6-Bromoisatin Found in Muricid Mollusc Extracts Inhibits Colon Cancer Cell Proliferation and Induces Apoptosis, Preventing Early Stage Tumor Formation in a Colorectal Cancer Rodent Model

 ,
,
Abstract
:
1. Introduction
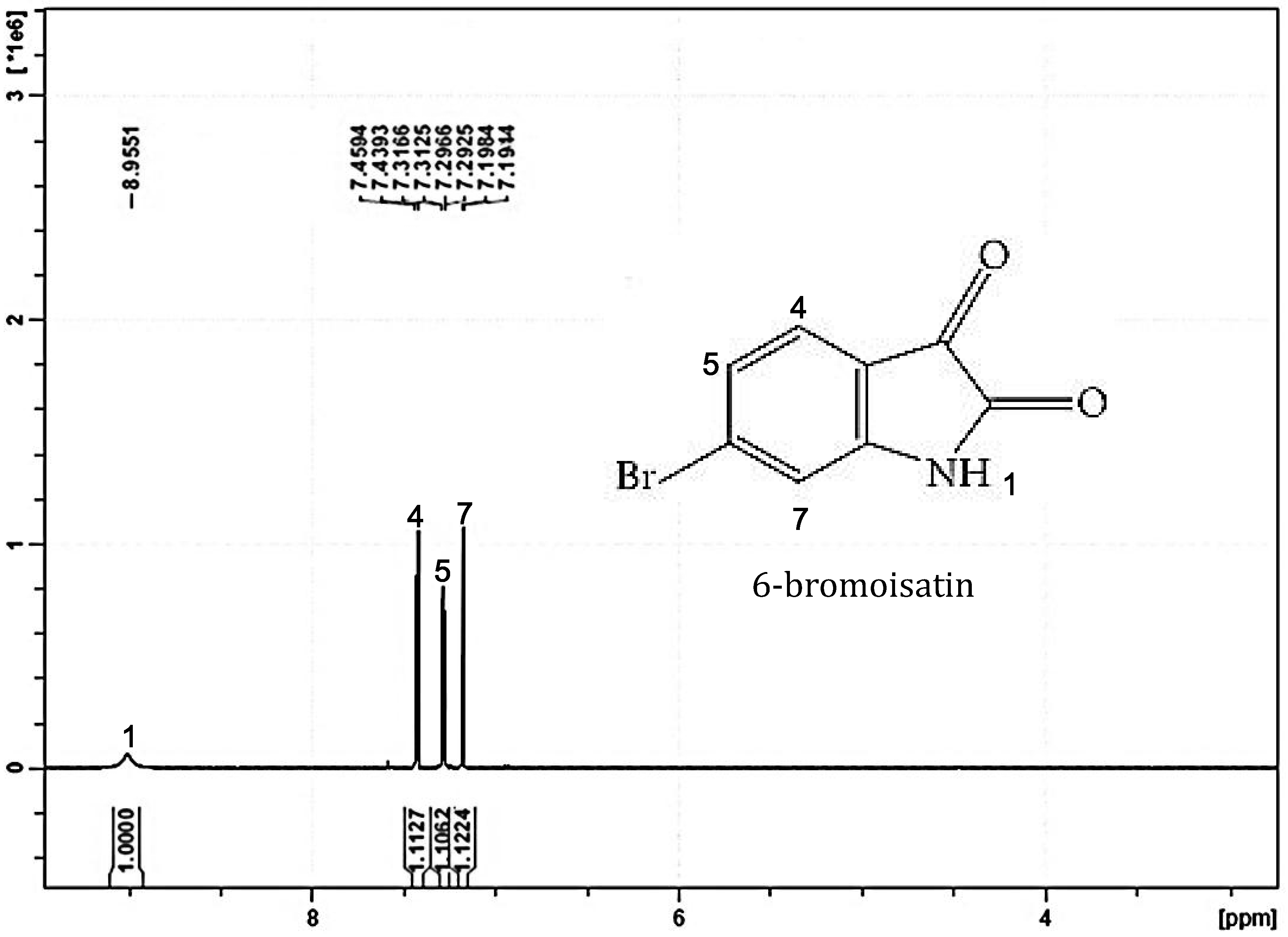
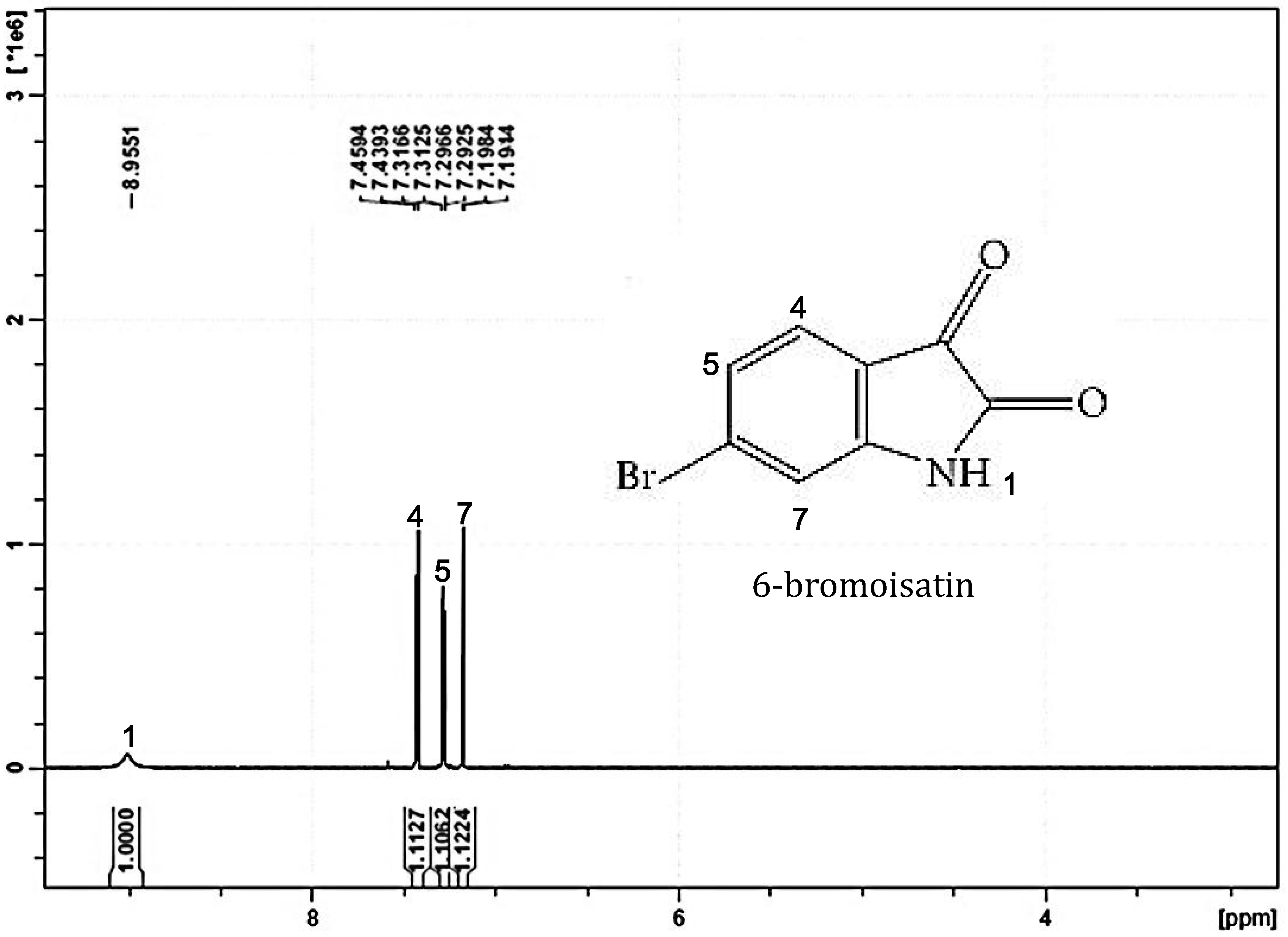
2. Results and Discussion
2.1. Chemical Analysis
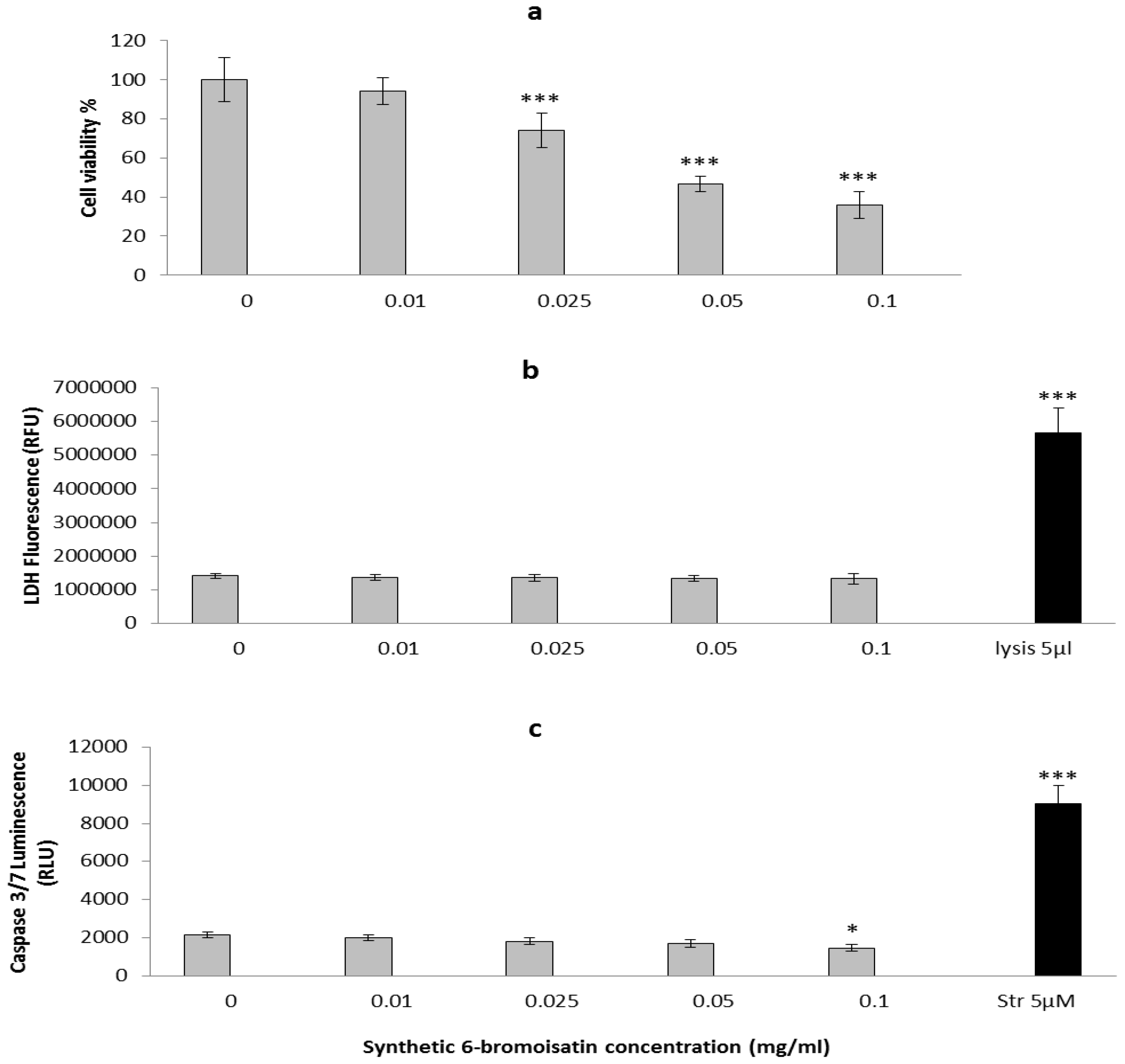
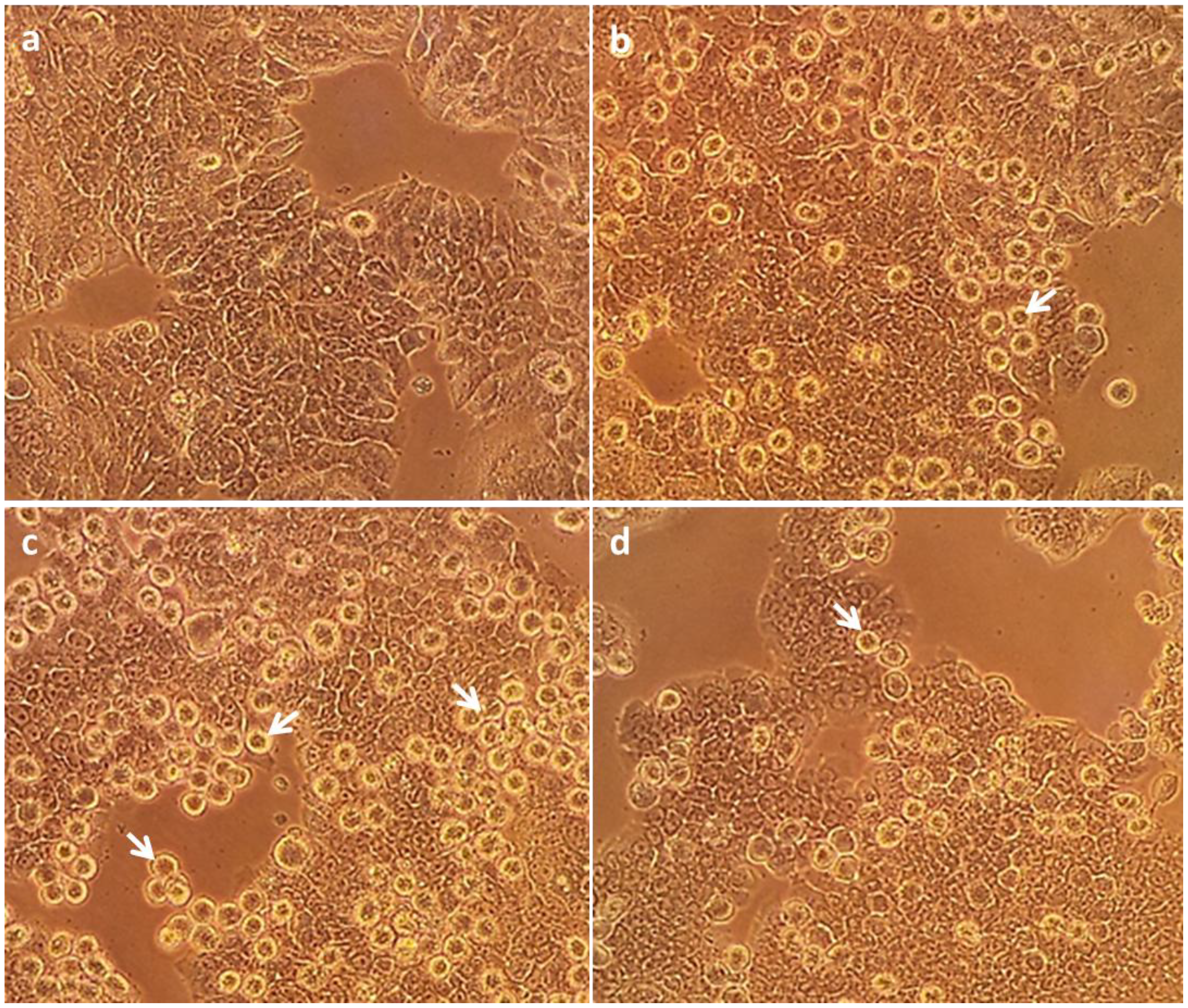
2.2. In Vitro Apoptosis, Necrosis and Cell Viability
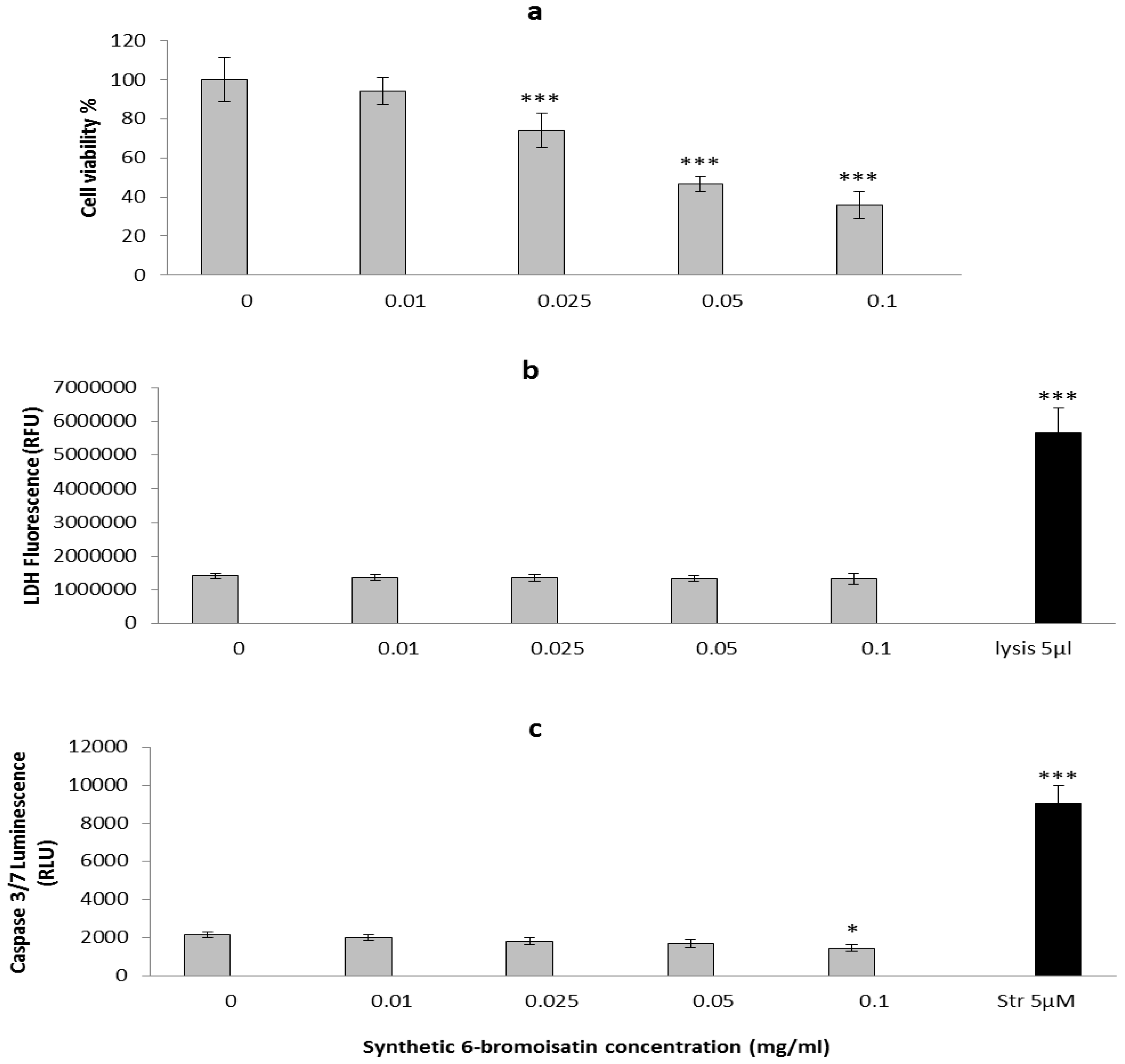
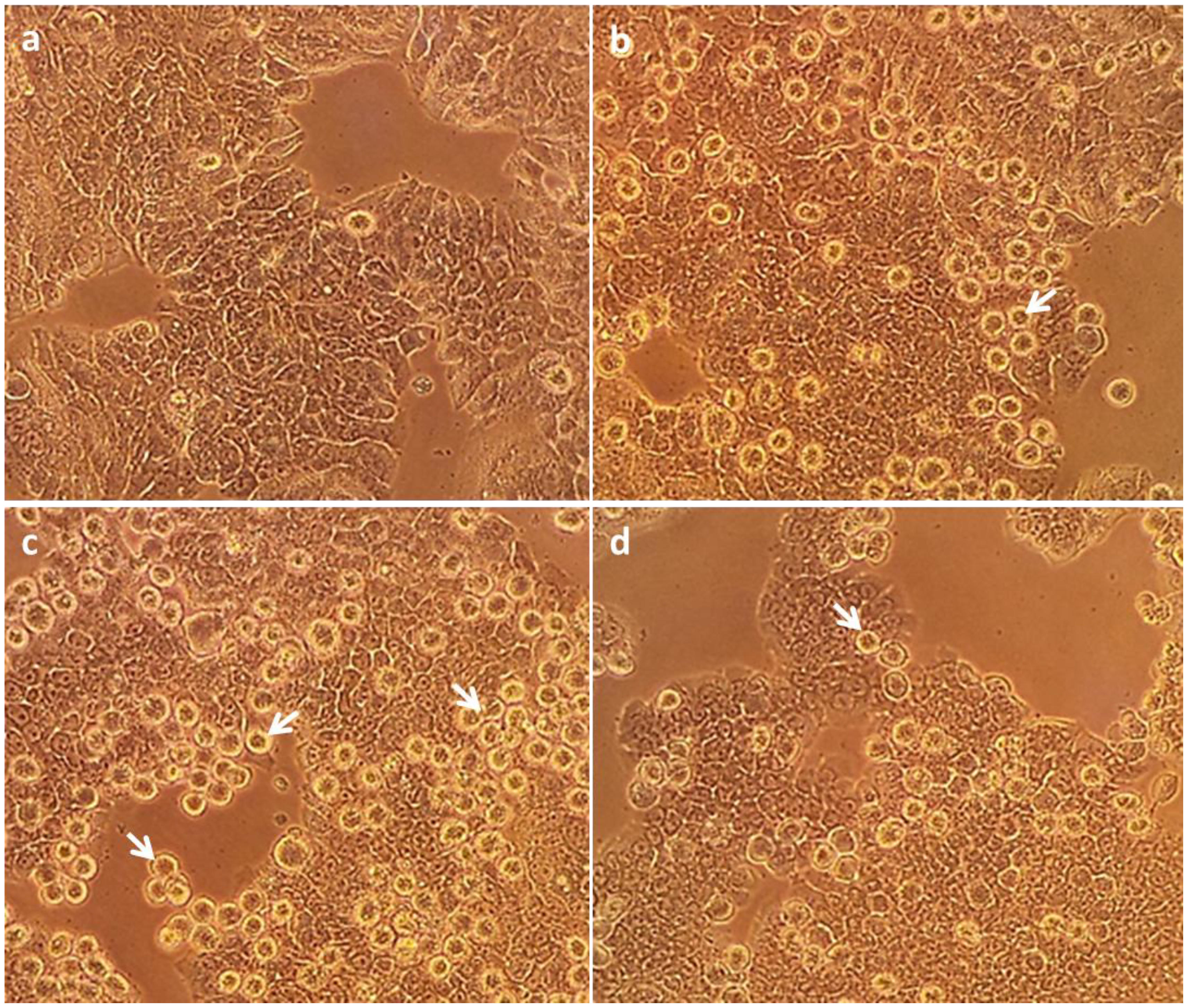
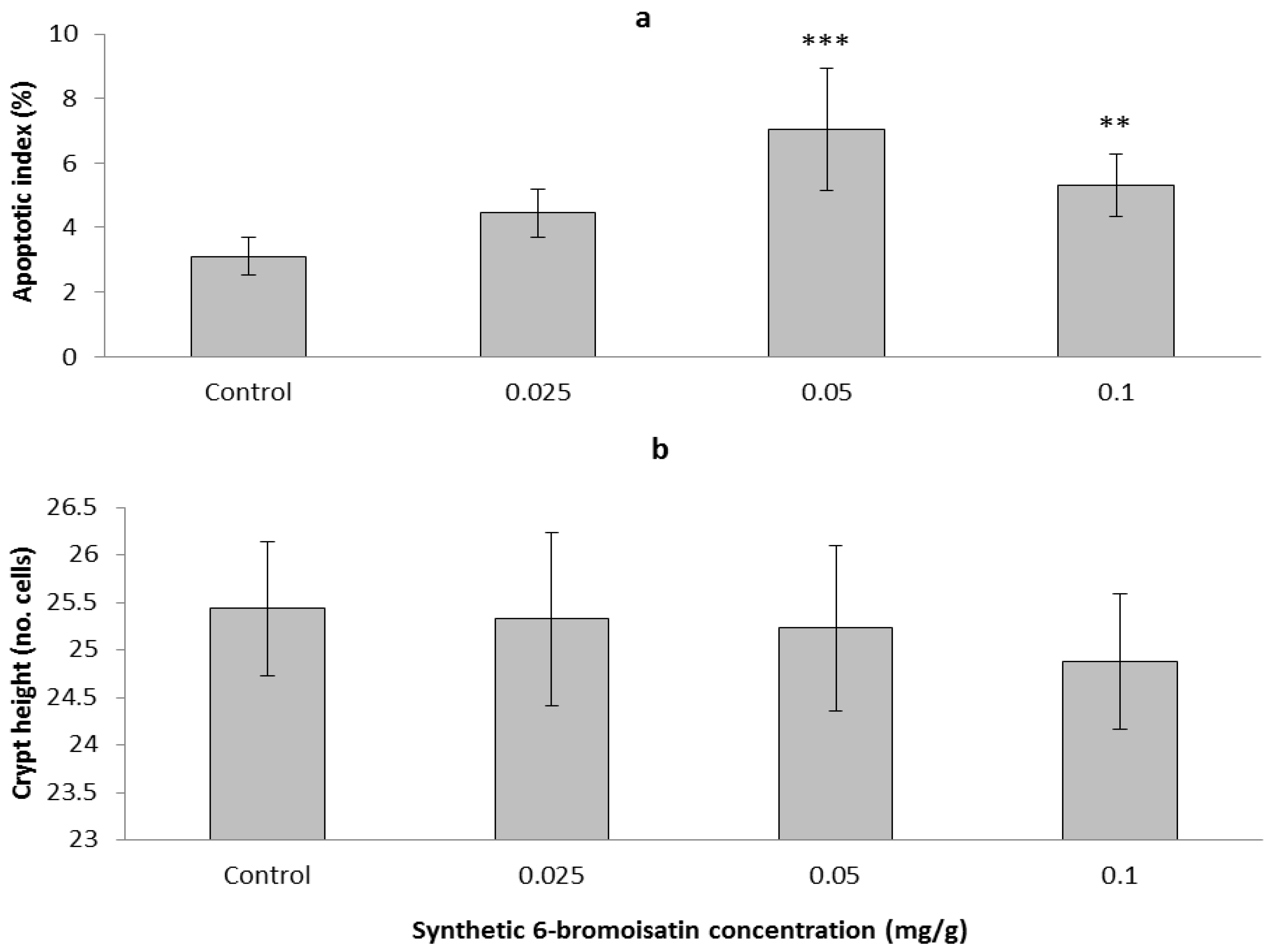
2.3. In Vivo Mouse Model
2.3.1. Mice; General Observations

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Weight (g) | |||||||
|---|---|---|---|---|---|---|---|
| Concentration | Body (Day1) | Body (Day5) | Body (Day10) | Body (Day14) | Total weight Gain | Liver | Liver/Body (%) |
| Control | 22.0 ± 1.6 | 22.6 ± 1.6 | 22.8 ± 1.9 | 23.5 ± 1.9 | 1.4 ± 0.7 | 1.1 ± 0.3 | 4.8 ± 1.2 |
| 0.025 mg/g | 22.2 ± 1.3 | 22.7 ± 1.3 | 23.0 ± 1.3 | 23.6 ± 1.4 | 1.4 ± 0.8 | 1.0 ± 0.1 | 4.4 ± 0.6 |
| 0.05 mg/g | 22.6 ± 1.2 | 23.4 ± 1.2 | 23.4 ± 1.5 | 24.1 ± 1.6 | 1.4 ± 1.0 | 1.2 ± 0.1 | 5.2 ± 0.6 |
| 0.1 mg/g | 22.3 ± 1.3 | 22.6 ± 1.3 | 23.3 ± 1.3 | 23.8 ± 1.6 | 1.4 ± 0.8 | 1.3 ± 0.1 | 5.3 ± 0.4 |
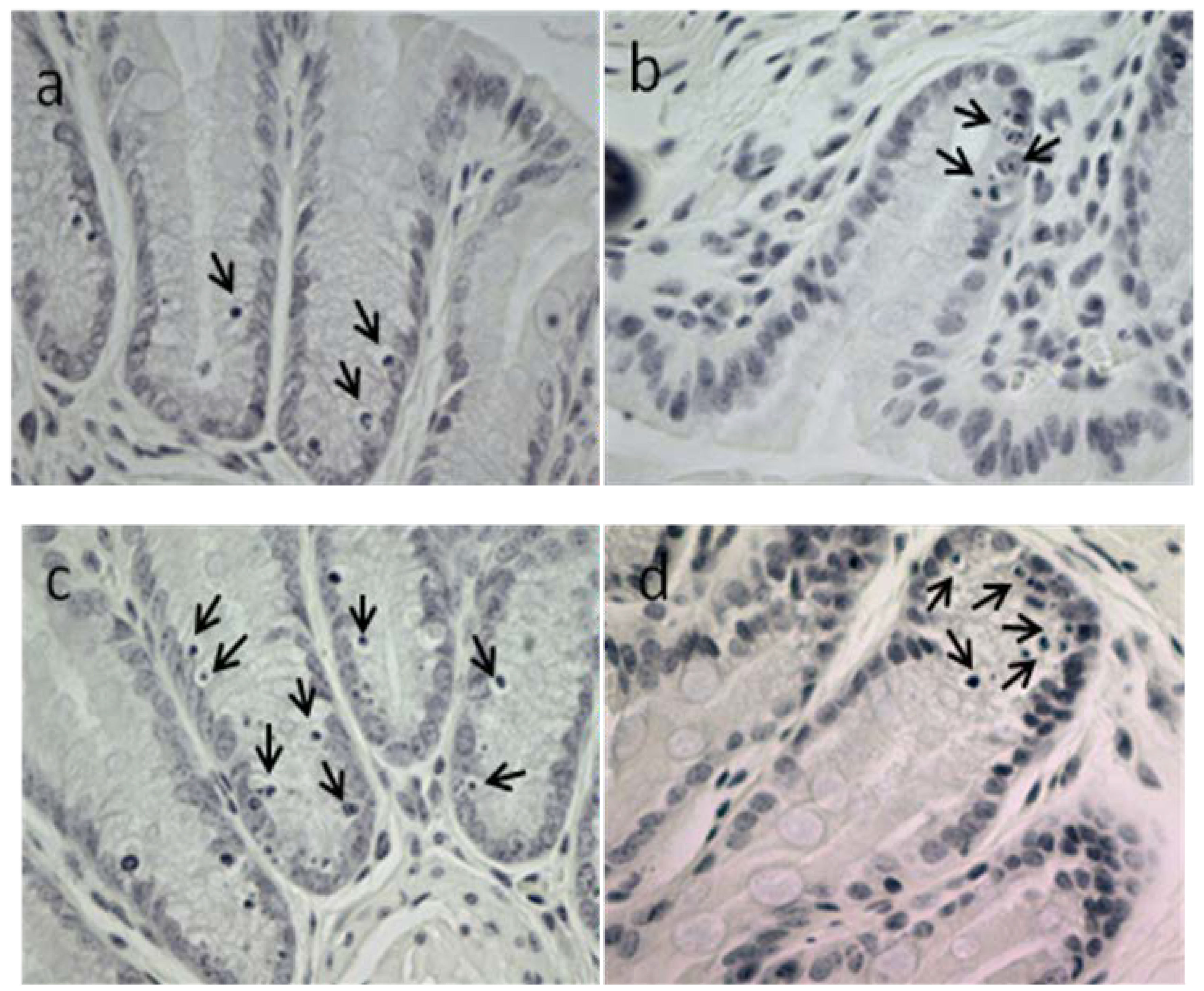
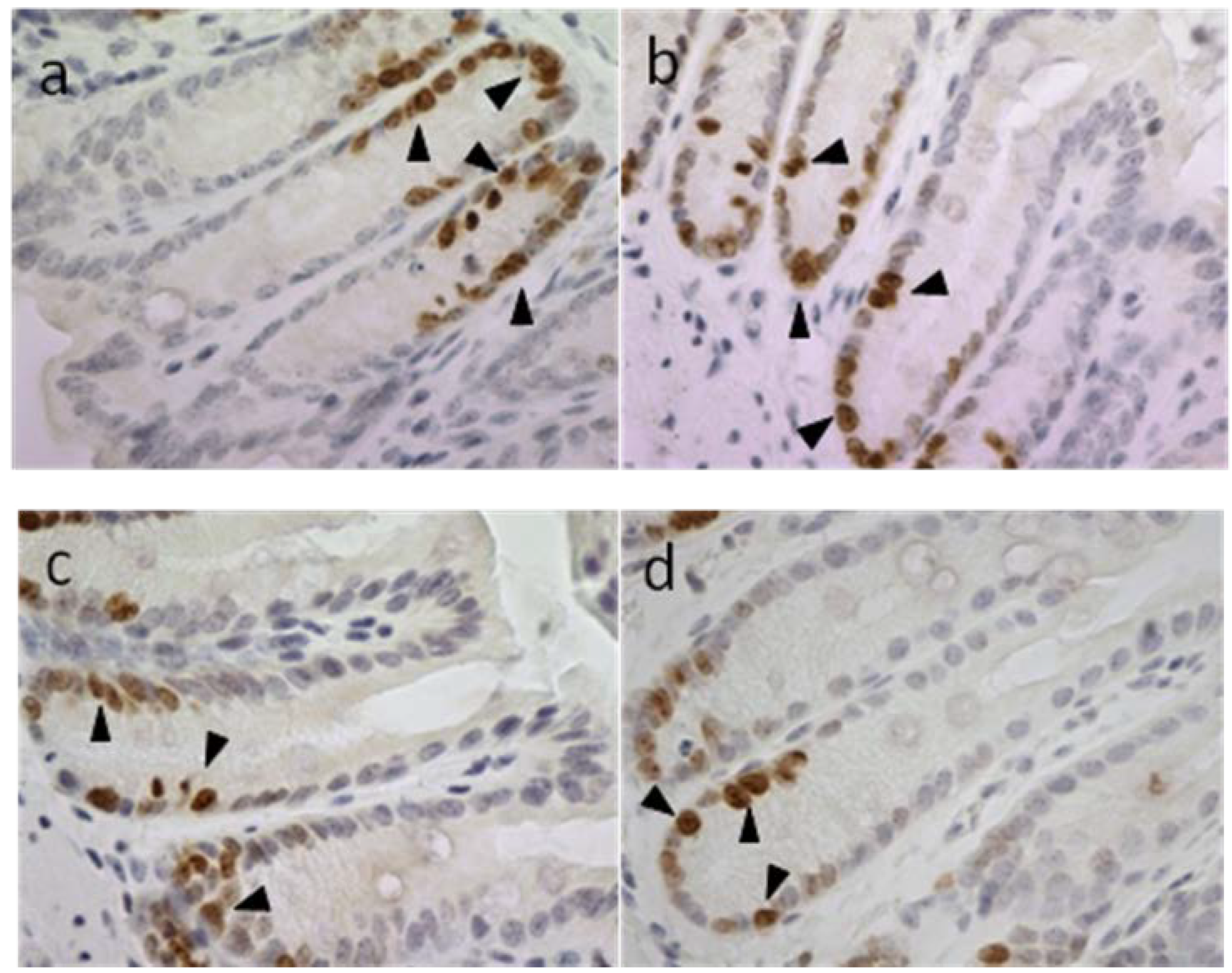
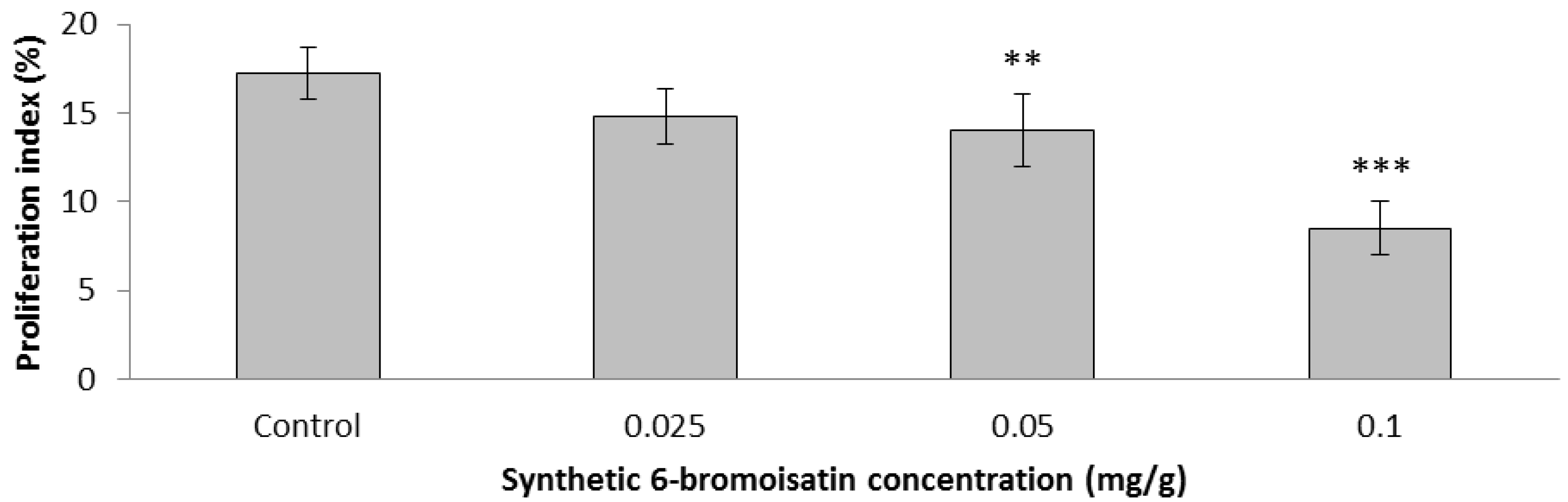
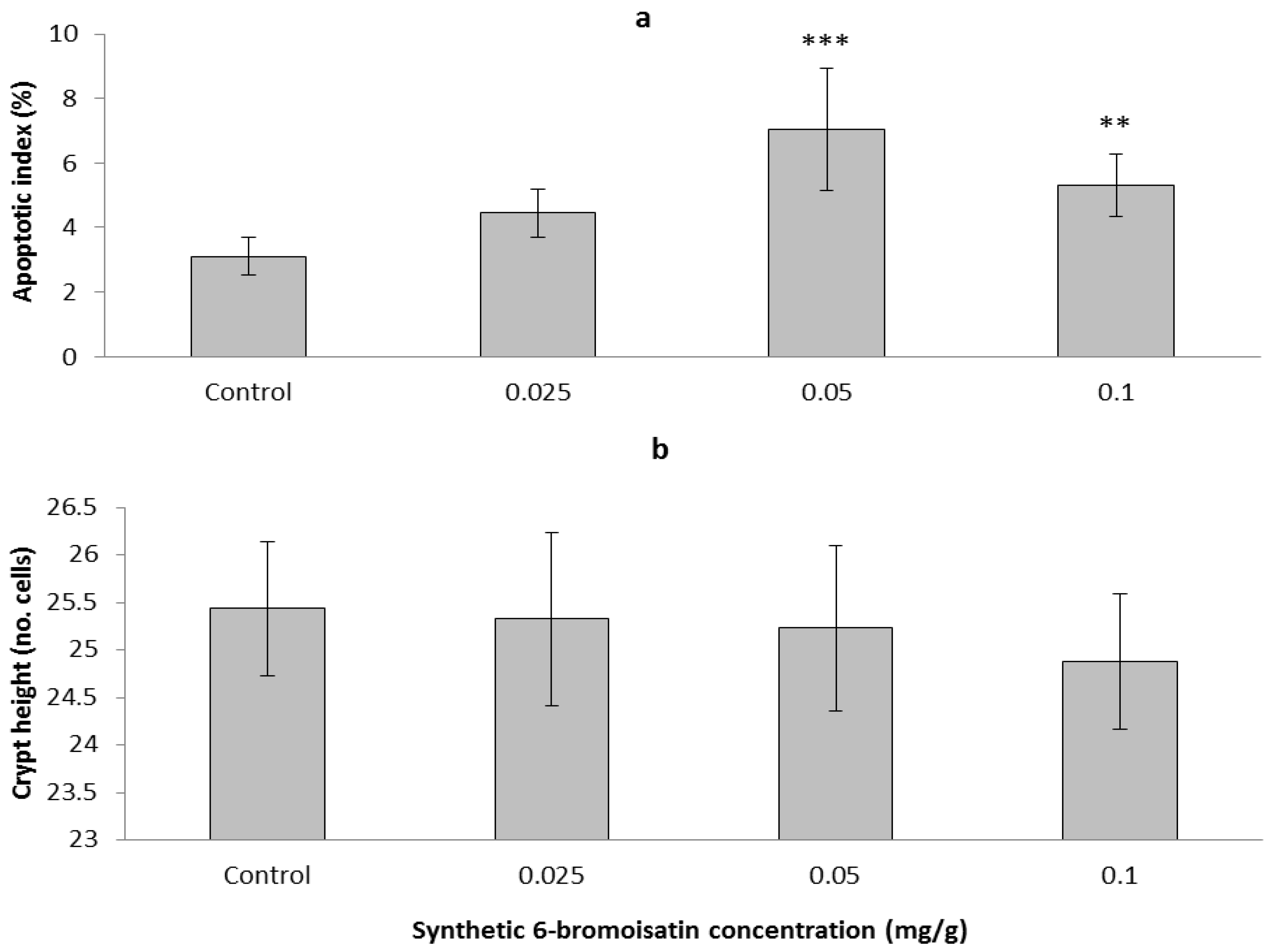
2.3.2. Apoptotic Index, Crypt Height and Cell Proliferation
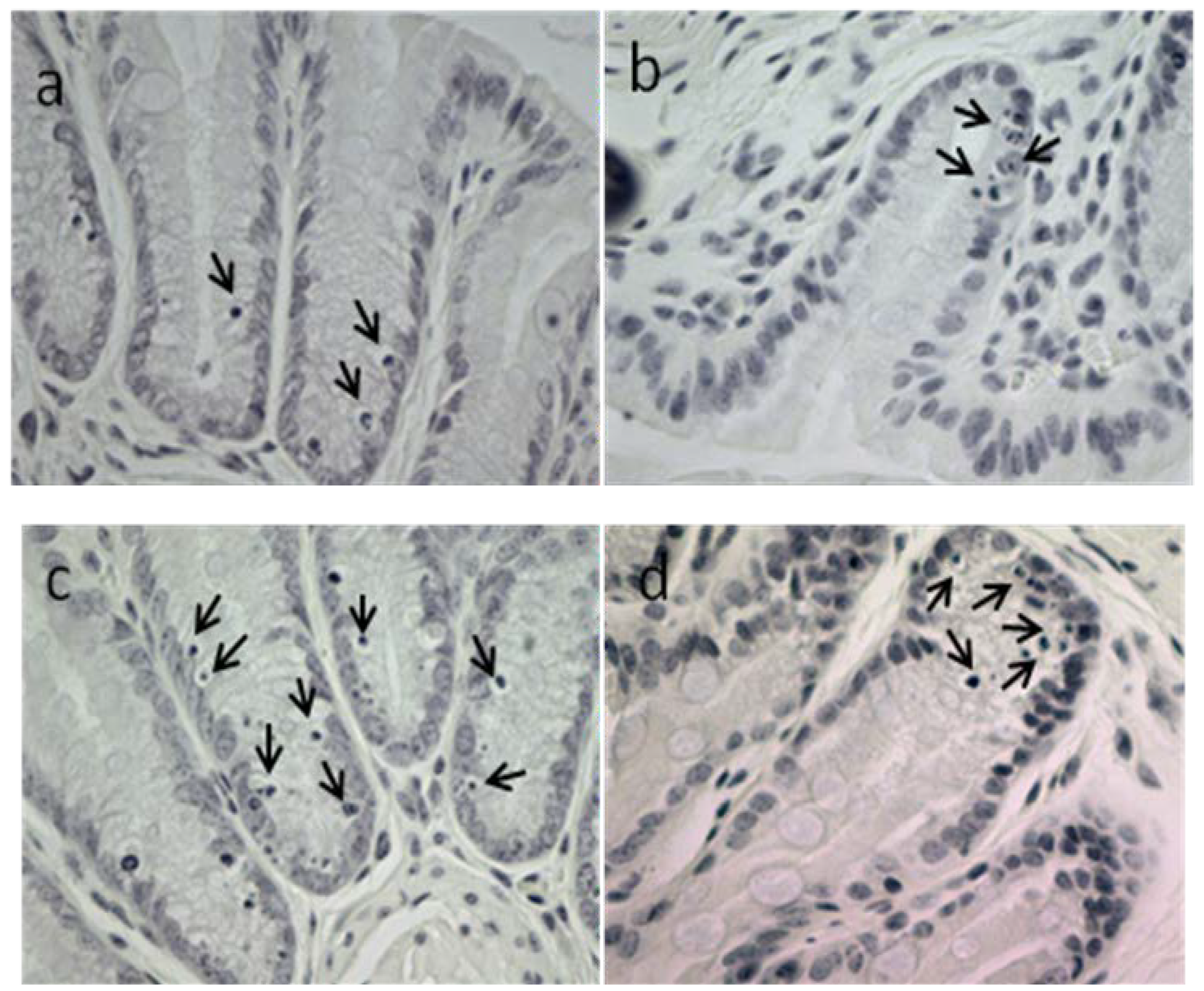
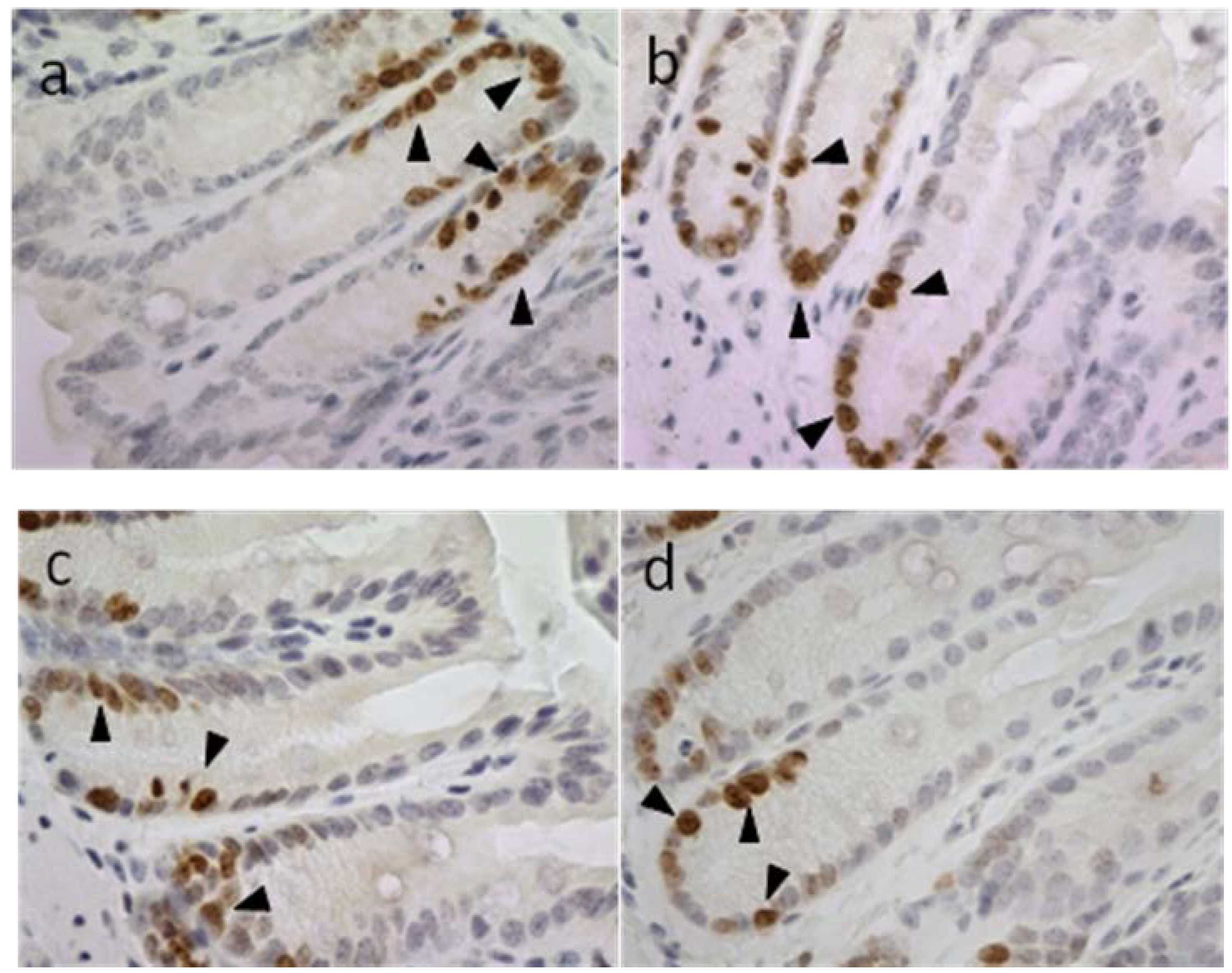

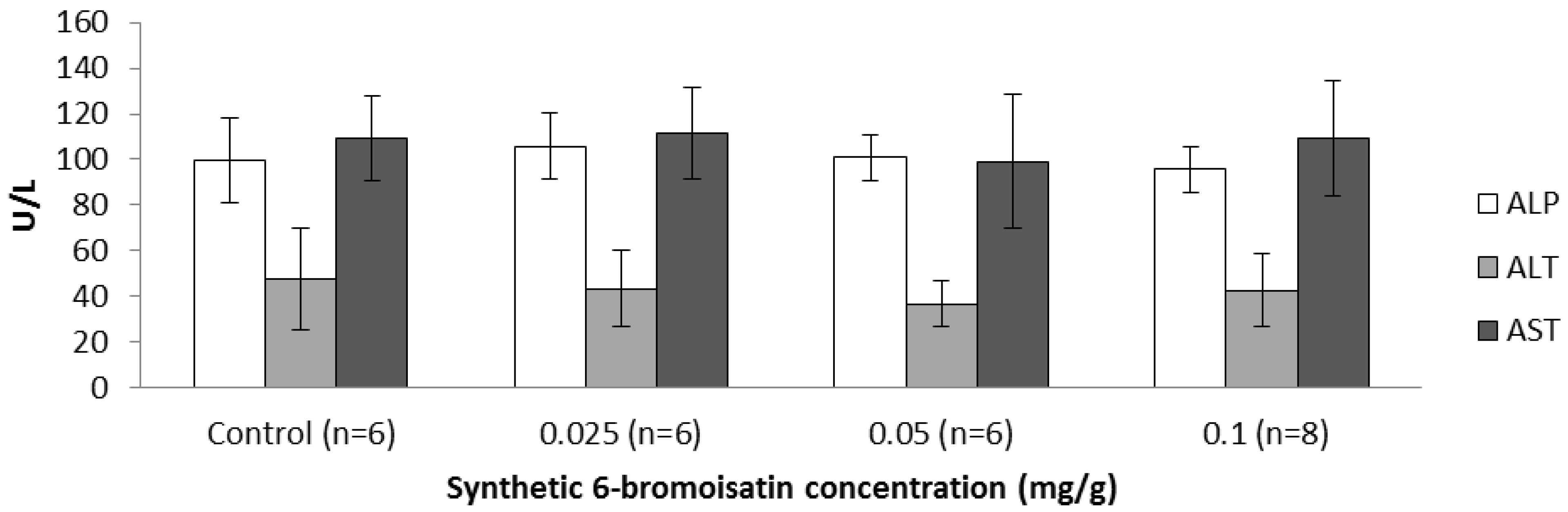
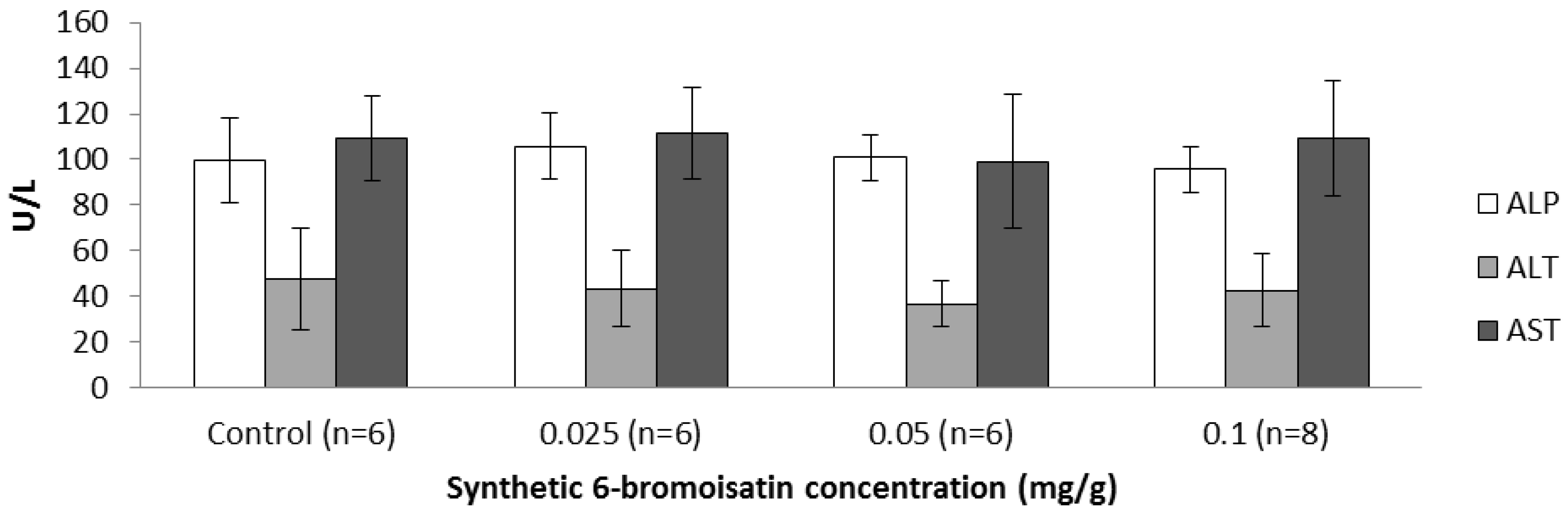
2.3.3. Blood Biochemistry and Hematology
| Oil Control (n = 6) | 6-Bromoisatin (0.025 mg/g, n = 6) | 6-Bromoisatin (0.05 mg/g, n = 6) | 6-Bromoisatin (0.1 mg/g, n = 8) | |
|---|---|---|---|---|
| Biochemistry | ||||
