Alternative Methods for the Detection of Emerging Marine Toxins: Biosensors, Biochemical Assays and Cell-Based Assays


Abstract
:1. Introduction
2. Biosensors, Biochemical Assays and Cell-Based Assays



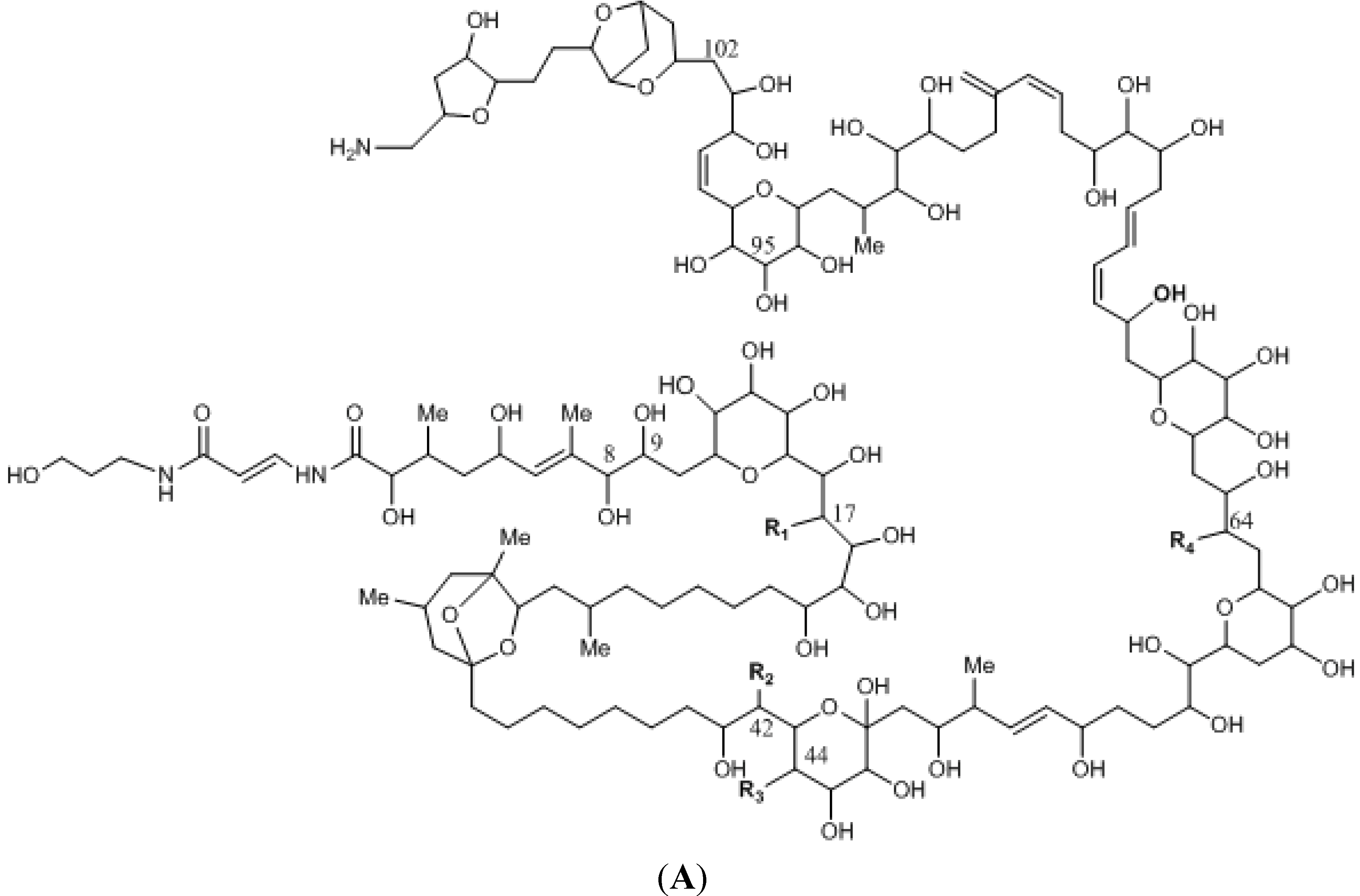{kind=link}
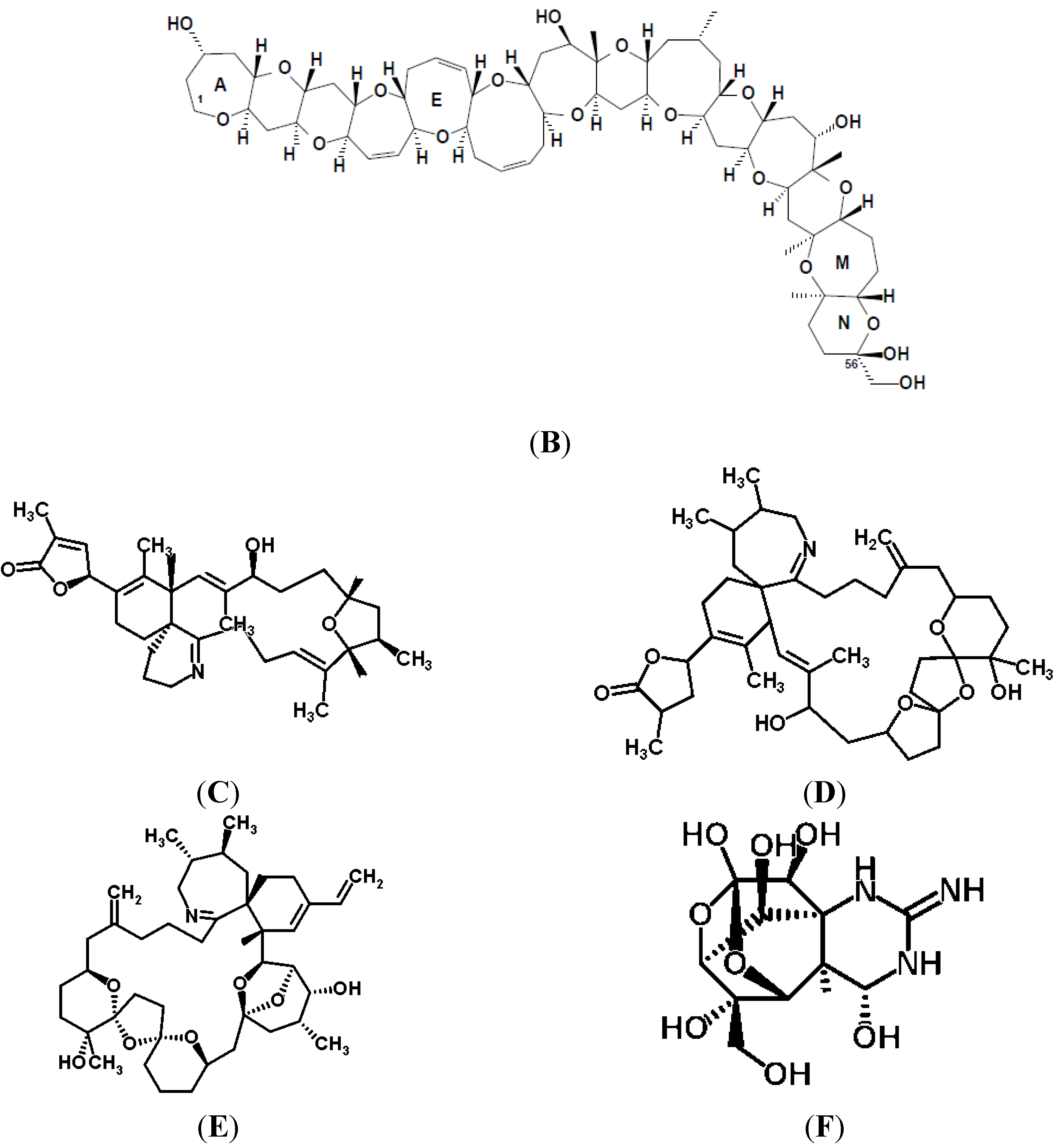{kind=link}
| Toxin | Principal Derivatives | Producer Organism | Vectors | Syndromes and Effects in Humans | Endemic Areas |
|---|---|---|---|---|---|
| PLTX | Palytoxins | Palythoa | Fish, Molluscs, Crustaceans, Echinoderms | Clupeotoxism Rhabdomyolisis Respiratory illness Skin illness Skeletal muscle injury Digestive distress Cardiac distress | Australia, New Zealand, French Polynesia, Cook Islands, Japan, Indonesia, Micronesia, Malaysia, Vietnam, Philippines, Singapore, Hawaii, Caribbean, Mexico, Brazil, Madagascar, Reunion Island, Mediterranean coastline of Europe |
| Putative PLTX Ovatoxins | Ostreopsis cf. ovata | ||||
| Ostreotoxins | Ostreopsis lenticularis | ||||
| Mascarenotoxins | Ostreopsis mascarenensis | ||||
| CTX | Ciguatoxins Gambiertoxins Maitotoxins | Gambierdiscus sp. | Fish | Gastrointestinal effects Neurological effects Cardiovascular effects | French Polynesia, Cook Islands, Hawaii, Japan, Mexico, Tokelau, North Marianna, Tuvalu, Marshall Islands, Fiji, New Caledonia, Tonga, Vanuatu, Samoa, Kiribati, Australia, Micronesia Sri Lanka, Hong Kong, Vietnam, Indonesia, Madagascar, Reunion Island, Maldive Islands, Gulf of Mexico, Guadeloupe, Florida, Virgin Islands, Puerto Rico, Brazil, Canary Islands, Madeira |
| CIs | Gymnodimines | Gymnondinium selliformis | Shellfish | Not reported | Canada, New Zealand, Japan, Denmark, Ireland, Norway, Spain, Italy, Tunisia |
| Spirolids | Alexandrium ostenfeldii | ||||
| Pinnatoxins | Vulcanodinium sp. | ||||
| Pteriatoxins | unknown | ||||
| Prorocentrolides | Prorocentrum sp. | ||||
| TTX | Tetrodotoxins | Vibrio sp. Pseudomonas sp. Alteromonas sp. | Pufferfish, Starfish, Blue-ring Octopus, Xanthid Crabs, Gastropods, Flatworm, Frogs, Goby, Newt Taricha | Ichthyosarcotoxism Numbness Respiratory paralysis Gastrointestinal effects | Japan, China, Taiwan, Madagascar, Australia, New Zealand, Korea, India, New Hampshire, New York, Pennsylvania, Viriginia, Chicago, California, Hawai, Bangladesh, Thailand, Norway, Meditaerranian region (Israel, Egypt and Greece), Spain (fish caught in Portugal) |
2.1. Palytoxins
2.1.1. Immunoassays and Biosensors for Palytoxins
2.1.2. Cell-Based Assays for Palytoxins
2.1.3. Receptor-Binding Assays and Biosensors for Palytoxins
| Assay/Biosensor | Detection Technique | Sample | Reference(s) |
|---|---|---|---|
| Immunoassay | Colorimetry | P. tuberculosa and spiked shellfish | [45] |
| - | [46] | ||
| Aeromonas sp. and Vibrio sp. | [47] | ||
| Spiked mussels, microalgae and seawater | [49] | ||
| Spiked mussels and clams | [50] | ||
| Immunosensor | ECL | Spiked mussels and microalgae | [48] |
| SPR | Grouper and clams | [51] | |
| CBA | Colorimetry | Ostreopsis sp. | [53,54,56] |
| Spiked and naturally-contaminated seafood | [54,56] | ||
| Cyanobacteria trichodesmium | [55] | ||
| - | [57] | ||
| Fluorescence | Spiked and naturally-contaminated seafood and Ostreopsis sp. | [60] | |
| Hemolytic assay | Colorimetry | Corals (Palythoa sp. and Zoanthus sp.) | [70,77,78] |
| Cyanobacteria | [81,85] | ||
| Ostreopsis sp. | [30,37,61,62,63,64,65,66,74,75,76] | ||
| Fish | [43,71,73,79,80,82] | ||
| Sponges, soft coral, gorgonians, crustaceans | [78] | ||
| Mouthed rock shells, sea urchin, mullet, sea-brams | [84] | ||
| Crabs | [77] | ||
| Clams | [37] | ||
| Mussels | [37,75,78] | ||
| Electrochemistry | Mussels | [87] | |
| RBA | FP | Mussels and Ostreopsis sp. | [88] |
| Receptor-based biosensor | SPR | Ostreopsis sp. | [89,90] |
2.2. Ciguatoxins
2.2.1. Immunoassays and Immunosensors for Ciguatoxins
2.2.2. Cell-Based Assays for Ciguatoxins
2.2.3. Receptor-Binding Assays for Ciguatoxins
| Assay | Detection Technique | Sample | Reference(s) |
|---|---|---|---|
| Immunoassays | Colorimetry | Fish | [124,134,135] |
| - | [133,136,137,138,139,140] | ||
| Radioactivity | Fish | [123] | |
| Immunostick test | Colorimetry | Fish | [125,126,127,131,132] |
| CBA | Colorimetry | - | [142] |
| Fish | [54,128,134,141,148,156,161,162,169,170] | ||
| Gambierdiscus sp. | [54,76,163] | ||
| Clams sea urchins and cyanobacteria | [54] | ||
| Rat/mouse blood and urine | [157,158,159,160] | ||
| Cyanobacteria | [171] | ||
| Luminescence | - | [143] | |
| Fluorescence | fish | [150] | |
| Hemolytic assay | Colorimetry | Prorocentrum sp. and fish | [164] |
| Gambierdiscus sp. | [165] | ||
| RBA | Radioactivity | - | [103] |
| Fish | [148,149,155,166] | ||
| [155] [98] | |||
| Fluorescence | - | [168] | |
| Chemiluminiscence | Gambierdiscus sp. | [167] |
2.3. Cyclic Imines
2.3.1. Cell-Based Assays for Cyclic Imines
2.3.2. Receptor-Binding Assays for Cyclic Imines
| Assay | Detection Technique | Sample | Reference(s) |
|---|---|---|---|
| CBA | Colorimetry | - | [195] |
| Vulcanodinium rugosum | [196] | ||
| Patch clamp (electrophisiology) | - | [194] | |
| Hemolytic assay | Colorimetry | Karenia selliformis | [198] |
| RBA | Colorimetry | Cockles | [204] |
| Clams, oysters, scallops and mussels | [206] | ||
| Fluorescence | Cockles | [204] | |
| Fluorescence (coupled to flow cytometry) | Scallops | [207] | |
| FP | Mussels | [200,201,203] | |
| Clams, cockles and scallops | [202] | ||
| Chemiluminiscence | Cockle | [204] | |
| Scallops | [205] |
2.4. Tetrodotoxins
2.4.1. Immunoassays and Immunosensors for Tetrodotoxins
2.4.2. Cell-Based Assays and Biosensors for Tetrodotoxins
| Assay/Biosensor | Detection Technique | Sample | Reference(s) |
|---|---|---|---|
| Immunoassay | Colorimetry | - | [222,224,226,227] |
| Puffer fish | [221,223,225] | ||
| Immunosensor | Electrochemistry | - | [224,228] |
| SPR | - | [233,237] | |
| Pufferfish | [232,234,235] | ||
| Sea snail | [236] | ||
| Human urine | [234] | ||
| Milk and apple juice | [232] | ||
| FFD | - | [238] | |
| Immunostick test | Visual | Spiked puffer fish | [229,230] |
| Fish | [231] | ||
| CBA | Colorimetry | - | [239] |
| Bacteria from freshwater sediments | [242] | ||
| Bacteria from pufferfish | [243] | ||
| Bacterial culture supernatants | [244,245] | ||
| Spiked E. coli | [246] | ||
| Newts | [250] | ||
| Fluorescence | - | [255] | |
| Patch clamp (Electrophysiology) | - | [252,253,254,255] | |
| Pufferfish | [251] |
3. Conclusions and Future Perspectives
Acknowledgments
Author Contributions
Conflicts of Interest
References
- European Food Safety Authority (EFSA). Scientific Opinion on marine biotoxins in shellfish: Cyclic imines (spirolides, gymnodimines, pinnatoxins and pteriatoxins). EFSA J. 2010, 8. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Scientific Opinion on marine biotoxins in shellfish—Emerging toxins: Ciguatoxin group. EFSA J. 2010, 8. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Scientific opinion on marine biotoxins in shellfish-palytoxin group. EFSA J. 2009, 7, 1–38. [Google Scholar]
