Hydrolysis of Fucoidan by Fucoidanase Isolated from the Marine Bacterium, Formosa algae

 ,
,
Abstract
:1. Introduction
2. Results and Discussion


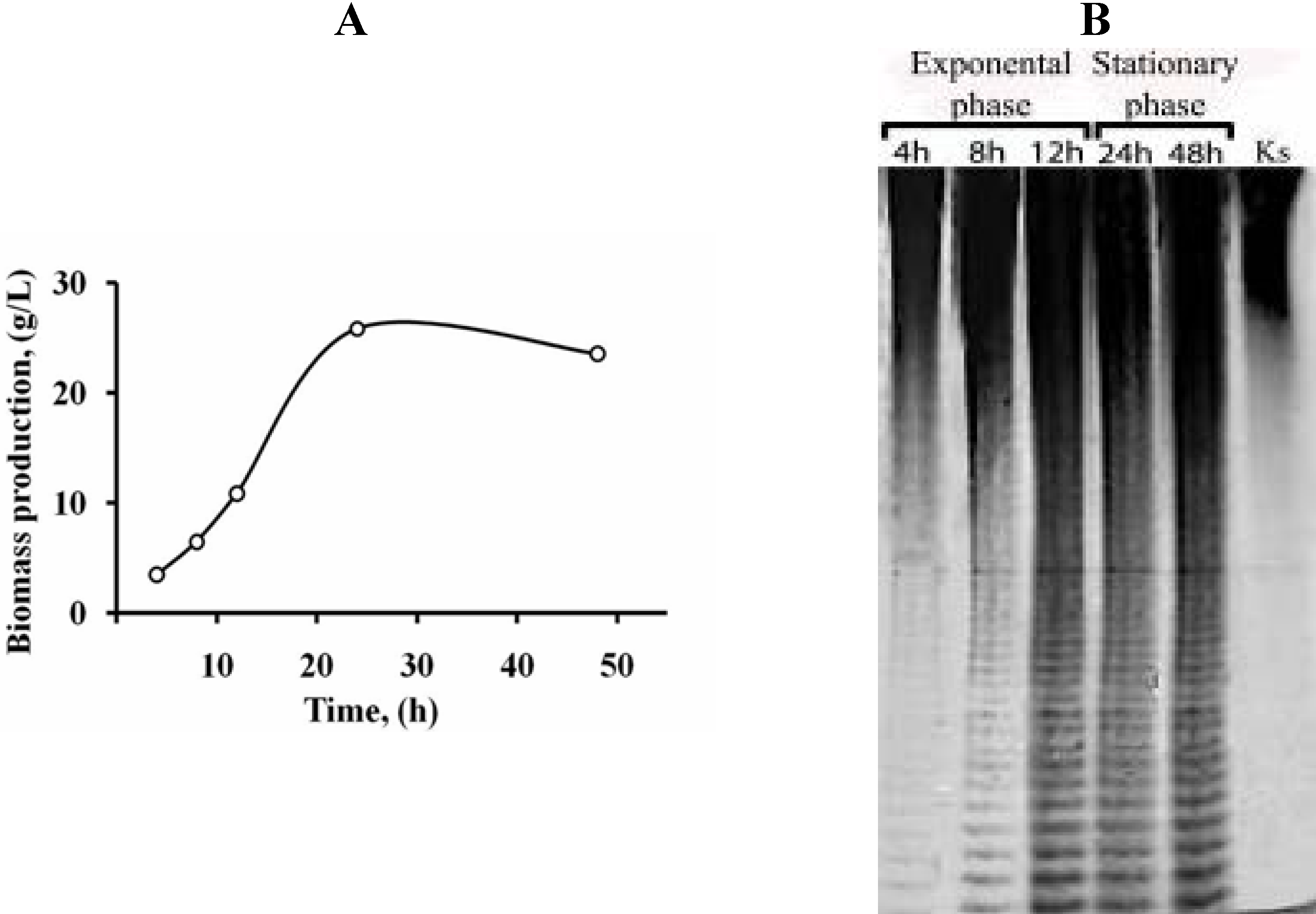{kind=link}
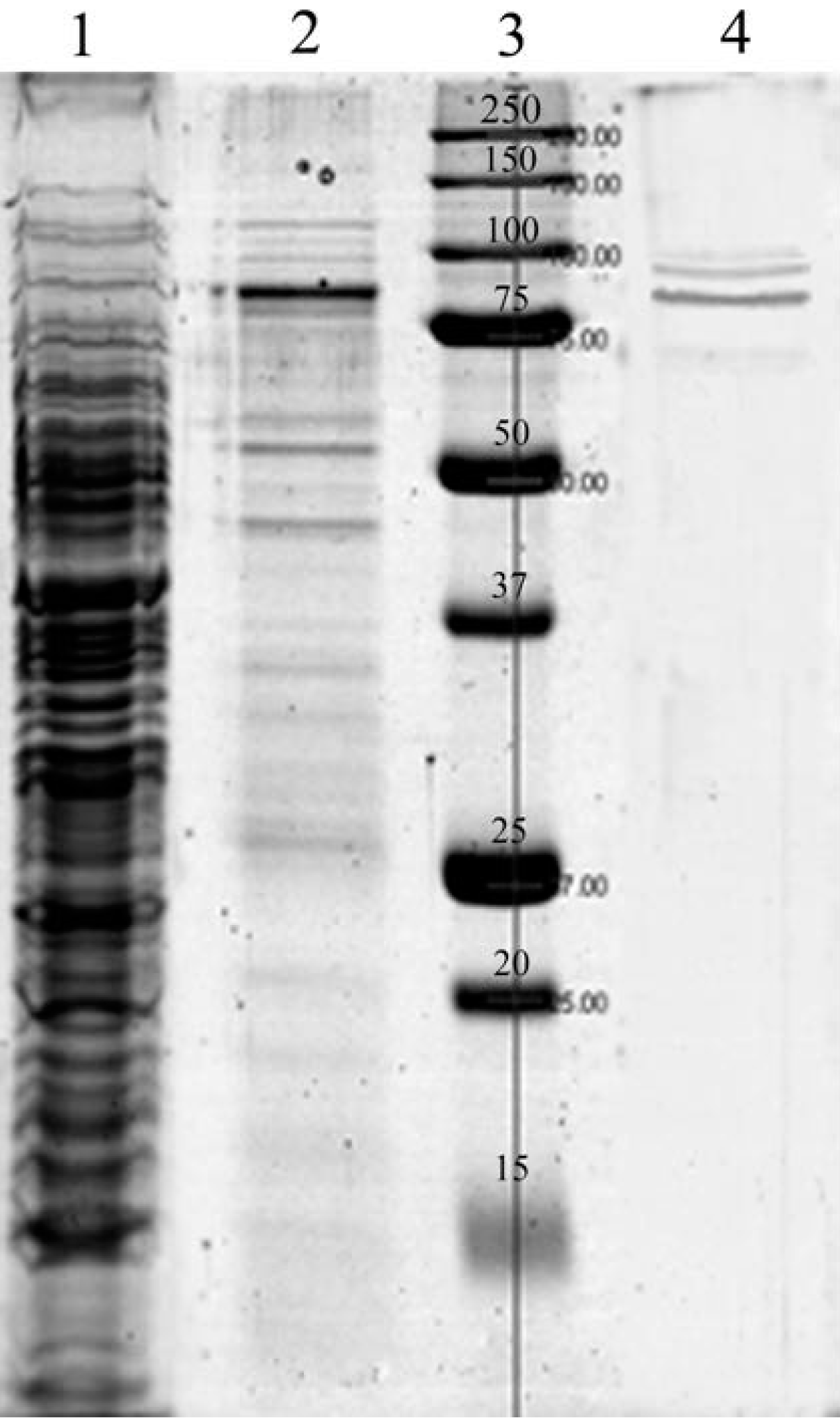{kind=link}
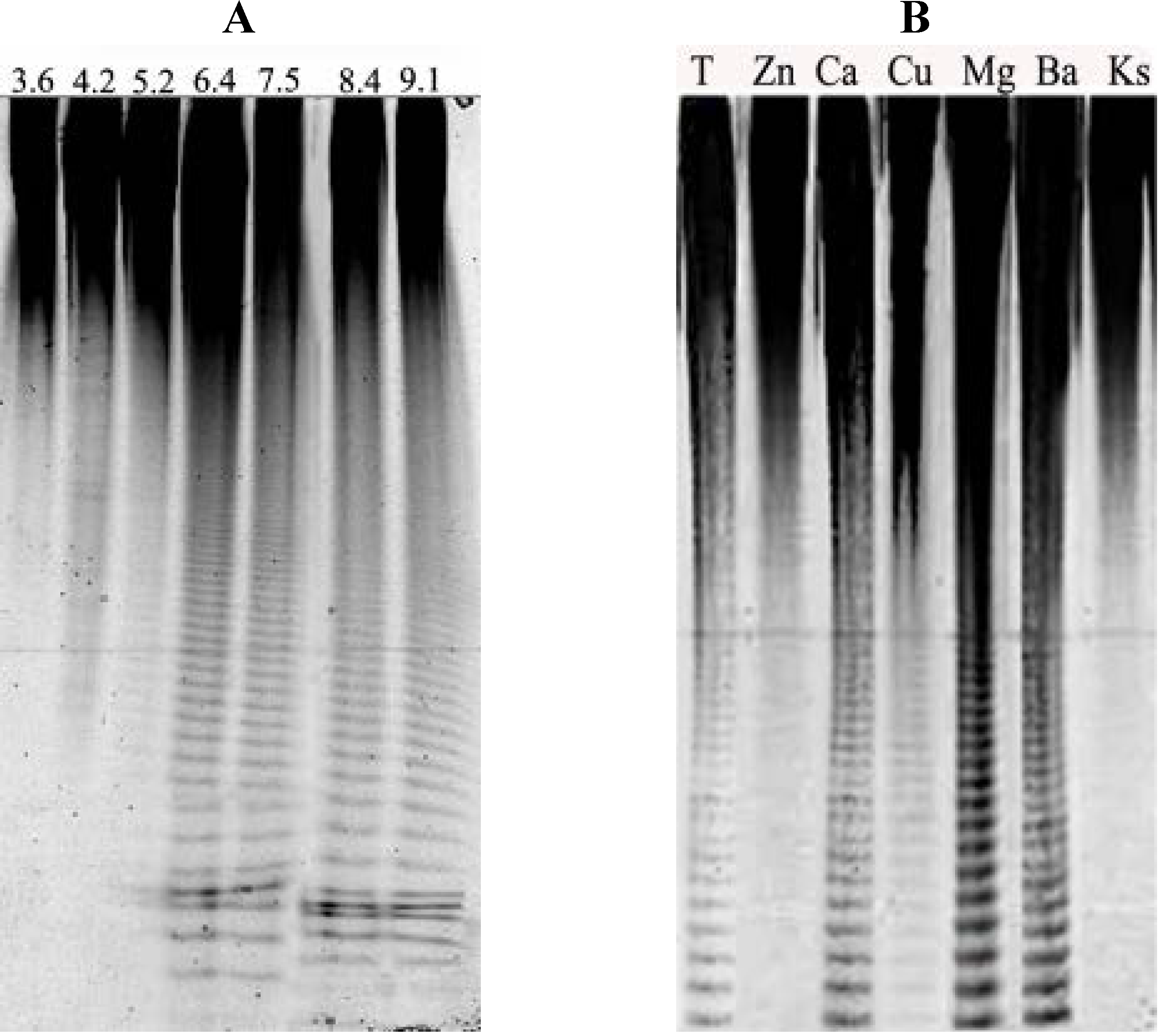{kind=link}
{kind=link}
{kind=link}
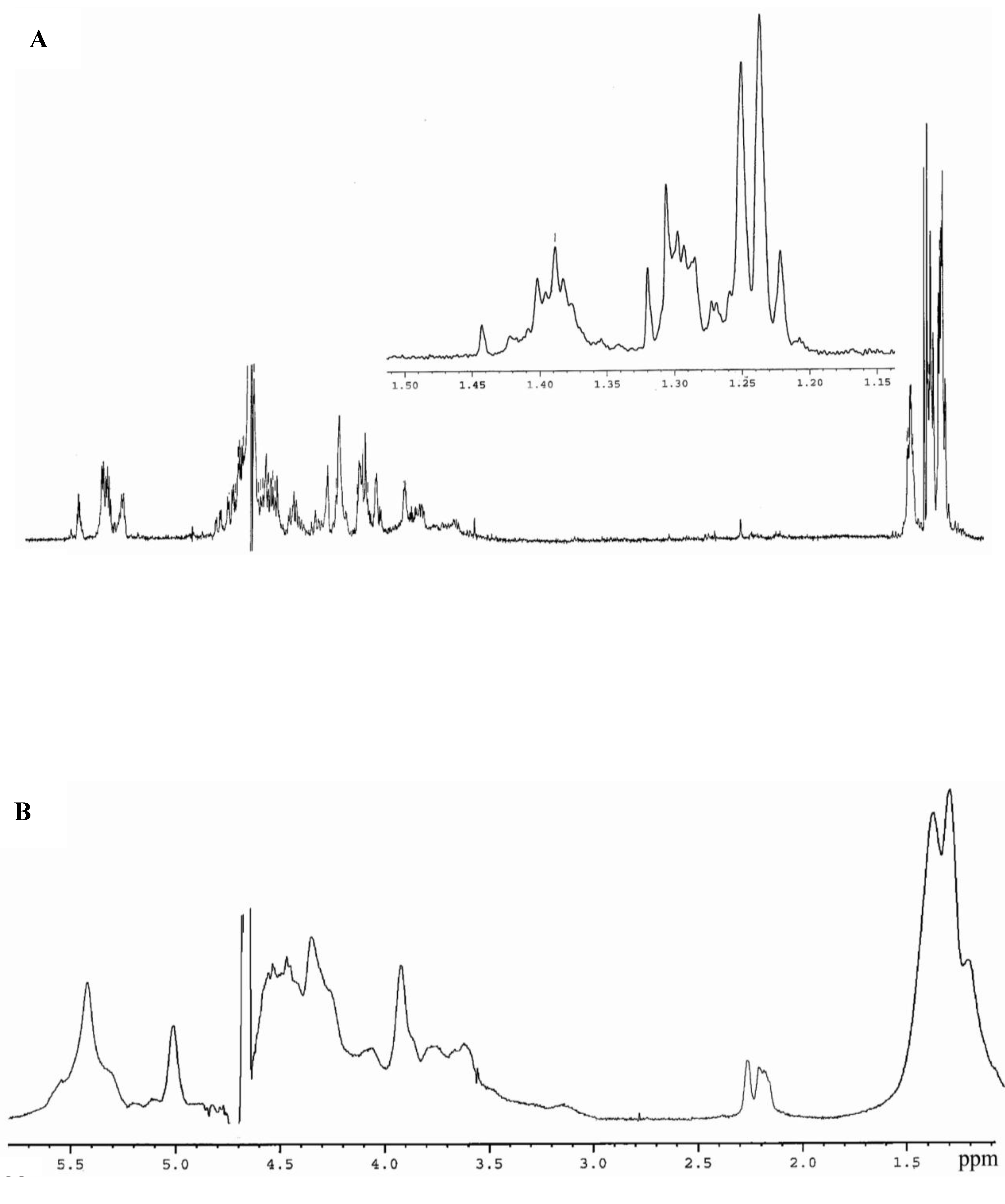{kind=link}
{kind=link}
| Enzyme substrate | Specific activity (U/mg of protein) * |
|---|---|
| Alginic acid | 6.4 |
| Amylopectin | 0 |
| СM-cellulose | 0 |
| Laminarin | 10.6 |
| Pustulan | 0 |
| p-Np-N-acetyl-β-d-glucosaminide | 20.7 |
| p-Np-β-d-galactopyranoside | 0 |
| p-Np-β-d-glucopyranoside | 9.5 |
| p-Np-β-d-mannopyranoside | 0 |
| p-Np-α-d-fucopyranoside | 0 |
| p-Np-sulfate | 0 |



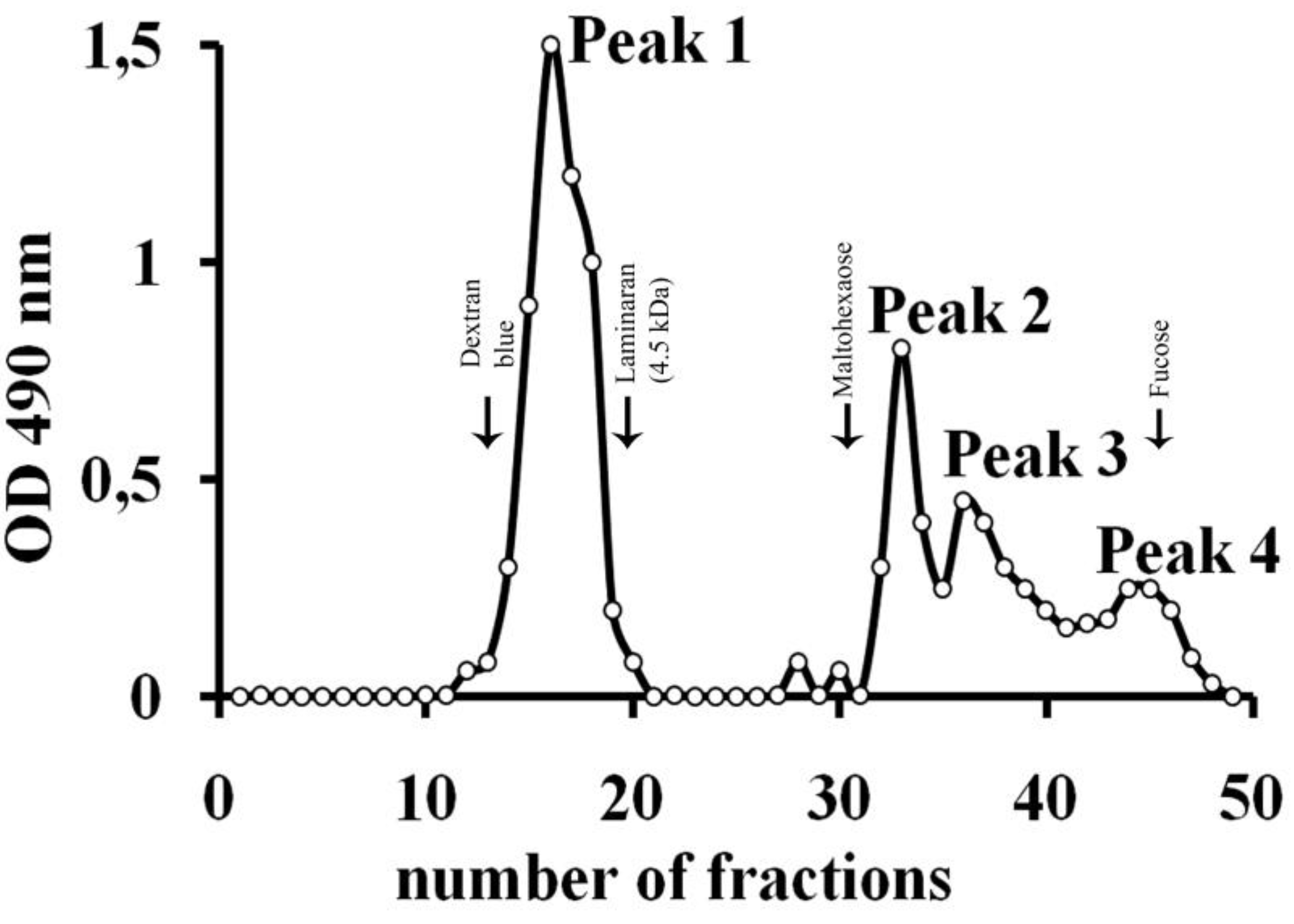
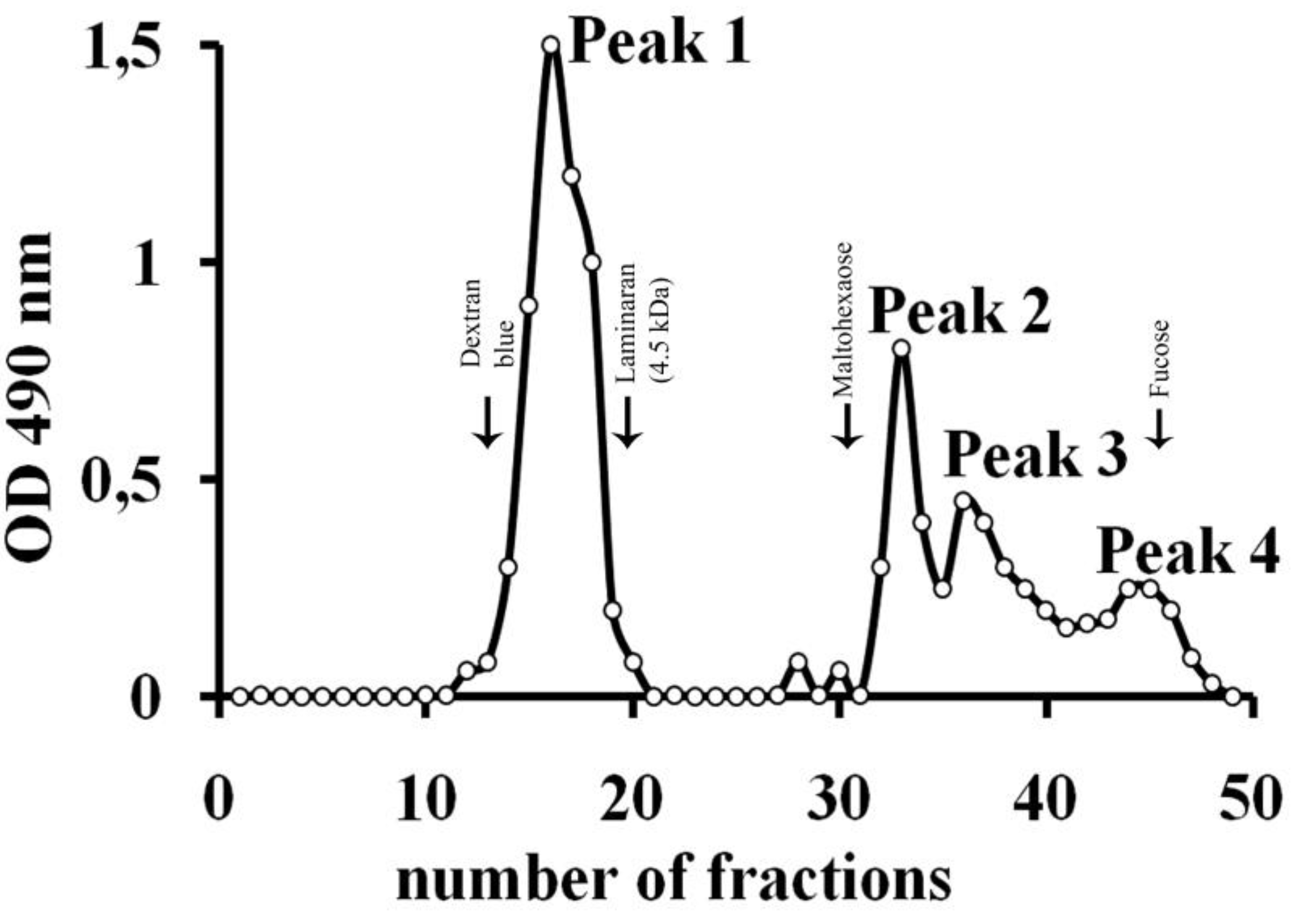
| Substrate | Yield of hydrolysis products, % |
|---|---|
| Fucoidan from F. evanescens | 7 |
| F. dAc* | 9.4 |
| F. dS* | 0.8 |
| Fucoidan from F. vesiculosus | 5.6 |
| Carrageenan | 0 |
| Fucoidan from S. cichorioides | 0 |
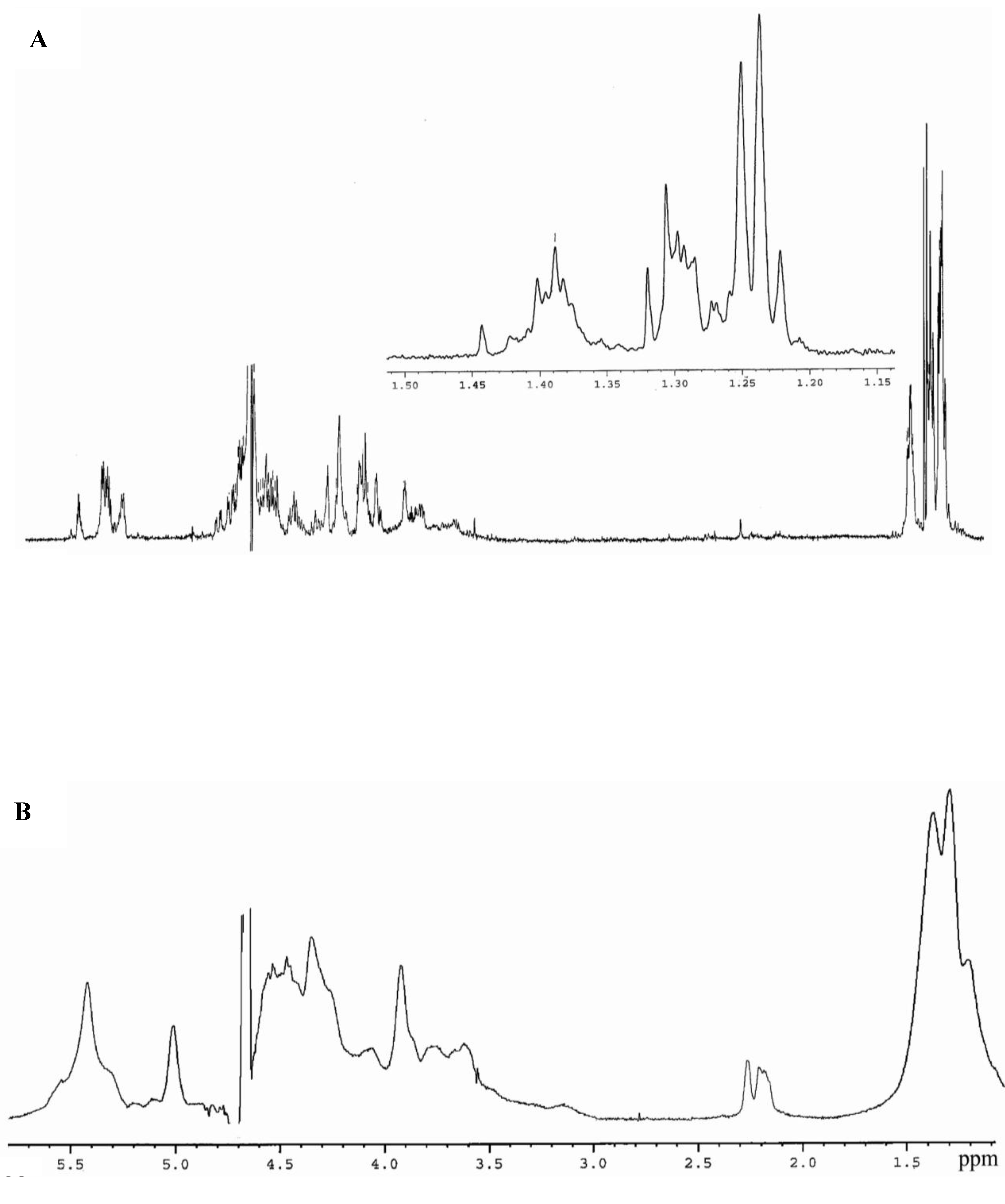
| Residue | H1 | H2 | H3 | H4 | H5 | H6 |
| A | 5.32 | 4.45 | 4.08 | 3.88 | 4.22 | 1.23 |
| B | 5.48 | 4.50 | 4.02 | 4.06 | 4.47 | 1.22 |
| C | 5.34 | 4.56 | 4.70 | 4.21 | 4.53 | 1.24 |
| D | 5.50 | 4.51 | 4.03 | 4.08 | 4.48 | 1.23 |
| Residue | C1 | C2 | C3 | C4 | C5 | C6 |
| A | 95.3 | 76.4 | 69.9 | 73.3 | 67.1 | 16.0 |
| B | 91.7 | 74.6 | 73.9 | 68.6 | 67.9 | 16.1 |
| C | 96.4 | 73.6 | 76.2 | 71.7 | 67.8 | 16.3 |
| D | 91.7 | 74.6 | 75.2 | 70.5 | 67.8 | 16.2 |
| Lymphocyte activation marker | Control M ± σ | Fucoidan from F. evanescens M ± σ | HMP M ± σ | “Peak 2” M ± σ |
|---|---|---|---|---|
| CD16+ CD69+ (percentage of CD16+-lymphocytes) | 33.4 ± 11.9 | 88.1 ± 6.5 p = 0.000 | 93.2 ± 4.4 p = 0.000 | 90.8 ± 5.1 p = 0.000 |
