Cyanobacteria from Terrestrial and Marine Sources Contain Apoptogens Able to Overcome Chemoresistance in Acute Myeloid Leukemia Cells

Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
2.1.1. Screening Cyanobacterial Extracts for Apoptosis-Inducing Activity

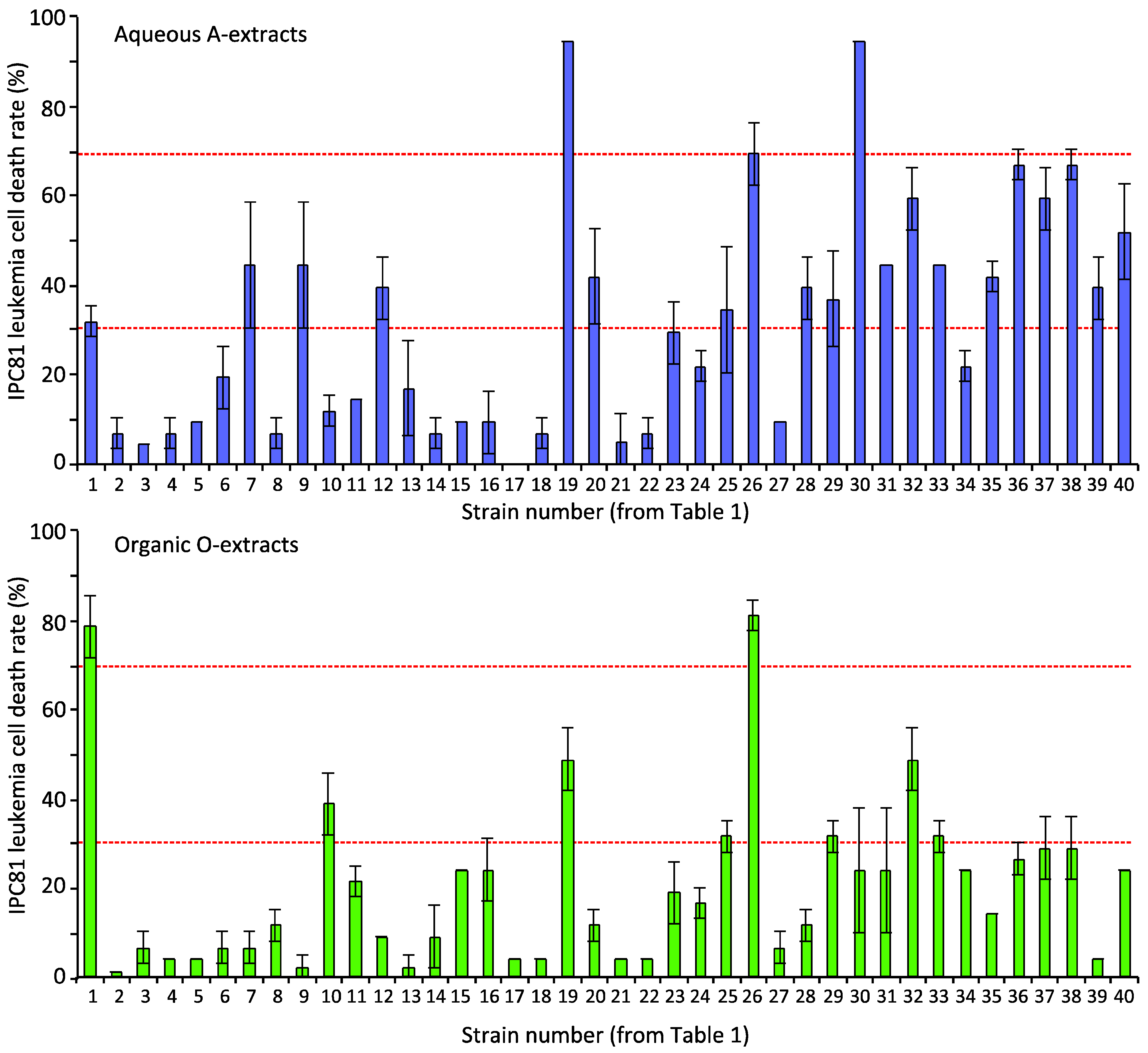{kind=link}
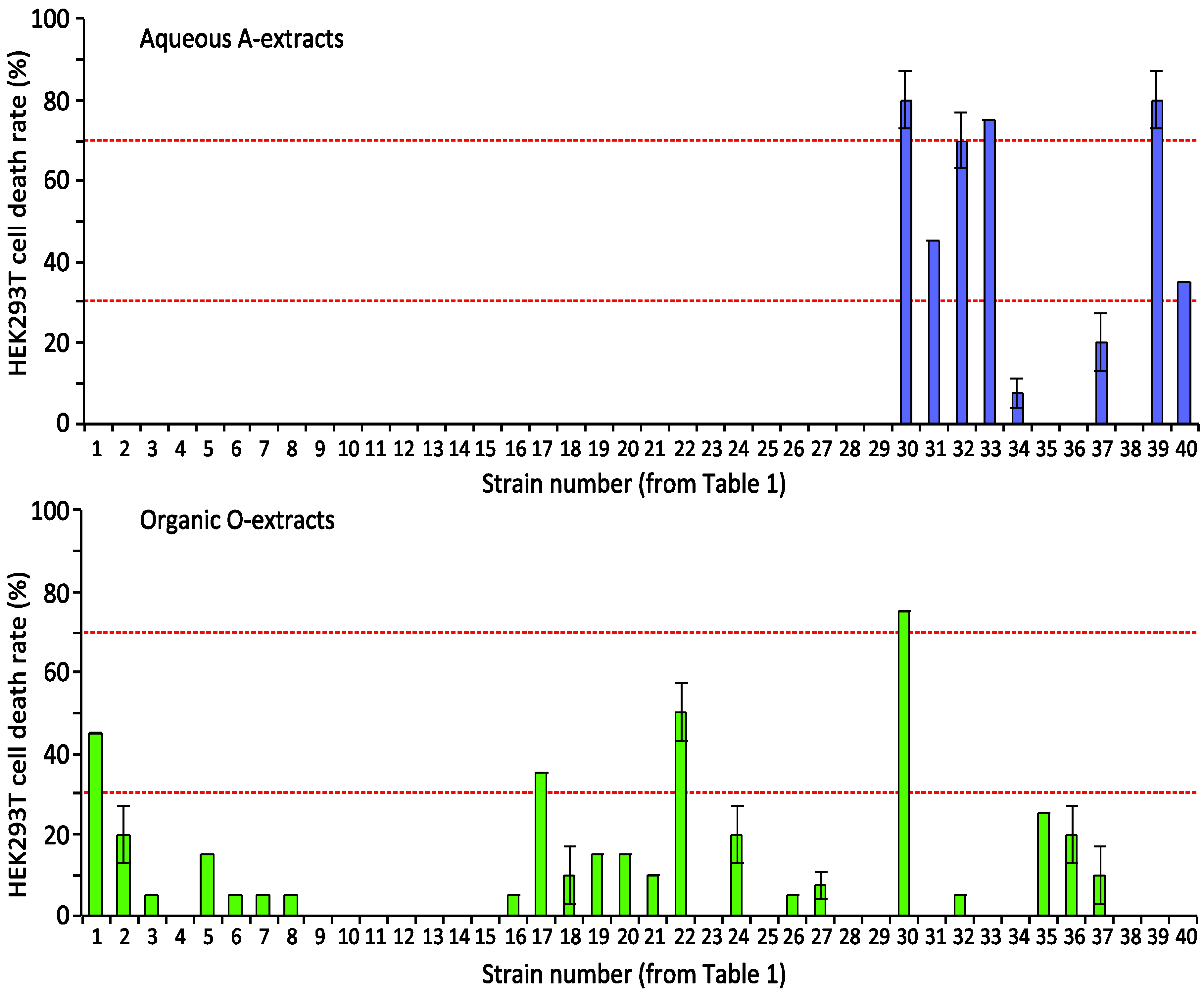{kind=link}
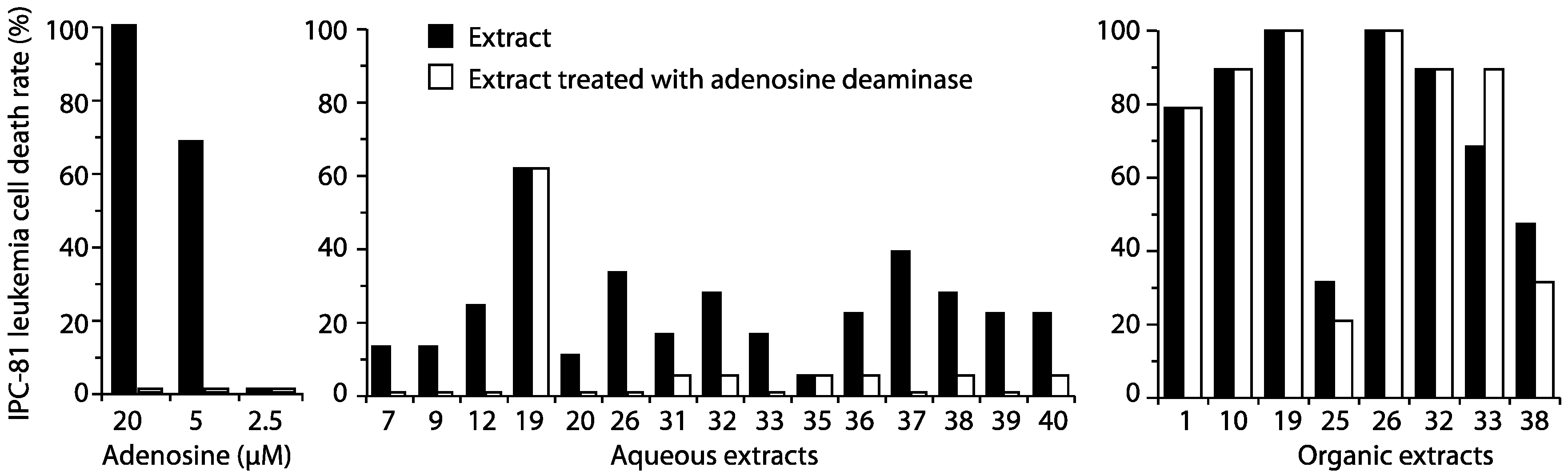{kind=link}
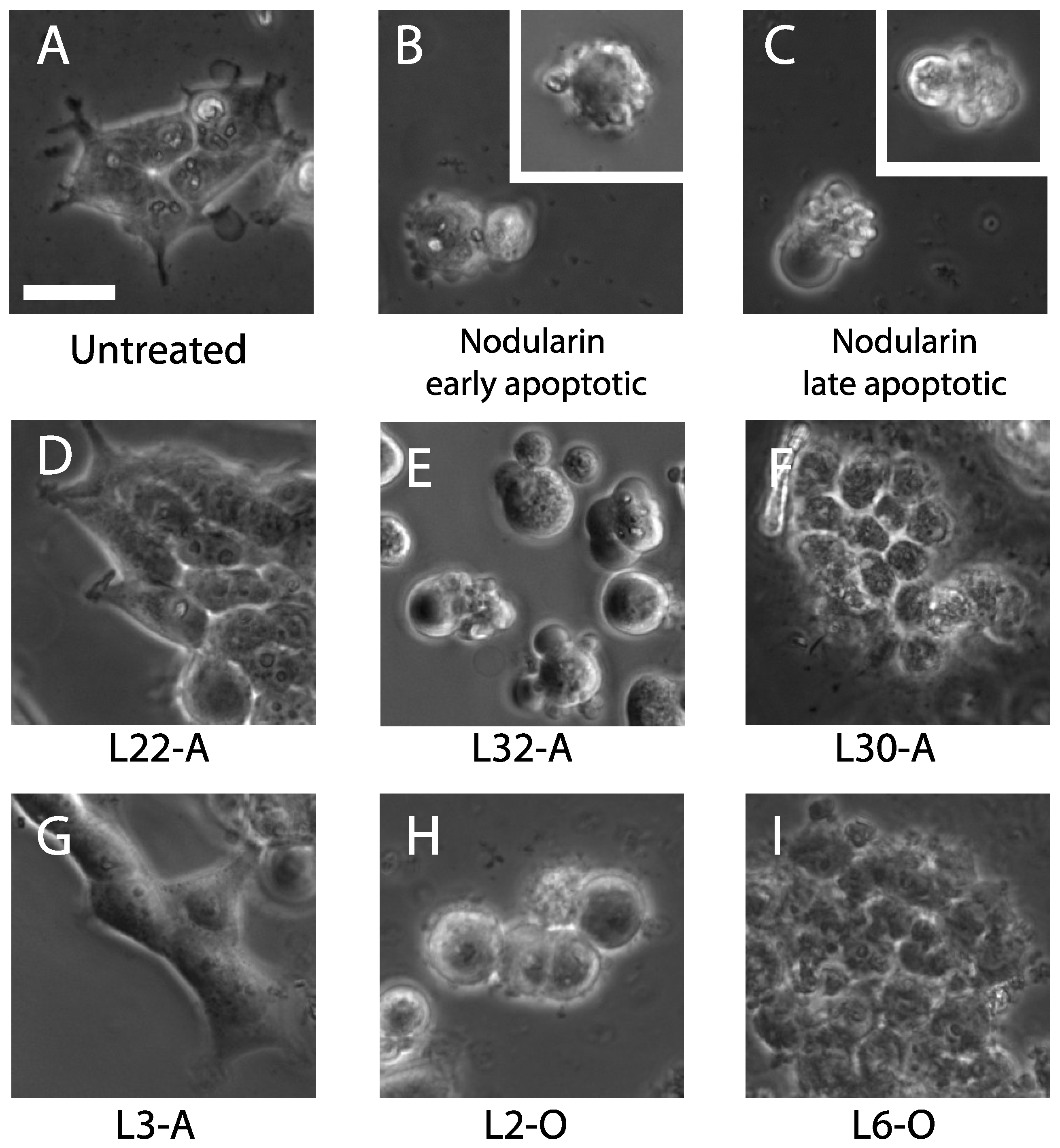{kind=link}
| Code | Genus | Strain | Live habitat; Location |
|---|---|---|---|
| L1 | Calothrix | HAN 24/1 | Rock pond water, Kobben, Hanko, Finland |
| L2 | Calothrix | HAN 33/2 | Brown/yellow biofilm, HC, Hanko, Finland |
| L3 | Calothrix | HAN 38/3 | Red biofilm, HC, Hanko, Finland |
| L4 | Calothrix | HAN 6/4 | Growth on rock waterline, Kobben, Hanko, Finland |
| L5 | Calothrix | HAN 17/1 | Red biofilm, Kobben, Hanko, Finland |
| L6 | Calothrix | HAN 21/4 | Gastropod 10 cm under water, Kobben, Hanko, Finland |
| L7 | Anabaena | HAN 15/1 | Gastropod from waterline, Kobben, Hanko, Finland |
| L8 | Calothrix | HAN 21/3 | Gastropod 10 cm under water, Kobben, Hanko, Finland |
| L9 | Calothrix | HAN 37/3 | Green biofilm, HC, Hanko, Finland |
| L10 | Calothrix | HAN 21/5 | Gastropod 10 cm under water, Kobben, Hanko, Finland |
| L11 | Calothrix | HAN 36/2 | Biofilm, HC, Hanko, Finland |
| L12 | Calothrix | HAN 22/1 | Black biofilm, Kobben, Hanko, Finland |
| L13 | Calothrix | HAN 22/2 | Black biofilm, Kobben, Hanko, Finland |
| L14 | Calothrix | HAN 6/3 | Growth on rock waterline, Kobben, Hanko, Finland |
| L15 | Calothrix | HAN 8/1 | Biofilm, Kobben, Hanko, Finland |
| L16 | Calothrix | HAN 38/2 | Red biofilm, HC, Hanko, Finland |
| L17 | Calothrix | HAN 3/19 | Green biofilm, Kobben, Hanko, Finland |
