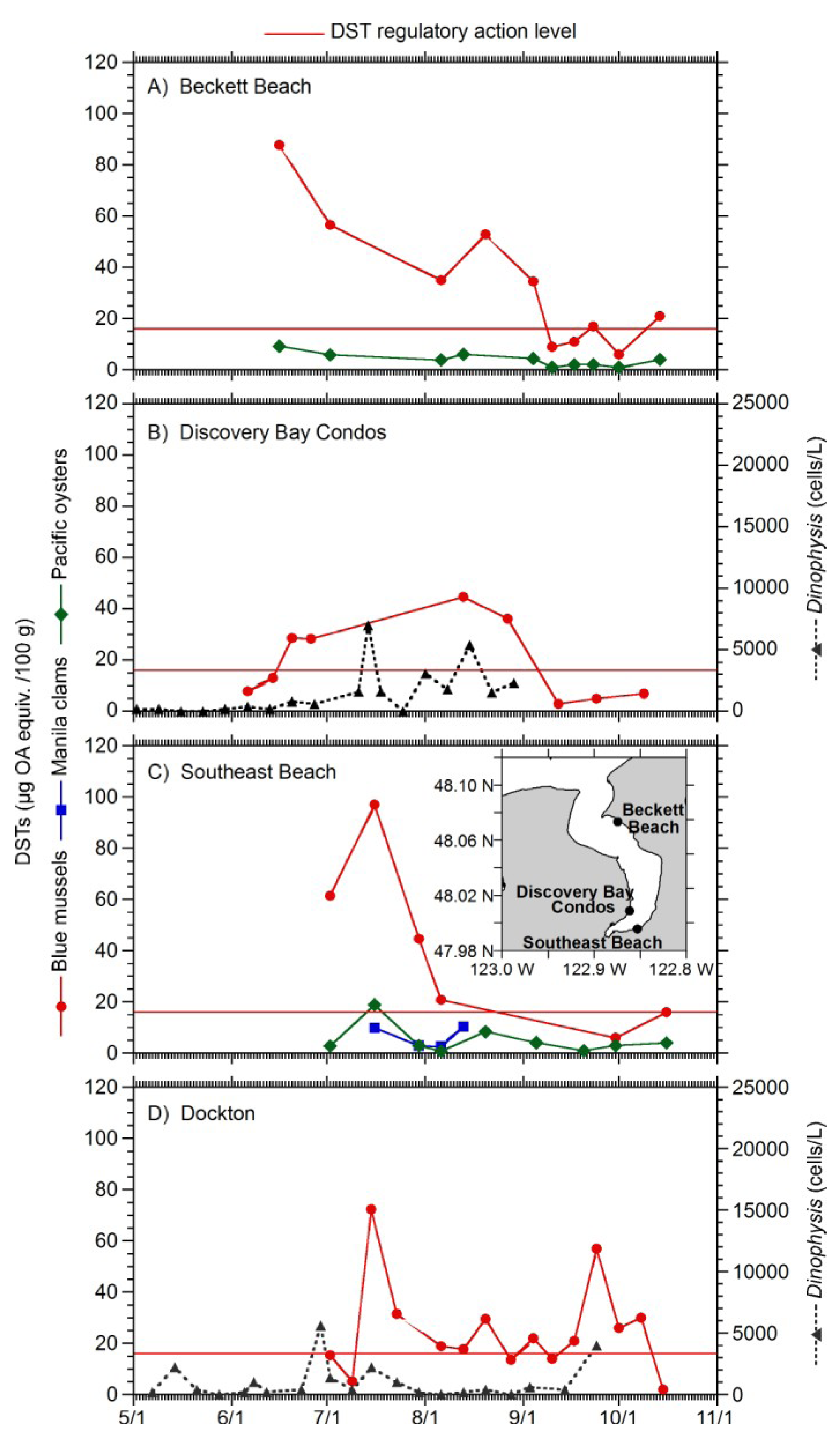{kind=link}
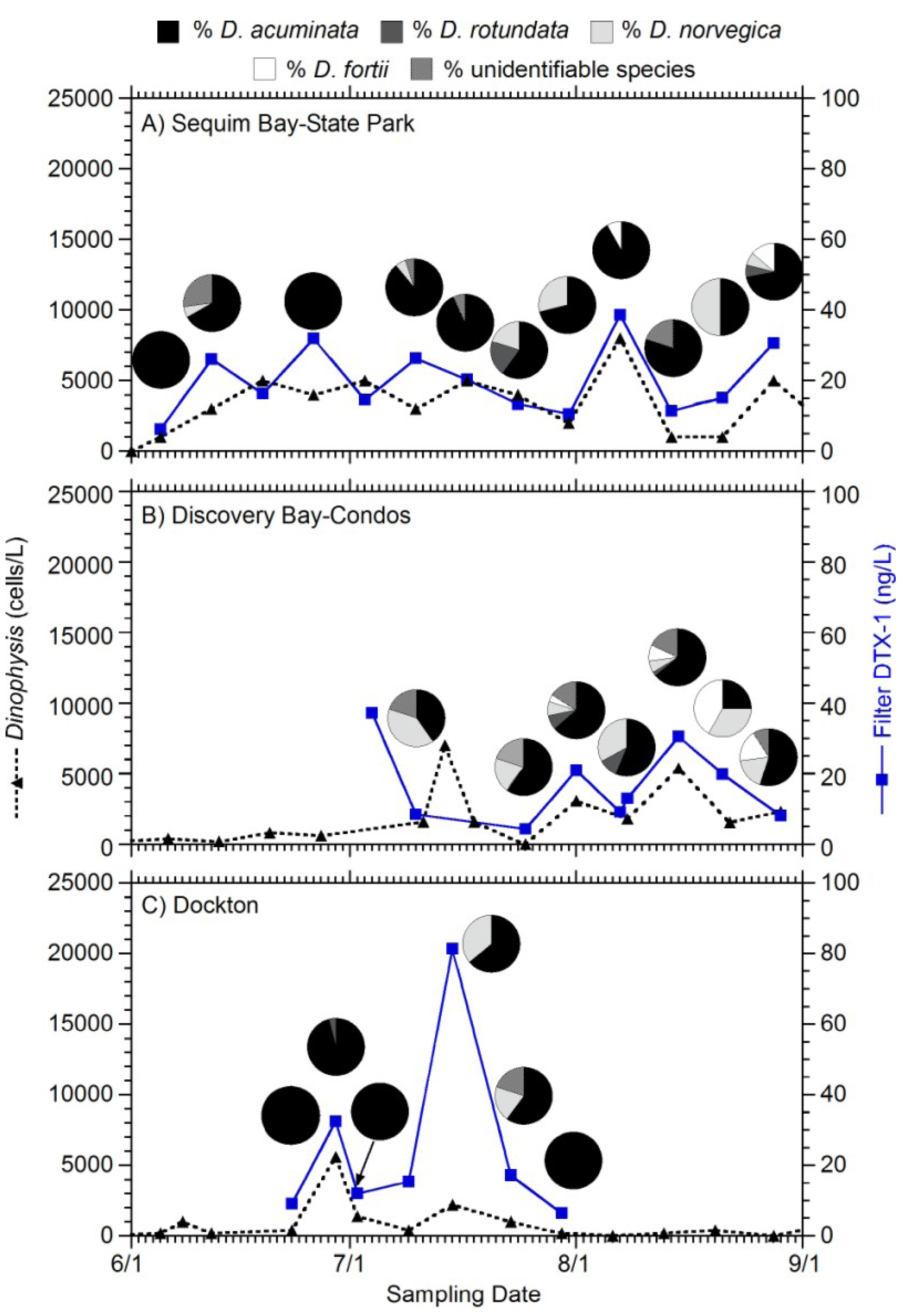{kind=link}
{kind=link}
| Date | Sample | Total OA equiv | DTX-1 | DTX-3 | YTX | PTX2 | AZA-2 | Dinophysis (cells/L) |
|---|---|---|---|---|---|---|---|---|
| 6/5/12 | Blue Mussel | 12.33 | 5.76 | 6.57 | na 2 | bd 3 | bd | 1000 |
| Pacific Oyster | 2.55 | 1.11 | 1.44 | na | bd | bd | ||
| Littleneck Clam | 1.84 | 0.00 | 1.84 | na | bd | bd | ||
| Plankton Filter | 6.24 | 3.88 | 2.36 | 2.95 | 5.60 | bd | ||
| 6/12/12 | Blue Mussel | 11.80 | 7.47 | 4.33 | na | bd | bd | 3000 |
| Pacific Oyster | 2.35 | 0.00 | 2.35 | na | bd | bd | ||
| Littleneck Clam | 3.99 | 0.00 | 3.99 | na | bd | bd | ||
| Plankton Filter | 26.21 | 19.62 | 6.59 | bd | 33.48 | bd | ||
| 6/19/12 | Blue Mussel | 28.72 | 28.72 | 0.00 | na | 65.09 | bd | 5000 |
| Pacific Oyster | 11.31 | 4.10 | 7.21 | na | 59.59 | bd | ||
| Littleneck Clam | 6.26 | 0.00 | 6.26 | na | 17.38 | bd | ||
| Plankton Filter | 16.37 | 12.42 | 3.95 | bd | 8.24 | bd | ||
| 6/26/12 | Blue Mussel | 92.69 | 54.52 | 38.17 | na | 57.16 | bd | 4000 |
| Pacific Oyster | 15.91 | 5.59 | 10.32 | na | 34.41 | bd | ||
| Littleneck Clam | 6.80 | 5.06 | 1.74 | na | 12.58 | bd | ||
| Plankton Filter | 32.05 | 23.14 | 8.91 | bd | 13.05 | bd | ||
| 7/3/12 | Blue Mussel | 69.35 | 48.09 | 21.26 | 27.32 | 3.30 | bd | 5000 |
| Pacific Oyster | 10.95 | 2.62 | 8.33 | 3.70 | bd | bd | ||
| Littleneck Clam | 5.08 | 0.00 | 5.08 | bd | bd | bd | ||
| Plankton Filter | 14.63 | 9.91 | 4.72 | bd | 12.44 | bd | ||
| 7/10/12 | Blue Mussel | 48.12 | 34.22 | 13.90 | 89.70 | 1.46 | bd | 3000 |
| Pacific Oyster | 23.10 | 4.24 | 18.86 | bd | 3.96 | bd | ||
| Littleneck Clam | 5.66 | 0.00 | 5.66 | bd | bd | bd | ||
| Plankton Filter | 26.41 | 20.58 | 5.83 | bd | 20.82 | bd | ||
| 7/17/12 | Blue Mussel | 28.29 | 13.64 | 14.65 | 22.78 | bd | bd | 5000 |
| Pacific Oyster | 19.38 | 2.94 | 16.44 | 1.78 | 2.06 | bd | ||