| CD16+ CD25+ (percentage of CD16+-lymphocytes) | 2.8 ± 2.0 | 17.4 ± 5.89 p = 0.000 | 21.9 ± 4.9 p = 0.000 | 19.3 ± 5.9 p = 0.000 |
| CD3+ CD69+ (percentage of CD3+-lymphocytes) | 14.9 ± 5.2 | 25.3 ± 12.9 p = 0.008 | 30.1 ± 13.0 p = 0.000 | 37.1 ± 15.3 p = 0.002 |
| CD3+ CD25+ (percentage of CD3+-lymphocytes) | 11.8 ± 3.1 | 16.6 ± 3.9 p = 0.000 | 19.2 ± 3.2 p = 0.000 | 19.1 ± 3.0 p = 0.000 |
3. Experimental Section
3.1. Reagents
3.2. General Methods
3.3. Enzyme Activity Assay
3.4. Bacterial Culture Conditions
3.5. NMR Spectroscopy
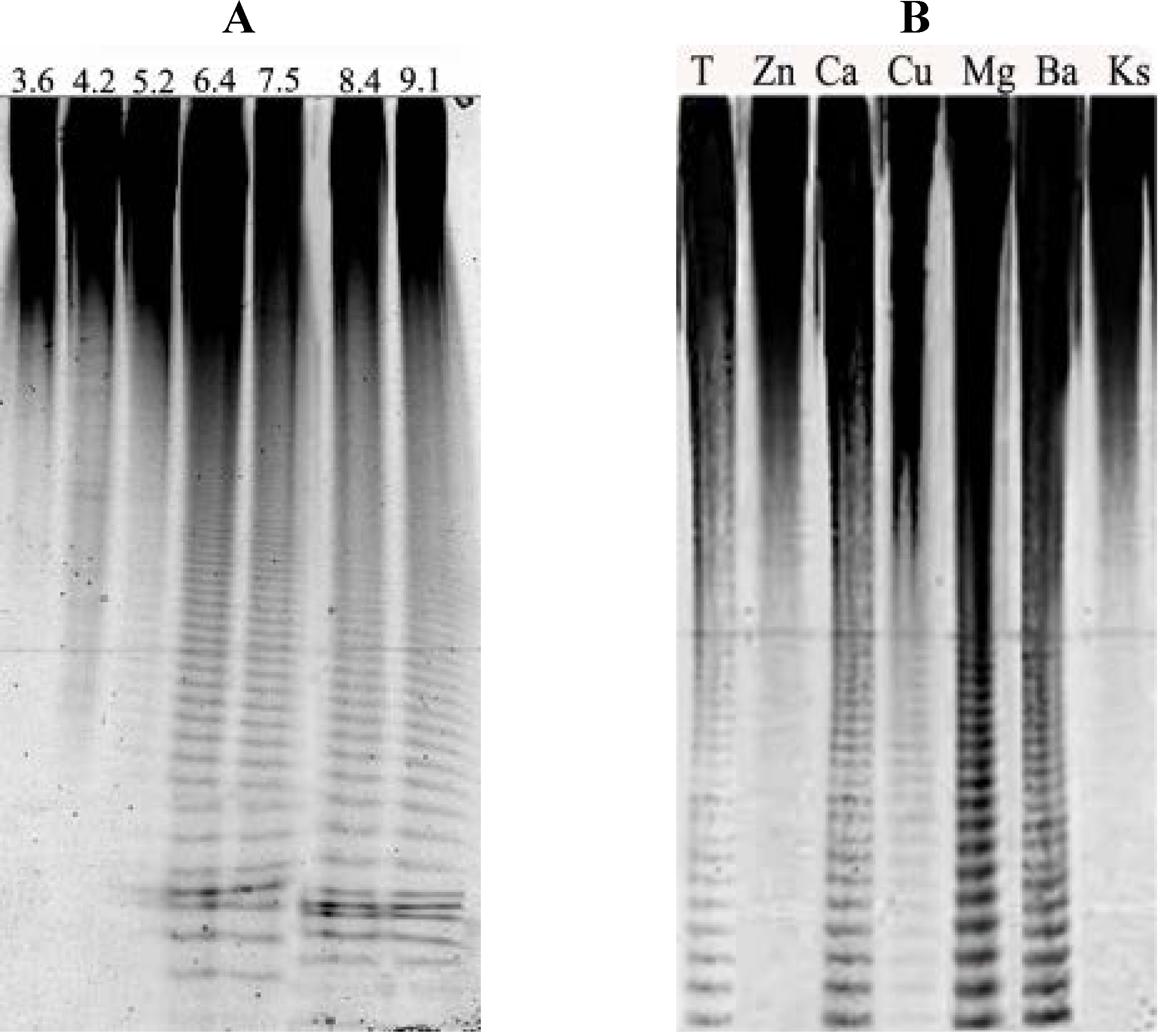
3.6. Purification of Fucoidanase
3.7. Determination of the Optimum pH for Fucoidanase
3.8. Determination of Fucoidanase Stability at Different Temperatures
3.9. Determination of the Yield of the Low Molecular Weight Products
3.10. Preparation of Fucoidan Oligosaccharides
3.11. Influence of Bivalent Metals
3.12. Kinetics of the Enzymatic Reaction
3.13. Flow Cytometric Measurement of CD69 and CD25
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Kusaykin, M.; Bakunina, I.; Sova, V.; Ermakova, S.; Kuznetsova, T.; Besednova, N.; Zaporozhets, T.; Zvyagintseva, T. Structure, biological activity, and enzymatic transformation of fucoidans from the brown seaweeds. Biotechnol. J. 2008, 3, 904–915. [Google Scholar] [CrossRef]
- Usov, A.I.; Bilan, M.I. Fucoidans—Sulfated polysaccharides of brown algae. Russ. Chem. Rev. 2009, 78, 785–799. [Google Scholar] [CrossRef]
- Li, B.; Lu, F.; Wei, X.; Zhao, R. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar] [CrossRef]
- Holtkamp, A.D.; Kelly, S.; Ulber, R.; Lang, S. Fucoidans and fucoidanases—Focus on techniques for molecular structure elucidation and modification of marine polysaccharides. Appl. Microbiol. Biotechnol. 2009, 82, 1–11. [Google Scholar] [CrossRef]
- Fitton, J.H. Fucoidans: Healthful saccharides from the sea. GlycoSci. Nutr. 2005, 6, 1–6. [Google Scholar]