| Sodium (mmol/L) | 146.5 ± 1.2 | 147.7 ± 0.5 | 149.7 ± 1.4 | 149.7 ± 1.2 |
| Potassium (mmol/L) | 5.4 ± 0.3 | 5.1 ± 0.2 # | 4.5 ± 0.4 * | 4.3 ± 0.4 ** |
| NA/K | 27.0 ± 2.2 | 29.1 ± 1.2 # | 33.5 ± 2.7 ** | 34.1 ± 3.6 *** |
| Urea (mmol/L) | 10.6 ± 1.2 | 9.7 ± 1.6 | 9.9 ± 1.4 | 9.6 ± 1.2 |
| Creat. (umol/L) | 14.2 ± 1.2 | 13.3 ± 0.8 | 14.7 ± 0.5 | 14.6 ± 1.2 |
| Calcium (mmol/L) | 2.2 ± 0.03 | 2.2 ± 0.1 | 2.2 ± 0.04 | 2.2 ± 0.1 |
| Protein (g/L) | 45.8 ± 3.0 | 46.7 ± 2.8 | 46.2 ± 1.9 | 46.1 ± 2.2 |
| Albumin (g/L) | 28.2 ± 1.7 | 28.7 ± 1.5 | 27.8 ± 1.5 | 28.2 ± 1.5 |
| Globulin (g/L) | 17.7 ± 1.5 | 18.0 ± 1.4 | 18.3 ± 0.8 | 17.9 ± 1.2 |
| Hematology | ||||
| Red cell count (×1012/L) | 9.0 ± 0.4 | 9.3 ± 0.4 | 9.1 ± 0.4 | 9.4 ± 0.5 |
| Hemoglobin (g/L) | 135.2 ± 2.7 | 139.7 ± 5.1 | 135.0 ± 3.5 | 138.4 ± 5.7 |
| Hct (L/L) | 0.4 ± 0.01 | 0.4 ± 0.01 | 0.4 ± 0.01 | 0.4 ± 0.02 |
| MCV (FL) | 46.0 ± 1.2 | 46.5 ± 0.8 | 46.7 ± 1.2 | 46.4 ± 0.7 |
| MCH (Pg) | 15.2 ± 0.4 | 15.0 ± 0.0 | 15.0 ± 0.0 | 15.0 ± 0.0 |
| MCHC (g/L) | 323.7 ± 4.5 | 322.7 ± 1.5 | 319.0 ± 2.8 | 319.8 ± 3.0 |
| White cell count (×109/L) | 4.8 ± 0.9 | 5.2 ± 1.8 | 5.8 ± 1.5 # | 3.2 ± 1.1 |
| Neutrophils (×109/L) | 1.9 ± 0.3 | 1.6 ± 0.8 | 2.2 ± 1.3 | 0.8 ± 0.7 |
| Lymphocytes (×109/L) | 2.8 ± 0.9 | 3.3 ± 1.1 | 3.4 ± 0.8 | 2.3 ± 0.9 |
| Monocytes (×109/L) | 0.2 ± 0.1 | 0.2 ± 0.3 | 0.2 ± 0.1 | 0.1 ± 0.1 |
3. Experimental Section
3.1. Synthetic 6-Bromoisatin and Chemical Analysis
3.2. In Vitro Experiments Using HT29 Colorectal Cancer Cells
3.3. In Vivo Model for Early Stage Colon Cancer Prevention
3.4. Liver Enzymes, Blood Biochemistry and Hematology
3.5. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Medvedev, A.; Buneeva, O.; Glover, V. Biological targets for isatin and its analogues: Implications for therapy. Biol. Targets Ther. 2007, 1, 151–162. [Google Scholar]
