Bioactive Compounds Offered in Microcapsules to Determine the Nutritional Value of Copepods’ Natural Diet

Abstract
:1. Introduction
2. Results and Discussion
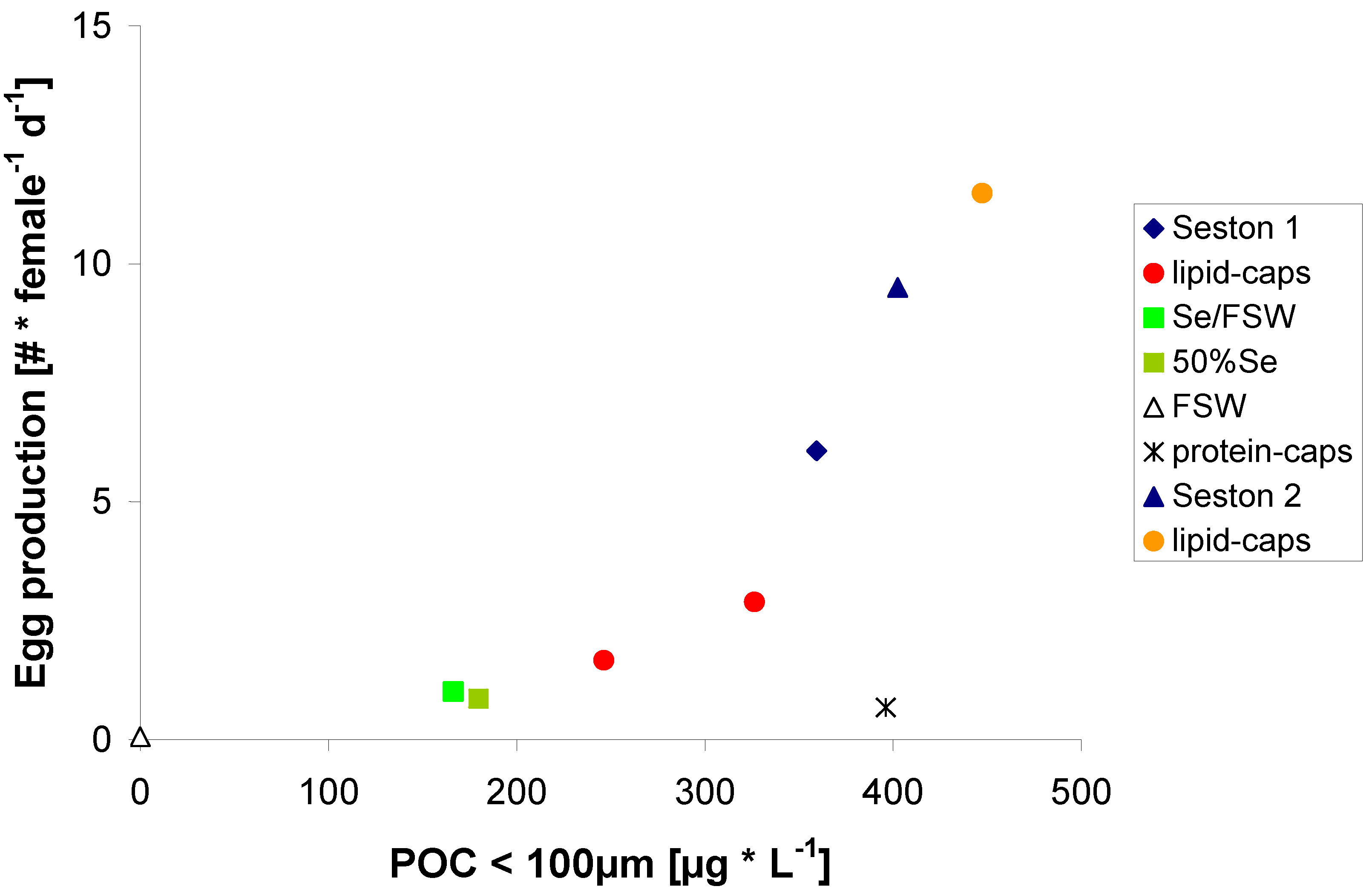
2.1. Uptake and Suitability of μ-Capsules
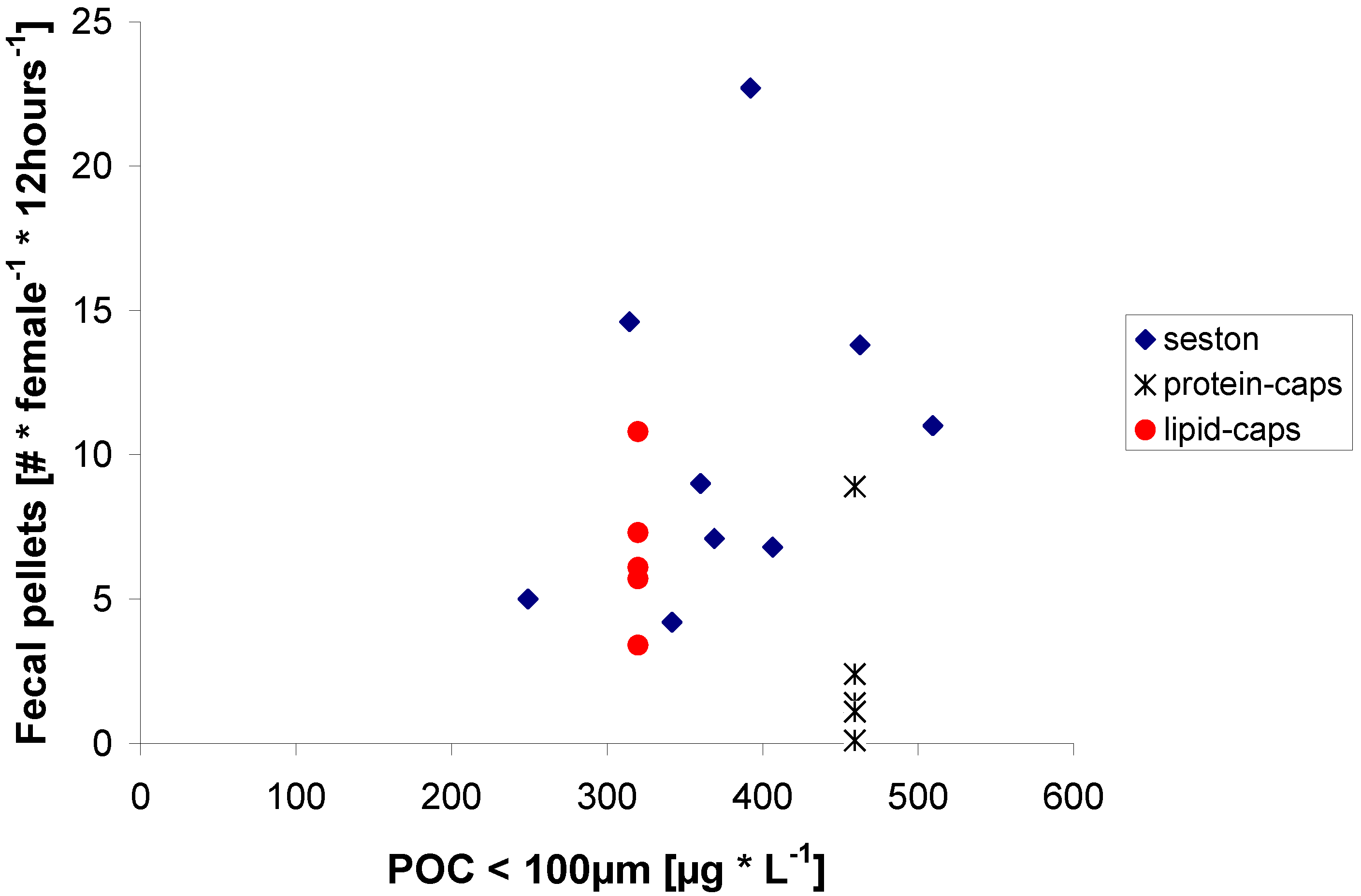
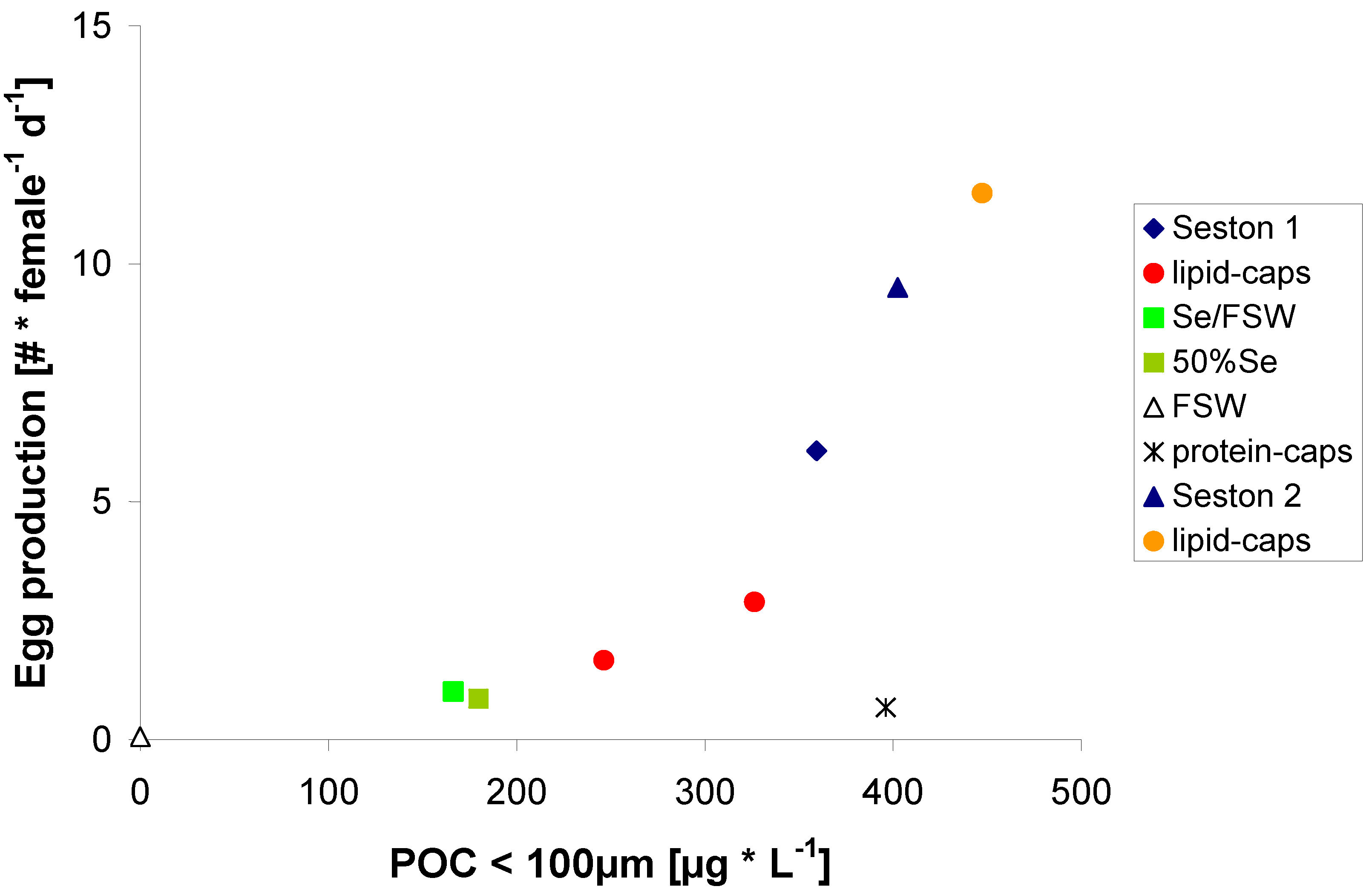
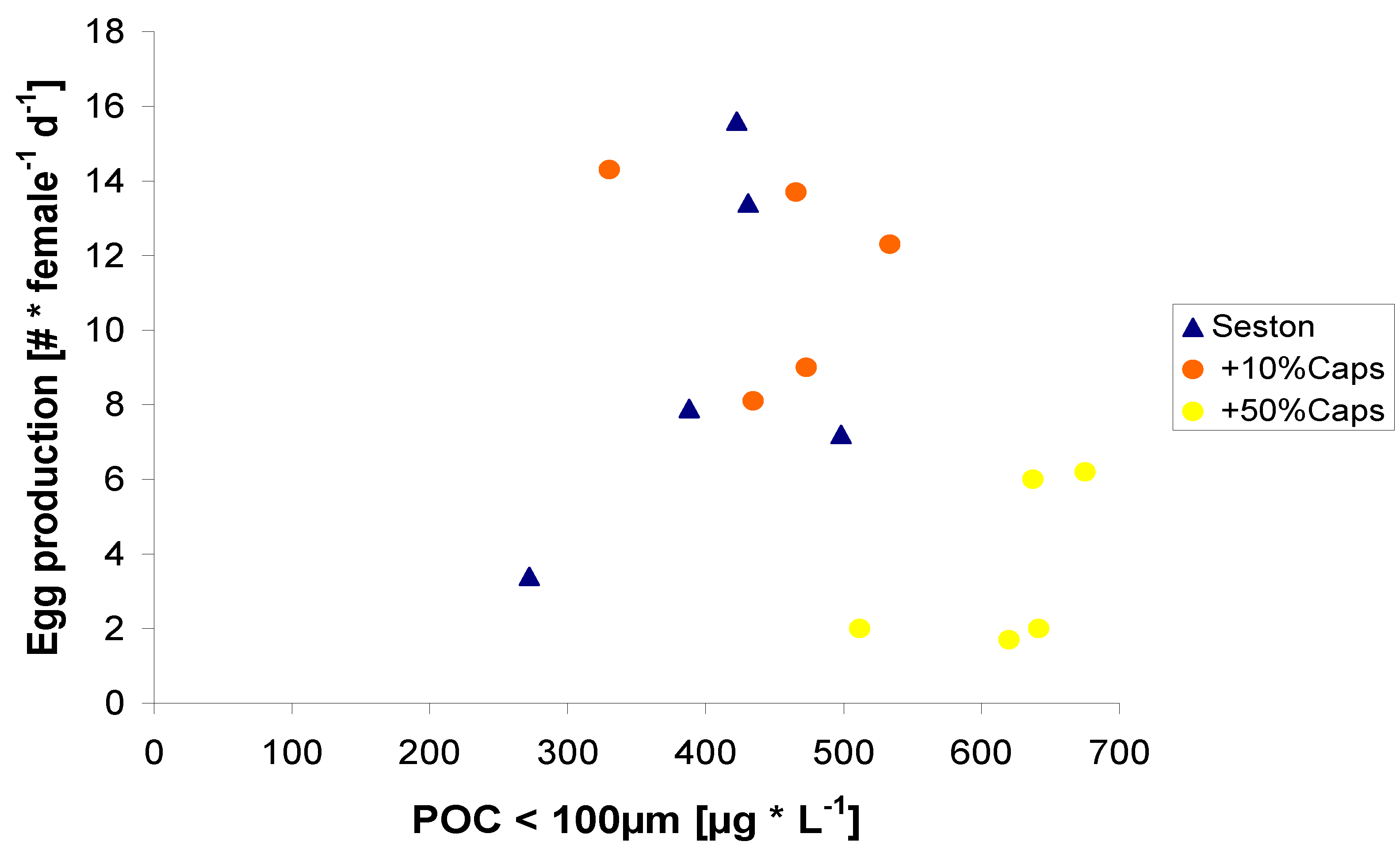
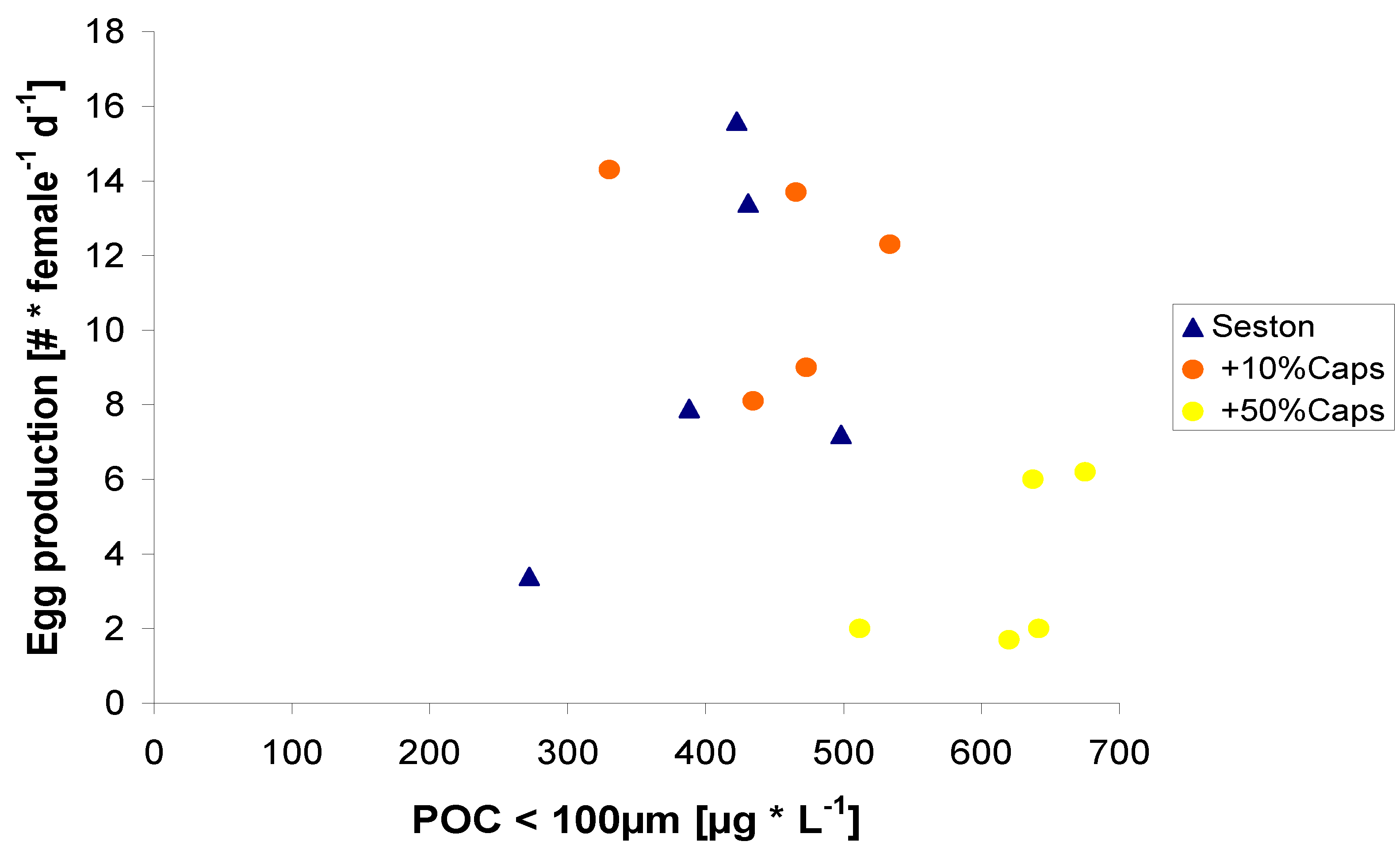
2.2. Egg Production in Relation to Food Concentration
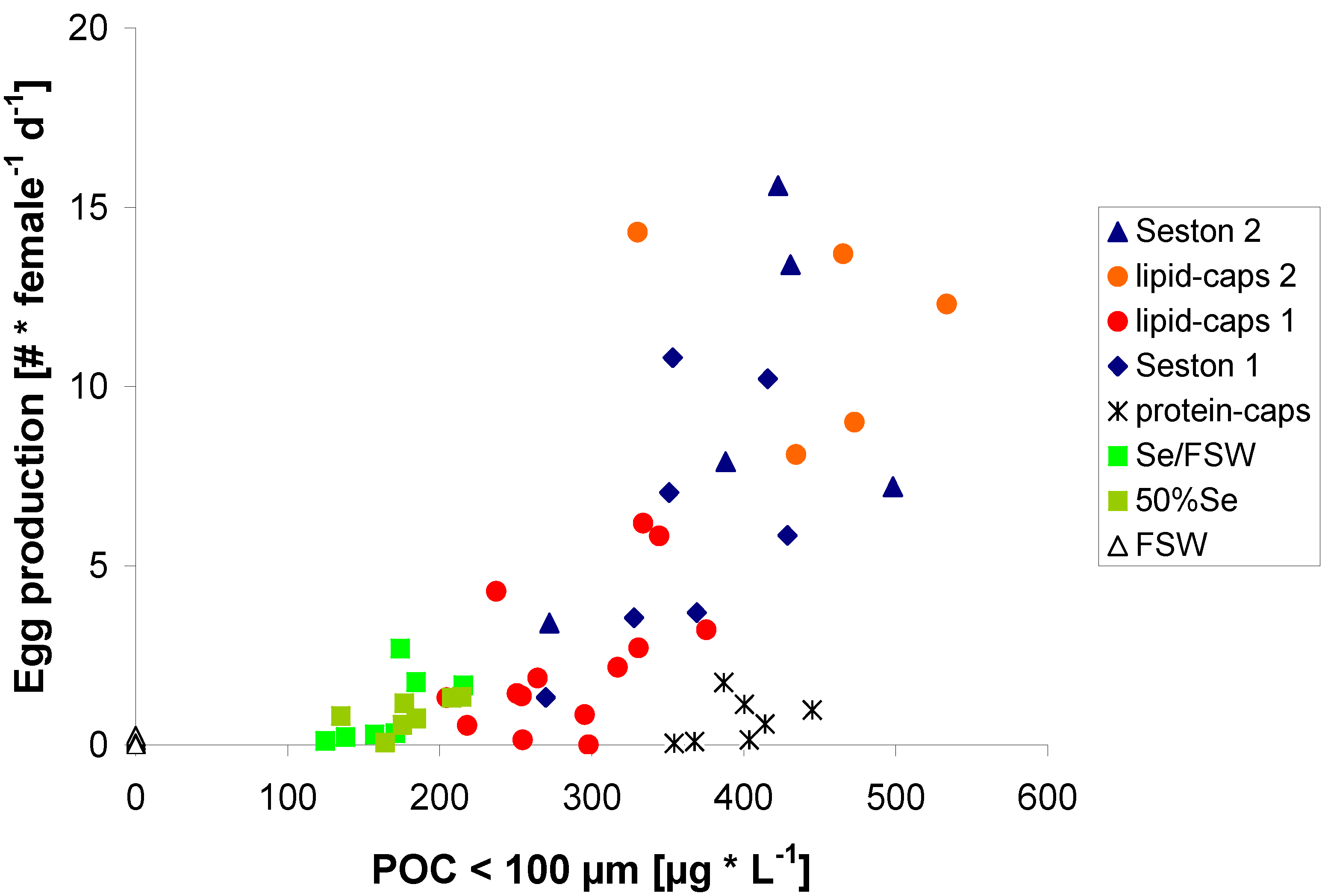

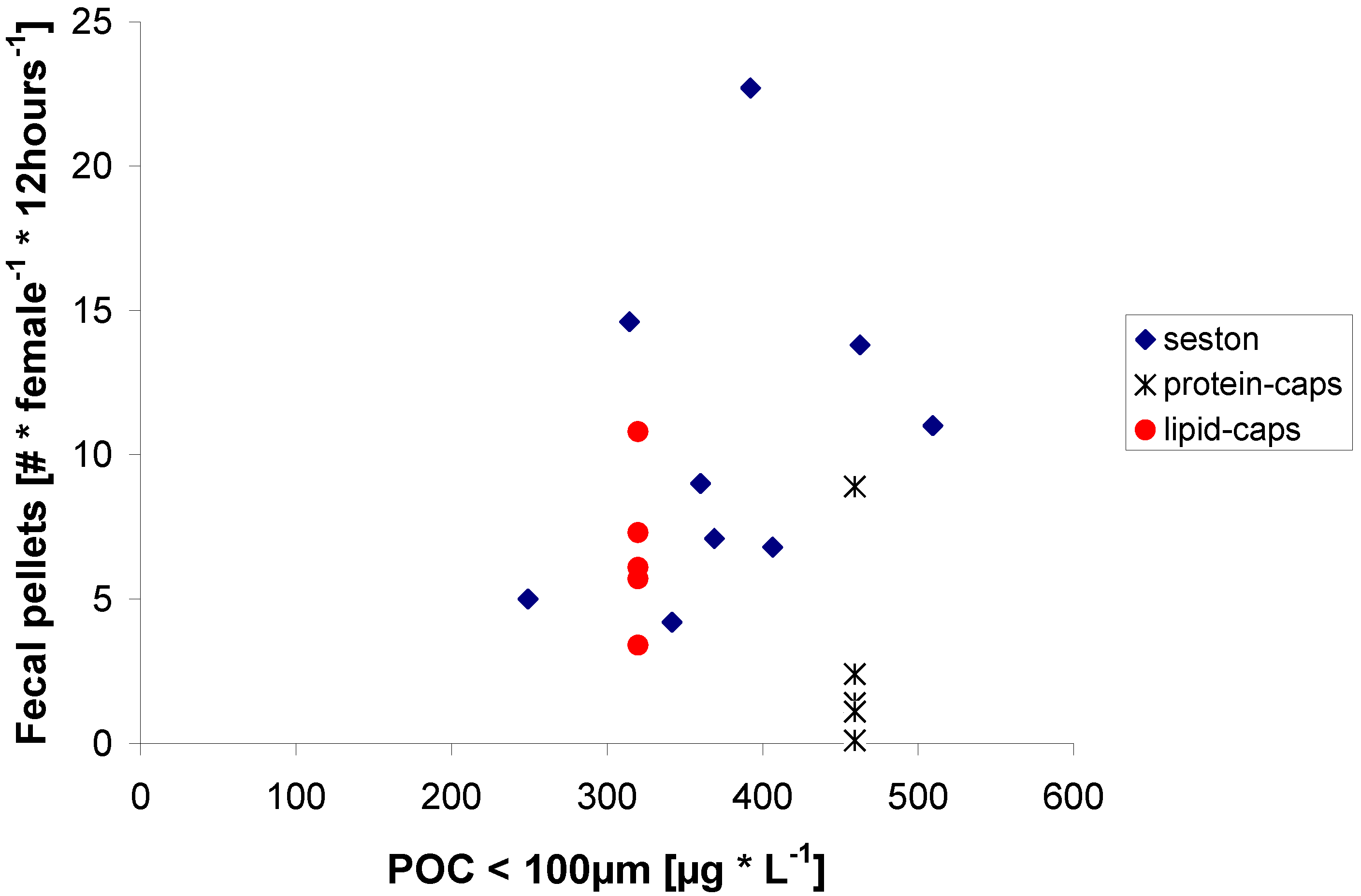{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Size | Respresentative organisms | Cell volume | Cell volume |
|---|---|---|---|
| [μm] | [μm3·mL−1] | % | |
| <4 | Flagellates | 56,651 | 56.8 |
| 4–8 | cryptophyte, thecate dinoflagellates | 3213 | 3.2 |
| 8–18 | Scrippsiella, Prorocentrum Diatoms *, ciliates | 5819 | 5.8 |
| >18 | Protoperidinium, Prorocentrum, Nitzschia | 34,106 | 34.2 |
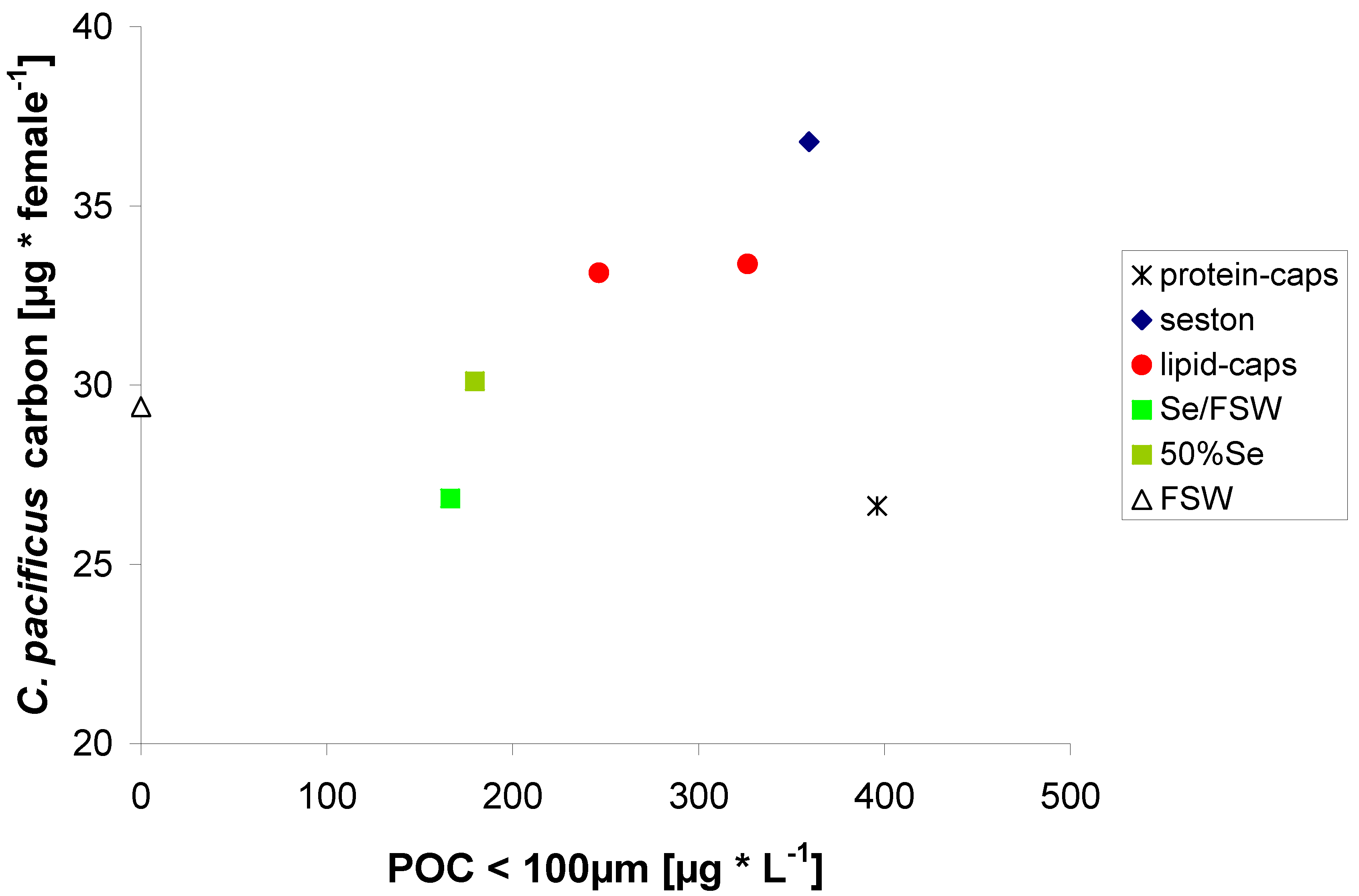
2.3. Egg Production on Protein μ-Caps
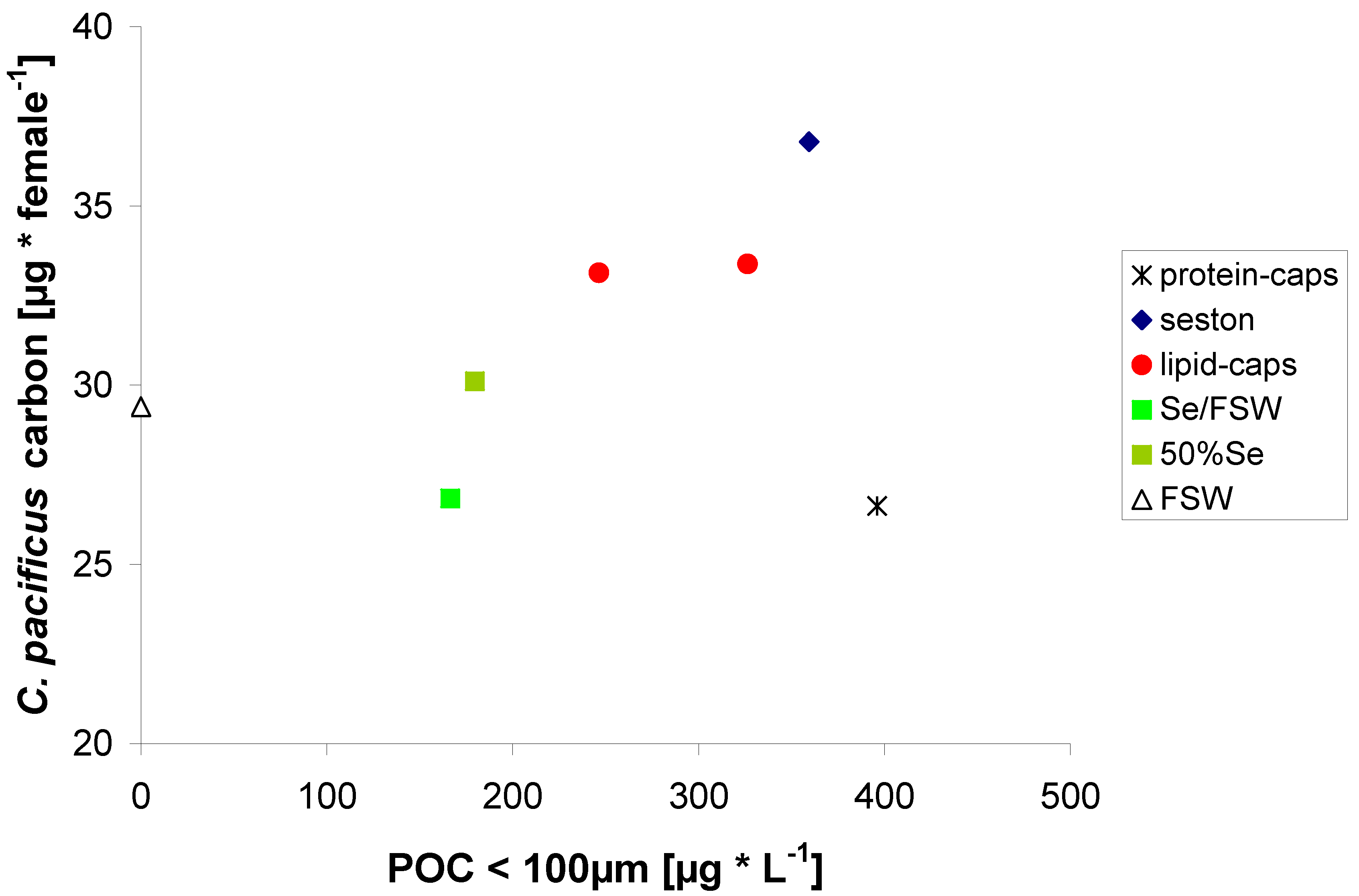
2.4. Egg Production on Lipid μ-Caps Rich in ω3-HUFA
| Fatty acid | % |
|---|---|
| C14:0 | 5.83 |
| C14:1ω? | 0.21 |
| C14:1ω5 | 0.13 |
| C15:0 | 0.47 |
| C16:0 | 13.98 |
| C16:1ω7 | 6.83 |
| C16:1ω5 | 0.51 |
| C16:2ω6 | 0.07 |
| C16:3ω4 | 1.24 |
| C16:4ω1 | 0.82 |
| C17:0 | 0.6 |
| C18:0 | 2.83 |
| C18:1ω12/ω9 | 14.86 |
| C18:1ω7 | 3.59 |
| C18:2ω6 | 2.95 |
| C18:3ω6 | 0.26 |
| C18:3ω3 | 1.74 |
| C18:4ω3 | 5.59 |
| C20:0 | 0.32 |
| C20:1ω9 | n.d. |
| C20:1ω7 | 0.45 |
| C20:2ω6 | 0.17 |
| C20:3ω6 | 0.17 |
| C20:4ω6 | 0.91 |
| C20:5ω3 | 13.36 |
| C22:0 | n.d. |
| C22:1ω9 | 10.71 |
| C22:6ω3 | 10.05 |
| C23:0 | 0.21 |
| C24:0 | 0.05 |
| C24:1ω9 | 0.79 |
| ∑HUFA | 24.7 |
2.5. Zooplankton Nutrition and the Coupling between Primary and Secondary Production
3. Experimental Section
3.1. Animal Collection and Experimental Protocol