- Bilan, M.I.; Grachev, A.A.; Shashkov, A.S.; Nifantiev, N.E.; Usov, A.I. Structure of a fucoidan from the brown seaweed Fucus serratus L. Carbohydr. Res. 2006, 341, 238–245. [Google Scholar] [CrossRef]
- Bilan, M.I.; Grachev, A.A.; Ustuzhanina, N.E.; Shashkov, A.S.; Nifantiev, N.E.; Usov, A.I. Structure of a fucoidan from the brown seaweed Fucus evanescens C. Ag. Carbohydr. Res. 2002, 337, 719–730. [Google Scholar] [CrossRef]
- Bilan, M.I.; Grachev, A.A.; Ustuzhanina, N.E.; Shashkov, A.S.; Nifantiev, N.E.; Usov, A.I. A highly regular fraction of a fucoidan from the brown seaweed Fucus distichus L. Carbohydr. Res. 2004, 339, 511–517. [Google Scholar] [CrossRef]
- Chevolot, L.; Mulloy, B.; Ratiskol, J.; Foucault, A.; Colliec-Jouault, S. A disaccharide repeat unit is the major structure in fucoidans from two species of brown algae. Carbohydr. Res. 2001, 330, 529–535. [Google Scholar] [CrossRef]
- Zvyagintseva, T.N.; Shevchenko, N.M.; Chizhov, A.O.; Krupnova, T.N.; Sundukova, E.V.; Isakov, V.V. Water-soluble polysaccharides of some far-eastern brown seaweeds. Distribution, structure, and their dependence on the developmental conditions. J. Exp. Mar. Biol. Ecol. 2003, 294, 1–13. [Google Scholar] [CrossRef]
- Chizhov, A.O.; Dell, A.; Morris, H.R.; Haslam, S.M.; McDowell, R.A.; Shashkov, A.S.; Nifant’ev, N.E.; Khatuntseva, E.A.; Usov, A.I. A study of fucoidan from the brown seaweed Chorda filum. Carbohydr. Res. 1999, 320, 108–119. [Google Scholar] [CrossRef]
- Ermakova, S.; Sokolova, R.; Kim, S.M.; Um, B.H.; Isakov, V.; Zvyagintseva, T. Fucoidans from brown seaweeds Sargassum hornery, Eclonia cava, Costaria costata: Structural characteristics and anticancer activity. Appl. Biochem. Biotechnol. 2011, 164, 841–850. [Google Scholar] [CrossRef]
