Palmitic Acid and Ergosta-7,22-dien-3-ol Contribute to the Apoptotic Effect and Cell Cycle Arrest of an Extract from Marthasterias glacialis L. in Neuroblastoma Cells

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
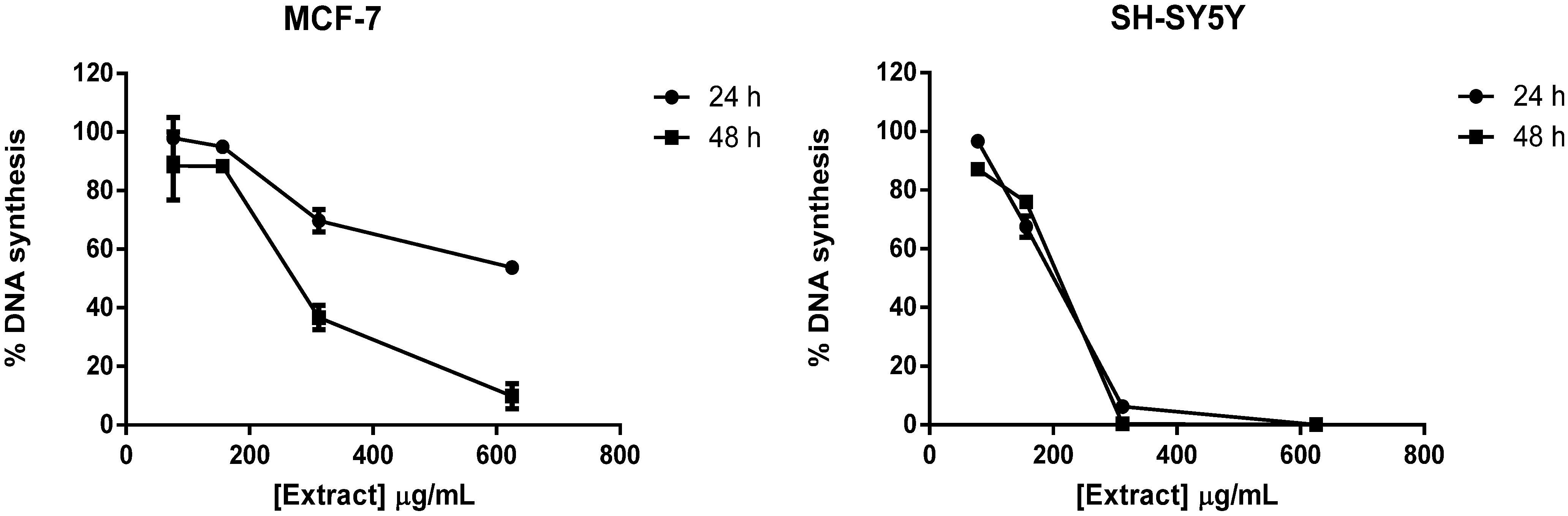
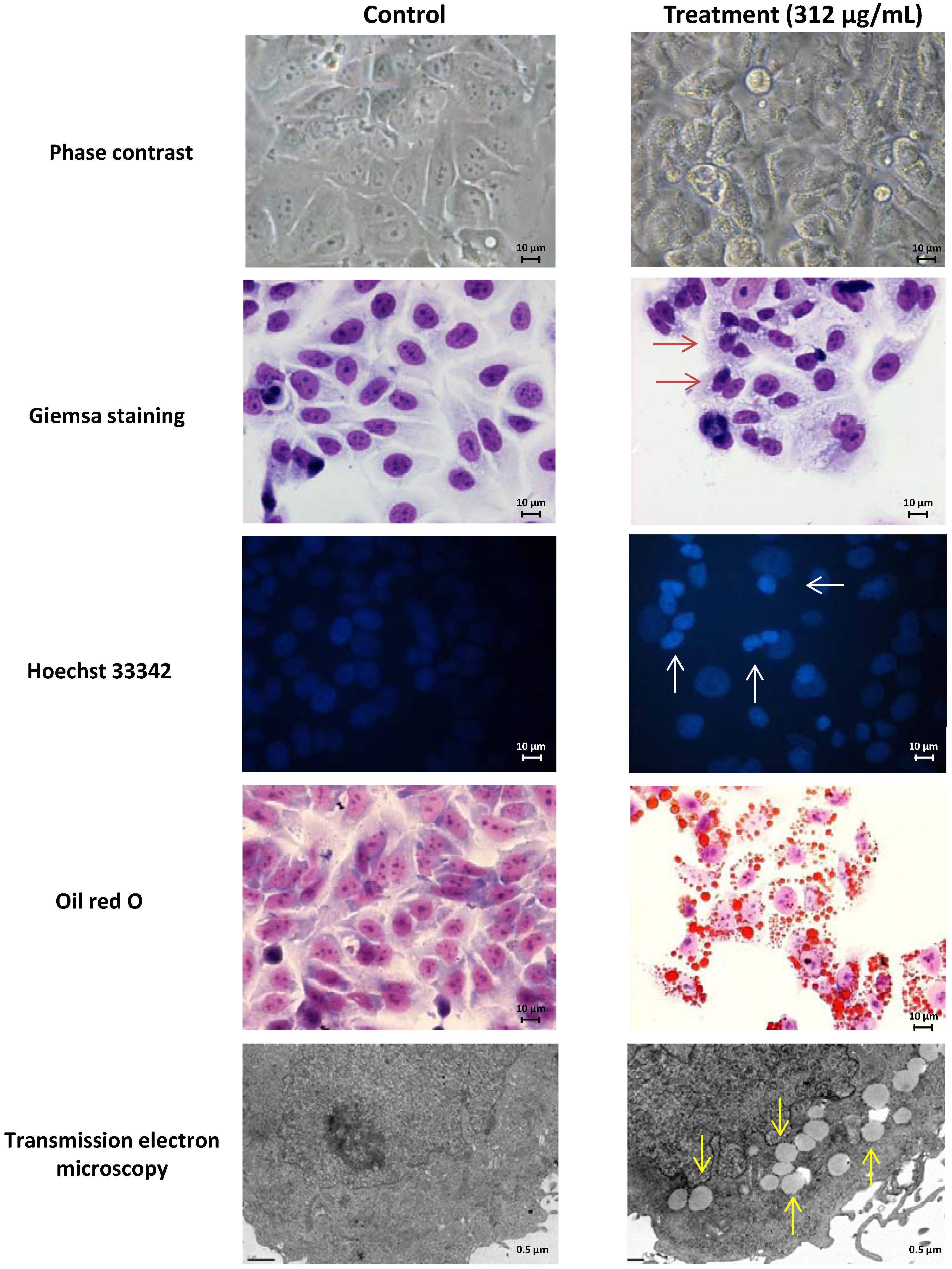
2.1. Screening of M. glacialis Lipophilic Extract Effect on Cell Morphology and DNA Synthesis
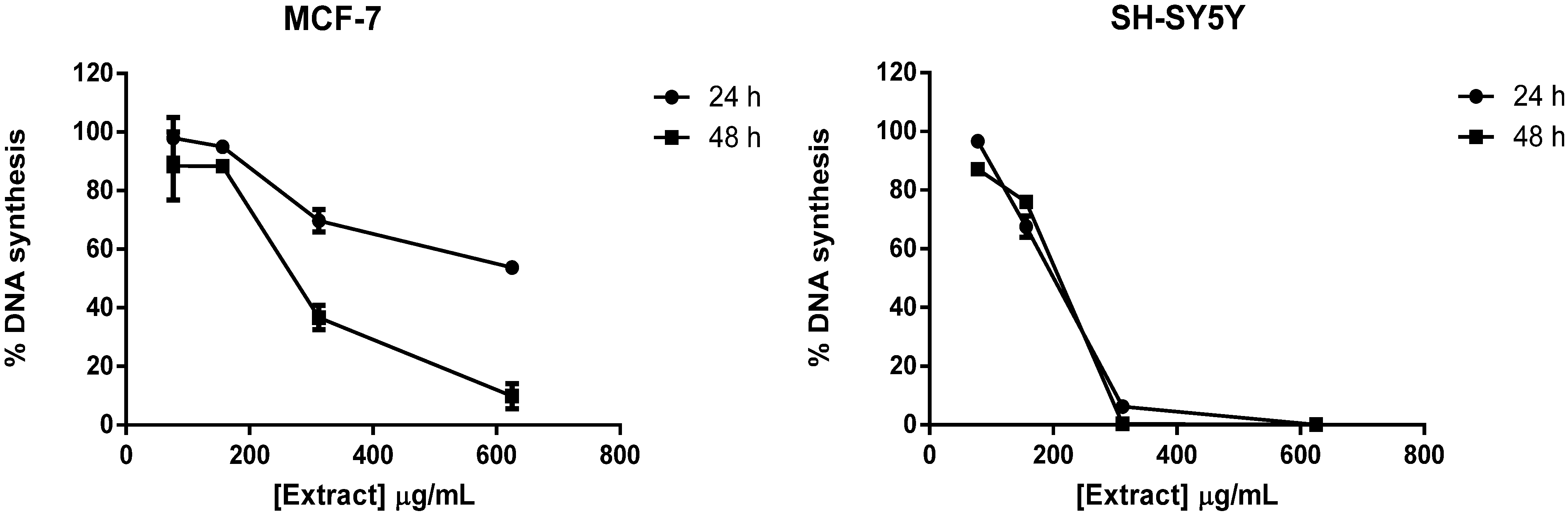
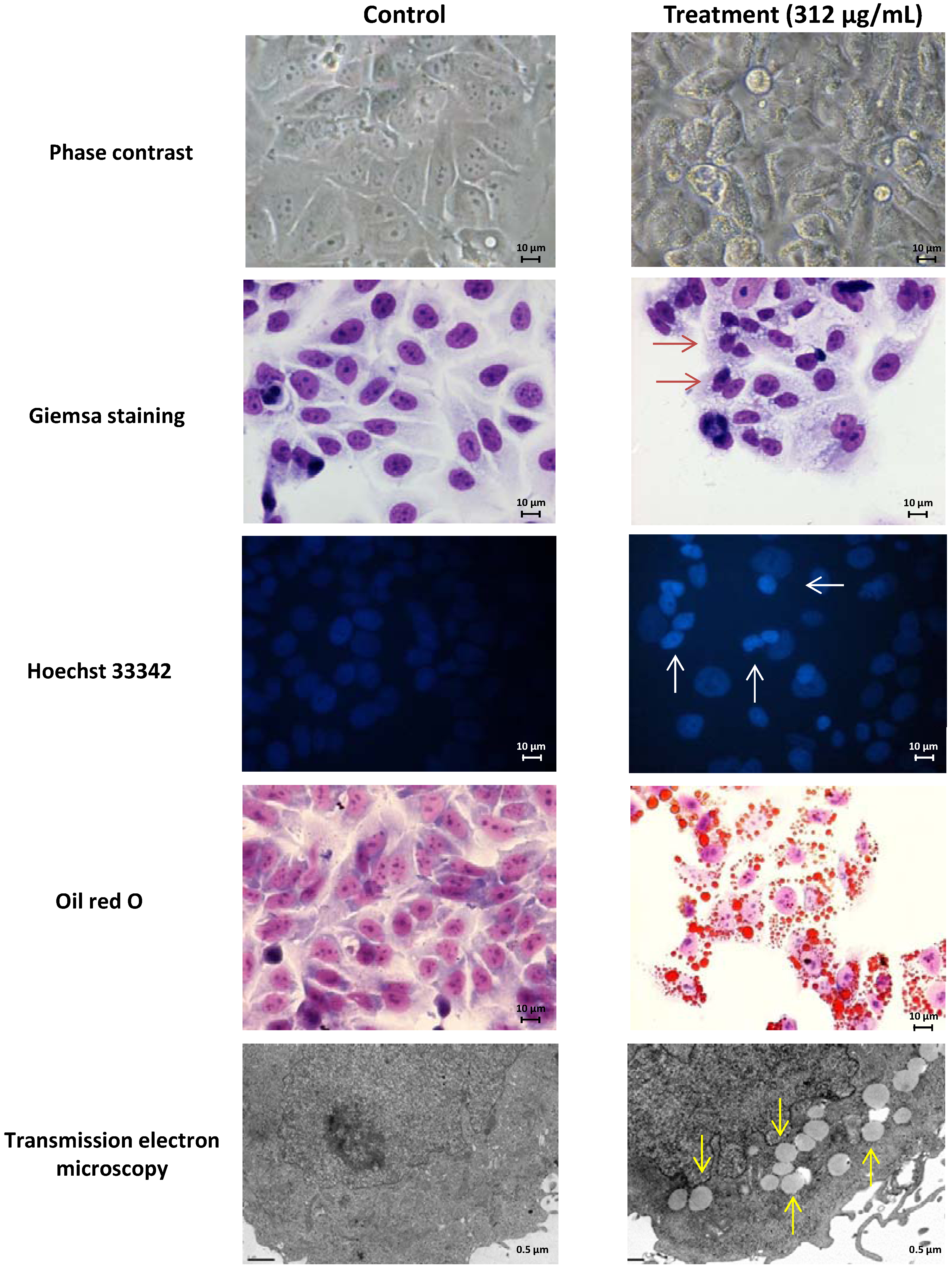
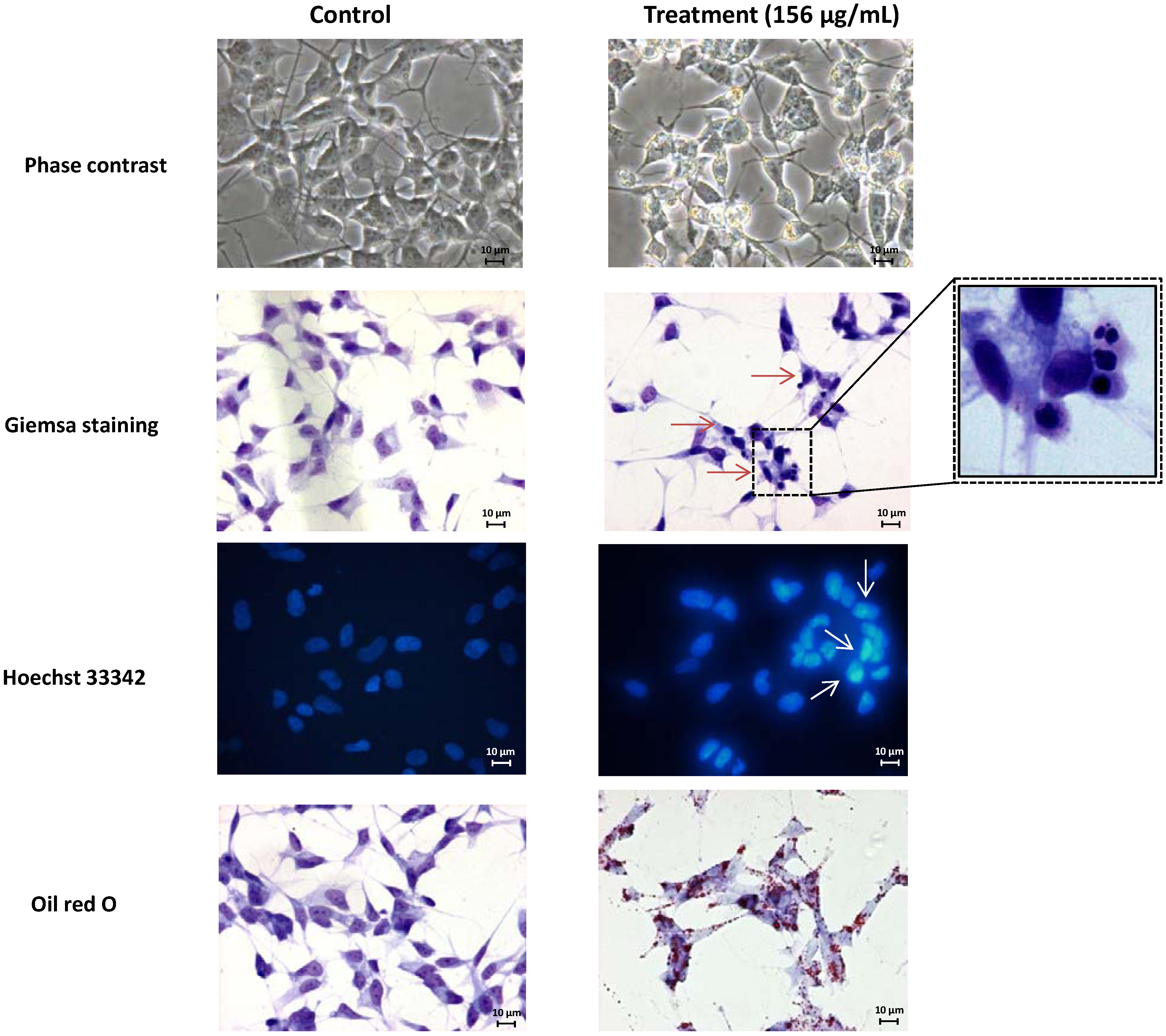
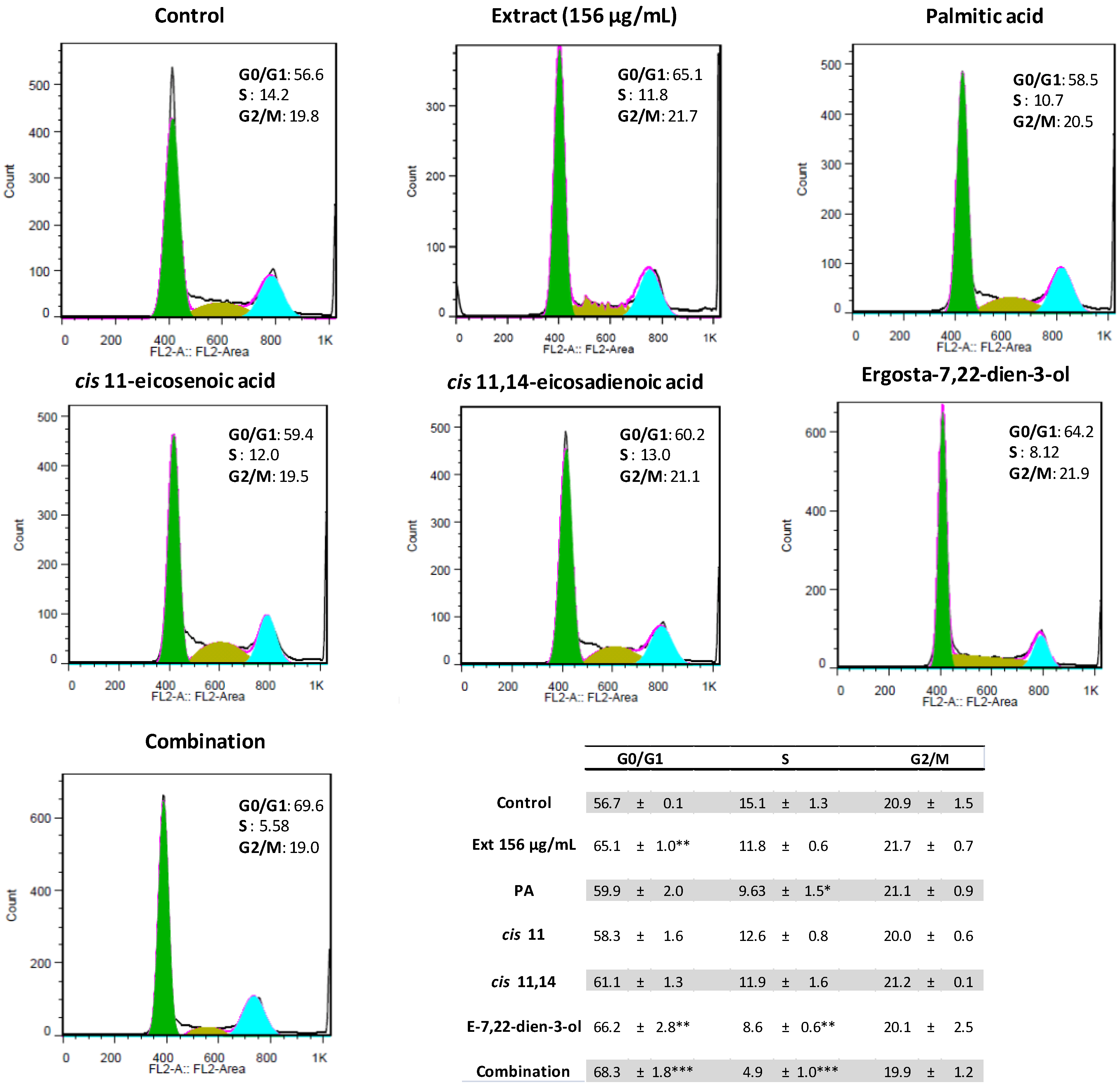
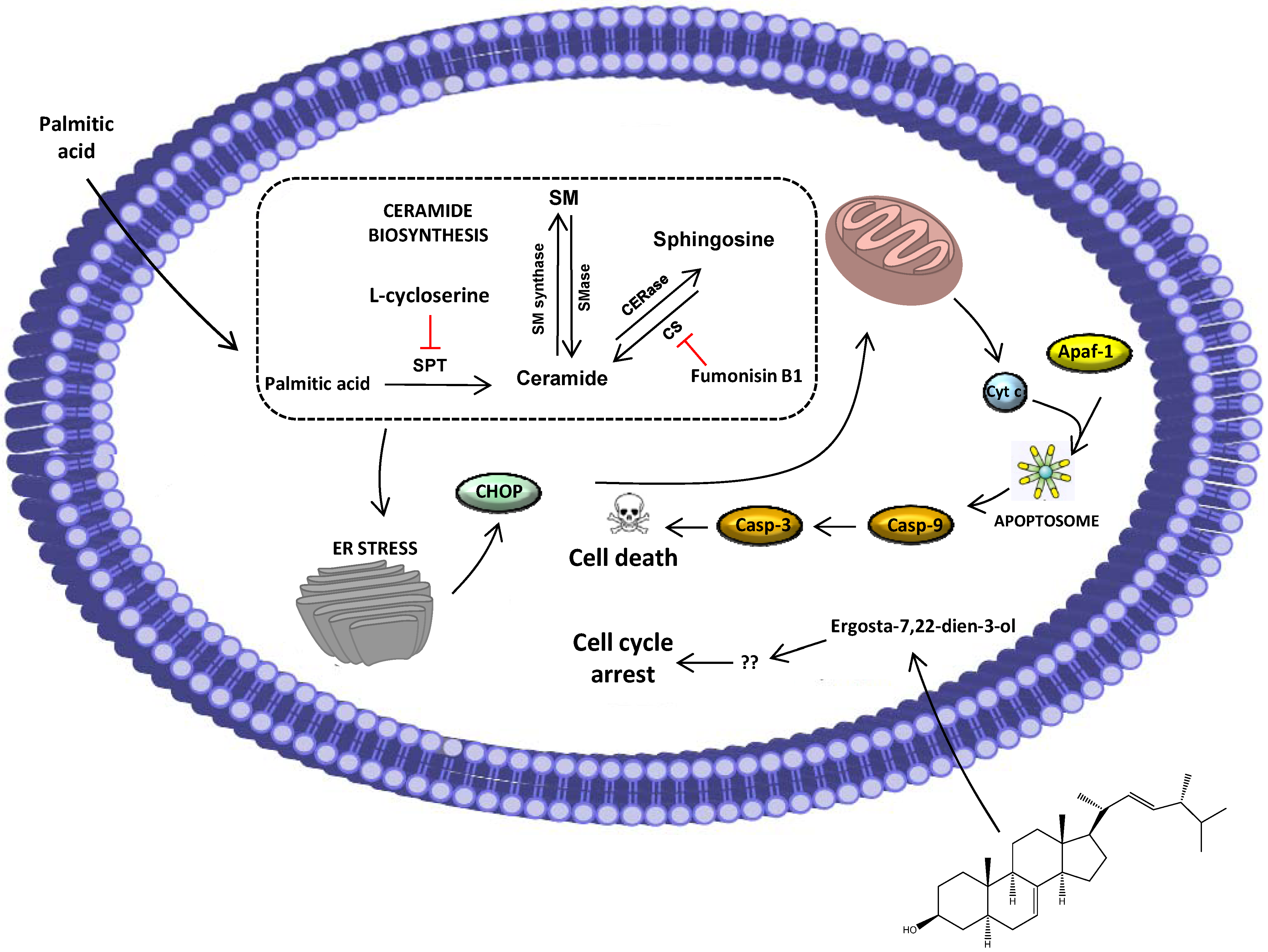
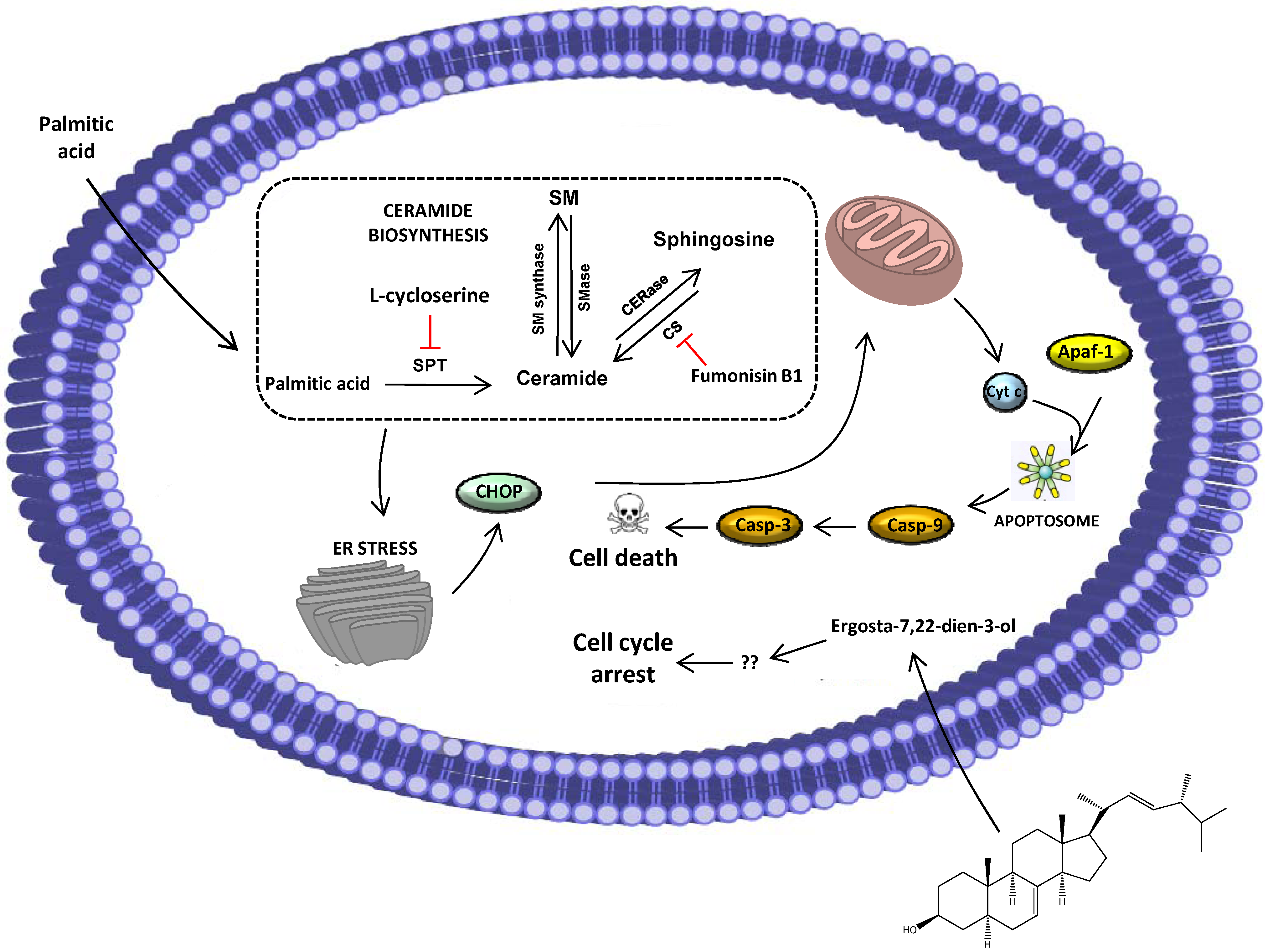
2.2. Ergosta-7,22-dien-3-ol Is Responsible for Cell Cycle Arrest
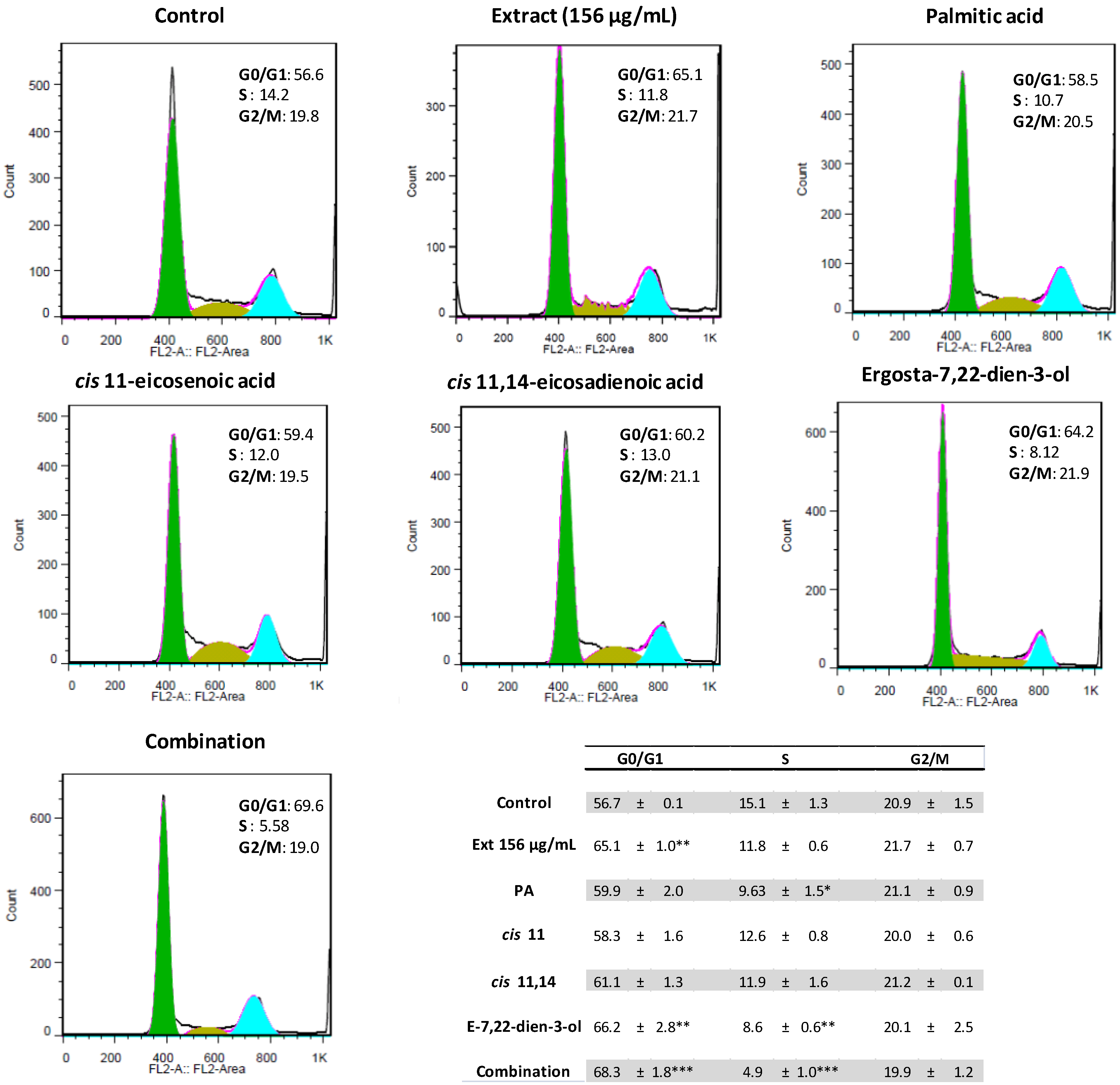
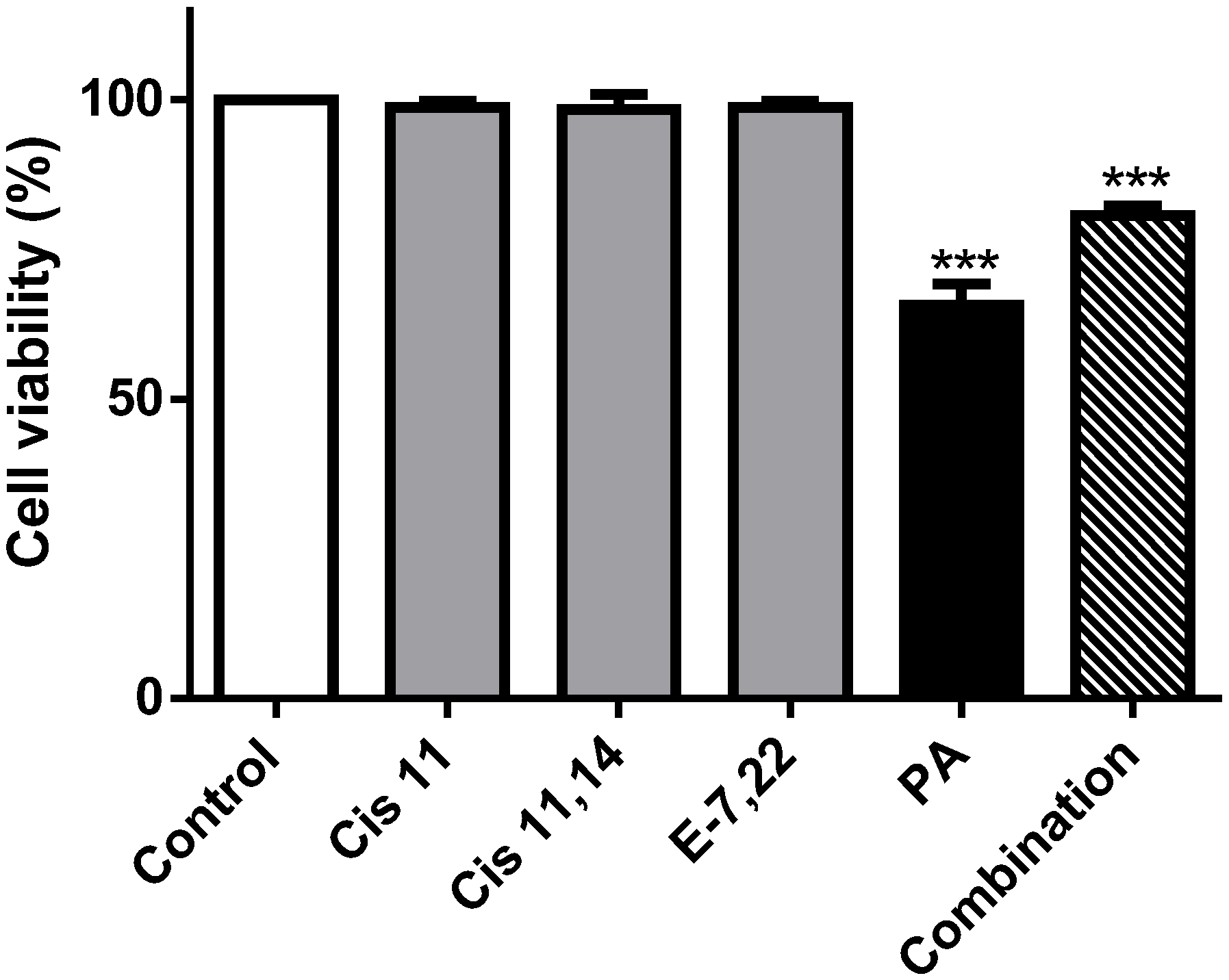
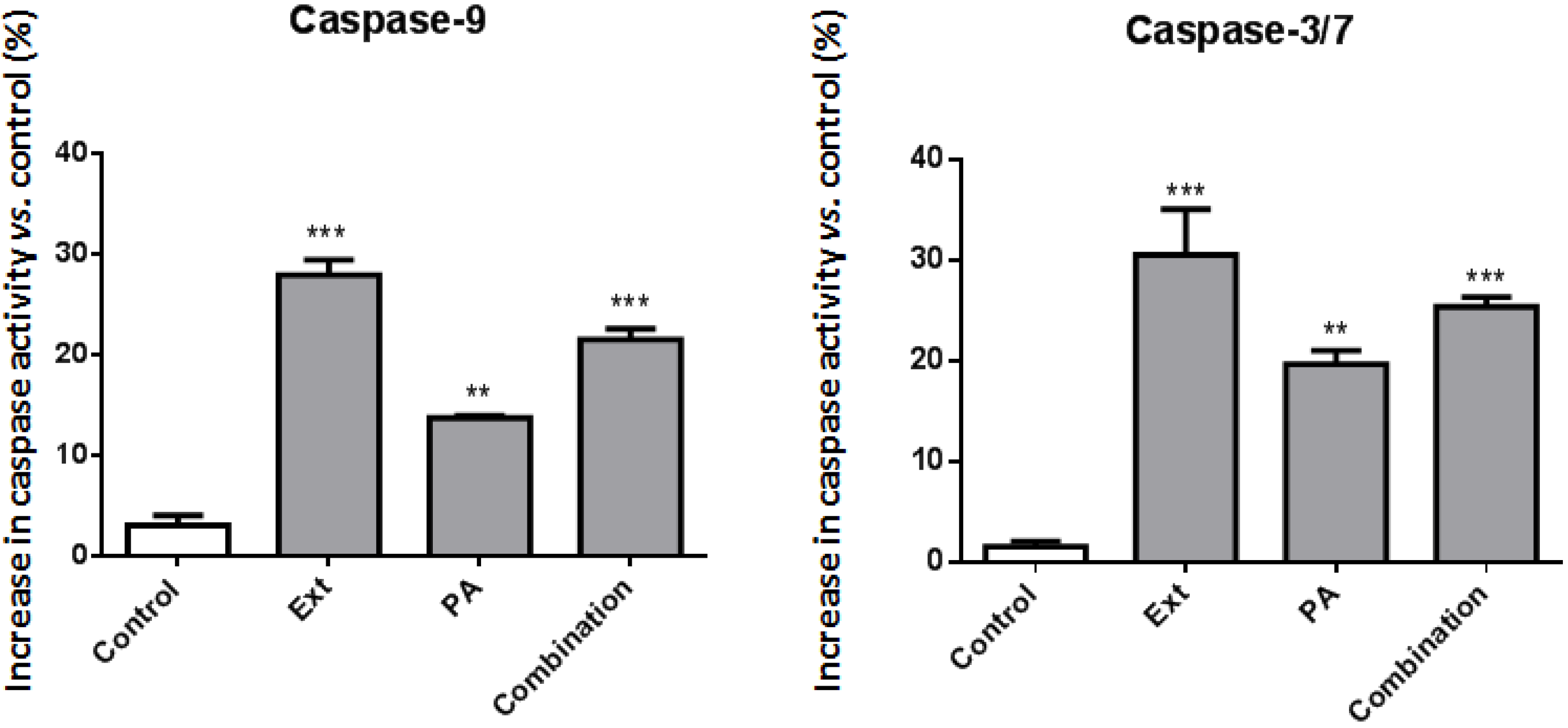
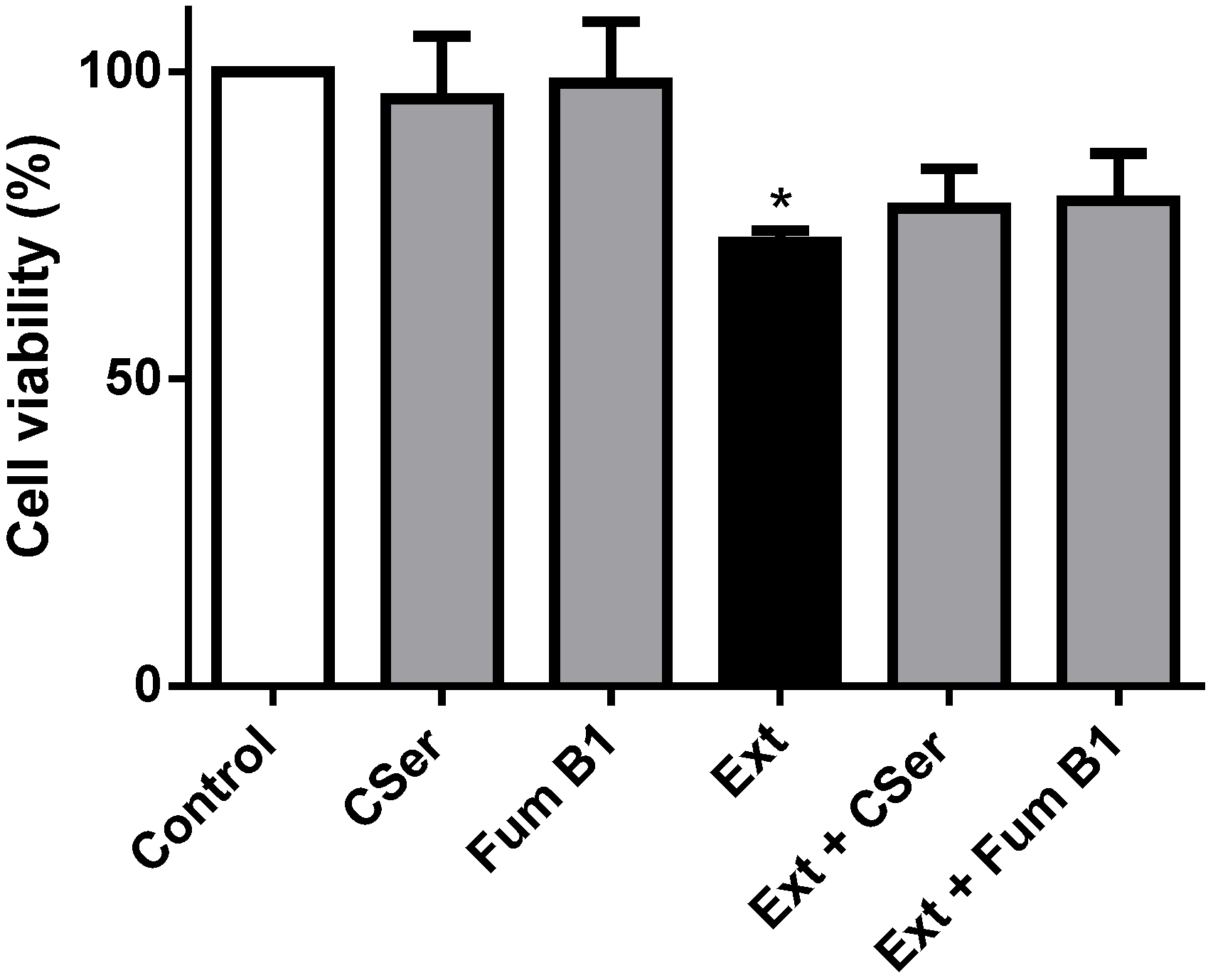
2.3. Palmitic Acid Is Responsible for the Extract-Induced Ceramide-Independent Apoptosis via the Intrinsic Pathway
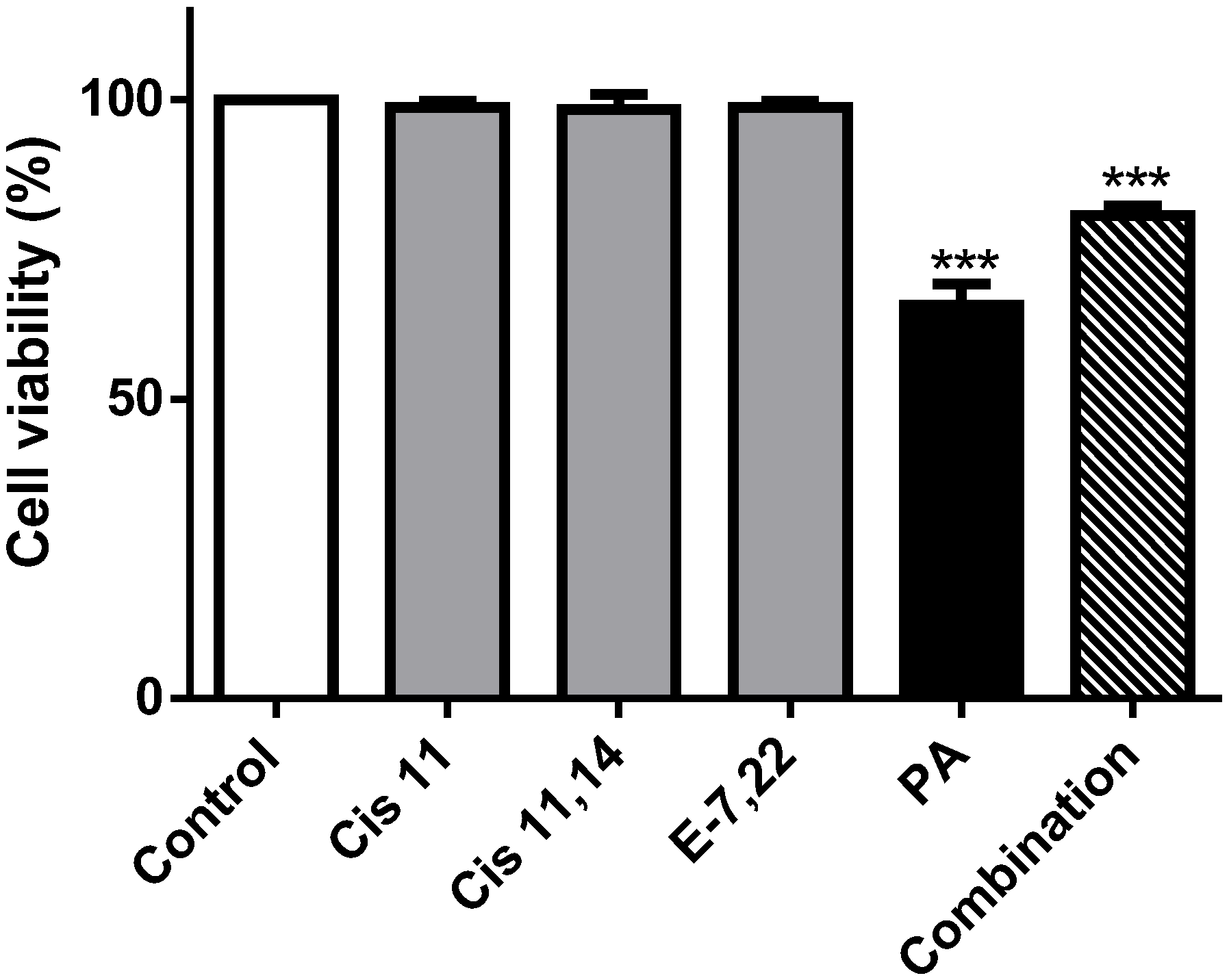