| Littleneck Clam | 6.77 | 1.02 | 5.75 | bd | 1.33 | bd | ||
| Plankton Filter | 20.36 | 15.48 | 4.88 | bd | 13.52 | bd | ||
| 7/24/12 | Blue Mussel | 59.90 | 44.38 | 15.52 | 58.48 | 1.57 | bd | 4000 |
| Pacific Oyster | 11.71 | 2.37 | 9.34 | bd | 1.33 | bd | ||
| Littleneck Clam | 4.51 | 0.00 | 4.51 | bd | 1.82 | bd | ||
| Plankton Filter | 13.35 | 8.78 | 4.57 | 3.00 | 8.55 | bd | ||
| 7/31/12 | Blue Mussel | 64.68 | 40.60 | 24.08 | 33.78 | bd | bd | 2000 |
| Pacific Oyster | 3.10 | 1.25 | 1.85 | 1.10 | bd | bd | ||
| Littleneck Clam | 7.79 | 1.28 | 6.51 | bd | 1.00 | bd | ||
| Plankton Filter | 10.58 | 7.88 | 2.70 | bd | 5.91 | 0.40 | ||
| 8/7/12 | Blue Mussel | 59.30 | 36.28 | 23.02 | 30.93 | 2.83 | bd | 8000 |
| Pacific Oyster | 14.27 | 3.65 | 10.62 | 1.51 | 1.70 | bd | ||
| Littleneck Clam | 3.99 | 0.00 | 3.99 | bd | bd | bd | ||
| Plankton Filter | 38.67 | 29.74 | 8.93 | bd | 33.72 | 0.46 | ||
| 8/14/12 | Blue Mussel | 66.61 | 42.76 | 23.85 | 30.20 | bd | bd | 1000 |
| Pacific Oyster | 5.67 | 1.96 | 3.71 | 2.88 | bd | bd | ||
| Littleneck Clam | 4.21 | 1.10 | 3.11 | 1.43 | bd | bd | ||
| Plankton Filter | 11.42 | 10.13 | 1.29 | bd | 8.55 | bd | ||
| 8/21/12 | Blue Mussel | 73.48 | 65.98 | 7.50 | 37.26 | bd | bd | 1000 |
| Pacific Oyster | 8.17 | 2.77 | 5.40 | 3.55 | 2.77 | bd | ||
| Littleneck Clam | 3.19 | 0.00 | 3.19 | 1.62 | bd | bd | ||
| Plankton Filter | 15.15 | 10.31 | 4.84 | 3.10 | 18.65 | 0.40 | ||
| 8/28/12 | Blue Mussel | 51.00 | 42.52 | 8.48 | 34.72 | 2.51 | bd | 5000 |
| Pacific Oyster | 26.20 | 5.59 | 20.61 | 4.88 | 3.49 | bd | ||
| Littleneck Clam | 6.69 | 1.69 | 5.00 | bd | 1.85 | bd | ||
| Plankton Filter | 30.66 | 22.62 | 8.04 | 6.79 | 38.36 | bd |
3. Discussion
3.1. Dinophysis Monitoring
3.2. Environmental Triggers
3.3. DSTs in the USA
3.4. Other Lipophilic Toxins
3.5. Lipophilic Toxin Regulation
4. Experimental Section
4.1. Sample Collection
4.2. Shellfish Preparation and Homogenization
4.3. Filter Preparation and Extraction