- Vine, K.L.; Matesic, L.; Locke, J.; Ranson, M.; Skropeta, D. Cytotoxic and anticancer activities of isatin and its derivatives: A comprehensive review from 2000–2008. Anticancer Agents Med. Chem. 2009, 9, 397–414. [Google Scholar] [CrossRef]
- Pal, M.; Sharma, N.K.; Priyanka, J.K. Synthetic and biological multiplicity of isatin: A review. J. Adv. Sci. Res. 2011, 2, 35–44. [Google Scholar] [CrossRef]
- Akgul, O.; Tarikogullari, A.H.; Kose, F.A.; Ballar, P.; Pabuccuoglu, V. Synthesis and cytotoxic activity of some 2-(2,3-dioxo-2, 3-dihydro-1H-indol-1-yl) acetamide derivatives. Turk. J. Chem. 2013, 37, 204–212. [Google Scholar]
- Benkendorff, K. The Australian Muricidae Dicathais orbita: A model species for marine natural product research. Mar. Drugs 2013, 11, 1370–1398. [Google Scholar] [CrossRef]
- Cane, A.; Tournaire, M.-C.; Barritault, D.; Crumeyrolle-Arias, M. The endogenous oxindoles 5-hydroxyoxindole and isatin are antiproliferative and proapoptotic. Biochem. Biophys. Res. Commun. 2000, 276, 379–384. [Google Scholar] [CrossRef]
- Igosheva, N.; Lorz, C.; O’Conner, E.; Glover, V.; Mehmet, H. Isatin, an endogenous monoamine oxidase inhibitor, triggers a dose-and time-dependent switch from apoptosis to necrosis in human neuroblastoma cells. Neurochem. Int. 2005, 47, 216–224. [Google Scholar] [CrossRef]
- Vine, K.L.; Locke, J.M.; Ranson, M.; Benkendorff, K.; Pyne, S.G.; Bremner, J.B. In vitro cytotoxicity evaluation of some substituted isatin derivatives. Bioorg. Med. Chem. 2007, 15, 931–938. [Google Scholar] [CrossRef]
- Motzer, R.J.; Michaelson, M.D.; Redman, B.G.; Hudes, G.R.; Wilding, G.; Figlin, R.A.; Ginsberg, M.S.; Kim, S.T.; Baum, C.M.; DePrimo, S.E. Activity of SU11248, a multitargeted inhibitor of vascular endothelial growth factor receptor and platelet-derived growth factor receptor, in patients with metastatic renal cell carcinoma. J. Clin. Oncol. 2006, 24, 16–24. [Google Scholar] [CrossRef]
