Marine Pseudovibrio sp. as a Novel Source of Antimicrobials

 ,
,
Abstract
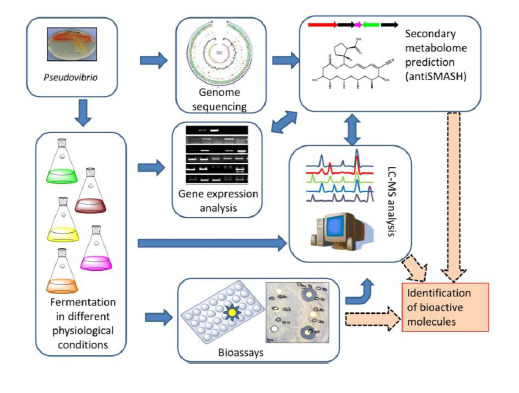
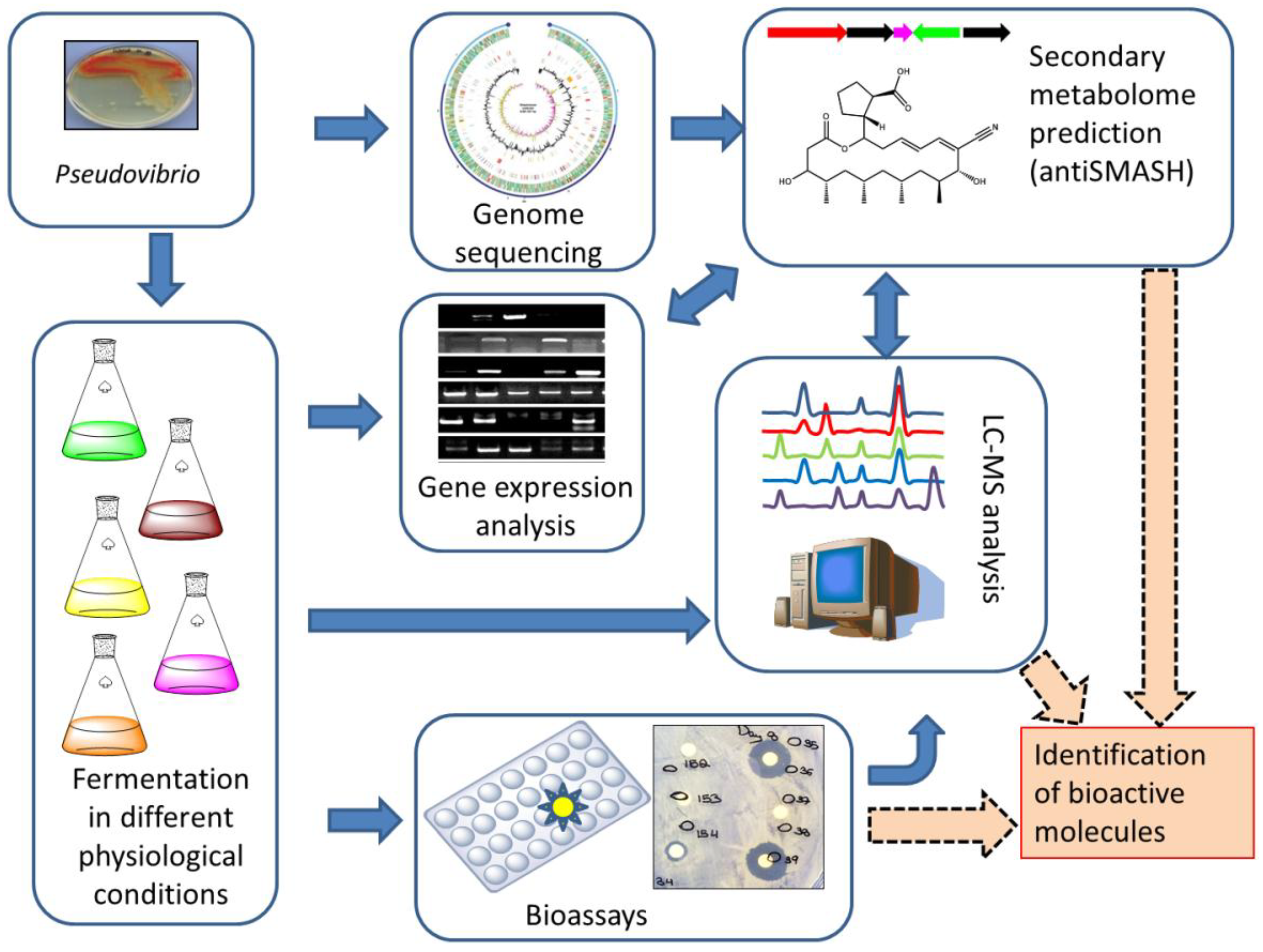
:
1. Introduction
2. Pseudovibrio sp.
2.1. Characterisation of the Genus Pseudovibrio

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | P. denitrificans | P. ascidiaceicola | P. japonicas | P. axinellae |
|---|---|---|---|---|
| Gram reaction | Negative | Negative | Negative | Negative |
| Oxidase | Positive | Positive | Positive | Positive |
| Catalase | Positive | Positive | Positive | Positive |
| Cell shape | Straight/curved rod (exponential-early stationary phase) | Straight/curved rod (exponential-early stationary phase) Predominantly cocci (late stationary phase) | Rod shaped | Rod shaped |
| Aerobic/anaerobic | Facultatively anaerobic | Facultatively anaerobic | Facultatively anaerobic | Facultatively anaerobic |
| Motile | Lateral or subpolar flagella | Subpolar flagella | Lateral or subpolar flagella | Subpolar flagella |
| Temperature tolerances | Grow at 20 °C–35 °C but not at 4 °C or 45 °C | Grow at 10 °C–30 °C | Grow at 15 °C–30 °C but not at 8 °C or 35 °C | Grow at 10 °C–30 °C |
| Halophilic | Grow at 2%–6% NaCl | Grow at 3%–5% NaCl | Grow at 1%–6% NaCl | Grow at 2%–4% NaCl |
| Substrates utilized for growth | Galactose, glucose, mannose, sucrose and trehalose but not d-arabinose, cellobiose, dulcitol, glycerol, inositol, lactose, mannitol, sorbitol or xylose. | Dextrin, Tween 80, N-acetyl-d-galactosamine, N-acetyl-d-glucosamine, l-fucose, d-galactose, α-d-glucose, myo-inositol, maltose, d-mannose, d-meli-biose, d-raffinose, sucrose, d-trehalose, turanose, d-gluconic acid, β-hydroxybutyric acid, dl-lactic acid, succinic acid, l-alanine, l-alanyl glycine, l-glutamic acid, glycyl-l-aspartic acid, glycyl-l-glutamic acid, hydroxyl-l-proline, l-proline, l-serine, inosine, uridine, thymidine, 2-amino ethanol, glycerol, d-glucose 6-phosphate. | d-glucose, maltose, d-mannose and sucrose, d-fructose, d-galactose, α-lactose or d-xylose negative results for arginine dihydrolase, lysine decarboxylase, ornithine decarboxylase, lipase (C4), cysteine arylamidase, chymotrypsin, α-galactosidase, β-glucuronidase, α-glucosidase, β-glucosidase, N-acetyl-β-glucosamidase, α-mannosidase and α-fucosidase utilisation. | Dextrin, Tween 40, Tween 80, l-fucose, α-d-glucose, myo-inositol, maltose, d-mannose, raffinose, sucrose, trehalose, turanose, pyruvic acid methyl ester, d-gluconic acid, d-glucuronic acid, β-hydroxybutyric acid, α-ketobutyric acid, α-ketoglutaric acid, dl-lactic acid, succinic acid, bromosuccinic acid, d-alanine, l-alanine, l-alanyl glycine, l-aspar-agine, l-aspartic acid, l-glutamic acid, glycyl l-aspartic acid, glycyl l-glut-amic acid, hydroxyl-l-proline, l-proline, l-serine, inosine, uridine, thymidine, 2-aminoethanol, glycerol. |
2.2. Bioactivity of the genus Pseudovibrio



3. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- United State Department of Health and Human Services. Antibiotic Resistance Threats in the Unites States; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2013. [Google Scholar]
- ECDC. Annual Epidemiological Report 2012; European Center for Disease Prevention and Control: Tomtebodavägen 11a, 17183, Stockholm, Sweden. Available online: http://www.ecdc.europa.eu/en/publications/Annual-Epidemiological-Report-2012.pdf (accessed on 14 February 2014).