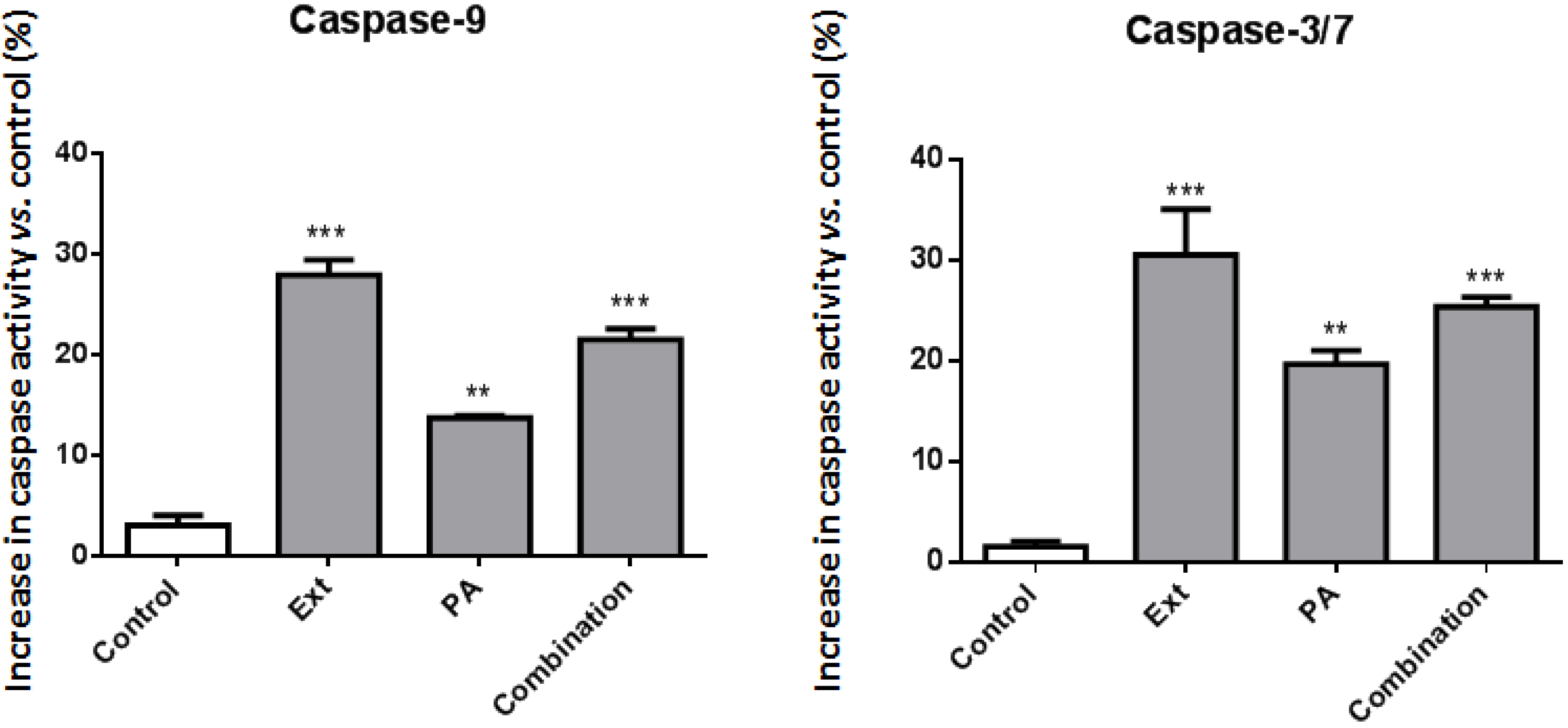
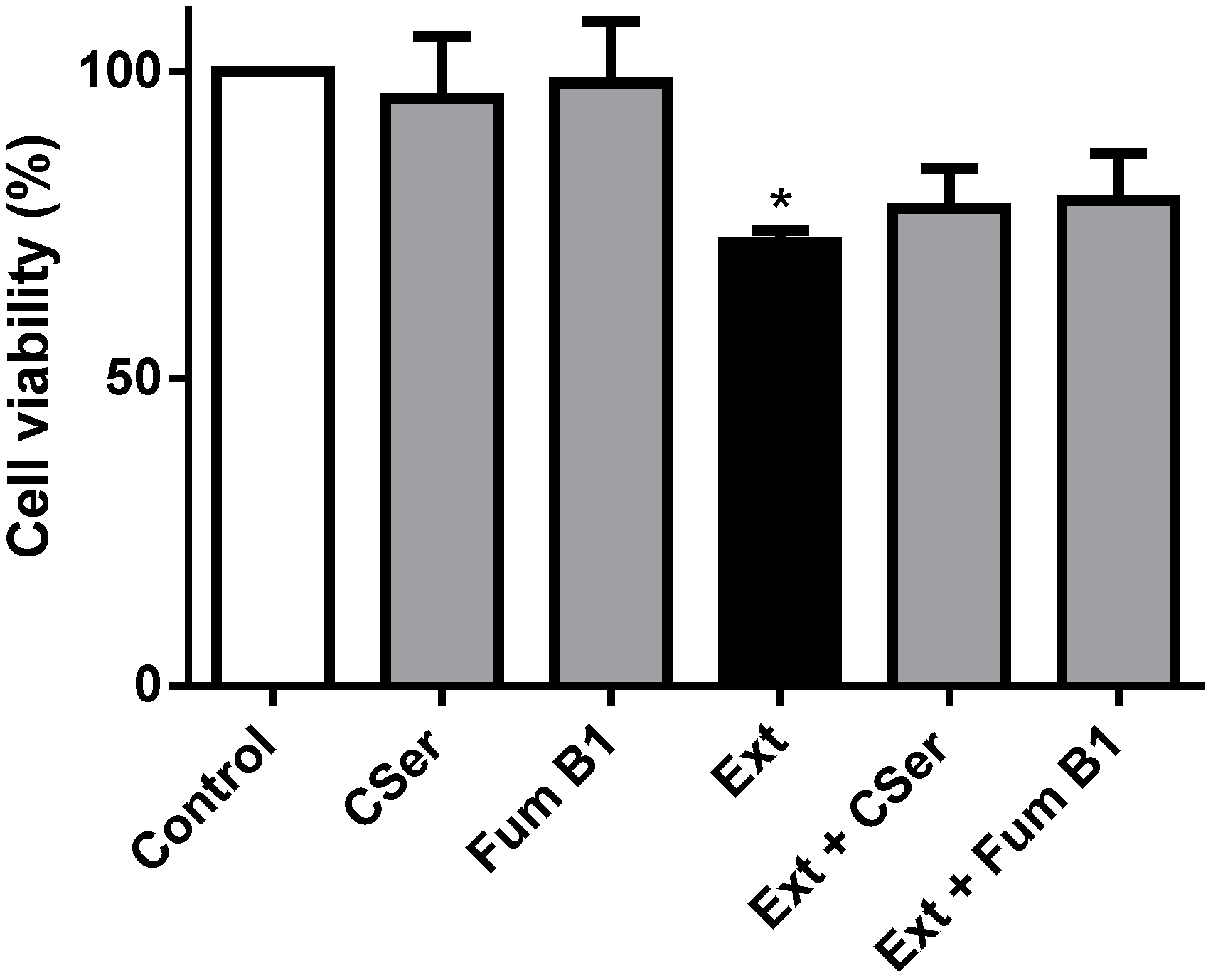
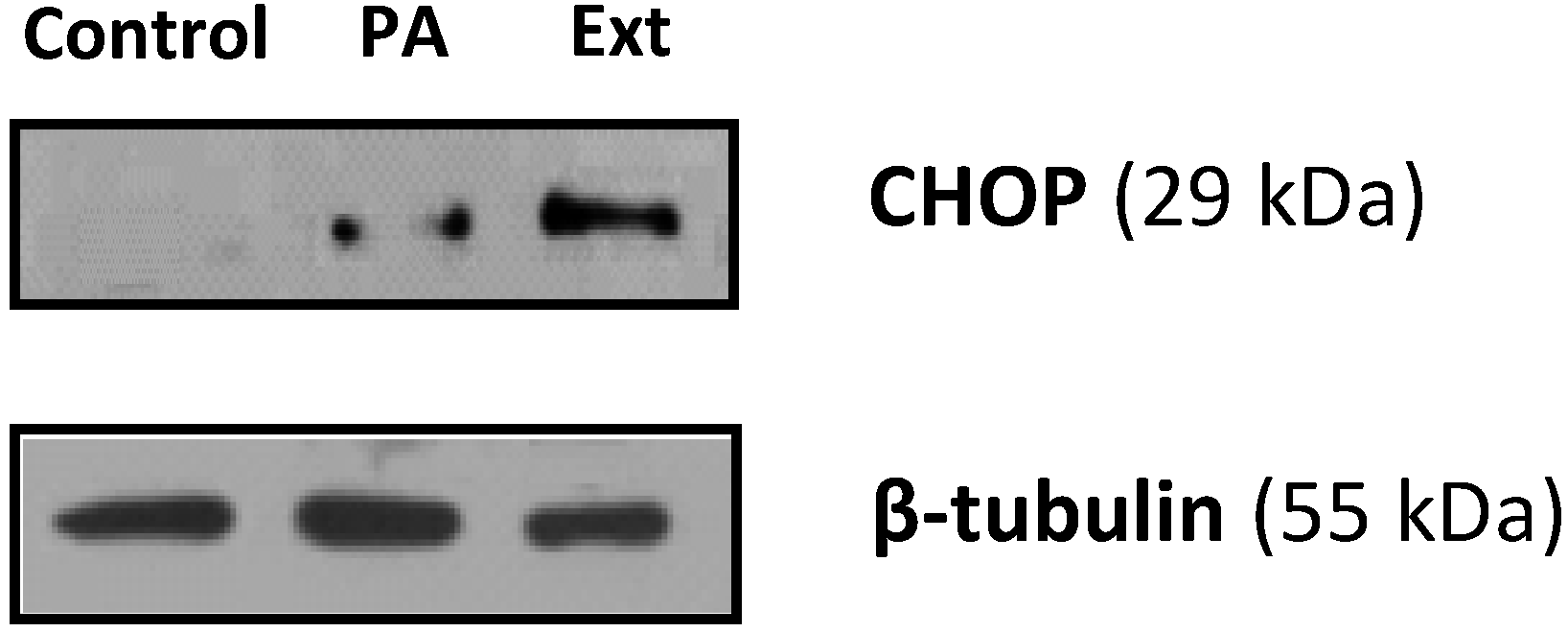
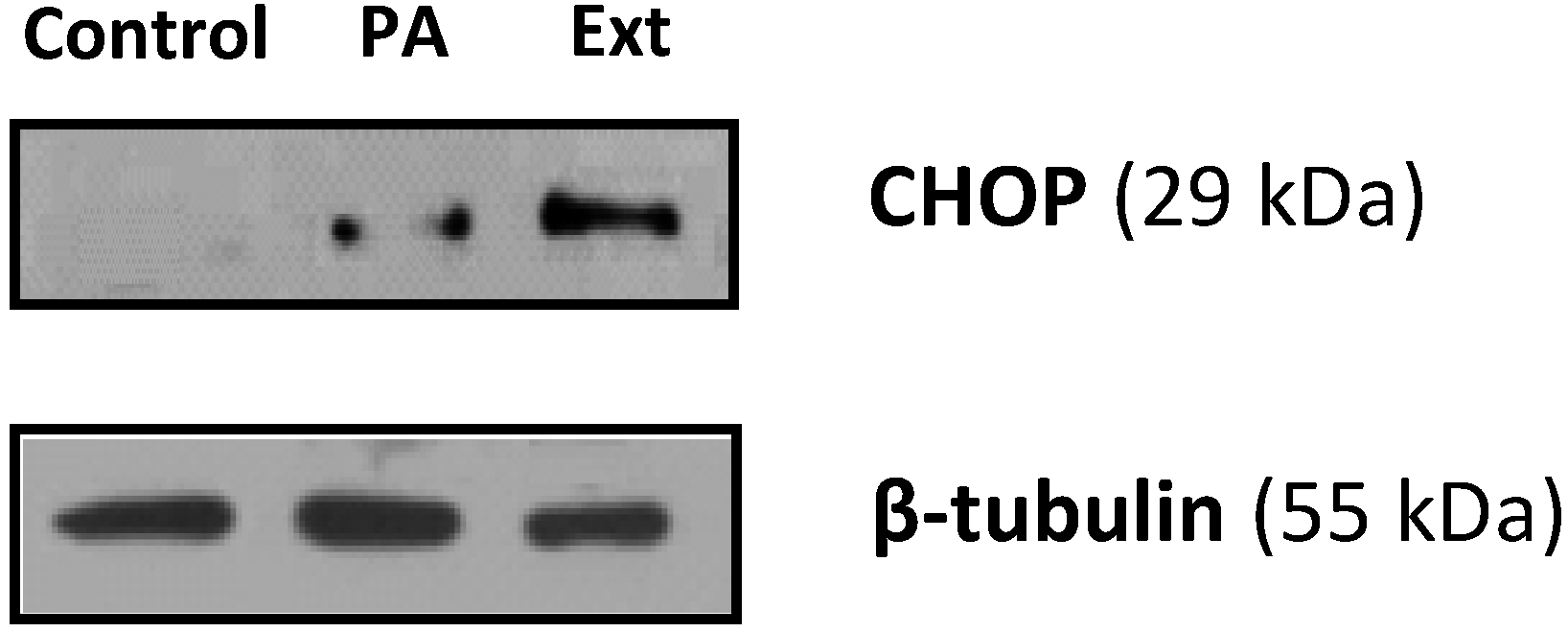
2.4. The Extract and Palmitic Acid Cause ER-Stress Involving CHOP
3. Experimental Section
3.1. Reagents and Standards
3.2. Extract Preparation
3.3. Compounds Preparation
3.4. Cell Culture
3.5. Morphological Studies
3.6. 3H-Thymidine Incorporation Assay
3.7. Cell Cycle Analysis
3.8. Cell Viability
3.9. Caspase-3/-7 and -9 Activity Assay
3.10. Western Blot
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Altmann, K.-H.; Gertsch, J. Anticancer drugs from nature—natural products as a unique source of new microtubule-stabilizing agents. Nat. Prod. Rep. 2007, 24, 327–357. [Google Scholar] [CrossRef]
- Itokawa, H.; Morris-Natschke, S.L.; Akiyama, T.; Lee, K.-H. Plant-derived natural product research aimed at new drug discovery. J. Nat. Med.-Tokyo 2008, 62, 263–280. [Google Scholar] [CrossRef]