| L18 | Calothrix | HAN 33/1 | Brown/yellow biofilm, HC, Hanko, Finland |
| L19 | Nodularia | HAN 37/1 | Green biofilm, HC, Hanko, Finland |
| L20 | Calothrix | HAN 16/1 | Waterplant from shallow water, Kobben, Hanko, Finland |
| L21 | Calothrix | HAN 37/2 | Green biofilm, HC, Hanko, Finland |
| L22 | Calothrix | HAN 26/2 | Black biofilm, HC, Hanko, Finland |
| L23 | Calothrix | HAN 30/2 | Green biofilm and Gastropod, HC, Hanko, Finland |
| L24 | Calothrix | HAN 20/2 | Black grains from rock surface, Kobben, Hanko, Finland |
| L25 | Anabaena | HAN 15/2 | Gastropod from waterline, Kobben, Hanko, Finland |
| L26 | Anabaena | HAN 21/1 | Gastropod 10 cm under water, Kobben, Hanko, Finland |
| L27 | Calothrix | HAN 16/2 | Waterplant from shallow water, Kobben, Hanko, Finland |
| L28 | Nostoc | UK 2aImII | Peltigera sp. Helsinki, Finland |
| L29 | Nostoc | UK 222IIc | Peltigera sp. Mikkeli, Finland |
| L30 | Nostoc | 113.5 | Nephroma arcticum, Finland |
| L31 | Nostoc | UK 92Ic | Peltigera sp. Hitonhaudan rotko, Finland |
| L32 | Nostoc | UK 89 | Peltigera sp. Hitonhaudan rotko, Finland |
| L33 | Nostoc | UK 81I | Peltigera sp. Scotland |
| L34 | Nostoc | UK 222Ib | Peltigera sp. Mikkeli, Finland |
| L35 | Nostoc | UK 60II | Peltigera sp. Scotland |
| L36 | Nostoc | N135.9.1 | lichen, unknown |
| L37 | Nostoc | N138 | lichen, unknown |
| L38 | Nostoc | UK 104 | Peltigera sp. Teeri-Lososuo, Kuhmo, Finland |
| L39 | Nostoc | UK 220Ib | Peltigera sp. Mikkeli, Finland |
| L40 | Nostoc | N134.1 | lichen, unknown |
2.1.2. The Detection of Known Bioactivities



2.1.3. Apoptosis-Inducing Features of Cyanobacteria Extracts

| Extract | Microcystin-like activity in OATP-transfected HEK293 cells | Microcystins by LC-MS (nM) |