3.2. Micro-Capsule Preparation and Their Ingestion
3.3. Egg Production Measurements
3.4. Seston Analyses
4. Conclusions
Acknowledgments
Abbreviations
| DHA | docosahexaenoic acid |
| DW | dry weight |
| E | egg production |
| EPA | eicosapentaenoic acid |
| FA | fatty acid |
| FAME | fatty acid methyl ester |
| FSW | filtered sea water |
| GC-FID | gas chromatography-flame-ionisation detector |
| HUFA | highly unsaturated fatty acid |
| μ-caps | microcapsules |
| POC | particulate organic carbon |
| PUFA | polyunsaturated fatty acid |
Conflict of Interest
References
- Ianora, A.; Miralto, A. Toxigenic effects of diatoms on grazers, phytoplankton and other microbes: A review. Ecotoxicoloy 2010, 19, 493–511. [Google Scholar] [CrossRef]
- Huntley, M.E.; Sykes, P.; Rohan, S.; Marin, V. Chemically mediated rejection of dinoflagellate prey by the copepod Calanus pacificus and Paracalanus parvus: Mechanisns, occurrence, significance. Mar. Ecol. Prog. Ser. 1986, 28, 105–120. [Google Scholar] [CrossRef]
- Koski, M.; Rosenberg, M.; Vitasalo, M.; Tanskanen, S.; Sjolund, U. Is Prymnesium patelliferum toxic for copepods? Grazing, egg production, and egestion of the calanoid copepod Eurytemora affinis in mixtures of “good” and “bad” food. ICES J. Mar. Sci. 1999, 56, 131–139. [Google Scholar] [CrossRef]
- Adolf, J.E.; Bachvaroff, T.R.; Krupatkina, D.N.; Nongaki, H.; Brown, P.J.P.; Lewitus, A.J.; Harvey, H.R.; Place, A.R. Species specificity and potential roles of Karlodinium micrum toxin. Afr. J. Mar. Sci. 2006, 28, 415–419. [Google Scholar] [CrossRef]
- Müller-Navarra, D.C. Food web paradigms: The biochemical view on trophic interactions. Int. Rev. Hydrobiol. 2008, 93, 489–505. [Google Scholar] [CrossRef]
- Guisande, C.; Rivero, I.; Maneiro, I. Comparison among amino acid composition of females, eggs and food to determine the relative importance of food quantity and food quality to copepod reproduction. Mar. Ecol. Prog. Ser. 2000, 202, 135–142. [Google Scholar] [CrossRef]