- Pereira, D.M.; Valentão, P.; Correia-da-Silva, G.; Teixeira, N.; Andrade, P.B. Plant secondary metabolites in cancer chemotherapy: Where are we? Curr. Pharm. Biotechnol. 2012, 13, 632–650. [Google Scholar] [CrossRef]
- Mayer, A.; Glaser, K.B.; Cuevas, C.; Jacobs, R.S.; Kem, W.; Little, R.D.; McIntosh, J.M.; Newman, D.J.; Potts, B.C.; Shuster, D.E. The odyssey of marine pharmaceuticals: A current pipeline perspective. Trends Pharmacol. Sci. 2010, 31, 255–265. [Google Scholar] [CrossRef]
- Montaser, R.; Luesch, H. Marine natural products: A new wave of drugs? Future Med. Chem. 2011, 3, 1475–1489. [Google Scholar] [CrossRef]
- Pereira, D.M.; Valentão, P.; Andrade, P.B. Lessons from the Sea: Distribution, SAR and Molecular Mechanisms of Anti-inflammatory Drugs from Marine Organisms. In Studies in Natural Products Chemistry (Bioactive Natural Products); Atta-ur-Rahman, Ed.; Elsevier Science Publishers: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Schumacher, M.; Kelkel, M.; Dicato, M.; Diederich, M. Gold from the sea: marine compounds as inhibitors of the hallmarks of cancer. Biotechnol. Adv. 2011, 29, 531–547. [Google Scholar] [CrossRef]