4.4. Tissue and Filter Extract Hydrolysis
4.5. LC-MS/MS Analysis
4.6. Dinophysis Cell Counts and Species Identification
4.7. Fraser River Data
5. Conclusions
Acknowledgements
Conflict of Interest
References
- Van Egmond, H.P.; van Apeldoorn, M.E.; Speijers, G.J.A. Marine Biotoxins. In Food and Nutrition Paper 80; FAO: Rome, Italy, 2004; pp. 53–96. [Google Scholar]
- Control of Communicable Diseases Manual, 19th ed.; Heymann, D.L. (Ed.) American Public Health Association Press: Washington, DC, USA; p. 2008.
- Cembella, A.D. Occurrence of okadaic acid, a major diarrheic shellfish toxin, in natural populations of Dinophysis spp. from the eastern coast of North America. J. Appl. Phycol. 1989, 1, 307–310. [Google Scholar] [CrossRef]
- Barceloux, D.G. Diarrhetic Shellfish Poisoning and Okadaic Acid. In Medical Toxicology of Natural Substances: Foods, Fungi, Medicinal Herbs, Plants, and Venomous Animals; John Wiley & Sons: Hoboken, NJ, USA, 2008; pp. 222–226. [Google Scholar]
- Burgess, V.; Shaw, G. Pectenotoxins—An issue for public health. A review of their comparative toxicology and metabolism. Environ. Int. 2001, 27, 275–283. [Google Scholar] [CrossRef]
- Stamman, E.; Segar, D.A.; Davis, P.G. A Preliminary Epidemiological Assessment of the Potential for Diarrhetic Shellfish Poisoning in the Northeast United States; NOAA Technical Memorandum NOS OMA 34; NOAA: Rockville, MD, USA, April 1987. Available online: http://docs.lib.noaa.gov/noaa_documents/NOS/OMA/TM_NOS_OMA/nos_oma_34.pdf (accessed on 9 May 2013).
- Manerio, E.; Rodas, V.L.; Costas, E.; Martinez, H.J. Shellfish consumption: A major risk factor for colorectal cancer. Med. Hypotheses 2008, 70, 409–412. [Google Scholar] [CrossRef]
- Cordier, S.; Monfort, C.; Miossec, L.; Richardson, S.; Belin, C. Ecological analysis of digestive cancer mortality related to contamination by diarrhetic shellfish poisoning toxins along the coasts of France. Environ. Res. 2000, 84, 145–150. [Google Scholar] [CrossRef]