- Anderson, T.R.; Boersma, M.; Raubenheimer, D. Stoichiometry: Linking elements to biochemicals. Ecology 2004, 85, 1193–1202. [Google Scholar] [CrossRef]
- Jónasdótir, S.H.; Fields, D.; Pantoja, S. Copepod egg production in Long Island Sound, USA, as a function of the chemical composition of seston. Mar. Ecol. Prog. Ser. 1995, 119, 87–98. [Google Scholar] [CrossRef]
- Müller-Navarra, D. Evidence that a highly unsaturated fatty acid limits Daphnia growth in nature. Arch. Hydrobiol. 1995, 132, 297–307. [Google Scholar]
- Müller-Navarra, D.C.; Brett, M.T.; Liston, A.M.; Goldman, C.R. A highly unsaturated fatty acid predicts carbon transfer between primary and secondary producers and consumers. Nature 2000, 403, 74–77. [Google Scholar] [CrossRef]
- Wacker, A.; von Elert, E. Polyunsaturated fatty acids: Evidence for non-substitutable bio-chemical resources in Daphnia galeata. Ecology 2001, 82, 2507–2520. [Google Scholar]
- Hazzard, S.E.; Kleppel, G.S. Egg production of the copepod Acartia tonsa in Florida Bay: Role of fatty acids in the nutritional composition of the food environment. Mar. Ecol. Prog. Ser. 2003, 252, 199–206. [Google Scholar] [CrossRef]
- Arendt, K.E.; Jónasdóttir, S.H.; Hansen, P.J.; Gärtner, S. Effects of dietary fatty acids on the reproductive success of the calanoid copepod Temora longicornis. Mar. Biol. 2005, 146, 513–530. [Google Scholar] [CrossRef]
- Evjerno, J.O.; Tokle, N.; Vadstein, O.; Olsen, Y. Effect of essential dietary fatty acids on egg production and hatching success of the marine copepd Temora longicornis. J. Exp. Mar. Biol. 2008, 365, 31–37. [Google Scholar] [CrossRef]