|---|---|---|
| L1-A | − | − |
| L7-A | − | 3.9 |
| L9-A | − | − |
| L12-A | − | − |
| L19-A | − | − |
| L20-A | − | − |
| L26-A | − | − |
| L30 A | ND | 3.8 |
| L31-A | + | 2100 |
| L32-A | ++ | 2700 |
| L33-A | + | 5100 |
| L35-A | / | 2600 |
| L36-A | − | 620 |
| L37-A | + | 13,000 |
| L38-A | − | 54 |
| L39-A | + | 2200 |
| L40-A | + | 1300 |
| L1-O | − | − |
| L10-O | − | − |
| L19-O | − | − |
| L25-O | − | − |
| L26-O | − | − |
| L32-O | − | 43 |
| L33-O | − | 89 |
| L38-O | − | 5.6 |
| Extract | EC50 (IPC-81) | Bcl-2 a | p53 b | EC50 (Molm-13) |
|---|---|---|---|---|
| L19-A | 2.4 | - | 0 | 2.1 |
| L26-A | >3.1 | - | 0 | >3.1 |
| L30-A | 0.8 | - | - | <0.5 |
| L36-A | >4.8 | - | 0 | >4.8 |
| L1-O | 0.3 | - | - | <0.3 |
| L26-O | <0.3 | + | 0 | <<0.3 |
| L30-O | ND | + | ND | ND |
2.2. Discussion
3. Experimental Section
3.1. Reagents
3.2. Isolation, Cultivation and Identification of Cyanobacteria
3.3. Preparation of Cyanobacterial Extracts for Cell Experiments
3.4. Cell Lines and Maintenance
3.5. Determination of Cell Death
3.6. Hydrolysis of Extracts with Adenosine Deaminase
3.7. Analysis of Microcystins with LC-MS
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mehta, A.; Hoffbrand, V. Haematology at a Glance, 3rd ed.; Wiley-Blackwell: Oxford, UK, 2010; pp. 60–63. [Google Scholar]
- Fischer, T.; Stone, R.M.; Deangelo, D.J.; Galinsky, I.; Estey, E.; Lanza, C.; Fox, E.; Ehninger, G.; Feldman, E.J.; Schiller, G.J.; et al. Phase IIB trial of oral midostaurin (PKC412), the FMS-like tyrosine kinase 3 receptor (FLT3) and multi-targeted kinase inhibitor, in patients with acute myeloid leukemia and high-risk myelodysplastic syndrome with either wild-type or mutated FLT3. J. Clin. Oncol. 2010, 28, 4339–4345. [Google Scholar] [CrossRef]