- Yasumoto, T.; Murata, M.; Oshima, Y.; Sano, M.; Matsumoto, G.; Clardy, J. Diarrhetic shellfish toxins. Tetrahedron 1985, 41, 1019–1025. [Google Scholar] [CrossRef]
- Yasumoto, T. The chemistry and biological function of natural marine toxins. Chem. Record 2001, 1, 228–242. [Google Scholar] [CrossRef]
- Hallegraeff, G.M. Harmful Algal Blooms: A Global Overview. In Manual on Harmful Marine Microalgae; Hallegraeff, G.M., Anderson, D.M., Cembella, A.D., Eds.; Intergovernmental Oceanographic Commission Manuals and Guides No. 33; United Nations Educational, Scientific and Cultural Organization, IOC Science and Communication Centre on Harmful Algae: Copenhagen, Denmark, 2003; pp. 1–22. [Google Scholar]
- Campbell, L.; Olson, R.J.; Sosik, H.M.; Abraham, A.; Henricks, D.W.; Hyatt, C.J.; Buskey, E.J. First harmful Dinophysis (Dionphyceae, Dinophysiales) bloom in the U.S. is revealed by automated imaging flow cytometry. J. Phycol. 2010, 46, 66–75. [Google Scholar] [CrossRef]
- Deeds, J.R.; Wiles, K.; Heideman, G.B., VI; White, K.D.; Abraham, A. First U.S. report of shellfish harvesting closures due to confirmed okadaic acid in Texas Gulf coast oysters. Toxicon 2010, 55, 1138–1146. [Google Scholar] [CrossRef]
- Taylor, M.; McIntyre, L.; Ritson, M.; Stone, J.; Bronson, R.; Bitzikos, O.; Rourke, W.; Galanis, E. Outbreak of diarrhetic shellfish poisoning associated with mussels. Mar. Drugs 2013. submitted for publication. [Google Scholar]
- Horner, R.; Garrison, D.; Plumley, F. Harmful algal blooms and red tide problems on the U.S. west coast. Limnol. Oceanogr. 1997, 42, 1076–1088. [Google Scholar] [CrossRef]
- Takai, A.; Bialojan, C.; Troschka, M.; Ruegg, J.C. Smooth muscle myosin phosphatase inhibition and force enhancement by black sponge toxin. FEBS Lett. 1987, 217, 81–84. [Google Scholar] [CrossRef]
- Fujiki, H.; Suganuma, M. Tumor promotion by inhibitors of protein phosphatases 1 and 2A: The okadaic acid class of compounds. Adv. Cancer Res. 1993, 61, 143–194. [Google Scholar] [CrossRef]