- Prenen, H.; Cools, J.; Mentens, N.; Folens, C.; Sciot, R.; Schöffski, P.; Van Oosterom, A.; Marynen, P.; Debiec-Rychter, M. Efficacy of the kinase inhibitor SU11248 against gastrointestinal stromal tumor mutants refractory to imatinib mesylate. Clin. Cancer Res. 2006, 12, 2622–2627. [Google Scholar] [CrossRef]
- Kapadia, G.; Shukla, Y.; Basak, S.; Sokoloski, E.; Fales, H. The melosatins—a novel class of alkaloids from Melochia tomentosa. Tetrahedron 1980, 36, 2441–2447. [Google Scholar] [CrossRef]
- Gräfe, U.; Radics, L. Isolation and structure elucidation of 6-(3′-methylbuten-2′-yl) isatin, an unusual metabolite from Streptomyces albus. J. Antibiot. 1986, 39, 162–163. [Google Scholar] [CrossRef]
- Cooksey, C.J. Tyrian purple: 6,6′-dibromoindigo and related compounds. Molecules 2001, 6, 736–769. [Google Scholar] [CrossRef]
- Edwards, V.; Benkendorff, K.; Young, F. Marine compounds selectively induce apoptosis in female reproductive cancer cells but not in primary-derived human reproductive granulosa cells. Mar. Drugs 2012, 10, 64–83. [Google Scholar] [CrossRef]
- Esmaeelian, B.; Benkendorff, K.; Johnston, R.M.; Abbott, C.A. Purified brominated indole derivatives from Dicathais orbita induce apoptosis and cell cycle arrest in colorectal cancer cell lines. Mar. Drugs 2013, 11, 3802–3822. [Google Scholar] [CrossRef]
- Westley, C.B.; McIver, C.M.; Abbott, C.A.; Le Leu, R.K.; Benkendorff, K. Enhanced acute apoptotic response to azoxymethane-induced DNA damage in the rodent colonic epithelium by Tyrian purple precursors: A potential colorectal cancer chemopreventative. Cancer Biol. Ther. 2010, 9, 371–379. [Google Scholar] [CrossRef]
- McLeod, R.; Schmocker, S.; Kennedy, E. Management of primary colon cancer in older adults. Geriat. Aging 2009, 12, 374–381. [Google Scholar]
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef]