- Kanerva, M.; Ollgren, J.; Hakanen, A.J.; Lyytikäinen, O. Estimating the burden of healthcare-associated infections caused by selected multidrug-resistant bacteria Finland, 2010. Antimicrob. Resist. Infect. Control 2012, 1. [Google Scholar] [CrossRef]
- Infectious Diseases Society of America. The 10 × '20 initiative: Pursuing a global commitment to develop 10 new antibacterial drugs by 2020. Clin. Infect. Dis. 2020, 50, 1081–1083. [Google Scholar]
- Talbot, G.H.; Bradley, J.; Edwards, J.E.; Gilbert, D.; Scheld, M.; Bartlett, J.G. Bad bugs need drugs: An update on the development pipeline from the antimicrobial availability task force of the infectious diseases society of America. Clin. Infect. Dis. 2006, 42, 657–668. [Google Scholar] [CrossRef] [PubMed]
- Penesyan, A.; Kjelleberg, S.; Egan, S. Development of novel drugs from marine surface associated microorganisms. Mar. Drugs 2010, 8, 438–459. [Google Scholar] [CrossRef] [PubMed]
- Penesyan, A.; Marshall-Jones, Z.; Holmstrom, C.; Kjelleberg, S.; Egan, S. Antimicrobial activity observed among cultured marine epiphytic bacteria refects their potential as a source of newdrugs. Fed. Eur. Microbiol. Soc. 2009, 69, 113–124. [Google Scholar]
- European Science Foundation. A New Vision and Strategy for Europe; Marine Board: Ostend, Belgium, 2012. [Google Scholar]
- Lam, K.S. Discovery of novel metabolites from marine actinomycetes. Curr. Opin. Microbiol. 2006, 9, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Prudhomme, J.; McDaniel, E.; Ponts, N.; Bertani, S.; Fenical, W.; Jensen, P.; le Roch, K. Marine actinomycetes: A new source of compounds against the human malaria parasite. PLoS One 2008. [Google Scholar] [CrossRef]
- Hentschel, U.; Schmid, M.; Wagner, M.; Fieseler, L.; Gernert, C.; Hacker, J. Isolation and phylogenetic analysis of bacteria with antimicrobial activities from the Mediterranean sponges Aplysina aerophoba and Aplysina cavernicola. FEMS Microbiol. Ecol. 2001, 35, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Viegelmann, C.; Margassery, L.; Kennedy, J.; Zhang, T.; OBrien, C.; O’Gara, F.; Morrisssey, J.; Dobson, A.; Edrada-Ebel, R. Metabolomic profiling and genomic study of a marine sponge-associated Streptomyces sp. Mar. Drugs 2014, 12, 3323–3351. [Google Scholar] [CrossRef]