- Tereo, K.; Ito, E.; Yasumoto, T.; Yamuguchi, K. Enterotoxic, Hepatotoxic and Immunotoxic Effects of Dinoflagellate Toxins on Mice. In Toxic Marine Phytoplankton; Granéli, E., Sundström, B., Edler, L., Anderson, D.M., Eds.; Elsevier Science Publishing Co., Inc.: New York, NY, USA, 1990; pp. 418–423. [Google Scholar]
- Draisci, R.; Lucentini, L.; Mascioni, A. Pectenotoxins and Yessotoxins: Chemistry, Toxicology, Pharmacology, and Analysis. In Seafood and Freshwater Toxins: Pharmacology, Physiology, and Detection; Botana, L.M., Ed.; Marcel Dekker, Inc.: New York, NY, USA, 2000; pp. 289–324. [Google Scholar]
- Aune, T. Risk Assessment of Toxins Associated with DSP, PSP and ASP in Seafood. In Mycotoxins and Phycotoxins in Perspective at the Turn of the Millennium, Proceedings of the X International Union of Pure and Applied Chemistry Symposium on Mycotoxins and Phycotoxins, Guarujá, Brazil, 21–25 May 2000; de Koe, W.J., Samson, R.A., van Egmond, H.P., Gilbert, J., Sabino, M., Eds.; IUPIC: Wageningen, The Netherlands, 2001; pp. 515–526. [Google Scholar]
- Aune, T.; Sorby, R.; Yasumoto, T.; Ramstad, H.; Landsverk, T. Comparison of oral and intraperitoneal toxicity of yessotoxin towards mice. Toxicon 2002, 40, 77–82. [Google Scholar] [CrossRef]
- Miles, C.O.; Wilkins, A.L.; Munday, R.; Dines, M.H.; Hawkes, A.D.; Briggs, L.R.; Sandvik, M.; Jensen, D.J.; Cooney, J.M.; Holland, P.T.; et al. Isolation of pectenotoxin-2 from Dinophysis acuta and its conversion to pectenotoxin-2 seco acid, and preliminary assessment of their acute toxicities. Toxicon 2004, 43, 1–9. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations, Intergovernmental Oceanographic Commission of UNESCO, World Health Organization; Report of the Joint FAO/IOC/WHO ad Hoc Expert Consultation on Biotoxins in Bivalve Molluscs; Food and Agriculture Organization: Oslo, Norway, 24–30 September 2004; p. 40.