- European Comission. Regulation (EC) No 854/2004 of the European Parliament and of the Council of 29 April 2004 laying down specific rules for the organisation of official controls on products of animal origin intended for human consumption. Off. J. Eur. Union 2004, L226, 83–127. [Google Scholar]
- European Comission. Regulation (EC) No. 853/2004 of the European Parliament and of the Council of 29 April 2004 laying down specific hygiene rules for food of animal origin. Off. J. Eur. Union 2004, L139, 22–82. [Google Scholar]
- European Comission. Regulation (EU) No 15/2011 of 10 January 2011 amending Regulation (EC) No 2074/2005 as regards recognised testing methods for detecting marine biotoxins in live bivalve molluscs. Off. J. Eur. Union 2011, L6, 3–6. [Google Scholar]
- Foodstuffs—Determination of domoic acid in shellfish and finfish by RP-HPLC using UV detection. Available online: http://aesan.msssi.gob.es/CRLMB/docs/docs/procedimientos/Foodstuffs.pdf (accessed on 11 November 2014).
- Lawrence, J.F.; Niedzwiadek, B.; Menard, C. Quantitative determination of paralytic shellfish poisoning toxins in shellfish using prechromatographic oxidation and liquid cromatography with fluorescence detection: Collaborative Study. J. AOAC Int. 2005, 88, 1714–1732. [Google Scholar]
- Van de Riet, J.M.; Gibbs, R.S.; Chou, F.W.; Muggah, P.M.; Rourke, W.A.; Burns, G.; Thomas, K.; Quilliam, M.A. Liquid chromatographic post-column oxidation method for analysis of paralytic shellfish toxins in mussels, clams, scallops, and oysters: Single-Laboratory Validation. J. AOAC Int. 2009, 92, 1690–1704. [Google Scholar]
- Van de Riet, J.; Gibbs, R.S.; Muggah, P.M.; Rourke, W.A.; MacNeil, J.D.; Quilliam, M.A. Liquid chromatography post-column oxidation (PCOX) method for the determination of paralytic shellfish toxins in mussels, clams, oysters, and scallops: Collaborative study. J. AOAC Int. 2011, 94, 1154–1176. [Google Scholar]
- Turner, A.D.; Lewis, A.M.; Rourke, W.A.; Higman, W.A. Inter laboratory comparison of two AOAC liquid chromatographic fluorescence detection methods for paralytic shellfish toxin analysis through characterization of an oyster reference material. J. AOAC Int. 2014, 97, 380–390. [Google Scholar] [CrossRef]
- Gerssen, A.; Mulder, P.; McElhinney, M.A.; de Boer, J. Liquid chromatography-tandem mass spectrometry method for the detection of marine lipophilic toxins under alkaline conditions. J. Chromatogr. A 2009, 1216, 1421–1430. [Google Scholar]
- Fux, E.; McMillan, D.; Bire, R.; Hess, P. Development of an ultra-performance liquid chromatography-mass spectrometry method for the detection of lipophilic marine toxins. J. Chromatogr. A 2007, 1157, 273–280. [Google Scholar]
- McNabb, P.; Selwood, A.I.; Holland, P.T. Multiresidue method for determination of algal toxins in shellfish: Single-laboratory validation and interlaboratory study. J. AOAC Int. 2005, 88, 761–772. [Google Scholar]
- Takada, N.; Umemura, N.; Suenaga, K.; Chou, T.; Nagatsu, A.; Haino, T.; Yamada, K.; Uemura, D. Pinnatoxins B and C, the most toxic components in the pinnatoxin series from the Okinawan bivalve Pinna muricata. Tetrahedron Lett. 2001, 42, 3491–3494. [Google Scholar] [CrossRef]
- Miles, C.O.; Rundberget, T.; Sandvik, M.; Aasen, J.; Selwood, A.I. The Presence of Pinnatoxins in Norwegian Mussels; National Veterinary Institute: Oslo, Norway, 2010. [Google Scholar]
- Lewis, R.J.; Jones, A.; Vernoux, J.-P. HPLC/Tandem Electrospray Mass Spectrometry for the determination of sub-ppb levels of Pacific and Caribbean ciguatoxins in crude extracts of fish. Anal. Chem. 1998, 71, 247–250. [Google Scholar]
- Hamilton, B.; Hurbungs, M.; Jones, A.; Lewis, R.J. Multiple ciguatoxins present in Indian Ocean reef fish. Toxicon 2002, 40, 1347–1353. [Google Scholar] [CrossRef]
- Munday, R. Palytoxin toxicology: Animal studies. Toxicon 2011, 57, 470–477. [Google Scholar] [CrossRef]
- Moore, R.E.; Scheuer, P.J. Palytoxin: A new marine toxin from a coelenterate. Science 1971, 172, 495–498. [Google Scholar] [CrossRef]
- Walsh, G.E.B.; Ralph, L. A review of Hawaiian zoanthids with descriptions of three new species. Zool. J. Linn. Soc. 1971, 50, 161–180. [Google Scholar] [CrossRef]
- Moore, R.E.; Bartolini, G. Structure of palytoxin. J. Am. Chem. Soc. 1981, 103, 2491–2494. [Google Scholar] [CrossRef]
- Uemura, D.; Ueda, K.; Hirata, Y.; Naoki, H.; Iwashita, T. Further studies on palytoxin. II. Structure of palytoxin. Tetrahedron Lett. 1981, 22, 2781–2784. [Google Scholar] [CrossRef]
- Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; Grauso, L.; Tartaglione, L.; Florio, C.; Lorenzon, P.; de Bortoli, M. Stereostructure and biological activity of 42-hydroxy-palytoxin: A new palytoxin analogue from Hawaiian Palythoa subspecies. Chem. Res. Toxicol. 2009, 22, 1851–1859. [Google Scholar] [CrossRef]
- Uemura, D.; Hirata, Y.; Iwashita, T.; Naoki, H. Studies on palytoxins. Tetrahedron 1985, 41, 1007–1017. [Google Scholar] [CrossRef]
- Inuzuka, T.; Uemura, D.; Arimoto, H. The conformational features of palytoxin in aqueous solution. Tetrahedron 2008, 64, 7718–7723. [Google Scholar] [CrossRef]
- Mercado, J.A.; Rivera-Rentas, A.L.; Gonzalez, I.; Tosteson, T.R.; Molgó, J.; Escalona de Motta, G. Neuro- and myo-toxicity of extracts from the benthic dinoflagellate Ostreopsis lenticularis is sensitive to μ-conotoxin. Soc. Neurosci. 1994, 20, 718. [Google Scholar]
- Ukena, T.; Satake, M.; Usami, M.; Oshima, Y.; Naoki, H.; Fujita, T.; Kan, Y.; Yasumoto, T. Structure elucidation of ostreocin D, a palytoxin analog isolated from the dinoflagellate Ostreopsis siamensis. Biosci. Biotechnol. Biochem. 2001, 65, 2585–2588. [Google Scholar] [CrossRef]
- Ukena, T.; Satake, M.; Usami, M.; Oshima, Y.; Fujita, T.; Naoki, H.; Yasumoto, T. Structural confirmation of ostreocin-D by application of negative-ion fast-atom bombardment collision-induced dissociation tandem mass spectrometric methods. Rapid Commun. Mass Spectrom. 2002, 16, 2387–2393. [Google Scholar] [CrossRef]
- Lenoir, S.; Ten-Hage, L.; Turquet, J.; Quod, J.P.; Bernard, C.; Hennion, M.C. First evidence of palytoxin analogues from an Ostreopsis mascarenensis (Dinophyceae) benthic bloom in Southwestern Indian Ocean. J. Phycol. 2004, 40, 1042–1051. [Google Scholar] [CrossRef]
- Tubaro, A.; Durando, P.; del Favero, G.; Ansaldi, F.; Icardi, G.; Deeds, J.R.; Sosa, S. Case definitions for human poisonings postulated to palytoxins exposure. Toxicon 2011, 57, 478–495. [Google Scholar] [CrossRef]
- Milandri, A.; Ceredi, A.; Riccardi, E.; Gasperetti, L.; Susini, F.; Casotti, M.; Faiman, L.; Pigozzi, S. Impact of Ostreopsis Ovata on Marine Benthic Communities: Accumulation of Palytoxins in Mussels, Sea Urchins and Octopuses from Italy, Proceedings of the ICHA14 Conference, Crete, Greek, 1–5 November 2010; pp. 1–5.