- Prieto, M.L.; O’Sullivan, L.; Tan, S.P.; McLoughlin, P.; Hughes, H.; O’Connor, P.M.; Cotter, P.D.; Lawlor, P.G.; Gardiner, G.E. Assessment of the bacteriocinogenic potential of marine bacteria reveals lichenicidin production by seaweed-derived Bacillus spp. Mar. Drugs 2012, 10, 2280–2299. [Google Scholar] [CrossRef]
- Desriac, F.; Jégou, C.; Balnois, E.; Brillet, B.; le Chevalier, P.; Fleury, Y. Antimicrobial peptides from marine proteobacteria. Mar. Drugs 2013, 11, 3632–3660. [Google Scholar] [PubMed]
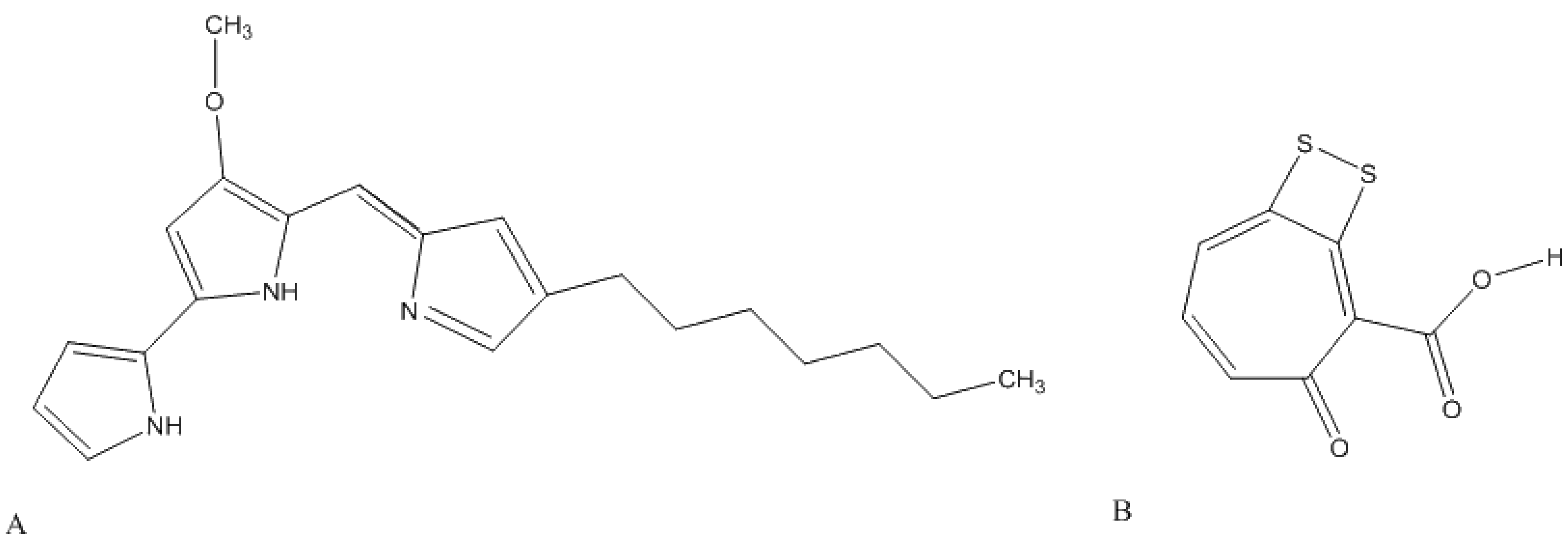
- Sertan-de Guzman, A.A.; Bernardo, R.Z.; Neilan, B.A.; Elardo, S.P.; Mangalindan, G.C.; Tasdemir, D.; Ireland, C.M.; Barraquio, W.L.; Concepcion, G.P. Pseudovibrio denitrificans strain Z143-1, a heptylprodigiosin-producing bacterium isolated from a Philippine tunicate. FEMS Microbiol. Lett. 2007, 277, 188–196. [Google Scholar]
- Geng, H.F.; Belas, R. Expression of tropodithietic acid biosynthesis is controlled by a novel autoinducer. J. Bacteriol. 2010, 192, 4377–4387. [Google Scholar] [CrossRef] [PubMed]
- Thiel, V.; Imhoff, J.F. Phylogenetic identification of bacteria with antimicrobial activities isolated from Mediterranean sponges. Biomol. Eng. 2003, 20, 421–423. [Google Scholar] [CrossRef] [PubMed]
- Muscholl-Silberhorn, A.; Thiel, V.; Imhoff, J.F. Abundance and bioactivity of cultured sponge-associated bacteria from the mediterranean sea. Microb. Ecol. 2008, 55, 94–106. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, J.; Baker, P.; Piper, C.; Cotter, P.D.; Walsh, M.; Mooij, M.J.; Bourke, M.B.; Rea, M.C.; O’Connor, P.M.; Ross, R.P.; et al. Isolation and analysis of bacteria with antimicrobial activities from the marine sponge Haliclona simulans collected from Irish waters. Mar. Biotechnol. 2009, 11, 384–396. [Google Scholar] [CrossRef] [PubMed]
- Fukunaga, Y.; Kurahashi, M.; Tanaka, K.; Yanavfbgi, K.; Yokota, A.; Harayama, S. Pseudovibrio ascidiaceicola sp. nov., isolated from ascidians (sea squirts). Int. J. Syst. Evol. Microbiol. 2006, 56, 343–347. [Google Scholar] [CrossRef] [PubMed]
- Vizcaino, M.; Moeller, P.; Morris, P. The Potential for the Marine Bacterium Pseudovibrio denitrificans to Produce Novel Antibiotics. Available online: https://www.musc.edu/mbes-ljg/MBESOpenHouse/2010/abstracts/Vizcaino-SROH_July_16_2010.pdf (accessed on 30 March 2014).