- Ferreres, F.; Pereira, D.M.; Gil-Izquierdo, A.; Valentao, P.; Botelho, J.; Mouga, T.; Andrade, P.B. HPLC-PAD-atmospheric pressure chemical ionization-MS metabolite profiling of cytotoxic carotenoids from the echinoderm Marthasterias glacialis (spiny sea-star). J. Sep. Sci. 2010, 33, 2250–2257. [Google Scholar] [CrossRef]
- Mariutti, L.R.B.; Pereira, D.M.; Mercadante, A.Z.; Valentão, P.; Teixeira, N.; Andrade, P.B. Further insights on the carotenoid profile of the echinoderm Marthasterias glacialis. Mar. Drugs 2012, 10, 1498–1510. [Google Scholar] [CrossRef]
- Pereira, D.M.; Vinholes, J.; Guedes de Pinho, P.; Valentão, P.; Mouga, T.; Teixeira, N.; Andrade, P.B. A gas chromatography-mass spectrometry multi-target method for the simultaneous analysis of three classes of metabolites in marine organisms. Talanta 2012, 100, 391–400. [Google Scholar] [CrossRef]
- Pereira, D.M.; Correia-da-Silva, G.; Valentão, P.; Mouga, T.; Teixeira, N.; Andrade, P.B. A lipidomic approach to drug discovery from marine organisms: Effect of a purified fraction of the lipidome of the echinoderm Marthasterias glacialis L. against human cancer cells. Comb. Chem. High Throughput Screen. 2013, in press. [Google Scholar]
- Ron, D.; Walter, P. Signal integration in the endoplasmic reticulum unfolded protein response. Nat. Rev. Mol. Cell Biol. 2007, 8, 519–529. [Google Scholar] [CrossRef]
- Schroder, M.; Kaufman, R.J. The mammalian unfolded protein response. Annu. Rev. Biochem. 2005, 74, 739–789. [Google Scholar] [CrossRef]
- Zhang, K.; Kaufman, R.J. From endoplasmic-reticulum stress to the inflammatory response. Nature 2008, 454, 455–462. [Google Scholar] [CrossRef]
- Kong, J.Y.; Rabkin, S.W. Palmitate-induced cardiac apoptosis is mediated through CPT-1 but not influenced by glucose and insulin. Am. J. Physiol.-Heart Circ. Physiol. 2002, 282, H717–H725. [Google Scholar]
- Karaskov, E.; Scott, C.; Zhang, L.; Teodoro, T.; Ravazzola, M.; Volchuk, A. Chronic palmitate but not oleate exposure induces endoplasmic reticulum stress, which may contribute to INS-1 pancreatic β-cell apoptosis. Endocrinology 2006, 147, 3398–3407. [Google Scholar] [CrossRef]