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Grillo, C.; Melchiorre, N. Putative palytoxin and its new analogue, Ovatoxin-a, in Ostreopsis ovata collected along the Ligurian Coasts during the 2006 toxic outbreak. J. Am. Soc. Mass Spectrom. 2008, 19, 111–120. [Google Scholar] [CrossRef]
- Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; Grauso, L.; Tartaglione, L.; Guerrini, F.; Pistocchi, R. Complex palytoxin-like profile of Ostreopsis ovata. Identification of four new ovatoxins by high-resolution liquid chromatography/mass spectrometry. Rapid Commun. Mass Spectrom. 2010, 24, 2735–2744. [Google Scholar]
- Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Battocchi, C.; Crinelli, R.; Carloni, E.; Magnani, M.; et al. Unique toxin profile of a Mediterranean Ostreopsis cf. ovata Strain: HR LC-MSn characterization of ovatoxin-f, a new palytoxin congener. Chem. Res. Toxicol. 2012, 25, 1243–1252. [Google Scholar]
- Munday, R. Occurrence and toxicology of palytoxins. Food Sci. Technol. 2008, 173, 693–713. [Google Scholar]
- Aligizaki, K.; Katikou, P.; Milandri, A.; Diogène, J. Occurrence of palytoxin-group toxins in seafood and future strategies to complement the present state of the art. Toxicon 2011, 57, 390–399. [Google Scholar] [CrossRef]
- Hilgemann, D.W. From a pump to a pore: How palytoxin opens the gates. Proc. Natl. Acad. Sci. USA 2003, 100, 386–388. [Google Scholar] [CrossRef]
- Artigas, P.; Gadsby, D.C. Na+/K+-pump ligands modulate gating of palytoxin-induced ion channels. Proc. Natl. Acad. Sci. USA 2003, 100, 501–505. [Google Scholar] [CrossRef]
- Shimahara, T.; Molgó, J. Palytoxin enhances quantal acetylcholine release from motor nerve terminals and increases cytoplasmic calcium levels in a neuronal hybrid cell line. Life Sci. Adv. Pharmacol. 1990, 9, 785–792. [Google Scholar]
- Satoh, E.; Ishii, T.; Nishimura, M. Palytoxin-induced increase in cytosolic-free Ca2+ in mouse spleen cells. Eur. J. Pharmacol. 2003, 465, 9–13. [Google Scholar] [CrossRef]
- Deeds, J.R.; Schwartz, M.D. Human risk associated with palytoxin exposure. Toxicon 2010, 56, 150–162. [Google Scholar] [CrossRef]
- Onuma, Y.; Satake, M.; Ukena, T.; Roux, J.; Chanteau, S.; Rasolofonirina, N.; Ratsimaloto, M.; Naoki, H.; Yasumoto, T. Identification of putative palytoxin as the cause of clupeotoxism. Toxicon 1999, 37, 55–65. [Google Scholar] [CrossRef]
- CRLMB (Community Reference Laboratory for Marine Biotoxins). Report on toxicology working group meeting, 24–25 October 2005; CRLMB: Cesenatico, Italy; pp. 24–25.
- Bignami, G.S.; Raybould, T.J.G.; Sachinvala, N.D.; Grothaus, P.G.; Simpson, S.B.; Lazo, C.B.; Byrnes, J.B.; Moore, R.E.; Vann, D.C. Monoclonal antibody-based enzyme-linked immunoassays for the measurement of palytoxin in biological samples. Toxicon 1992, 30, 687–700. [Google Scholar] [CrossRef]
- Levine, L.; Fujiki, H.; Gjika, H.B.; Vanvunakis, H. Production of antibodies to palytoxin—neutralization of several biological properties of playtoxin. Toxicon 1987, 25, 1273–1282. [Google Scholar] [CrossRef]
- Frolova, G.; Kuznetsova, T.; Mikhailov, V.; Elyakov, G. An enzyme linked immunosorbent assay for detecting palytoxin-producing bacteria. Russ. J. Bioorg. Chem. 2000, 26, 285–289. [Google Scholar] [CrossRef]
- Zamolo, V.A.; Valenti, G.; Venturelli, E.; Chaloin, O.; Marcaccio, M.; Boscolo, S.; Castagnola, V.; Sosa, S.; Berti, F.; Fontanive, G.; et al. Highly sensitive electrochemiluminescent nanobiosensor for the detection of palytoxin. ACS Nano 2012, 6, 7989–7997. [Google Scholar] [CrossRef]
- Boscolo, S. Sandwich ELISA assay for the quantitation of palytoxin and its analogs in natural samples. Environ. Sci. Technol. 2013, 47, 2034–2042. [Google Scholar] [CrossRef]
- Garet, E.; Cabado, A.G.; Vieites, J.M.; Gonzalez-Fernandez, A. Rapid isolation of single-chain antibodies by phage display technology directed against one of the most potent marine toxins: Palytoxin. Toxicon 2010, 55, 1519–1526. [Google Scholar] [CrossRef]
- Yakes, B.; DeGrasse, S.; Poli, M.; Deeds, J. Antibody characterization and immunoassays for palytoxin using an SPR biosensor. Anal. Bioanal. Chem. 2011, 400, 2865–2869. [Google Scholar] [CrossRef] [PubMed]
- Wattenberg, E.V. Modulation of protein kinase signaling cascades by palytoxin. Toxicon 2011, 57, 440–448. [Google Scholar] [CrossRef]
- Bellocci, M.; Ronzitti, G.; Milandri, A.; Melchiorre, N.; Grillo, C.; Poletti, R.; Yasumoto, T.; Rossini, G.P. A cytolytic assay for the measurement of palytoxin based on a cultured monolayer cell line. Anal. Biochem. 2008, 374, 48–55. [Google Scholar] [CrossRef]
- Pawlowiez, R.; Darius, H.T.; Cruchet, P.; Rossi, F.; Caillaud, A.; Laurent, D.; Chinain, M. Evaluation of seafood toxicity in the Australes archipelago (French Polynesia) using the neuroblastoma cell-based assay. Food Add. Contam. A 2013, 30, 567–586. [Google Scholar] [CrossRef]
- Kerbrat, A.S.; Amzil, Z.; Pawlowiez, R.; Golubic, S.; Sibat, M.; Darius, H.T.; Chinain, M.; Laurent, D. First evidence of palytoxin and 42-hydroxy-palytoxin in the marine cyanobacterium Trichodesmium. Mar. Drugs 2011, 9, 543–560. [Google Scholar] [CrossRef]
- Ledreux, A.; Krys, S.; Bernard, C. Suitability of the Neuro-2a cell line for the detection of palytoxin and analogues (neurotoxic phycotoxins). Toxicon 2009, 53, 300–308. [Google Scholar] [CrossRef]
- Cañete, E.; Diogène, J. Comparative study of the use of neuroblastoma cells (Neuro-2a) and neuroblastoma-glioma hybrid cells (NG108-15) for the toxic effect quantification of marine toxins. Toxicon 2008, 52, 541–550. [Google Scholar] [CrossRef]
- Rossini, G.P.; Bigiani, A. Palytoxin action on the Na+, K+-ATPase and the disruption of ion equilibria in biological systems. Toxicon 2011, 57, 429–439. [Google Scholar] [CrossRef]
- Bellocci, M.; Sala, G.L.; Prandi, S. The cytolytic and cytotoxic activities of palytoxin. Toxicon 2011, 57, 449–459. [Google Scholar] [CrossRef]
- Espiña, B.; Cagide, E.; Louzao, M.; Fernandez, M.; Vieytes, M.; Katikou, P.; Villar, A.; Jaen, D.; Maman, L.; Botana, L. Specific and dynamic detection of palytoxins by in vitro microplate assay with human neuroblastoma cells. Biosci. Rep. 2009, 29, 13–23. [Google Scholar] [CrossRef]
- Penna, A.; Fraga, S.; Battocchi, C.; Casabianca, S.; Giacobbe, M.G.; Riobo, P.; Vernesi, C. A phylogeographical study of the toxic benthic dinoflagellate genus Ostreopsis Schmidt. J. Biogeogr. 2010, 37, 830–841. [Google Scholar] [CrossRef] [Green Version]
- Pezzolesi, L.; Guerrini, F.; Ciminiello, P.; Dell’Aversano, C.; Iacovo, E.D.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Pistocchi, R. Influence of temperature and salinity on Ostreopsis cf. ovata growth and evaluation of toxin content through HR LC-MS and biological assays. Water Res. 2012, 46, 82–92. [Google Scholar]
- Nascimento, S.M.; Correa, E.V.; Menezes, M.; Varela, D.; Paredes, J.; Morris, S. Growth and toxin profile of Ostreopsis cf. ovata (Dinophyta) from Rio de Janeiro, Brazil. Harmful Algae 2012, 13, 1–9. [Google Scholar]
- Vidyarathna, N.K.; Graneli, E. Influence of temperature on growth, toxicity and carbohydrate production of a Japanese Ostreopsis ovata strain, a toxic-bloom-forming dinoflagellate. Aquat. Microb. Ecol. 2011, 65, 261–270. [Google Scholar]
- Vidyarathna, N.K.; Graneli, E. Physiological responses of Ostreopsis ovata to changes in N and P availability and temperature increase. Harmful Algae 2013, 21–22, 54–63. [Google Scholar]
- Riobo, P.; Paz, B.; Franco, J.M. Analysis of palytoxin-like in Ostreopsis cultures by liquid chromatography with precolumn derivatization and fluorescence detection. Anal. Chim. Acta 2006, 566, 217–223. [Google Scholar] [CrossRef]
- Habermann, E.; Ahnerthilger, G.; Chhatwal, G.S.; Beress, L. Delayed hemolytic action of palytoxin. General characteristics. General characteristics. Biochim. Biophys. Acta 1981, 649, 481–486. [Google Scholar] [CrossRef]
- Habermann, E.; Chhatwal, G.S.; Hessler, H.J. Palytoxin raises the nonspecific permeability of erythrocytes in an ouabain-sensitive manner. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1981, 317, 374. [Google Scholar]
- Habermann, E.; Chhatwal, G.S. Ouabain inhibits the increase due to palytoxin of cation permeability of erythrocytes. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1982, 319, 101–107. [Google Scholar]
- Bignami, G.S. A rapid and sensitive hemolysis neutralization assay for palytoxin. Toxicon 1993, 31, 817–820. [Google Scholar] [CrossRef]
- Taniyama, S.; Arakawa, O.; Terada, M.; Nishio, S.; Takatani, T.; Mahmud, Y.; Noguchi, T. Ostreopsis sp., a possible origin of palytoxin (PTX) in parrotfish Scarus ovifrons. Toxicon 2003, 42, 29–33. [Google Scholar] [CrossRef]
- Riobo, P.; Paz, B.; Franco, J.M.; Vazquez, J.A.; Murado, M.A. Proposal for a simple and sensitive haemolytic assay for palytoxin. Toxicological dynamics, kinetics, ouabain inhibition and thermal stability. Harmful Algae 2008, 7, 415–429. [Google Scholar]
- Taniyama, S.; Mahmud, Y.; Terada, M.; Takatani, T.; Arakawa, O.; Noguchi, T. Occurrence of a food poisoning incident by palytoxin from a serranid Epinephelus sp. in Japan. J. Nat. Toxins 2002, 11, 277–282. [Google Scholar]