- Vishchuk, O.S.; Ermakova, S.P.; Zvyagintseva, T.N. Sulfated polysaccharides from brown seaweeds Saccharina japonica and Undaria pinnatifida: Isolation, structural characteristics, and antitumor activity. Carbohydr. Res. 2011, 346, 2769–2776. [Google Scholar] [CrossRef]
- Zapopozhets, T.S.; Besednova, N.N.; Loenko Iu, N. Antibacterial and immunomodulating activity of fucoidan. Antibiot. Khimioter. 1995, 40, 9–13. [Google Scholar]
- Zaporozhets, T.S.; Ivanushko, L.A.; Zvyagintseva, T.A.; Molchanova, V.I.; Besednova, N.N.; Epstein, L.M. Cytokine inducing activity of biopolymers from sea hydrobionts. Med. Immunol. 2004, 6, 89–96. [Google Scholar]
- Zaporozhets, T.S.; Kuznetsova, T.A.; Smolina, T.P.; Shevchenko, N.M.; Zvyagintseva, T.N.; Besednova, N.N. Immunotropic and аnticoagulant activity of fucoidan from brown seaweed Fucus evanescens: Prospects of application in medicine. J. Microbiol. 2006, 54–58. [Google Scholar]
- Hirmo, S.; Utt, M.; Ringner, M.; Wadstrom, T. Inhibition of heparan sulphate and other glycosaminoglycans binding to Helicobacter pylori by various polysulphated carbohydrates. FEMS Immunol. Med. Microbiol. 1995, 10, 301–306. [Google Scholar] [CrossRef]
- Shibata, H.; KimuraTakagi, I.; Nagaoka, M.; Hashimoto, S.; Sawada, H.; Ueyama, S.; Yokokura, T. Inhibitory effect of Cladosiphon fucoidan on the adhesion of Helicobacter pylori to human gastric cells. J. Nutr. Sci. Vitaminol.(Tokyo) 1999, 45, 325–336. [Google Scholar] [CrossRef]
- Adhikari, U.; Mateu, C.G.; Chattopadhyay, K.; Pujol, C.A.; Damonte, E.B.; Ray, B. Structure and antiviral activity of sulfated fucans from Stoechospermum marginatum. Phytochemistry 2006, 67, 2474–2482. [Google Scholar] [CrossRef]
- Lee, J.B.; Hayashi, K.; Hashimoto, M.; Nakano, T.; Hayashi, T. Novel antiviral fucoidan from sporophyll of Undaria pinnatifida (Mekabu). Chem. Pharm. Bull. (Tokyo) 2004, 52, 1091–1094. [Google Scholar] [CrossRef]