- Koski, M.; Dutz, J.; Klein-Breteler, W.; Rampen, S.; Noordeloos, A. Seasonal changes in food quantity and quality of the common North Sea copepods Temora longicornis and Pseudocalanus elongatus: A bioassay approach. Mar. Ecol. Prog. Ser. 2010, 399, 141–155. [Google Scholar] [CrossRef]
- Jónasdóttir, S.H.; Visser, A.W.; Jespersen, C. Assessing the role of food quality in the production and hatching of Temora longicornis eggs. Mar. Ecol. Prog. Ser. 2009, 382, 139–150. [Google Scholar] [CrossRef]
- Chen, M.R.; Liu, H.B.; Chen, B.Z. Effects of dietary essential fatty acids on reproduction rates of a subtropical calanoid copepod, Acartia erythraea. Mar. Ecol. Prog. Ser. 2012, 255, 95–110. [Google Scholar] [CrossRef]
- Pond, D.; Harris, R.; Head, R.; Harbour, D. Environmental and nutritional factors determining seasonal variability in the fecundity and egg viability of Calanus helgolandicus in coastal waters off Plymouth, UK. Mar. Ecol. Prog. Ser. 1996, 143, 45–63. [Google Scholar] [CrossRef]
- Jónasdóttir, S.H.; Dutz, J.; Koski, M.; Vidoudez, C.; Jakobsen, H.H.; Pohnert, G.; Nejstgaard, J.G. Extensive cross disciplinary analysis of biological and chemical control of Calanus finmarchicus reproduction during an aldehyde forming diatom bloom in mesocosms. Mar. Biol. 2011, 158, 1943–1963. [Google Scholar] [CrossRef]
- Koski, M.; Yebra, L.; Dutz, J.; Jónasdóttir, S.H.; Vidoudez, C.; Jakobsen, H.H.; Pohnert, G.; Nejstgaard, J.C. The effect of egg versus seston quality on hatching success, naupliar metabolism and survival of Calanus finmarchicus in mesocosms dominated by Phaeocystis and diatoms. Mar. Biol. 2012, 159, 643–660. [Google Scholar] [CrossRef]
- Kleppel, G.S. On the diets of calanoid copepods. Mar. Ecol. Progr. Ser. 1993, 99, 183–195. [Google Scholar] [CrossRef]
- Kleppel, G.S.; Burkart, C.A.; Houchin, L. Nutrition and the regulation of egg production in the calanoid copepod Acartia tonsa. Limnol. Oceanogr. 1998, 43, 1000–1007. [Google Scholar] [CrossRef]