- Rhodes, L.; Adamson, J.; Suzuki, T.; Briggs, L.; Garthwaite, I. Toxic marine epiphytic dinoflagellates, Ostreopsis siamensis and Coolia monotis (Dinophyceae), in New Zealand. N. Z. J. Mar. Freshw. Res. 2000, 34, 371–383. [Google Scholar] [CrossRef]
- Rhodes, L.; Towers, N.; Briggs, L.; Munday, R.; Adamson, J. Uptake of palytoxin-like compounds by shellfish fed Ostreopsis siamensis (Dinophyceae). N. Z. J. Mar. Freshw. Res. 2002, 36, 631–636. [Google Scholar] [CrossRef]
- Rhodes, L.L.; Smith, K.F.; Munday, R.; Selwood, A.I.; McNabb, P.S.; Holland, P.T.; Bottein, M.-Y. Toxic dinoflagellates (Dinophyceae) from Rarotonga, Cook Islands. Toxicon 2010, 56, 751–758. [Google Scholar] [CrossRef]
- Gleibs, S.; Mebs, D.; Werding, B. Studies on the origin and distribution of palytoxin in a Caribbean coral reef. Toxicon 1995, 33, 1531–1537. [Google Scholar] [CrossRef]
- Gleibs, S.; Mebs, D. Distribution and sequestration of palytoxin in coral reef animals. Toxicon 1999, 37, 1521–1527. [Google Scholar] [CrossRef]
- Wachi, K.M.; Hokama, Y.; Haga, L.S.; Shiraki, A.; Takenaka, W.E.; Bignami, G.S.; Levine, L. Evidence for palytoxin as one of the sheep erythrocyte lytic in lytic factors in crude extracts of ciguateric and non-ciguateric reef fish tissue. J. Nat. Toxins 2000, 9, 139–146. [Google Scholar]
- Wachi, K.M.; Hokama, Y. Diversity of marine biotoxins in the near-shore ocean area: Presence of a palytoxin-like entity at Barber’s Point Harbor, Oahu. J. Nat. Toxins 2001, 10, 317–333. [Google Scholar]
- Seemann, P.; Gernert, C.; Schmitt, S.; Mebs, D.; Hentschel, U. Detection of hemolytic bacteria from Palythoa caribaeorum (Cnidaria, Zoantharia) using a novel palytoxin-screening assay. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2009, 96, 405–411. [Google Scholar] [CrossRef]
- Huang, K.M.; Liu, S.M.; Huang, Y.W.; Huang, K.L.; Hwang, D.F. Food poisoning caused by Sunfish Masturus lanceolatus in Taiwan. J. Food Drug Anal. 2011, 19, 191–196. [Google Scholar]
- Bire, R.; Trotereau, S.; Lemee, R.; Delpont, C.; Chabot, B.; Aumond, Y.; Krys, S. Occurrence of palytoxins in marine organisms from different trophic levels of the French Mediterranean coast harvested in 2009. Harmful Algae 2013, 28, 10–22. [Google Scholar] [CrossRef]
- Brissard, C.; Herrenknecht, C.; Sechet, V.; Herve, F.; Pisapia, F.; Harcouet, J.; Lemee, R.; Chomerat, N.; Hess, P.; Amzil, Z. Complex toxin profile of French Mediterranean Ostreopsis cf. ovata strains, seafood accumulation and ovatoxins prepurification. Mar. Drugs 2014, 12, 2851–2876. [Google Scholar]
- Pagliara, P.; Caroppo, C. Cytotoxic and antimitotic activities in aqueous extracts of eight cyanobacterial strains isolated from the marine sponge Petrosia ficiformis. Toxicon 2011, 57, 889–896. [Google Scholar] [CrossRef]
- Igarashi, T.; Aritake, S.; Yasumoto, T. Mechanisms underlying the hemolytic and ichthyotoxic activities of maitotoxin. Nat. Toxins 1999, 7, 71–79. [Google Scholar] [CrossRef]
- Volpe, G.; Cozzi, L.; Migliorelli, D.; Croci, L.; Palleschi, G. Development of a haemolytic-enzymatic assay with mediated amperometric detection for palytoxin analysis: Application to mussels. Anal. Bioanal. Chem. 2014, 406, 2399–2410. [Google Scholar] [CrossRef]
- Alfonso, A.; Fernandez-Araujo, A.; Alfonso, C.; Carames, B.; Tobio, A.; Louzao, M.C.; Vieytes, M.R.; Botana, L.M. Palytoxin detection and quantification using the fluorescence polarization technique. Anal. Biochem. 2012, 424, 64–70. [Google Scholar] [CrossRef]
- Vale-Gonzalez, C.; Pazos, M.; Alfonso, A.; Vieytes, M.; Botana, L. Study of the neuronal effects of ouabain and palytoxin and their binding to Na+, K+-ATPases using an optical biosensor. Toxicon 2007, 50, 541–552. [Google Scholar] [CrossRef]
- Alfonso, A.; Pazos, M.-J.; Fernández-Araujo, A.; Tobio, A.; Alfonso, C.; Vieytes, M.R.; Botana, L.M. Surface plasmon resonance biosensor method for palytoxin detection based on Na+, K+-ATPase affinity. Toxins 2014, 6, 96–107. [Google Scholar]
- Scheuer, P.; Takahaski, W.; Tsutsumi, J.; Yoshida, T. Ciguatoxin: Isolation and chemical structure. Science 1967, 155, 1267–1268. [Google Scholar] [CrossRef]
- Lange, W.R. Ciguatera fish poisoning. Am. Fam. Phys. 1994, 50, 579–584. [Google Scholar]
- Yasumoto, T.; Nakajima, I.; Bagnis, R.; Adachi, R. Finding of a dinoflagellate as a likely culprit of ciguatera. Bull. Jpn. Soc. Sci. Fish. 1977, 43, 1021–1026. [Google Scholar] [CrossRef]
- Bagnis, R.; Hurtel, J.M.; Chanteau, S.; Chungue, E.; Inoue, A.; Yasumoto, T. Dinoflagellate Gambierdiscus—Toxicus Adachi et Fukuyo—Probable cause of ciguatera. Comptes Rendus Hebd. Seances L’Acad. Sci. D 1979, 289, 671–674. [Google Scholar]
- Bagnis, R.; Chanteau, S.; Chungue, E.; Hurtel, J.M.; Yasumoto, T.; Inoue, A. Origins of ciguatera fish poisoning—A new Dinoflagellate, Gambierdiscus-Toxicus Adachi and Fukuyo, definitively involved as a causal agent. Toxicon 1980, 19, 199–208. [Google Scholar] [CrossRef]
- Sperr, A.E.; Doucette, G.J. Variation in growth rate and ciguatera toxin production among geographically distinct isolates of Gambierdiscus toxicus. In Harmful and Toxic Algal Blooms; Yasumoto, T., Oshima, Y., Fukuyo, Y., Eds.; IOC of UNESCO: Paris, France, 1996; pp. 309–312. [Google Scholar]
- Litaker, R.W.; Vandersea, M.W.; Faust, M.A.; Kibler, S.R.; Nau, A.W.; Holland, W.C.; Chinain, M.; Holmes, M.J.; Tester, P.A. Global distribution of ciguatera causing dinoflagellates in the genus Gambierdiscus. Toxicon 2010, 56, 711–730. [Google Scholar] [CrossRef]
- Chinain, M.; Darius, H.T.; Ung, A.; Cruchet, P.; Wang, Z.; Ponton, D.; Laurent, D.; Pauillac, S. Growth and toxin production in the ciguatera-causing dinoflagellate Gambierdiscus polynesiensis (Dinophyceae) in culture. Toxicon 2010, 56, 739–750. [Google Scholar] [CrossRef]
- Mills, A.R. Poisonous fish in the South Pacific. J. Trop. Med. Hyg. 1956, 59, 99–103. [Google Scholar]
- Vernoux, J.P.; Lahlou, N.; Elandaloussi, S.A.; Riyeche, N.; Magras, L.P. A study of the distribution of ciguatoxin in indiviual Caribbean fish. Acta Trop. 1985, 42, 225–233. [Google Scholar]
- Murata, M.; Legrand, A.M.; Ishibashi, Y.; Yasumoto, T. Structures of ciguatoxin and its congener. J. Am. Chem. Soc. 1989, 111, 8929–8931. [Google Scholar] [CrossRef]
- Murata, M.; Legrand, A.N.; Ishibashi, Y.; Fukui, M.; Yasumoto, T. Conformations of ciguatoxin and related polyethers. Abstr. Pap. Am. Chem. Soc. 1990, 200. 54-AGFD. [Google Scholar]
- Lewis, R.J.; Sellin, M.; Poli, M.A.; Norton, R.S.; MacLeod, J.K.; Sheil, M.M. Purification and characterization of ciguatoxins from moray eel (Lycodontis javanicus, Muraenidae). Toxicon 1991, 29, 1115–1127. [Google Scholar] [CrossRef]
- Satake, M.; Ishimaru, T.; Legrand, A.M.; Yasumoto, T. Isolation of a ciguatoxin analog from cultures of Gambierdiscus toxicus. In Toxic Pyhtoplankton Blooms in the Sea; Samyda, T.J., Shimizu, Y., Eds.; Elsevier: New York, NY, USA, 1993; Volume 3; pp. 575–579. [Google Scholar]
- Satake, M.; Murata, M.; Yasumoto, T. The structure of CTX3C, a ciguatoxin congener isolated from cultured Gambierdiscus toxicus. Tetrahedron Lett. 1993, 34, 1975–1978. [Google Scholar] [CrossRef]
- Satake, M.; Morohashi, A.; Oguri, H.; Oishi, T.; Hirama, M.; Harada, N.; Yasumoto, T. The absolute configuration of ciguatoxin. J. Am. Chem. Soc. 1997, 119, 11325–11326. [Google Scholar] [CrossRef]
- Satake, M.; Fukui, M.; Legrand, A.M.; Cruchet, P.; Yasumoto, T. Isolation and structures of new ciguatoxin analogs, 2,3-dihydroxyCTX3C and 51-hydroxyCTX3C, accumulated in tropical reef fish. Tetrahedron Lett. 1998, 39, 1197–1198. [Google Scholar] [CrossRef]
- Lehane, L.; Lewis, R.J. Ciguatera: Recent advances but the risk remains. Int. J. Food Microbiol. 2000, 61, 91–125. [Google Scholar] [CrossRef]
- Lewis, R.J.; Vernoux, J.P.; Brereton, I.M. Structure of Caribbean ciguatoxin isolated from Caranx latus. J. Am. Chem. Soc. 1998, 120, 5914–5920. [Google Scholar] [CrossRef]
- Hamilton, B.; Hurbungs, M.; Vernoux, J.P.; Jones, A.; Lewis, R.J. Isolation and characterisation of Indian Ocean ciguatoxin. Toxicon 2002, 40, 685–693. [Google Scholar] [CrossRef]
- Bidard, J.N.; Vijverberg, H.P.M.; Frelin, C.; Chungue, E.; Legrand, A.M.; Bagnis, R.; Lazdunski, M. Ciguatoxin is a novel type of Na+ channel toxin. J. Biol. Chem. 1984, 259, 8353–8357. [Google Scholar]
- Molgo, J.; Comella, J.X.; Legrand, A.M. Ciguatoxin enhances quantal transmitter release from frog motor-nerve terminals. Br. J. Pharmacol. 1990, 99, 695–700. [Google Scholar] [CrossRef]
- Molgo, J.; Gaudrytalarmain, Y.M.; Legrand, A.M.; Moulian, N. Ciguatoxin extracted from poisonous moray eels gymnothorax-javanicus triggers acetylcholine-release from torpedo cholinergic synaptosomes via reversed Na+/Ca2+ exchange. Neurosci. Lett. 1993, 160, 65–68. [Google Scholar] [CrossRef]
- Benoit, E.; Juzans, P.; Legrand, A.M.; Molgo, J. Nodal swelling produced by ciguatoxin-induced selective activation of sodium channels in myelinated nerve fibers. Neuroscience 1996, 71, 1121–1131. [Google Scholar] [CrossRef] [PubMed]
- Mattei, C.; Dechraoui, M.Y.; Molgo, J.; Meunier, F.A.; Legrand, A.M.; Benoit, E. Neurotoxins targetting receptor site 5 of voltage-dependent sodium channels increase the nodal volume of myelinated axons. J. Neurosci. Res. 1999, 55, 666–673. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J. Ion channel toxins and therapeutics: From cone snail venoms to ciguatera. Ther. Drug Monit. 2000, 22, 61–64. [Google Scholar] [CrossRef]