- Shukla, N.; Kobos, R.; Renaud, T.; Steinherz, L.J.; Steinherz, P.G. Phase II trial of clofarabine with topotecan, vinorelbine, and thiotepa in pediatric patients with relapsed or refractory acute leukemia. Pediatr. Blood Cancer 2014, 61, 431–435. [Google Scholar] [CrossRef]
- Nand, S.; Othus, M.; Godwin, J.E.; Willman, C.L.; Norwood, T.H.; Howard, D.S.; Coutre, S.E.; Erba, H.P.; Appelbaum, F.R. A phase II trial of azacitidine and gemtuzumab ozogamicin therapy in older patients with acute myeloid leukemia. Blood 2013, 122, 3432–3439. [Google Scholar] [CrossRef]
- Craddock, C.; Quek, L.; Goardon, N.; Freeman, S.; Siddique, S.; Raghavan, M.; Aztberger, A.; Schuh, A.; Grimwade, D.; Ivey, A.; et al. Azacitidine fails to eradicate leukemic stem/progenitor cell populations in patients with acute myeloid leukemia and myelodysplasia. Leukemia 2013, 27, 1028–1036. [Google Scholar] [CrossRef]
- August, K.J.; Narendran, A.; Neville, K.A. Pediatric relapsed or refractory leukemia: New pharmacotherapeutic developments and future directions. Drugs 2013, 73, 439–461. [Google Scholar] [CrossRef]
- Lücking, R.; Lawrey, J.D.; Sikaroodi, M.; Gillevet, P.M.; Chaves, J.L.; Sipman, H.J.; Bungartz, F. Do lichens domesticate photobionts like farmers domesticate crops? Evidence from a previously unrecognized lineage of filamentous cyanobacteria. Am. J. Bot. 2009, 96, 1409–1418. [Google Scholar] [CrossRef]
- Kuiper-Goodman, T.; Falconer, I.; Fitzgerald, J. Human Health Aspects. In Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring, and Management; Chorus, I., Bartram, J., Eds.; E&FN Spon: London, UK, 1999; pp. 113–153. [Google Scholar]