- Chan, A.T.; Giovannucci, E.L. Primary prevention of colorectal cancer. Gastroenterology 2010, 138, 2029–2043.e10. [Google Scholar] [CrossRef]
- ACS, American Cancer Society. Colorectal Cancer Facts & Figures 2011–2013; American Cancer Society: Atlanta, GA, USA, 2013. [Google Scholar]
- IARC, International Agency for Research on Cancer (IARC). World Cancer Report 2008; World Health Organization Press: Geneva, Switzerland, 2008; pp. 192–193. [Google Scholar]
- Rajamanickam, S.; Agarwal, R. Natural products and colon cancer: Current status and future prospects. Drug Dev. Res. 2008, 69, 460–471. [Google Scholar] [CrossRef]
- Hong, M.Y.; Chapkin, R.S.; Wild, C.P.; Morris, J.S.; Wang, N.; Carroll, R.J.; Turner, N.D.; Lupton, J.R. Relationship between DNA adduct levels, repair enzyme, and apoptosis as a function of DNA methylation by azoxymethane. Cell Growth Differ. 1999, 10, 749–758. [Google Scholar]
- Reddy, B.S.; Hirose, Y.; Cohen, L.A.; Simi, B.; Cooma, I.; Rao, C.V. Preventive potential of wheat bran fractions against experimental colon carcinogenesis: implications for human colon cancer prevention. Cancer Res. 2000, 60, 4792–4797. [Google Scholar]
- Hu, Y.; Martin, J.; Le Leu, R.; Young, G. The colonic response to genotoxic carcinogens in the rat: regulation by dietary fibre. Carcinogenesis 2002, 23, 1131–1137. [Google Scholar] [CrossRef]
- Le Leu, R.; Hu, Y.; Young, G. Effects of resistant starch and nonstarch polysaccharides on colonic luminal environment and genotoxin-induced apoptosis in the rat. Carcinogenesis 2002, 23, 713–719. [Google Scholar] [CrossRef]
- Le Leu, R.K.; Brown, I.L.; Hu, Y.; Young, G.P. Effect of resistant starch on genotoxin-induced apoptosis, colonic epithelium, and lumenal contents in rats. Carcinogenesis 2003, 24, 1347–1352. [Google Scholar] [CrossRef]
- Vinson, J.; Bose, P. Comparative bioavailability of synthetic and natural Vitamin C in guinea pigs. Nutr. Rep. Int. 1983, 27, 1–5. [Google Scholar]