- Caillaud, A.; de la Iglesia, P.; Darius, H.T.; Pauillac, S.; Aligizaki, K.; Fraga, S.; Chinain, M.; Diogene, J. Update on methodologies available for ciguatoxin determination: Perspectives to confront the onset of ciguatera fish poisoning in Europe. Mar. Drugs 2010, 8, 1838–1907. [Google Scholar] [CrossRef]
- Perez-Arellano, J.L.; Luzardo, O.P.; Brito, A.P.; Cabrera, M.H.; Zumbado, M.; Carranza, C.; Angel-Moreno, A.; Dickey, R.W.; Boada, L.D. Ciguatera fish poisoning, Canary Islands. Emerg. Infect. Dis. 2005, 11, 1981–1982. [Google Scholar] [CrossRef]
- Gouveia, N.; Delgado, J.; Vale, P. Primeiro registo da ocorrência de episódios do tipo ciguatérico no arquipélago da Madeira. In Abstract Book of X Reuniao Oberica, Fitoplancton Toxico e Biotoxinas; IPIMAR: Lisbon, Portugal, 2009. [Google Scholar]
- Fraga, S. Global climate change and harmful algal blooms (HABs). In Abstract Book of 4th European Phycological Congress; Elsevier Science: Oviedo, Spain, 2007; p. 41. [Google Scholar]
- Stebbing, A.R.D.; Turk, S.M.T.; Wheeler, A.; Clarke, K.R. Immigration of southern fish species to south-west England linked to warming of the North Atlantic (1960–2001). J. Mar. Biol. Assoc. UK 2002, 82, 177–180. [Google Scholar] [CrossRef]
- CDC (Centers for Disease Control and Prevention). Cluster of Ciguatera Fish Poisoning,2007. Morbidity and Mortality Weekly Report (MMWR); CDC: North Carolina, NC, USA, 2009; pp. 283–285. [Google Scholar]
- Hokama, Y.; Banner, A.H.; Boylan, D.B. A radioimmunoassay for the detection of ciguatoxin. Toxicon 1977, 15, 317–325. [Google Scholar] [CrossRef]
- Hokama, Y.; Abad, M.A.; Kimura, L.H. A rapid enzyme-immunoassay for the detection of ciguatoxin in contaminated fish-tissues. Toxicon 1983, 21, 817–824. [Google Scholar] [CrossRef]
- Hokama, Y. A rapid, simplified enzyme-immunoassay stick test for the detecction of ciguatoxin and related polyethers from fish-tissues. Toxicon 1985, 23, 939–946. [Google Scholar] [CrossRef]
- Hokama, Y.; Shirai, L.K.; Iwamoto, L.M.; Kobayashi, M.N.; Goto, C.S.; Nakagawa, L.K. Assessment of a rapid enzyme-immunoassay stick test for the detection of ciguatoxin and related polyether toxins in fish-tissues. Biol. Bull. 1987, 172, 144–153. [Google Scholar] [CrossRef]
- Park, D.L. Detection of ciguatera and diarrheic shellfish toxins in finfish and shellfish with ciguatect kit. J. AOAC Int. 1995, 78, 533–537. [Google Scholar]
- Bienfang, P.; DeFelice, S.; Dowling, A. Quantitative evaluation of commercially available test kit for ciguatera in fish. Food Nutr. Sci. 2011, 2, 594–598. [Google Scholar] [CrossRef]
- Ebesu, J.S.; Campora, C.E. Comment on “Quantitative evaluation of commercially available test kit for ciguatera in fish”. Food Nutr. Sci. 2012, 3, 1233–1237. [Google Scholar] [CrossRef]
- Dickey, R.W.; Granade, H.R.; McClure, F.D. uatect Solid-Phase Immunobead Assay for the Detection of Ciguatera-Related Biotoxins in Caribbean Finfish. In Proceedings of the International Workshop on Ciguatera Management, Brisbane, Australia, 13–16 April 1994; Memoirs of the Queensland Museum: Brisbane, Australia, 1994; pp. 481–488. [Google Scholar]
- Hokama, Y.; Honda, S.A.A.; Kobayashi, M.N.; Nakagawa, L.K.; Asahina, A.Y.; Miyahara, J.T. Monoclonal-antibody (MAb) in detection of ciguatoxin (CTX) and related polyethers by the stick-enzyme immunoassay (S-EIA) in fish-tissues associated with ciguatera poisoning. In Mycotoxins and Phycotoxins; Natori, S., Hashimoto, K., Ueno, Y., Eds.; Elsevier: Amsterdam, The Netherlands, 1989; Volume 10; pp. 303–310. [Google Scholar]
- Campora, C.E.; Hokama, Y.; Ebesu, J.S.M. Comparative analysis of purified pacific and Caribbean ciguatoxin congeners and related marine toxins using a modified ELISA technique. J. Clin. Lab. Anal. 2006, 20, 121–125. [Google Scholar] [CrossRef]
- Pauillac, S.; Sasaki, M.; Inoue, M.; Naar, J.; Branaa, P.; Chinain, M.; Tachibana, K.; Legrand, A.M. Characterization of mice antisera elicited with a ciguatoxin tetracyclic synthetic ring fragment (JKLM) conjugated to carrier proteins. Toxicon 2000, 38, 669–685. [Google Scholar] [CrossRef]
- Campora, C.E.; Dierking, J.; Tamaru, C.S.; Hokama, Y.; Vincent, D. Detection of ciguatoxin in fish tissue using sandwich ELISA and neuroblastoma cell bioassay. J. Clin. Lab. Anal. 2008, 22, 246–253. [Google Scholar] [CrossRef]
- Campora, C.E.; Hokama, Y.; Yabusaki, K.; Isobe, M. Development of an enzyme-linked immunosorbent assay for the detection of ciguatoxin in fish tissue using chicken immunoglobulin Y. J. Clin. Lab. Anal. 2008, 22, 239–245. [Google Scholar] [CrossRef]
- Oguri, H.; Hirama, M.; Tsumuraya, T.; Fujii, I.; Maruyama, M.; Uehara, H.; Nagumo, Y. Synthesis-based approach toward direct sandwich immunoassay for ciguatoxin CTX3C. J. Am. Chem. Soc. 2003, 125, 7608–7612. [Google Scholar] [CrossRef]
- Tsumuraya, T.; Fujii, I.; Inoue, M.; Tatami, A.; Miyazaki, K.; Hirama, M. Production of monoclonal antibodies for sandwich immunoassay detection of ciguatoxin 51-hydroxyCTX3C. Toxicon 2006, 48, 287–294. [Google Scholar] [CrossRef]
- Tsumuraya, T.; Fujii, I.; Hirama, M. Production of monoclonal antibodies for sandwich immunoassay detection of Pacific ciguatoxins. Toxicon 2010, 56, 797–803. [Google Scholar] [CrossRef]
- Tsumuraya, T.; Takeuchi, K.; Yamashita, S.; Fujii, I.; Hirama, M. Development of a monoclonal antibody against the left wing of ciguatoxin CTX1B: Thiol strategy and detection using a sandwich ELISA. Toxicon 2012, 60, 348–357. [Google Scholar] [CrossRef]
- Tsumuraya, T.; Fujii, I.; Hirama, M. Preparation of anti-ciguatoxin monoclonal antibodies using synthetic haptens: Sandwich ELISA detection of ciguatoxins. J. AOAC Int. 2014, 97, 373–379. [Google Scholar] [CrossRef]
- Manger, R.L.; Leja, L.S.; Lee, S.Y.; Hungerford, J.M.; Wekell, M.M. Tetrazolium-based cell bioassay for neurotoxins active on voltage-sensitive sodium channels: Semiautomated assay for saxitoxins, brevetoxins, and ciguatoxins. Anaytical Biochem. 1993, 214, 190–194. [Google Scholar] [CrossRef]
- Cañete, E.; Diogène, J. Improvements in the use of neuroblastomaxglioma hybrid cells (NG108-15) for the toxic effect quantification of marine toxins. Toxicon 2010, 55, 381–389. [Google Scholar] [CrossRef]
- Fairey, E.R.; Edmunds, J.; Deamer-Melia, N.J.; Glasgow, H., Jr.; Johnson, F.M.; Moeller, P.R.; Burkholder, J.; Ramsdell, J.S. Reporter gene assay for fish-killing activity produced by Pfiesteria piscicida. Environ. Health Perspect. 1999, 107, 711–714. [Google Scholar] [CrossRef]
- Landsberg, J.; Flewelling, L.; Naar, J. Karenia brevis red tides, brevetoxins in the food web, and impacts on natural resources: Decadal advancements. Harmful Algae 2009, 8, 598–607. [Google Scholar] [CrossRef]
- McFarren, E.; Tanabe, H.; Silva, F.; Wilson, W.; Campbell, J.; Lewis, K. The occurrence of a ciguatera-like poison in oysters, clams, and Gymnodinium breve cultures. Toxicon 1965, 3, 111–123. [Google Scholar] [CrossRef]
- Watkins, S.M.; Reich, A.; Fleming, L.E.; Hammond, R. Neurotoxic shellfish poisoning. Mar. Drugs 2008, 6, 431–455. [Google Scholar] [CrossRef]
- Woofter, R.T.; Brendtro, K.; Ramsdell, J.S. Uptake and elimination of brevetoxin in blood of striped mullet (Mugil cephalus) after aqueous exposure to Karenia brevis. Environ. Health Perspect. 2005, 113, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Dechraoui, M.; Bottein, Y.; Tiedeken, J.A.; Persad, R.; Wang, Z.; Granade, H.; Dickey, R.W.; Ramsdell, J.S. Use of two detection methods to discriminate ciguatoxins from brevetoxins: Application to great barracuda from Florida Keys. Toxicon 2005, 46, 261–270. [Google Scholar] [CrossRef]
- Bottein Dechraoui, M.-Y.; Wacksman, J.J.; Ramsdell, J.S. Species selective resistance of cardiac muscle voltage gated sodium channels: Characterization of brevetoxin and ciguatoxin binding sites in rats and fish. Toxicon 2006, 48, 702–712. [Google Scholar] [CrossRef]
- Manger, R.; Woodle, D.; Berger, A.; Dickey, R.W.; Jester, E.; Yasumoto, T.; Lewis, R.; Hawryluk, T.; Hungerford, J. Flow cytometric-membrane potential detection of sodium channel active marine toxins: Application to ciguatoxins in fish muscle and feasibility of automating saxitoxin detection. J. AOAC Int. 2014, 97, 299–306. [Google Scholar] [CrossRef]
- Lewis, R.J. Detection of toxins associated with ciguatera fish poisoning. In Manual on Harmful Marine Microalgae; Hallegraeff, G.M., Anderson, D.M., Cembella, A.D., Eds.; UNESCO: Paris, France, 2003; p. 267. [Google Scholar]
- Yasumoto, T.; Fukui, M.; Sasaki, K.; Sugiyama, K. Determinations of marine toxins in foods. J. AOAC Int. 1995, 78, 574–582. [Google Scholar]
- Campora, C.E.; Hokama, Y.; Tamaru, C.S.; Anderson, B.; Vincent, D. Evaluating the risk of ciguatera fish poisoning from reef fish grown at marine aquaculture facilities in Hawaii. J. World Aquac. Soc. 2010, 41, 61–70. [Google Scholar] [CrossRef]
- Dickey, R.W.; Granade, H.R.; Jester, E.L.E.; Abraham, A.; El Said, K.R.; Plakas, S.M. termination of Caribbean and Pacific ciguatoxins in fish and formulation of regulatory advisory levels. In Proceedings of the Ciguatera and Related Biotoxins Workshop, Noumea, New Caledonia, 27–31 October 2008.