- McClure, M.O.; Moore, J.P.; Blanc, D.F.; Scotting, P.; Cook, G.M.; Keynes, R.J.; Weber, J.N.; Davies, D.; Weiss, R.A. Investigations into the mechanism by which sulfated polysaccharides inhibit HIV infection in vitro. AIDS Res. Hum. Retroviruses 1992, 8, 19–26. [Google Scholar] [CrossRef]
- Ponce, N.M.; Pujol, C.A.; Damonte, E.B.; Flores, M.L.; Stortz, C.A. Fucoidans from the brown seaweed Adenocystis utricularis: Extraction methods, antiviral activity and structural studies. Carbohydr. Res. 2003, 338, 153–165. [Google Scholar] [CrossRef]
- Cumashi, A.; Ushakova, N.A.; Preobrazhenskaya, M.E.; D’Incecco, A.; Piccoli, A.; Totani, L.; Tinari, N.; Morozevich, G.E.; Berman, A.E.; Bilan, M.I.; et al. A comparative study of the antiinflammatory, anticoagulant, antiangiogenic, and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology 2007, 17, 541–552. [Google Scholar]
- Mourao, P.A. Use of sulfated fucans as anticoagulant and antithrombotic agents: Future perspectives. Curr. Pharm. Des. 2004, 10, 967–981. [Google Scholar] [CrossRef]
- Bakunina, I.; Nedashkovskaia, O.I.; Alekseeva, S.A.; Ivanova, E.P.; Romanenko, L.A.; Gorshkova, N.M.; Isakov, V.V.; Zviagintseva, T.N.; Mikhailov, V.V. Degradation of fucoidan by the marine proteobacterium Pseudoalteromonas citrea. Mikrobiologiia (Moscow) 2002, 71, 49–55. [Google Scholar]
- Bakunina, I.; Shevchenko, L.S.; Nedashkovskaia, O.I.; Shevchenko, N.M.; Alekseeva, S.A.; Mikhailov, V.V.; Zviagintseva, T.N. Screening of marine bacteria for fucoidan hydrolases. Mikrobiologiia (Moscow) 2000, 69, 370–376. [Google Scholar]
- Descamps, V.; Colin, S.; Lahaye, M.; Jam, M.; Richard, C.; Potin, P.; Barbeyron, T.; Yvin, J.C.; Kloareg, B. Isolation and culture of a marine bacterium degrading the sulfated fucans from marine brown algae. Mar. Biotechnol. (N. Y.) 2006, 8, 27–39. [Google Scholar] [CrossRef]