- Shieh, W.Y.; Lin, Y.T.; Jean, W.D. Pseudovibrio denitrificans gen. nov., sp. nov., a marine, facultatively anaerobic, fermentative bacterium capable of denitrification. Int. J. Syst. Evol. Microbiol. 2004, 54, 2307–2312. [Google Scholar] [CrossRef] [PubMed]
- Penesyan, A.; Tebben, J.; Lee, M.; Thomas, T.; Kjelleberg, S.; Harder, T.; Egan, S. Identification of the antibacterial compound produced by the marine epiphytic bacterium Pseudovibrio sp. D323 and related sponge-associated bacteria. Mar. Drugs 2011, 9, 1391–1402. [Google Scholar] [CrossRef]
- Enticknap, J.J.; Kelly, M.; Peraud, O.; Hill, R. Characterization of a culturable alphaproteobacterial symbiont common to many marine sponges and evidence for vertical transmission via sponge larvae. Appl. Environ. Microbiol. 2006, 72, 3724–3732. [Google Scholar] [CrossRef] [PubMed]
- Graca, A.P.; Bondoso, J.; Gaspar, H.; Xavier, J.R.; Monteiro, M.C.; de la Cruz, M.; Oves-Costales, D.; Vicente, F.; Lage, O.M. Antimicrobial Activity of Heterotrophic Bacterial Communities from the Marine Sponge Erylus discophorus (Astrophorida, Geodiidae). PLoS One 2013, 8, e78992. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.; Radax, R.; Seger, D.; Wagner, M. Sponge-associated microorganisms: Evolution, ecology, and biotechnological potential. Microbiol. Mol. Biol. Rev. 2007, 71, 295–347. [Google Scholar] [CrossRef] [PubMed]
- Esteves, A.I.; Hardoim, C.C.; Xavier, J.R.; Goncalves, J.M.; Costa, R. Molecular richness and biotechnological potential of bacteria cultured from Irciniidae sponges in the north-east Atlantic. FEMS Microbiol. Ecol. 2013, 85, 519–536. [Google Scholar] [CrossRef]
- Santos, O.C.; Pontes, P.V.; Santos, J.F.; Muricy, G.; Giambiagi-deMarval, M.; Laport, M.S. Isolation, characterization and phylogeny of sponge-associated bacteria with antimicrobial activities from Brazil. Res. Microbiol. 2010, 161, 604–612. [Google Scholar] [CrossRef] [PubMed]
- Margassery, L.M.; Kennedy, J.; O’Gara, F.; Dobson, A.D.; Morrissey, J.P. Diversity and antibacterial activity of bacteria isolated from the coastal marine sponges Amphilectus fucorum and Eurypon major. Lett. Appl. Microbiol. 2012, 55, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Hosoya, S.; Yokota, A. Pseudovibrio japonicus sp. nov., isolated from coastal seawater in Japan. Int. J. Syst. Evol. Microbiol. 2007, 57, 1952–1955. [Google Scholar] [CrossRef] [PubMed]
- O’Halloran, J.A.; Barbosa, T.M.; Morrissey, J.P.; Kennedy, J.; Dobson, A.D.; O’Gara, F. Pseudovibrio axinellae sp. nov., isolated from an Irish marine sponge. Int. J. Syst. Evol. Microbiol. 2013, 63, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, J.; Codling, C.E.; Jones, B.V.; Dobson, A.D.; Marchesi, J.R. Diversity of microbes associated with the marine sponge, Haliclona simulans, isolated from Irish waters and identification of polyketide synthase genes from the sponge metagenome. Environ. Microbiol. 2008, 10, 1888–1902. [Google Scholar] [CrossRef] [PubMed]
- O’Halloran, J.A.; Barbosa, T.M.; Morrissey, J.P.; Kennedy, J.; O’Gara, F.; Dobson, A.D. Diversity and antimicrobial activity of Pseudovibrio spp. from Irish marine sponges. J. Appl. Microbiol. 2011, 110, 1495–1508. [Google Scholar] [CrossRef] [PubMed]
- Flemer, B.; Kennedy, J.; Margassery, L.M.; Morrissey, J.P.; O’Gara, F.; Dobson, A.D. Diversity and antimicrobial activities of microbes from two Irish marine sponges, Suberites carnosus and Leucosolenia sp. J. Appl. Microbiol. 2011, 112, 289–301. [Google Scholar] [CrossRef]
- Bruhn, J.B.; Gram, L.; Belas, R. Production of antibacterial compounds and biofilm formation by Roseobacter species are influenced by culture conditions. Appl. Environ. Microbiol. 2007, 73, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Gómez, B.; Fernàndez-Guerra, A.; Casamayor, E.O.; González, J. M.; Pedrós-Alió, C.; Acinas, S.G. Patterns and architecture of genomic islands in marine bacteria. BMC Genomics 2012, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.Q. Genome-based studies of marine of microorganisms to maximize the diversity of natural products discovery for medical treatments. Evid.-Based Complement. Altern. Med. 2011, 2011. [Google Scholar] [CrossRef]
- Bondarev, V.; Rcchter, M.; Romano, S.; Piel, J.; Schwedt, A.; Schulz-Vogt, H.N. The genus Pseudovibrio contains metabolically versatile bacteria adapted for symbiosis. Environ. Microbiol. 2013, 15, 2095–2113. [Google Scholar] [CrossRef]
- CambridgeSoft Corporation, ChemBioDraw Software. 2012. PerkinElmer, 100 Cambridge Park Drive, Cambridge, MA 02140. Available online: http://www.cambridgesoft.com/Ensemble_for_Biology/ChemBioDraw/Default.aspx. Analysis and visualisation of molecular structures (Accessed on 23 November 2014).