- Langdon, C.J.; Waldock, M.J. The effect of algal and artificial diets on the growth and fatty acid composition of Crassostrea gigas spat. J. Mar. Biol. Assoc. UK 1981, 61, 431–448. [Google Scholar] [CrossRef]
- Kreeger, D.A.; Hawkins, J.S.; Bayne, B.L. Use of dual-labeled microcapsules to discern the physiological fates of assimilated carbohydrate, protein carbon, and protein nitrogen in suspension-feeding organisms. Limnol. Oceanogr. 1996, 41, 208–215. [Google Scholar] [CrossRef]
- Checkley, D.M. Food limitation of egg production by a marine, planktonic copepod in the sea off southern California. Limnol. Oceanogr. 1980, 25, 991–998. [Google Scholar] [CrossRef]
- Durbin, E.G.; Durbin, A.G.; Smayda, T.J.; Verity, P.G. Food limitation of production by adult Acartia tonsa in Narragansett Bay, Rhode Island. Limnol. Oceanogr. 1983, 28, 1199–1213. [Google Scholar] [CrossRef]
- Runge, J.A. Relationship of egg production of Calanus pacificus to seasonal changes in phytoplankton availability in Puget Sound, Washington. Limnol. Oceanogr. 1985, 30, 382–396. [Google Scholar] [CrossRef]
- Ambler, J.W. Effect of food quantity and quality on egg production of Acartia tonsa Dana from East Lagoon, Galveston, Texas. Estuar. Coast. Shelf Sci. 1986, 23, 183–196. [Google Scholar] [CrossRef]
- Peterson, W.T. Rates of egg production by the copepod Calanus marshallae in the laboratory and in the sea off Oregon, USA. Mar. Ecol. Prog. Ser. 1988, 47, 229–237. [Google Scholar] [CrossRef]
- Mullin, M.M. Production of eggs by the copepod Calanus pacificus in the southern California sector of the California current system. CalCoFi Rep. 1991, 32, 65–90. [Google Scholar]
- Kleppel, G.S. Environmental regulation of feeding and egg production by Acartia tonsa off southern California. Mar. Biol. 1992, 112, 57–65. [Google Scholar] [CrossRef]