- Bilan, M.I.; Kusaykin, M.I.; Grachev, A.A.; Tsvetkova, E.A.; Zvyagintseva, T.N.; Nifantiev, N.E.; Usov, A.I. Effect of enzyme preparation from the marine mollusk Littorina kurila on fucoidan from the brown alga Fucus distichus. Biochemistry (Moscow) 2005, 70, 1321–1326. [Google Scholar] [CrossRef]
- Daniel, R.; Berteau, O.; Jozefonvicz, J.; Goasdoue, N. Degradation of algal (Ascophyllum nodosum) fucoidan by an enzymatic activity contained in digestive glands of the marine mollusk Pecten maximus. Carbohydr. Res. 1999, 322, 291–297. [Google Scholar] [CrossRef]
- Kitamura, K.; Matsuo, M.; Yasui, T. Enzymatic degradation of fucoidan by fucoidanase from the hepatopancreas of Patinopecten yessoensis. Biosci. Biotechnol. Biochem. 1992, 56, 490–494. [Google Scholar] [CrossRef]
- Kusaykin, M.I.; Burtseva, Y.V.; Svetasheva, T.G.; Sova, V.V.; Zvyagintseva, T.N. Distribution of O-glycosylhydrolases in marine invertebrates. Enzymes of the marine mollusk Littorina kurila that catalyze fucoidan transformation. Biochemistry (Moscow) 2003, 68, 317–324. [Google Scholar]
- Thanassi, N.M.; Nakada, H.I. Enzymatic dagradation of fucoidan by enzymes from the hepatopancreas of abalone Haliotis species. Arch. Biochem. Biophys. 1967, 118, 172–177. [Google Scholar] [CrossRef]
- Rodriguez-Jasso, R.M.; Mussatto, S.I.; Pastrana, L.; Aguilar, C.N.; Teixeira, J.A. Fucoidan-degrading fungal strains: Screening, morphometric evaluation, and influence of medium composition. Appl. Biochem. Biotechnol. 2010, 162, 2177–2188. [Google Scholar] [CrossRef] [Green Version]
- Colin, S.; Deniaud, E.; Jam, M.; Descamps, V.; Chevolot, Y.; Kervarec, N.; Yvin, J.C.; Barbeyron, T.; Michel, G.; Kloareg, B. Cloning and biochemical characterization of the fucanase FcnA: Definition of a novel glycoside hydrolase family specific for sulfated fucans. Glycobiology 2006, 16, 1021–1032. [Google Scholar] [CrossRef]
- Takayama, M.; Koyama, N.; Sakai, T.; Kato, I. Enzymes Capable of Degrading a Sulfated-Fucose-Containing Polysaccharide and Their Encoding Genes. U.S. Patent 6,489,155, 2002. [Google Scholar]