- Kolensnick, R.; Hannun, Y.A. Ceramide and apoptosis. Trends Biochem. Sci. 1999, 24, 224–225. [Google Scholar] [CrossRef]
- Morad, S.A.F.; Cabot, M.C. Ceramide-orchestrated signalling in cancer cells. Nat. Rev. Cancer 2013, 13, 51–65. [Google Scholar] [CrossRef]
- Movsesyan, V.A.; Yakovlev, A.G.; Dabaghyan, E.A.; Stoica, B.A.; Faden, A.I. Ceramide induces neuronal apoptosis through the caspase-9/caspase-3 pathway. Biochem. Biophys. Res. Commun. 2002, 299, 201–207. [Google Scholar] [CrossRef]
- Suzuki, J.; Akahane, K.; Nakamura, J.; Naruse, K.; Kamiya, H.; Himeno, T.; Nakamura, N.; Shibata, T.; Kondo, M.; Nagasaki, H.; et al. Palmitate induces apoptosis in Schwann cells via both ceramide-dependent and independent pathways. Neuroscience 2011, 176, 188–198. [Google Scholar] [CrossRef]
- Holland, W.L.; Summers, S.A. Sphingolipids, insulin resistance, and metabolic disease: New insights from in vivo manipulation of sphingolipid metabolism. Endocr. Rev. 2008, 29, 381–402. [Google Scholar] [CrossRef]