- Sivonen, K.; Jones, G. Cyanobacterial Toxins. In Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring, and Management; Chorus, I., Bartram, J., Eds.; E&FN Spon: London, UK, 1999; pp. 41–111. [Google Scholar]
- Sivonen, K. Cyanobacterial Toxins. In Encyclopedia of Microbiology; Schaechter, M., Ed.; Elsevier: Oxford, UK, 2009; pp. 290–307. [Google Scholar]
- Zanchett, G.; Oliveira-Filho, E.C. Cyanobacteria and cyanotoxins: From impacts on aquatic ecosystems and human health to anticarcinogenic effects. Toxins 2013, 5, 1896–1917. [Google Scholar] [CrossRef]
- Sainis, I.; Fokas, D.; Vareli, K.; Tzakos, A.G.; Kounnis, V.; Briasoulis, E. Cyanobacterial cyclopeptides as lead compounds to novel targeted cancer drugs. Mar. Drugs 2010, 8, 629–657. [Google Scholar] [CrossRef]
- Dixit, R.B.; Suseela, M.R. Cyanobacteria: Potential candidates for drug discovery. Antonie Van Leeuwenhoek 2013, 103, 947–961. [Google Scholar] [CrossRef]
- Burja, A.M.; Banaigs, B.; Abou-Mansour, E.; Burgess, J.G.; Wright, P.C. Marine cyanobacteria—A prolific source of natural products. Tetrahedron 2001, 57, 9347–9377. [Google Scholar] [CrossRef]
- Sivonen, K.; Börner, T. Bioactive compounds produced by cyanobacteria. In The Cyanobacteria: Molecular Biology, Genomics and Evolution; Herrero, A., Flores, E., Eds.; Caister Academic Press: Linton, UK, 2008; pp. 159–197. [Google Scholar]
- D’Agostino, G.; del Campo, J.; Mellado, B.; Izquierdo, M.A.; Minarik, T.; Cirri, L.; Marini, L.; Perez-Gracia, J.L.; Scambia, G. A multicenter phase II study of the cryptophycin analog LY355703 in patients with platinum-resistant ovarian cancer. Int. J. Gynecol. Cancer 2006, 16, 71–76. [Google Scholar] [CrossRef]