- Kiørboe, T.; Nielsen, T.G. Regulation of zooplankton biomass and production in a temperature, coastal ecosystem. I. Copepods. Limnol. Oceanogr. 1994, 39, 493–507. [Google Scholar] [CrossRef]
- Bell, M.V.; Dick, J.R.; Anderson, T.R.; Pond, D.W. Application of liposome and stable isotope tracer techniques to study polyunsaturated fatty acid biosynthesis in marine zooplankton. J. Plankton Res. 2007, 29, 417–422. [Google Scholar] [CrossRef]
- Ravet, J.L.; Brett, M.T.; Müller-Navarra, D.C. A test of the role of poly-unsaturated fatty acids in phytoplankton food quality for Daphnia using liposome supplementation. Limnol. Oceanogr. 2003, 48, 1938–1947. [Google Scholar] [CrossRef]
- Bartram, W.C. Experimental development of a model for the feeding of neritic copepods on phytoplankton. J. Plankton Res. 1981, 3, 25–51. [Google Scholar] [CrossRef]
- Frost, B. Effects of size and concentration of food particles on the feeding behavior of the marine planktonic copepod Calanus pacificus. Limnol. Oceanogr. 1972, 17, 805–815. [Google Scholar] [CrossRef]
- Runge, J.A. Egg production of the marine, planktonic copepod, Calanus pacificus Brodsky: Laboratory observations. J. Exp. Mar. Biol. Ecol. 1984, 74, 53–66. [Google Scholar] [CrossRef]
- Mayzaud, P.; Poulet, S.A. The importance of the time factor in the response of zooplankton to varying concentrations of naturally occurring particulate matter. Limnol. Oceanogr. 1978, 23, 1144–1154. [Google Scholar] [CrossRef]
- Kivi, K.; Kuosa, H.; Tanskanen, S. An experimental study on the role of crustacean and microprotozoan grazers in the planktonic food web. Mar. Ecol. Prog. Ser. 1996, 136, 59–68. [Google Scholar] [CrossRef]
- Kleppel, G.S.; Burkart, C.A. Egg production and the nutritional environment of Acartia tonsa: The role of food quality in copepod nutrition. ICES J. Mar. Sci. 1995, 52, 297–304. [Google Scholar] [CrossRef]