- Lodge, J.K. Vitamin E bioavailability in humans. J. Plant Physiol. 2005, 162, 790–796. [Google Scholar] [CrossRef]
- Constantinou, C.; Papas, K.; Constantinou, A. Caspase-independent pathways of programmed cell death: The unraveling of new targets of cancer therapy? Curr. Cancer Drug Targets 2009, 9, 717–728. [Google Scholar] [CrossRef]
- Zhuang, S.; Schnellmann, R.G. A death-promoting role for extracellular signal-regulated kinase. J. Pharmacol. Exp. Ther. 2006, 319, 991–997. [Google Scholar] [CrossRef]
- Wang, X.; Martindale, J.L.; Holbrook, N.J. Requirement for ERK activation in cisplatin-induced apoptosis. J. Biol. Chem. 2000, 275, 39435–39443. [Google Scholar] [CrossRef]
- Kim, Y.K.; Kim, H.J.; Kwon, C.H.; Kim, J.H.; Woo, J.S.; Jung, J.S.; Kim, J.M. Role of ERK activation in cisplatin-induced apoptosis in OK renal epithelial cells. J. Appl. Toxicol. 2005, 25, 374–382. [Google Scholar] [CrossRef]
- Jo, S.-K.; Cho, W.Y.; Sung, S.A.; Kim, H.K.; Won, N.H. MEK inhibitor, U0126, attenuates cisplatin-induced renal injury by decreasing inflammation and apoptosis. Kidney Int. 2005, 67, 458–466. [Google Scholar] [CrossRef]
- Sinha, D.; Bannergee, S.; Schwartz, J.H.; Lieberthal, W.; Levine, J.S. Inhibition of ligand-independent ERK1/2 activity in kidney proximal tubular cells deprived of soluble survival factors up-regulates Akt and prevents apoptosis. J. Biol. Chem. 2004, 279, 10962–10972. [Google Scholar] [CrossRef]
- Hu, Y.; McIntosh, G.H.; Le Leu, R.K.; Woodman, R.; Young, G.P. Suppression of colorectal oncogenesis by selenium-enriched milk proteins: apoptosis and K-ras mutations. Cancer Res. 2008, 68, 4936–4944. [Google Scholar] [CrossRef]
- Le Leu, R.K.; Hu, Y.; Brown, I.L.; Woodman, R.J.; Young, G.P. Synbiotic intervention of Bifidobacterium lactis and resistant starch protects against colorectal cancer development in rats. Carcinogenesis 2010, 31, 246–251. [Google Scholar] [CrossRef]