- Ermakova, S.P.; Ivanova, E.P.; Bakunina, I.; Mikhailov, V.V.; Zviagintseva, T.N. Brown algae metabolites influence on o-glycoside hydrolases synthesis of bacteria degrading Fucus evanescens tallom. Mikrobiologiia (Moscow) 2012, 81, 396–402. [Google Scholar]
- Ivanova, E.P.; Alexeeva, Y.V.; Flavier, S.; Wright, J.P.; Zhukova, N.V.; Gorshkova, N.M.; Mikhailov, V.V.; Nicolau, D.V.; Christen, R. Formosa algae gen. nov., sp nov., a novel member of the family Flavobacteriaceae. Int. J. Syst. Evol. Microbiol. 2004, 54, 705–711. [Google Scholar] [CrossRef]
- Furukawa, S.; Fujikawa, T.; Koga, D.; Ide, A. Purification and some properties of exo-type fucoidanases from Vibrio sp. N-5. Biosci. Biotechnol. Biochem. 1992, 56, 1829–1834. [Google Scholar] [CrossRef]
- Sakai, T.; Kawai, T.; Kato, I. Isolation and characterization of a fucoidan-degrading marine bacterial strain and its fucoidanase. Mar. Biotechnol. (N. Y.) 2004, 6, 335–346. [Google Scholar] [CrossRef]
- Sakai, T.; Ishizuka, K.; Shimanaka, K.; Ikai, K.; Kato, I. Structures of oligosaccharides derived from Cladosiphon okamuranus fucoidan by digestion with marine bacterial enzymes. Mar. Biotechnol. (N. Y.) 2003, 5, 536–544. [Google Scholar] [CrossRef]
- Khilchenko, S.R.; Zaporozhets, T.S.; Shevchenko, N.M.; Zvyagintseva, T.N.; Vogel, U.; Seeberger, P.; Lepenies, B. Immunostimulatory activity of fucoidan from the brown alga Fucus evanescens: Role of sulfates and acetates. J. Carbohydr. Chem. 2011, 30, 291–305. [Google Scholar]
- Jin, J.O.; Park, H.Y.; Xu, Q.; Park, J.I.; Zvyagintseva, T.; Stonik, V.A.; Kwak, J.Y. Ligand of scavenger receptor class A indirectly induces maturation of human blood dendritic cells via production of tumor necrosis factor-alpha. Blood 2009, 113, 5839–5847. [Google Scholar] [CrossRef]