- Arzul, G.; Gentiaen, P.; Bodennec, G. Potential Toxicity of Microalgal Polyunsaturated Fatty Acids (PUFAs). In Marine Lipids; Baudimant, G., Guezennec, J.H., Roy, P., Samain, J.-F., Eds.; IFREMER Publisher: Plouzané, France, 1998; pp. 53–62. [Google Scholar]
- Williams, P.M. Fatty acids derived from lipids of marine origin. J. Fish. Res. Board Can. 1965, 22, 1107–1122. [Google Scholar] [CrossRef]
- Jeffries, H.P. Seasonal composition of temperate plankton communities: Fatty acids. Limnol. Oceanogr. 1970, 15, 419–426. [Google Scholar] [CrossRef]
- Kattner, G.; Gercken, G.; Eberlein, K. Development of lipids during a spring plankton bloom in the North Sea. I. Particulate fatty acids. Mar. Chem. 1983, 14, 149–162. [Google Scholar] [CrossRef]
- Hama, T. Production and turnover rates of fatty acids in marine particulate matter through phytoplankton photosynthesis. Mar. Chem. 1991, 33, 213–227. [Google Scholar] [CrossRef]
- Müller-Navarra, D.C. Biochemical versus mineral limitation in Daphnia. Limnol. Oceanogr. 1995, 40, 1209–1214. [Google Scholar]
- Chen, X.; Wakeham, S.G.; Fisher, N.S. Influence of iron on fatty acid and sterol composition of marine phytoplankton and copepod consumers. Limnol. Oceanogr. 2011, 56, 716–724. [Google Scholar] [CrossRef]
- Koski, M.; Klein-Breteler, W.; Shogt, N. Effect of food quality on rate of growth and development of the pelagic copepod Pseudocalanus elongatus (Copepods, Calanoida). Mar. Ecol. Prog. Ser. 1998, 170, 169–187. [Google Scholar] [CrossRef]
- Sverdrup, H.U.; Allen, W.E. Distribution of diatoms in relation to the character of water masses and currents off Southern California in 1938. J. Mar. Res. 1939, 2, 131–144. [Google Scholar] [CrossRef]
- Dortch, Q.; Packard, T.T. Differences in biomass structure between oligotrophic and eutrophic marine ecosystems. Deep Sea Res. 1989, 36, 223–240. [Google Scholar] [CrossRef]