- Darius, H.T.; Ponton, D.; Revel, T.; Cruchet, P.; Ung, A.; Fouc, M.T.; Chinain, M. Ciguatera risk assessment in two toxic sites of French Polynesia using the receptor-binding assay. Toxicon 2007, 50, 612–626. [Google Scholar] [CrossRef]
- Wu, J.J.; Mak, Y.L.; Murphy, M.B.; Lam, J.C.W.; Chan, W.H.; Wang, M.; Chan, L.L.; Lam, P.K.S. Validation of an accelerated solvent extraction liquid chromatography-tandem mass spectrometry method for Pacific ciguatoxin-1 in fish flesh and comparison with the mouse neuroblastoma assay. Anal. Bioanal. Chem. 2011, 400, 3165–3175. [Google Scholar] [CrossRef]
- Bottein Dechraoui, M.Y.; Wang, Z.; Turquet, J.; Chinain, M.; Darius, T.; Cruchet, P.; Radwan, F.F.; Dickey, R.W.; Ramsdell, J.S. Biomonitoring of ciguatoxin exposure in mice using blood collection cards. Toxicon 2005, 46, 243–251. [Google Scholar] [CrossRef]
- Bottein Dechraoui, M.Y.; Wang, Z.; Ramsdell, J.S. Optimization of ciguatoxin extraction method from blood for Pacific ciguatoxin (P-CTX-1). Toxicon 2007, 49, 100–105. [Google Scholar] [CrossRef]
- Ledreux, A.; Ramsdell, J.S. Bioavailability and intravenous toxicokinetic parameters for Pacific ciguatoxin P-CTX-1 in rats. Toxicon 2013, 64, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Bottein Dechraoui, M.Y.; Wang, Z.; Ramsdell, J.S. Toxicokinetics of the ciguatoxin P-CTX-1 in rats after intraperitoneal or oral administration. Toxicology 2011, 284, 1–6. [Google Scholar] [CrossRef]
- O’Toole, A.C.; Dechraoui Bottein, M.-Y.; Danylchuk, A.J.; Ramsdell, J.S.; Cooke, S.J. Linking ciguatera poisoning to spatial ecology of fish: A novel approach to examining the distribution of biotoxin levels in the great barracuda by combining non-lethal blood sampling and biotelemetry. Sci. Total Environ. 2012, 427, 98–105. [Google Scholar] [CrossRef]
- Fraga, S.; Rodríguez, F.; Caillaud, A.; Diogène, J.; Raho, N.; Zapata, M. Gambierdiscus excentricus sp. nov. (Dinophyceae), a benthic toxic dinoflagellate from the Canary Islands (NE Atlantic Ocean). Harmful Algae 2011, 11, 10–22. [Google Scholar]
- Caillaud, A.; de la Iglesia, P.; Barber, E.; Eixarch, H.; Mohammad-Noor, N.; Yasumoto, T.; Diogene, J. Monitoring of dissolved ciguatoxin and maitotoxin using solid-phase adsorption toxin tracking devices: Application to Gambierdiscus pacificus in culture. Harmful Algae 2011, 10, 433–446. [Google Scholar] [CrossRef]
- Shimojo, R.Y.; Iwaoka, W.T. A rapid hemolysis assay for the detection of sodium channel-specific marine toxins. Toxicology 2000, 154, 1–7. [Google Scholar] [CrossRef]
- Holland, W.C.; Litaker, R.W.; Tomas, C.R.; Kibler, S.R.; Place, A.R.; Davenport, E.D.; Tester, P.A. Differences in the toxicity of six Gambierdiscus (Dinophyceae) species measured using an in vitro human erythrocyte lysis assay. Toxicon 2013, 65, 15–33. [Google Scholar] [CrossRef]
- Poli, M.A.; Lewis, R.J.; Dickey, R.W.; Musser, S.M.; Buckner, C.A.; Carpenter, L.G. Identification of Caribbean ciguatoxins as the cause of an outbreak of fish poisoning among US soldiers in Haiti. Toxicon 1997, 35, 733–741. [Google Scholar] [CrossRef]
- Yasumoto, T. Search for ciguatoxin producing clones of Gambierdiscus toxicus on Okinawa coastal reefs. In Abstract book of the Ciguatera and related biotoxins workshop, 27–31 October 2008; Institut de recherche pour le développement (IRD): Noumea, New Caledonia.
- McCall, J.R.; Jacocks, H.M.; Niven, S.C.; Poli, M.A.; Baden, D.G.; Bourdelais, A.J. Development and utilization of a fluorescence-based receptor-binding assay for the site 5 voltage-sensitive sodium channel ligands brevetoxin and ciguatoxin. J. AOAC Int. 2014, 97, 307–315. [Google Scholar] [CrossRef]
- Caillaud, A.; Eixarch, H.; de la Iglesia, P.; Rodriguez, M.; Dominguez, L.; Andree, K.; Diogene, J. Towards the standardisation of the neuroblastoma (neuro-2a) cell-based assay for ciguatoxin-like toxicity detection in fish: Application to fish caught in the Canary Islands. Food Add. Contam. A 2012, 29, 1000–1001. [Google Scholar] [CrossRef]
- Robertson, A.; Garcia, A.C.; Quintana, H.A.F.; Smith, T.B.; Castillo, B.F.; Reale-Munroe, K.; Gulli, J.A.; Olsen, D.A.; Hooe-Rollman, J.I.; Jester, E.L.E.; et al. Invasive lionfish (Pterois volitans): A potential human health threat for ciguatera fish poisoning in tropical waters. Mar. Drugs 2014, 12, 88–97. [Google Scholar]
- Kerbrat, A.-S.; Darius, H.T.; Pauillac, S.; Chinain, M.; Laurent, D. Detection of ciguatoxin-like and paralysing toxins in Trichodesmium spp. from New Caledonia lagoon. from New Caledonia lagoon. Mar. Pollut. Bull. 2010, 61, 360–366. [Google Scholar] [CrossRef]
- Otero, A.; Chapela, M.-J.; Atanassova, M.; Vieites, J.M.; Cabado, A.G. Cyclic Imines: Chemistry and mechanism of action: A review. Chem. Res. Toxicol. 2011, 24, 1817–1829. [Google Scholar] [CrossRef]
- Chatzianastasiou, M.; Katikou, P.; Zacharaki, T.; Papazachariou, A.; McKevitt, A. Cyclic imines, as emerging marine toxins: Chemical properties, distribution, toxicological aspects and detection methods. J. Hell. Vet. Med. Soc. 2011, 62, 240–248. [Google Scholar]
- Cembella, A.; Lewis, N.; Quilliam, M. The marine dinoflagellate Alexandrium ostenfeldii (Dinophyceae) as the causative organism of spirolide shellfish toxins. Phycologia 2000, 39, 67–74. [Google Scholar] [CrossRef]
- Touzet, N.; Franco, J.M.; Raine, R. Morphogenetic diversity and biotoxin composition of Alexandrium (Dinophyceae) in Irish coastal waters. Harmful Algae 2008, 7, 782–797. [Google Scholar] [CrossRef]
- Seki, T.; Satake, M.; Mackenzie, L.; Kaspar, H.F.; Yasumoto, T. Gymnodimine, a new marine toxin of unprecedented structure isolated from New-Zeland oysters and the dinofalgellate Gymnnodinium sp. Tetrahedron Lett. 1995, 36, 7093–7096. [Google Scholar] [CrossRef]
- Rhodes, L.; Smith, K.; Selwood, A.; McNabb, P.; van Ginkel, R.; Holland, P.; Munday, R. Production of pinnatoxins by a peridinoid dinoflagellate isolated from Northland, New Zealand. Harmful Algae 2010, 9, 384–389. [Google Scholar] [CrossRef]
- Nezan, E.; Chomerat, N. Vulcanodinium rugosum gen. nov., sp. nov. (Dinophyceae): A new marine dinoflagellate from the French Mediterranean coast. Cryptogam. Algol. 2011, 32, 3–18. [Google Scholar]
- Torigoe, K.; Murata, M.; Yasumoto, T.; Iwashita, T. Prorocentrolide, a toxic nitrogenous macrocycle from a marine dinoflagellate, Prorocentrum lima. J. Am. Chem. Soc. 1988, 110, 7876–7877. [Google Scholar] [CrossRef]
- Lu, C.K.; Lee, G.H.; Huang, R.; Chou, H.N. Spiro-prorocentrimine, a novel macrocyclic lactone from a benthic Prorocentrum sp. of Taiwan. Tetrahedron Lett. 2001, 42, 1713–1716. [Google Scholar] [CrossRef]
- Miles, C.O.; Wilkins, A.L.; Stirling, D.J.; MacKenzie, A.L. Gymnodimine C, an isomer of gymnodimine B, from Karenia selliformis. J. Agric. Food Chem. 2003, 51, 4838–4840. [Google Scholar] [CrossRef]
- Van Wagoner, R.M.; Misner, I.; Tomas, C.R.; Wright, J.L. Occurrence of 12-methylgymnodimine in a spirolide-producing dinoflagellate Alexandrium peruvianum and the biogenetic implications. Tetrahedron Lett. 2011, 52, 4243–4246. [Google Scholar] [CrossRef]
- Gueret, S.M.; Brimble, M.A. Spiroimine shellfish poisoning (SSP) and the spirolide family of shellfish toxins: Isolation, structure, biological activity and synthesis. Nat. Prod. Rep. 2010, 27, 1350–1366. [Google Scholar] [CrossRef]
- Selwood, A.I.; Miles, C.O.; Wilkins, A.L.; van Ginkel, R.; Munday, R.; Rise, F.; McNabb, P. Isolation, structural determination and acute toxicity of pinnatoxins E, F and G. J. Agric. Food Chem. 2010, 58, 6532–6542. [Google Scholar] [CrossRef]
- Molgó, J.; Girard, E.; Benoit, E. 18 Cyclic Imines: An insight into this emerging group of bioactive marine toxins. In Phycotoxins: Chemistry and Biochemistry; Botana, L.M., Ed.; Wiley-Blackwell: Ames, IA, USA, 2008; pp. 319–335. [Google Scholar]
- Aasen, J.A.; Hardstaff, W.; Aune, T.; Quilliam, M.A. Discovery of fatty acid ester metabolites of spirolide toxins in mussels from Norway using liquid chromatography/tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2006, 20, 1531–1537. [Google Scholar] [CrossRef]
- McCarron, P.; Rourke, W.A.; Hardstaff, W.; Pooley, B.; Quilliam, M.A. Identification of pinnatoxins and discovery of their fatty acid ester metabolites in mussels (Mytilus edulis) from Eastern Canada. J. Agric. Food Chem. 2012, 60, 1437–1446. [Google Scholar] [CrossRef]