- Reguera, B.; Pizarro, G. Planktonic Dinoflagellates That Contain Polyether Toxins of the Old “DSP Complex”. In Seafood and Freshwater Toxins: Pharmacology, Physiology, and Detection; Botana, L.M., Ed.; Marcel Dekker Inc.: New York, NY, USA, 2000; pp. 257–287. [Google Scholar]
- Krock, B.; Tillmann, U.; John, U.; Cembella, A.D. Characterization of azaspiracids in plankton size-fractions and isolation of an azaspiracid-producing dinoflagellate from the North Sea. Harmful Algae 2009, 8, 254–263. [Google Scholar] [CrossRef]
- Tillmann, U.; Elbrachter, M.; Krock, B.; John, U.; Cembella, A. Azadinium spinosum gen. et sp. nov. (Dinophyceae) identified as a primary producer of azaspiracid toxins. Eur. J. Phycol. 2009, 44, 63–79. [Google Scholar] [CrossRef]
- Tango, P.; Butler, W.; Lacouture, R.; Goshorn, D.; Magnien, R.; Michael, B. An Unprecedented Bloom of Dinophysis acuminata in Chesapeake Bay. In Proceedings of the Xth International Conference on Harmful Algae; Steidinger, K.A., Landsberg, J.H., Tomas, C.R., Vargo, G.A., Eds.; Florida Fish and Wildlife Conservation Commission, Florida Institute of Oceanography, IOC-UNESCO: St. Petersburg, FL, USA, 2004; pp. 358–360. [Google Scholar]
- Viviani, R. Eutrophication, marine biotoxins, human health. Sci. Total Environ. Suppl. 1992, 631–662. [Google Scholar]
- Morton, S.; Moeller, P.D.R.; Young, K.; Lanoue, B. Okadaic acid production from the marine dinoflagellate Prorocentrum belizeanum Faust isolated from the Belize coral reef ecosystem. Toxicon 1998, 36, 201–206. [Google Scholar] [CrossRef]
- Hackett, J.D.; Tong, M.; Kulis, D.M.; Fux, E.; Hess, P.; Bire, R.; Anderson, D.M. DSP toxin production de novo incultures of Dinophysis acuminata (Dinophyceae) from North America. Harmful Algae 2009, 8, 873–879. [Google Scholar] [CrossRef]
- Kamiyama, T.; Suzuki, T. Production of dinophysistoxin-1 and pectenotoxin-2 by a culture of Dinophysis acuminata (Dinophyceae). Harmful Algae 2009, 8, 312–317. [Google Scholar] [CrossRef]
- González-Gil, S.; Pizarro, G.; Paz, B.; Velo-Suárez, L.; Reguera, B. Is Dinophysis rotundata a Toxigenic Species? International Society for the Study of Harmful Algae, Environmental Science Publisher: Hong Kong, China, 2008. [Google Scholar]
- Botana, L.M.; Rodriguez-Vieytes, M.; Alfonso, A.; Louzao, M.C. Phycotoxins: Paralytic Shellfish Poisoning and Diarrhetic Shellfish Poisoning. In Handbook of Food Analysis—Residues and Other Food Component Analysis; Nollet, L.M.L., Ed.; Marcel Dekker Inc.: New York, NY, USA, 1996; Volume 2, pp. 1147–1169. [Google Scholar]
- Anderson, D.M.; Andersen, P.; Bricelj, M.V.; Cullen, J.J.; Rensel, J.E.J. Monitoring and Management Strategies for Harmful Algal Blooms in Coastal Waters; APEC #201-MR-01.1/IOC Technical Series No. 59; Asia Pacific Economic Program, Singapore, and Intergovernmental Oceanographic Commission: Paris, France, 2001. [Google Scholar]
- Reguera, B.; Velo-Suárez, L.; Raine, R.; Park, M.G. Harmful Dinophysis species: A review. Harmful Algae 2012, 14, 87–106. [Google Scholar] [CrossRef]
- Franks, P.J.S.; Anderson, D.M. Sampling Coastal Dinoflagellate Blooms: Equipment, Strategies, and Data Processing. In Biology, Epidemiology and Management of Pyrodinium Red Tides; Hallegraeff, G.M., Maclean, J.L., Eds.; International Center for Living Aquatic Resources Management: Manila, Philippines, 1989; pp. 235–254. [Google Scholar]
- Lloyd, J.; Duchin, J.; Borchert, J.; Flores-Quintana, H.; Robertson, A. Diarrhetic shellfish poisoning in Washington State, USA, 2011. Emerg. Infect. Dis. 2013, in press. [Google Scholar]
- Jester, R.; Lefebvre, K.; Langlois, G.; Vigilant, V.; Baugh, K.; Silver, M.W. A shift in the dominant toxin-producing algal species in central California alters phycotoxins in food webs. Harmful Algae 2009, 8, 291–298. [Google Scholar] [CrossRef]
- Rourke, W. Personal communication, Canadian Food Inspection Agency: Dartmouth, Canada, 2012.