- Zvyagintseva, T.N.; Shevchenko, N.M.; Popivnich, I.B.; Isakov, V.V.; Scobun, A.S.; Sundukova, E.V.; Elyakova, L.A. A new procedure for the separation of water-soluble polysaccharides from brown seaweeds. Carbohydr. Res. 1999, 322, 32–39. [Google Scholar] [CrossRef]
- Kusaykin, M.I.; Chizhov, A.O.; Grachev, A.A.; Alekseeva, S.A.; Yu Bakunina, I.; Nedashkovskaya, O.I.; Sova, V.V.; Zvyagintseva, T.N. A comparative study of specificity of fucoidanases from marine microorganisms and invertebrates. J. Appl. Phycol. 2006, 18, 369–373. [Google Scholar] [CrossRef]
- Zvyagintseva, T.N.; Elyakova, L.A.; Sundukova, E.V.; Mishenko, N.P.; Krivoschekova, V.E. Method for Producing of Pustulan. SU Patent 1,227,199, 1986. [Google Scholar]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Nelson, N. A photometric adaptation of the Somogyi method for the determination of glucose. J. Biol. Chem. 1944, 153, 375–380. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Silchenko, A.S.; Kusaykin, M.I.; Kurilenko, V.V.; Zakharenko, A.M.; Isakov, V.V.; Zaporozhets, T.S.; Gazha, A.K.; Zvyagintseva, T.N. Hydrolysis of Fucoidan by Fucoidanase Isolated from the Marine Bacterium, Formosa algae. Mar. Drugs 2013, 11, 2413-2430. https://doi.org/10.3390/md11072413
Silchenko AS, Kusaykin MI, Kurilenko VV, Zakharenko AM, Isakov VV, Zaporozhets TS, Gazha AK, Zvyagintseva TN. Hydrolysis of Fucoidan by Fucoidanase Isolated from the Marine Bacterium, Formosa algae. Marine Drugs. 2013; 11(7):2413-2430. https://doi.org/10.3390/md11072413
Chicago/Turabian StyleSilchenko, Artem S., Mikhail I. Kusaykin, Valeriya V. Kurilenko, Alexander M. Zakharenko, Vladimir V. Isakov, Tatyana S. Zaporozhets, Anna K. Gazha, and Tatyana N. Zvyagintseva. 2013. "Hydrolysis of Fucoidan by Fucoidanase Isolated from the Marine Bacterium, Formosa algae" Marine Drugs 11, no. 7: 2413-2430. https://doi.org/10.3390/md11072413