- Brett, M.T.; Müller-Navarra, D.C. The role of highly unsaturated fatty acids in aquatic food web processes. Freshw. Biol. 1997, 38, 483–499. [Google Scholar] [CrossRef]
- Ryther, J.H. Photosynthesis and fish production in the sea. Science 1969, 166, 72–76. [Google Scholar]
- Legendre, L. The significance of microalgal blooms for fisheries and for the export of particulate organic carbon in oceans. J. Plankton Res. 1990, 12, 681–699. [Google Scholar] [CrossRef]
- Marshall, S.M.; Orr, A.P. On the biology of Calanus finmarchicus. VII. Factors affecting egg production. J. Mar. Biol. Assoc. UK 1952, 30, 527–547. [Google Scholar] [CrossRef]
- Tester, P.A.; Turner, J.T. How long does it take copepods to make eggs? J. Exp. Mar. Biol. Ecol. 1990, 141, 169–182. [Google Scholar] [CrossRef]
- Cary, S.C.; Lovette, J.T.; Perl, P.J.; Huntley, M.E. A micro-encapsulation technique for introducing pure compounds in zooplankton diets. Limnol. Oceanogr. 1992, 37, 404–413. [Google Scholar] [CrossRef]
- Langdon, C.J.; DeBovoise, A.E. Effect of microcapsule type on delivery of dietary protein to a marine suspension-feeder, the oyster Crassostrea gigas. Mar. Biol. 1990, 105, 437–443. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Müller-Navarra, D.C.; Huntley, M.E. Bioactive Compounds Offered in Microcapsules to Determine the Nutritional Value of Copepods’ Natural Diet. Mar. Drugs 2013, 11, 2459-2471. https://doi.org/10.3390/md11072459
Müller-Navarra DC, Huntley ME. Bioactive Compounds Offered in Microcapsules to Determine the Nutritional Value of Copepods’ Natural Diet. Marine Drugs. 2013; 11(7):2459-2471. https://doi.org/10.3390/md11072459
Chicago/Turabian StyleMüller-Navarra, Dörthe C., and Mark E. Huntley. 2013. "Bioactive Compounds Offered in Microcapsules to Determine the Nutritional Value of Copepods’ Natural Diet" Marine Drugs 11, no. 7: 2459-2471. https://doi.org/10.3390/md11072459