- Konoki, K.; Onoda, T.; Watanabe, R.; Cho, Y.; Kaga, S.; Suzuki, T.; Yotsu-Yamashita, M. In vitro acylation of okadaic acid in the presence of various bivalves’ extracts. Mar. Drugs 2013, 11, 300–315. [Google Scholar] [CrossRef]
- Deeds, J. Personal communication, US Food and Drug Administration: College Park, MD, USA, 2011.
- Mountfort, D.O.; Suzuki, T.; Truman, P. Protein phosphatase inhibition assay adapted for determination of total DSP in contaminated mussels. Toxicon 2001, 39, 383–390. [Google Scholar] [CrossRef]
- Hess, P.; McMahon, T.; Slattery, D.; Swords, D.; Dowling, G.; McCarron, M.; Clarke, D.; Devilly, L.; Gibbons, W.; Silke, J.; et al. Biotoxin Chemical Monitoring in Ireland. In Proceedings of the 2nd Irish Marine Science Biotoxin Workshop, Marine Institute, Galway, Ireland, 11 October 2001; pp. 8–18.
- Furey, A.; Moroney, C.; Braña Magdalena, A.; Saez, M.J.F.; Lehane, M.; James, K.J. Geographical, temporal, and species variation of the polyether toxins, azaspiracids, in shellfish. Environ. Sci. Technol. 2003, 37, 3078–3084. [Google Scholar] [CrossRef]
- Hess, P.; McMahon, T.; Slattery, D.; Swords, D.; Dowling, G.; McCarron, M.; Clarke, D.; Gibbons, W.; Silke, J.; O’Cinneide, M. Use of LC-MS Testing to Identify Lipophilic Toxins, to Establish Local Trends and Interspecies Differences and to Test the Comparability of LC-MS Testing with the Mouse Bioassay: An Example from the Irish Biotoxin Monitoring Programme 2001. In Proceeding of the 4th International Conference Molluscan Shellfish Safety; Villalba, A., Reguera, B., Romalde, J.L., Beiras, R., Eds.; Consellería de Pesca e Asuntos Marítimos da Xunta de Galicia and Intergovernmental Oce anographic Commission of UNESCO: Santiago de Compostela, Spain, 2003; pp. 57–65. [Google Scholar]
- Satake, M.; Ofuji, K.; James, K.J.; Furey, A.; Yasumoto, T. New Toxic Event Caused by Irish Mussels. In Harmful Algae; Reguera, B., Blanco, J., Fernandez, M.L., Wyatt, T., Eds.; Xunta de Galicia and Intergovernmental Oceanographic Commission of UNESCO: Santiago de Compostela, Spain, 1998; pp. 468–469. [Google Scholar]
- Ito, E.; Satake, M.; Ofuji, K.; Higashi, M.; Harigaya, K.; McMahon, T.; Yasumoto, T. Chronic effects in mice caused by oral administration of sublethal doses of azaspiracid, a new marine toxin isolated from mussels. Toxicon 2002, 40, 193–203. [Google Scholar] [CrossRef]
- James, K.J.; Furey, A.; Lehane, M.; Ramstad, H.; Aune, T.; Hovgaard, P.; Morris, S.; Higman, W.; Satake, M.; Yasumoto, T. First evidence of an extensive northern European distribution of azaspiracid poisoning (AZP) toxins in shellfish. Toxicon 2002, 40, 909–915. [Google Scholar] [CrossRef]
- Braña Magdalena, A.; Lehane, M.; Krys, S.; Fernandez, M.L.; Furey, A.; James, K.J. The first identification of azaspiracids in shellfish from France and Spain. Toxicon 2003, 42, 105–108. [Google Scholar] [CrossRef]
- Twiner, M.J.; Rehmann, N.; Hess, P.; Doucette, G.J. Azaspiracid shellfish poisoning: A review on the chemistry, ecology, and toxicology with an emphasis on human health impacts. Mar. Drugs 2008, 6, 39–72. [Google Scholar] [CrossRef]
- Taleb, H.; Vale, P.; Amanhir, R.; Benhadouch, A.; Sagou, R.; Chafik, A. First detection of azaspiracids in mussels in northwest Africa. J. Shellfish Res. 2006, 25, 1067–1070. [Google Scholar]
- Klontz, K.C.; Abraham, A.; Plakas, S.M.; Dickey, R.W. Mussel-associated azaspiracid intoxication in the United States. Annal. Intern. Med. 2009, 150, 361. [Google Scholar]
- Dickey, R. Personal communication, US Food and Drug Administration: College Park, MD, USA, 2009.