- Verdier-Pinard, P.; Lai, J.Y.; Yoo, H.D.; Yu, J.; Marquez, B.; Nagle, D.G.; Nambu, M.; White, J.D.; Falck, J.R.; Gerwick, W.H.; et al. Structure-activity analysis of the interaction of curacin A, the potent colchicine site antimitotic agent, with tubulin and effects of analogs on the growth of MCF-7 breast cancer cells. Mol. Pharmacol. 1998, 53, 62–76. [Google Scholar]
- Perez, E.A.; Hillman, D.W.; Fishkin, P.A.; Krook, J.E.; Tan, W.W.; Kuriakose, P.A.; Alberts, S.R.; Dakhil, S.R. Phase II trial of dolastatin-10 in patients with advanced breast cancer. Invest. New Drugs 2005, 23, 257–261. [Google Scholar] [CrossRef]
- Gunasekera, S.P.; Owle, C.S.; Montaser, R.; Luesch, H.; Paul, V.J. Malyngamide 3 and cocosamides A and B from the marine cyanobacterium Lyngbya majuscula from Cocos Lagoon, Guam. J. Nat. Prod. 2011, 74, 871–876. [Google Scholar] [CrossRef]
- Luesch, H.; Yoshida, W.Y.; Moore, R.E.; Paul, V.J.; Corbett, T.H. Total structure determination of apratoxin A, a potent novel cytotoxin from the marine cyanobacterium Lyngbya majuscula. J. Am. Chem. Soc. 2001, 123, 5418–5423. [Google Scholar] [CrossRef]
- Teruya, T.; Sasaki, H.; Fukazawa, H.; Suenaga, K. Bisebromoamide, a potent cytotoxic peptide from the marine cyanobacterium Lyngbya sp.: Isolation, stereostructure, and biological activity. Org. Lett. 2009, 11, 5062–5065. [Google Scholar] [CrossRef]
- Hau, A.M.; Greenwood, J.A.; Lohr, C.V.; Serrill, J.D.; Proteau, P.J.; Ganley, I.G.; McPhail, K.L.; Ishmael, J.E. Coibamide a induces mTOR-independent autophagy and cell death in human glioblastoma cells. PLoS One 2013, 8, e5250. [Google Scholar]
- Hong, J.Y.; Luesch, H. Largazole: From discovery to broad-spectrum therapy. Nat. Prod. Rep. 2012, 29, 449–456. [Google Scholar] [CrossRef]