- Saini, K.S.; Thompson, C.; Winterford, C.M.; Walker, N.I.; Cameron, D.P. Streptozotocin at low doses induces apoptosis and at high doses causes necrosis in a murine pancreatic β cell line, INS-1. IUBMB Life 1996, 39, 1229–1236. [Google Scholar] [CrossRef]
- Hewawasam, R.; Jayatilaka, K.; Pathirana, C.; Mudduwa, L. Hepatoprotective effect of Epaltes divaricata extract on carbon tetrachloride induced hepatotoxicity in mice. Indian J. Med. Res. 2004, 120, 30–34. [Google Scholar]
- Westley, C.B.; Benkendorff, K.; McIver, C.M.; Le Leu, R.K.; Abbott, C.A. Gastrointestinal and hepatotoxicity assessment of an anticancer extract from muricid molluscs. Evid.-Based Comp. Alt. Med. 2013, 2013, 1–12. [Google Scholar]
- Lindeman, R.D. Hypokalemia: causes, consequences and correction. Am. J. Med. Sci. 1976, 272, 5–17. [Google Scholar] [CrossRef]
- Weiner, I.D.; Wingo, C.S. Hypokalemia—consequences, causes, and correction. J. Am. Soc. Nephrol. 1997, 8, 1179–1188. [Google Scholar]
- Gennari, F.J. Disorders of potassium homeostasis: Hypokalemia and hyperkalemia. Crit. Care Clin. 2002, 18, 273–288. [Google Scholar] [CrossRef]
- Nataraj, K.S.; Rao, J.V.; Jayaveera, K.N. Diuretic activity of some novel isatin derivatives. J. Pharm. Res. 2010, 3, 863. [Google Scholar]
- Matheus, M.E.; de Almeida Violante, F.; Garden, S.J.; Pinto, A.C.; Dias Fernandes, P. Isatins inhibit cyclooxygenase-2 and inducible nitric oxide synthase in a mouse macrophage cell line. Eur. J. Pharm. 2007, 556, 200–206. [Google Scholar] [CrossRef]
- Kim, J.-K.; Park, G.-M. Indirubin-3-monoxime exhibits anti-inflammatory properties by down-regulating NF-κB and JNK signaling pathways in lipopolysaccharide-treated RAW264.7 cells. Inflamm. Res. 2012, 61, 319–325. [Google Scholar] [CrossRef]
- Jung, H.-L.; Nam, K.N.; Son, M.S.; Kang, H.; Hong, J.W.; Kim, J.W.; Lee, E.H. Indirubin-3′-oxime inhibits inflammatory activation of rat brain microglia. Neurosci. Lett. 2011, 487, 139–143. [Google Scholar] [CrossRef]