- Twiner, M.J.; Hess, P.; Dechraoui, M.Y.; McMahon, T.; Samons, M.S.; Satake, M.; Yasumoto, T.; Ramsdell, J.S.; Doucette, G.J. Cytotoxic and cytoskeletal effects of azaspiracid-1 on mammalian cell lines. Toxicon 2005, 45, 891–900. [Google Scholar] [CrossRef]
- Colman, J.R.; Twiner, M.J.; Hess, P.; McMahon, T.; Satake, M.; Yasumoto, T.; Doucette, G.J.; Ramsdell, J.S. Teratogenic effects of azaspiracid-1 identified by microinjection of Japanese medaka (Oryzias latipes) embryos. Toxicon 2005, 45, 881–890. [Google Scholar] [CrossRef]
- Rundberget, T.; Sandvik, M.; Hovgaard, P.; Nguyen, L.; Aasen, J.A.B.; Castberg, T.; Gustad, E.; Miles, C.O. Use of SPATT disks in Norway: Detection of AZAs & DTXs and comparison with algal cell counts and toxin profiles in Shellfish. In Marine Biotoxin Science Workshop No. 23; New Zealand Food Safety Authority: Wellington, NZ, USA, 2006; pp. 35–38. [Google Scholar]
- Regulation (EC) no. 853/2004 of the European Parliament and of the Council of 29 April 2004. Off. J. Eur. Commun. 2004, L139, 55–205.
- SoundToxins Website. Available online: http://www.soundtoxins.org (accessed on 9 May 2013).
- ORHAB Website. Available online: http://www.orhab.org (accessed on 9 May 2013).
- EU-Harmonized Standard Operation Procedure for Determination of Lipophilic Marine Biotoxins in Molluscs by LC-MS/MS; Version 4; European Union Reference Laboratory for Marine Biotoxins (EU-RL-MB), Agencia Esponola de Seguridad Alimentaria y Nutricion (AESAN): Vigo, Spain, 2011; Available online: http://www.aesan.msps.es/CRLMB/docs/docs/metodos_analiticos_de_desarrollo/EU-Harmonised-SOP-LIPO-LCMSMS_Version4.pdf (accessed on 10 May 2013).
- Omni International Website. Available online: http://www.omni-inc.com (accessed on 9 May 2013).
- McNabb, P.; Sellwood, A.I.; Holland, P.T.; Aasen, J.; Aune, T.; Eaglesham, G.; Hess, P.; Igarishi, M.; Quilliam, M.; Slattery, D.; et al. Multiresidue method for determination of algal toxins in shellfish: Single-laboratory validation and interlaboratory study. J. AOAC Int. 2005, 88, 761–772. [Google Scholar]
- McCarron, P.; Giddings, S.; Quilliam, M. A mussel tissue certified reference material for multiple phycotoxins. Part 2: Liquid chromatography-mass spectroscopy, sample extraction and quantitation procedures. Anal. Bioanal. Chem. 2011, 400, 835–846. [Google Scholar] [CrossRef]
- Horner, R. A Taxonomic Guide to Some Common Marine Phytoplankton; Biopress Ltd.: Bristol, UK, 2002; p. 195. [Google Scholar]
- Environment Canada Website. Available online: http://www.wateroffice.ec.gc.ca/index_e.html (accessed on 9 May 2013).
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Trainer, V.L.; Moore, L.; Bill, B.D.; Adams, N.G.; Harrington, N.; Borchert, J.; Da Silva, D.A.M.; Eberhart, B.-T.L. Diarrhetic Shellfish Toxins and Other Lipophilic Toxins of Human Health Concern in Washington State. Mar. Drugs 2013, 11, 1815-1835. https://doi.org/10.3390/md11061815
Trainer VL, Moore L, Bill BD, Adams NG, Harrington N, Borchert J, Da Silva DAM, Eberhart B-TL. Diarrhetic Shellfish Toxins and Other Lipophilic Toxins of Human Health Concern in Washington State. Marine Drugs. 2013; 11(6):1815-1835. https://doi.org/10.3390/md11061815
Chicago/Turabian StyleTrainer, Vera L., Leslie Moore, Brian D. Bill, Nicolaus G. Adams, Neil Harrington, Jerry Borchert, Denis A. M. Da Silva, and Bich-Thuy L. Eberhart. 2013. "Diarrhetic Shellfish Toxins and Other Lipophilic Toxins of Human Health Concern in Washington State" Marine Drugs 11, no. 6: 1815-1835. https://doi.org/10.3390/md11061815