- Berger, M.; Neumann, A.; Schulz, S.; Simon, M.; Brinkhoff, T. Tropodithietic acid production in Phaeobacter gallaeciensis is regulated by N-acyl homoserine lactone-mediated quorum sensing. J. Bacteriol. 2011, 193, 6576–6585. [Google Scholar] [CrossRef] [PubMed]
- Crowley, S.P.; O’Gara, F.; O’Sullivan, O.; Dobson, A.D.W.; Cotter, P.C. Pseudovibrio isolates with anti-Salmonella activity. In progress.
- Geng, H.; Bruhn, J.B.; Nielsen, K.F.; Gram, L.; Belas, R. Genetic dissection of tropodithietic acid biosynthesis by marine roseobacters. Appl. Environ. Microbiol. 2008, 74, 1535–1545. [Google Scholar] [CrossRef] [PubMed]
- Putze, J.; Hennequin, C.; Nougayrede, J.P.; Zhang, W.L.; Homburg, S.; Karch, H.; Bringer, M.A.; Fayolle, C.; Carniel, E.; Rabsch, W.; et al. Genetic structure and distribution of the colibactin genomic island among members of the family enterobacteriaceae. Infect. Immun. 2009, 77, 4696–4703. [Google Scholar] [CrossRef] [PubMed]
- Nougayrède, J.P.; Homburg, S.; Taieb, F.; Boury, M.; Brzuszkiewicz, E.; Gottschalk, G.; Buchrieser, C.; Hacker, J.; Dobrindt, U.; Oswald, E. Escherichia coli induces DNA double-strand breaks in eukaryotic cells. Science 2006, 313, 848–851. [Google Scholar] [CrossRef] [PubMed]
- Romano, S.; Dittmar, T.; Bondarev, V.; Weber, R.J.M.; Viant, M.R.; Schulz-Vogt, H.N. Exo-Metabolome of Pseudovibrio sp. FO-BEG1 Analyzed by Ultra-High Resolution Mass Spectrometry and the Effect of Phosphate Limitation. PLoS One 2014, 9, e96038. [Google Scholar] [CrossRef] [PubMed]
- Genomes. Metabolic pathways—Pseudovibrio sp. FO-BEG1. Available online: http://www.genome.jp/kegg-bin/show_pathway?psf01100 (accessed on 20 August 2014).
- Blin, K.; Medema, M.H.; Kazempour, D.; Fischbach, M.A.; Breitling, R.; Takano, E.; Weber, T. AntiSMASH 2.0—A versatile platform for genome mining of secondary metabolite producers. Nucleic Acids Res. 2013, 41, 204–212. [Google Scholar] [CrossRef]
- De Jong, A.; van Hijum, S.A.F.T.; Bijlsma, J.J.E.; Kok, J.; Kuipers, O.P. BAGEL: A web-based bacteriocin genome mining tool. Nucleic Acids Res. 2006, 34, 273–279. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crowley, S.P.; O'Gara, F.; O'Sullivan, O.; Cotter, P.D.; Dobson, A.D.W. Marine Pseudovibrio sp. as a Novel Source of Antimicrobials. Mar. Drugs 2014, 12, 5916-5929. https://doi.org/10.3390/md12125916
Crowley SP, O'Gara F, O'Sullivan O, Cotter PD, Dobson ADW. Marine Pseudovibrio sp. as a Novel Source of Antimicrobials. Marine Drugs. 2014; 12(12):5916-5929. https://doi.org/10.3390/md12125916
Chicago/Turabian StyleCrowley, Susan P., Fergal O'Gara, Orla O'Sullivan, Paul D. Cotter, and Alan D. W. Dobson. 2014. "Marine Pseudovibrio sp. as a Novel Source of Antimicrobials" Marine Drugs 12, no. 12: 5916-5929. https://doi.org/10.3390/md12125916