- Hu, Y.; Le Leu, R.K.; Young, G.P. Sulindac corrects defective apoptosis and suppresses azoxymethane-induced colonic oncogenesis in p53 knockout mice. Int. J. Cancer 2005, 116, 870–875. [Google Scholar] [CrossRef]
- Potten, C.S.; Li, Y.; O’Connor, P.J.; Winton, D. A possible explanation for the differential cancer incidence in the intestine, based on distribution of the cytotoxic effects of carcinogens in the murine large bowel. Carcinogenesis 1992, 13, 2305–2312. [Google Scholar] [CrossRef]
- Cordes, C.; Munzel, A.N.N.K.; Rudolph, P.; Hoffmann, M.; Leuschner, I.; Gottschlich, S. Immunohistochemical staining of Ki-67 using the monoclonal antibody Ki-S11 is a prognostic indicator for laryngeal squamous cell carcinoma. Anticancer Res. 2009, 29, 1459–1465. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Esmaeelian, B.; Abbott, C.A.; Le Leu, R.K.; Benkendorff, K. 6-Bromoisatin Found in Muricid Mollusc Extracts Inhibits Colon Cancer Cell Proliferation and Induces Apoptosis, Preventing Early Stage Tumor Formation in a Colorectal Cancer Rodent Model. Mar. Drugs 2014, 12, 17-35. https://doi.org/10.3390/md12010017
Esmaeelian B, Abbott CA, Le Leu RK, Benkendorff K. 6-Bromoisatin Found in Muricid Mollusc Extracts Inhibits Colon Cancer Cell Proliferation and Induces Apoptosis, Preventing Early Stage Tumor Formation in a Colorectal Cancer Rodent Model. Marine Drugs. 2014; 12(1):17-35. https://doi.org/10.3390/md12010017
Chicago/Turabian StyleEsmaeelian, Babak, Catherine A. Abbott, Richard K. Le Leu, and Kirsten Benkendorff. 2014. "6-Bromoisatin Found in Muricid Mollusc Extracts Inhibits Colon Cancer Cell Proliferation and Induces Apoptosis, Preventing Early Stage Tumor Formation in a Colorectal Cancer Rodent Model" Marine Drugs 12, no. 1: 17-35. https://doi.org/10.3390/md12010017