- Hu, T.; deFreitas, A.S.; Curtis, J.M.; Oshima, Y.; Walter, J.A.; Wright, J.L. Isolation and structure of prorocentrolide B, a fast-acting toxin from Prorocentrum maculosum. J. Nat. Prod. 1996, 59, 1010–1014. [Google Scholar] [CrossRef]
- MacKenzie, L.A.; Haywood, A.J.; Adamson, J.; Truman, P.; Till, D.; Seki, T.; Satake, M.; Yasumoto, T. Gymnodimine contamination in shellfish in New Zealand. In Harmful and Toxic Algal Blooms; Yasumoto, T., Oshima, Y., Fukuyo, Y., Eds.; The Intergovernmental Oceanographic Commission of UNESCO: Paris, France, 1996; pp. 97–100. [Google Scholar]
- Uemura, D.; Chou, T.; Haino, T.; Nagatsu, A.; Fukuzawa, S.; Zheng, S.Z.; Chen, H.S. Pinnatoxin A: A toxic amphoteric macrocycle from the okinawan bivalve Pinna muricata. J. Am. Chem. Soc. 1995, 117, 1155–1156. [Google Scholar]
- Bourne, Y.; Radic, Z.; Aráoz, R.; Talley, T.T.; Benoit, E.; Servent, D.; Taylor, P.; Molgó, J.; Marchot, P. Structural determinants in phycotoxins and AChBP conferring high affinity binding and nicotinic AChR antagonism. Proc. Natl. Acad. Sci. USA 2010, 107, 6076–6081. [Google Scholar] [CrossRef]
- Stivala, C.E.; Benoit, E.; Araoz, R.; Servent, D.; Novikov, A.; Molgo, J.; Zakarian, A. Synthesis and biology of cyclic imine toxins, an emerging class of potent, globally distributed marine toxins. Nat. Prod. Rep. 2014. [Google Scholar] [CrossRef]
- Pigozzi, S.; Bianchi, L.; Boschetti, L.; Cangini, M.; Ceredi, A.; Magnani, F.; Milandri, A.; Montanari, S.; Pompei, M.; Riccardi, E. First evidence of spirolide accumulation in northwestern Adriatic shellfish. In Proceedings of the 12th ICHA, 4–8 September 2006; ISSHA and IOC of UNESCO: Copenhagen, Denmark, 2008; pp. 319–322. [Google Scholar]
- Kharrat, R.; Servent, D.; Girard, E.; Ouanounou, G.; Amar, M.; Marrouchi, R.; Benoit, E.; Molgo, J. The marine phycotoxin gymnodimine targets muscular and neuronal nicotinic acetylcholine receptor subtypes with high affinity. J. Neurochem. 2008, 107, 952–963. [Google Scholar]
- Dragunow, M.; Trzoss, M.; Brimble, M.A.; Cameron, R.; Beuzenberg, V.; Holland, P.; Mountfort, D. Investigations into the cellular actions of the shellfish toxin gymnodimine and analogues. Environ. Toxicol. Pharmacol. 2005, 20, 305–312. [Google Scholar] [CrossRef]
- Geiger, M.; Desanglois, G.; Hogeveen, K.; Fessard, V.; Lepretre, T.; Mondeguer, F.; Guitton, Y.; Herve, F.; Sechet, V.; Grovel, O.; et al. Cytotoxicity, fractionation and dereplication of extracts of the Dinoflagellate Vulcanodinium rugosum, a producer of pinnatoxin G. Mar. Drugs 2013, 11, 3350–3371. [Google Scholar] [CrossRef] [Green Version]
- Hellyer, S.D.; Selwood, A.I.; van Ginkel, R.; Munday, R.; Sheard, P.; Miles, C.O.; Rhodes, L.; Kerr, D.S. In vitro labelling of muscle type nicotinic receptors using a fluorophore-conjugated pinnatoxin F derivative. Toxicon 2014, 87, 17–25. [Google Scholar] [CrossRef]
- Tatters, A.O.; Muhlstein, H.I.; Tomas, C.R. The hemolytic activity of Karenia selliformis and two clones of Karenia brevis throughout a growth cycle. J. Appl. Phycol. 2010, 22, 435–442. [Google Scholar] [CrossRef]
- Eschbach, E.; Scharsack, J.P.; John, U.; Medlin, L.K. Improved erythrocyte lysis assay in microtitre plates for sensitive detection and efficient measurement of haemolytic compounds from ichthyotoxic algae. J. Appl. Toxicol. 2001, 21, 513–519. [Google Scholar] [CrossRef]
- Vilariño, N.; Fonfria, E.S.; Molgo, J.; Araoz, R.; Botana, L.M. Detection of gymnodimine-A and 13-desmethyl C spirolide phycotoxins by fluorescence polarization. Anal. Chem. 2009, 81, 2708–2714. [Google Scholar] [CrossRef]
- Fonfria, E.S.; Vilarino, N.; Molgo, J.; Araoz, R.; Otero, P.; Espina, B.; Carmen Louzao, M.; Alvarez, M.; Botana, L.M. Detection of 13,19-didesmethyl C spirolide by fluorescence polarization using Torpedo electrocyte membranes. Anal. Biochem. 2010, 403, 102–107. [Google Scholar] [CrossRef]
- Fonfria, E.S.; Vilarino, N.; Espina, B.; Carmen Louzao, M.; Alvarez, M.; Molgo, J.; Araoz, R.; Botana, L.M. Feasibility of gymnodimine and 13-desmethyl C spirolide detection by fluorescence polarization using a receptor-based assay in shellfish matrixes. Anal. Chim. Acta 2010, 657, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Otero, P.; Alfonso, A.; Alfonso, C.; Araoz, R.; Molgo, J.; Vieytes, M.R.; Botana, L.M. First direct fluorescence polarization assay for the detection and quantification of spirolides in mussel samples. Anal. Chim. Acta 2011, 701, 200–208. [Google Scholar] [CrossRef]
- Rodriguez, L.P.; Vilarino, N.; Molgo, J.; Araoz, R.; Antelo, A.; Vieytes, M.R.; Botana, L.M. Solid-phase receptor-based assay for the detection of cyclic imines by chemiluminescence, fluorescence, or colorimetry. Anal. Chem. 2011, 83, 5857–5863. [Google Scholar] [CrossRef]
- Rodriguez, L.P.; Vilarino, N.; Molgo, J.; Araoz, R.; Botana, L.M. High-throughput receptor-based assay for the detection of spirolides by chemiluminescence. Toxicon 2013, 75, 35–43. [Google Scholar] [CrossRef]
- Araoz, R.; Ramos, S.; Pelissier, F.; Guerineau, V.; Benoit, E.; Vilarino, N.; Botana, L.M.; Zakarian, A.; Molgo, J. Coupling the Torpedo microplate-receptor binding assay with mass spectrometry to detect cyclic imine neurotoxins. Anal. Chem. 2012, 84, 10445–10453. [Google Scholar] [CrossRef]
- Rodriguez, L.P.; Vilarino, N.; Molgo, J.; Araoz, R.; Carmen Louzao, M.; Taylor, P.; Talley, T.; Botana, L.M. Development of a solid-phase receptor-based assay for the detection of cyclic imines using a microsphere-flow cytometry system. Anal. Chem. 2013, 85, 2340–2347. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Sugimoto, A.; Takai, A.; Yasumoto, T. Effects of specific modifications of several hydroxyls of tetrodotoxin on its affinity to rat brain membrane. J. Pharmacol. Exp. Ther. 1999, 289, 1688–1696. [Google Scholar]
- Bane, V.; Lehane, M.; Dikshit, M.; O’Riordan, A.; Furey, A. Tetrodotoxin: Chemistry, toxicity, source, distribution and detection. Toxins 2014, 6, 693–755. [Google Scholar] [CrossRef]
- Tahara, Y.; Hirata, Y. Studies on the puffer fish toxin. J. Pharm. Soc. 1909, 29, 587–625. [Google Scholar]
- Yokoo, A. Chemical studies on pufferfish toxin (3)-separation of spheroidine. J. Chem. Soc. Jpn. 1950, 71, 590–592. [Google Scholar]
- Tsuda, K.; Tachikawa, R.; Sakai, K.; Tamura, C.; Amakasu, O.; Kawamura, M.; Ikuma, S. On the structure of tetrodotoxin. Chem. Pharm. Bull. 1964, 12, 642–645. [Google Scholar] [CrossRef]
- Woodward, R.; Gougoutas, J.Z. The structure of tetrodotoxin. Pure Appl. Chem. 1964, 9, 49–74. [Google Scholar]
- Goto, T.; Kishi, Y.; Takahashi, S.; Hirata, Y. Tetrodotoxin. Tetrahedron 1965, 21, 2059–2088. [Google Scholar] [CrossRef]
- Noguchi, T.; Arakawa, O.; Takatani, T. TTX accumulation in pufferfish. Comp. Biochem. Physiol. D 2006, 1, 145–152. [Google Scholar]
- Chau, R.; Kalaitzis, J.A.; Neilan, B.A. On the origins and biosynthesis of tetrodotoxin. Aquat. Toxicol. 2011, 104, 61–72. [Google Scholar] [CrossRef]
- Pratheepa, V.; Vasconcelos, V. Microbial diversity associated with tetrodotoxin production in marine organisms. Environ. Toxicol. Pharmacol. 2013, 36, 1046–1054. [Google Scholar] [CrossRef]
- Rodriguez, P.; Alfonso, A.; Vale, C.; Alfonso, C.; Vale, P.; Tellez, A.; Botana, L.M. First toxicity report of tetrodotoxin and 5,6,11-trideoxyTTX in the trumpet shell Charonia lampas lampas in Europe. Anal. Chem. 2008, 80, 5622–5629. [Google Scholar] [CrossRef]
- Bentur, Y.; Ashkar, J.; Lurie, Y.; Levy, Y.; Azzam, Z.S.; Litmanovich, M.; Golik, M.; Gurevych, B.; Golani, D.; Eisenman, A. Lessepsian migration and tetrodotoxin poisoning due to Lagocephalus sceleratus in the eastern Mediterranean. Toxicon 2008, 52, 964–968. [Google Scholar] [CrossRef]
- Silva, M.; Azevedo, J.; Rodriguez, P.; Alfonso, A.; Botana, L.M.; Vasconcelos, V. New gastropod vectors and tetrodotoxin potential expansion in temperate waters of the Atlantic Ocean. Mar. Drugs 2012, 10, 712–726. [Google Scholar] [CrossRef]
- Kawatsu, K.; Hamano, Y.; Yoda, T.; Terano, Y.; Shibata, T. Rapid and highly sensitive enzyme immunoassay for quantitative determination of tetrodotoxin. Jpn. J. Med. Sci. Biol. 1997, 50, 133–150. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, Y.; Pan, F.; Liu, Z.; Wang, Z. Identification of tetrodotoxin antigens and a monoclonal antibody. Food Chem. 2009, 112, 582–586. [Google Scholar] [CrossRef]
- Raybould, T.J.G.; Bignami, G.S.; Inouye, L.K.; Simpson, S.B.; Byrnes, J.B.; Grothaus, P.G.; Vann, D.C. A monoclonal antibody-based immunoassay for detecting tetrodotoxin in biological samples. J. Clin. Lab. Anal. 1992, 6, 65–72. [Google Scholar] [CrossRef]