- Wei, Y.; Wang, D.; Topczewski, F.; Pagliassotti, M.J. Saturated fatty acids induce endoplasmic reticulum stress and apoptosis independently of ceramide in liver cells. Am. J. Physiol.-Endocrinol. Metab. 2006, 291, E275–E281. [Google Scholar] [CrossRef]
- Bolognesi, A.; Chatgilialoglu, A.; Polito, L.; Ferreri, C. Membrane lipidome reorganization correlates with the fate of neuroblastoma cells supplemented with fatty acids. PLoS One 2013, 8, e55537. [Google Scholar]
- Saleh, M.; Vaillancourt, J.P.; Graham, R.K.; Huyck, M.; Srinivasula, S.M.; Alnemri, E.S.; Steinberg, M.H.; Nolan, V.; Baldwin, C.T.; Hotchkiss, R.S. Differential modulation of endotoxin responsiveness by human caspase-12 polymorphisms. Nature 2004, 429, 75–79. [Google Scholar] [CrossRef]
- Yavari, M.; Brinkley, G.; Klapstein, K.; Hartwig, W.; Rao, R.; Hermel, E. Presence of the functional CASPASE-12 allele in Indian subpopulations. Int. J. Immunogol. 2012, 39, 389–393. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Pereira, D.M.; Correia-da-Silva, G.; Valentão, P.; Teixeira, N.; Andrade, P.B. Palmitic Acid and Ergosta-7,22-dien-3-ol Contribute to the Apoptotic Effect and Cell Cycle Arrest of an Extract from Marthasterias glacialis L. in Neuroblastoma Cells. Mar. Drugs 2014, 12, 54-68. https://doi.org/10.3390/md12010054
Pereira DM, Correia-da-Silva G, Valentão P, Teixeira N, Andrade PB. Palmitic Acid and Ergosta-7,22-dien-3-ol Contribute to the Apoptotic Effect and Cell Cycle Arrest of an Extract from Marthasterias glacialis L. in Neuroblastoma Cells. Marine Drugs. 2014; 12(1):54-68. https://doi.org/10.3390/md12010054
Chicago/Turabian StylePereira, David M., Georgina Correia-da-Silva, Patrícia Valentão, Natércia Teixeira, and Paula B. Andrade. 2014. "Palmitic Acid and Ergosta-7,22-dien-3-ol Contribute to the Apoptotic Effect and Cell Cycle Arrest of an Extract from Marthasterias glacialis L. in Neuroblastoma Cells" Marine Drugs 12, no. 1: 54-68. https://doi.org/10.3390/md12010054