- Neagu, D.; Micheli, L.; Palleschi, G. Study of a toxin-alkaline phosphatase conjugate for the development of an immunosensor for tetrodotoxin determination. Anal. Bioanal. Chem. 2006, 385, 1068–1074. [Google Scholar] [CrossRef]
- Tao, J.; Wei, W.; Nan, L.; Lei, L.; Hui, H.; Fen, G.; Jun, L.; Jing, Z.; Rong, J. Development of competitive indirect ELISA for the detection of tetrodotoxin and a survey of the distribution of tetrodotoxin in the tissues of wild puffer fish in the waters of south-east China. Food Add. Contam. 2010, 27, 1589–1597. [Google Scholar] [CrossRef]
- Stokes, A.N.; Williams, B.L.; French, S.S. An improved competitive inhibition enzymatic immunoassay method for tetrodotoxin quantification. Biol. Proced. Online 2012, 14, 1–5. [Google Scholar]
- Wang, R.; Huang, A.; Liu, L.; Xiang, S.; Li, X.; Ling, S.; Wang, L.; Lu, T.; Wang, S. Construction of a single chain variable fragment antibody (scFv) against tetrodotoxin (TTX) and its interaction with TTX. Toxicon 2014, 83, 22–34. [Google Scholar] [CrossRef]
- Kreuzer, M.P.; Pravda, M.; O’Sullivan, C.K.; Guilbault, G.G. Novel electrochemical immunosensors for seafood toxin analysis. Toxicon 2002, 40, 1267–1274. [Google Scholar] [CrossRef]
- Yu, C.-H.; Yu, C.-F.; Tam, S.; Hoi-Fu Yu, P. Rapid screening of tetrodotoxin in urine and plasma of patients with puffer fish poisoning by HPLC with creatinine correction. Food Add. Contam. 2010, 27, 89–96. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, Y.; Lu, S.; Ren, H.; Li, Z.; Zhang, Y.; Pan, F.; Liu, W.; Zhang, J.; Liu, Z. Gold nanoparticle probe-based immunoassay as a new tool for tetrodotoxin detection in puffer fish tissues. Sens. Actuators B 2010, 146, 368–372. [Google Scholar] [CrossRef]
- Thattiyaphong, A.; Unahalekhaka, J.; Mekha, N.; Nispa, W.; Kluengklangdon, P.; Rojanapantip, L. Efficiency of a rapid test for detection of tetrodotoxin in puffer fish. J. Immunoass. Immunochem. 2013, 35, 111–119. [Google Scholar]
- Yakes, B.J.; Deeds, J.; White, K.; DeGrasse, S.L. Evaluation of surface plasmon resonance biosensors for detection of tetrodotoxin in food matrices and comparison to analytical methods. J. Agric. Food Chem. 2011, 59, 839–846. [Google Scholar]
- Taylor, A.D.; Ladd, J.; Etheridge, S.; Deeds, J.; Hall, S.; Jiang, S. Quantitative detection of tetrodotoxin (TTX) by a surface plasmon resonance (SPR) sensor. Sens. Actuators B 2008, 130, 120–128. [Google Scholar] [CrossRef]
- Taylor, A.D.; Vaisocherová, H.; Deeds, J.; DeGrasse, S.; Jiang, S. Tetrodotoxin detection by a surface plasmon resonance sensor in pufferfish matrices and urine. J. Sens. 2011. [Google Scholar] [CrossRef]
- Vaisocherova, H.; Taylor, A.D.; Jiang, S.; Hegnerova, K.; Vala, M.; Homola, J.; Yakes, B.J.; Deeds, J.; DeGrasse, S. Surface plasmon resonance biosensor for determination of tetrodotoxin: Prevalidation study. J. AOAC Int. 2011, 94, 596–604. [Google Scholar]
- Campbell, K.; Barnes, P.; Haughey, S.; Higgins, C.; Kawatsu, K.; Vasconcelos, V.; Elliott, C. Development and single laboratory validation of an optical biosensor assay for tetrodotoxin detection as a tool to combat emerging risks in European seafood. Anal. Bioanal. Chem. 2013, 405, 7753–7763. [Google Scholar] [CrossRef]
- Yakes, B.J.; Kanyuck, K.M.; DeGrasse, S.L. First report of a direct surface plasmon resonance immunosensor for a small molecule seafood toxin. Anal. Chem. 2014, 86, 9251–9255. [Google Scholar] [CrossRef] [PubMed]
- Yakes, B.J.; Etheridge, S.M.; Mulvaney, S.P.; Tamanaha, C.R. Fluidic force discrimination assays: A new technology for tetrodotoxin detection. Mar. Drugs 2010, 8, 565–576. [Google Scholar] [CrossRef]
- Kogure, K.; Tamplin, M.L.; Simidu, U.; Colwell, R.R. A tissue culture assay for tetrodotoxin, saxitoxin and related toxins. Toxicon 1988, 26, 191–197. [Google Scholar] [CrossRef]
- Narahashi, T.; Moore, J.W.; Scott, W.R. Tetrodotoxin blockage of sodium conductance increase in lobster giant axons. J. Gen. Physiol. 1964, 47, 965–974. [Google Scholar] [CrossRef]
- Catterall, W.A.; Nirenberg, M. Sodium uptake associated with activation of action potential ionophores of cultured neuroblastoma and muscle cells. Proc. Natl. Acad. Sci. USA 1973, 70, 3759–3763. [Google Scholar] [CrossRef]
- Do, H.; Hamasaki, K.; Ohwada, K.; Simidu, U.; Noguchi, T.; Shida, Y.; Kogure, K. Presence of tetrodotoxin and tetrodotoxin-producing bacteria in freshwater sediments. Appl. Environ. Microbiol. 1993, 59, 3934–3937. [Google Scholar]
- Wu, Z.; Xie, L.; Xia, G.; Zhang, J.; Nie, Y.; Hu, J.; Wang, S.; Zhang, R. A new tetrodotoxin-producing actinomycete, Nocardiopsis dassonvillei, isolated from the ovaries of puffer fish Fugu rubripes. Toxicon 2005, 45, 851–859. [Google Scholar] [CrossRef]
- Gallacher, S.; Birkbeck, T. A tissue culture assay for direct detection of sodium channel blocking toxins in bacterial culture supernates. FEMS Microbiol. Lett. 1992, 92, 101–107. [Google Scholar] [CrossRef]
- Hamasaki, K.; Kogure, K.; Ohwada, K. A biological method for the quantitative measurement of tetrodotoxin (TTX): Tissue culture bioassay in combination with a water-soluble tetrazolium salt. Toxicon 1996, 34, 490–495. [Google Scholar] [CrossRef]
- Hamasaki, K.; Kogure, K.; Ohwada, K. An improved method of tissue culture bioassay for tetrodotoxin. Fish. Sci. 1996b, 62, 825–829. [Google Scholar]
- Kodama, M.; Ogata, T.; Noguchi, T.; Maruyama, J.; Hashimoto, K. Occurrence of saxitoxin and other toxins in the liver of the pufferfish Takifugu pardalis. Toxicon 1983, 21, 897–900. [Google Scholar] [CrossRef]
- Jang, J.; Yotsu-Yamashita, M. Distribution of tetrodotoxin, saxitoxin, and their analogs among tissues of the puffer fish Fugu pardalis. Toxicon 2006, 48, 980–987. [Google Scholar] [CrossRef]
- Jen, H.-C.; Lin, S.-J.; Lin, S.-Y.; Huang, Y.-W.; Liao, I.; Arakawa, O.; Hwang, D.-F. Occurrence of tetrodotoxin and paralytic shellfish poisons in a gastropod implicated in food poisoning in southern Taiwan. Food Add. Contam. 2007, 24, 902–909. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Mebs, D. Occurrence of 11-oxotetrodotoxin in the red-spotted newt, Notophthalmus viridescens, and further studies on the levels of tetrodotoxin and its analogues in the newt’s efts. Toxicon 2003, 41, 893–897. [Google Scholar] [CrossRef]
- Cheun, B.; Endo, H.; Hayashi, T.; Nagashima, Y.; Watanabe, E. Development of an ultra high sensitive tissue biosensor for determination of swellfish poisoning, tetrodotoxin. Biosens. Bioelectron. 1996, 11, 1185–1191. [Google Scholar] [CrossRef]
- Pancrazio, J.J.; Gray, S.A.; Shubin, Y.S.; Kulagina, N.; Cuttino, D.S.; Shaffer, K.M.; Eisemann, K.; Curran, A.; Zim, B.; Gross, G.W. A portable microelectrode array recording system incorporating cultured neuronal networks for neurotoxin detection. Biosens. Bioelectron. 2003, 18, 1339–1347. [Google Scholar] [CrossRef]
- Charkhkar, H.; Knaack, G.L.; Gnade, B.E.; Keefer, E.W.; Pancrazio, J.J. Development and demonstration of a disposable low-cost microelectrode array for cultured neuronal network recording. Sens. Actuators B 2012, 161, 655–660. [Google Scholar] [CrossRef]
- Mohan, D.K.; Molnar, P.; Hickman, J.J. Toxin detection based on action potential shape analysis using a realistic mathematical model of differentiated NG108-15 cells. Biosens. Bioelectron. 2006, 21, 1804–1811. [Google Scholar] [CrossRef]
- Weiser, T. A novel toxicity-based assay for the identification of modulators of voltage-gated Na+ channels. J. Neurosci. Methods 2004, 137, 79–85. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reverté, L.; Soliño, L.; Carnicer, O.; Diogène, J.; Campàs, M. Alternative Methods for the Detection of Emerging Marine Toxins: Biosensors, Biochemical Assays and Cell-Based Assays. Mar. Drugs 2014, 12, 5719-5763. https://doi.org/10.3390/md12125719
Reverté L, Soliño L, Carnicer O, Diogène J, Campàs M. Alternative Methods for the Detection of Emerging Marine Toxins: Biosensors, Biochemical Assays and Cell-Based Assays. Marine Drugs. 2014; 12(12):5719-5763. https://doi.org/10.3390/md12125719
Chicago/Turabian StyleReverté, Laia, Lucía Soliño, Olga Carnicer, Jorge Diogène, and Mònica Campàs. 2014. "Alternative Methods for the Detection of Emerging Marine Toxins: Biosensors, Biochemical Assays and Cell-Based Assays" Marine Drugs 12, no. 12: 5719-5763. https://doi.org/10.3390/md12125719