- Lacaze, N.; Gombaud-Saintonge, G.; Lanotte, M. Conditions controlling long-term proliferation of Brown Norway rat promyelocytic leukemia in vitro: Primary growth stimulation by microenvironment and establishment of an autonomous Brown Norway “leukemic stem cell line”. Leuk. Res. 1983, 7, 145–154. [Google Scholar] [CrossRef]
- McCormack, E.; Bruserud, O.; Gjertsen, B.T. Animal models of acute myelogenous leukaemia development, application and future perspectives. Leukemia 2005, 19, 687–706. [Google Scholar] [CrossRef]
- Eriksson, J.E.; Toivola, D.; Meriluoto, J.A.; Karaki, H.; Han, Y.G.; Hartshorne, D. Hepatocyte deformation induced by cyanobacterial toxins reflects inhibition of protein phosphatases. Biochem. Biophys. Res. Commun. 1990, 173, 1347–1353. [Google Scholar] [CrossRef]
- Runnegar, M.T.; Kong, S.; Berndt, N. Protein phosphatase inhibition and in vivo hepatotoxicity of microcystins. Am. J. Physiol. 1993, 265, 224–230. [Google Scholar]
- Herfindal, L.; Selheim, F. Microcystin produces disparate effects on liver cells in a dose dependent manner. Mini-Rev. Med. Chem. 2006, 6, 109–120. [Google Scholar] [CrossRef]
- Herfindal, L.; Kasprzykowski, F.; Schwede, F.; Łankiewicz, L.; Fladmark, K.E; Łukomska, J.; Wahlsten, M.; Sivonen, K.; Grzonka, Z.; Jastorff, B.; et al. Acyloxymethyl esterification of nodularin-R and microcystin-LA produces inactive protoxins that become reactivated and produce apoptosis inside intact cells. J. Med. Chem. 2009, 52, 5758–5762. [Google Scholar] [CrossRef]
- Prestegard, S.K.; Oftedal, L.; Coyne, R.T.; Nygaard, G.; Skjaerven, K.H.; Knutsen, G.; Døskeland, S.O.; Herfindal, L. Marine benthic diatoms contain compounds able to induce leukemia cell death and modulate blood platelet activity. Mar. Drugs 2009, 7, 605–623. [Google Scholar] [CrossRef]
- Oftedal, L.; Herfindal, L.; Døskeland, S.O.; University of Bergen, Bergen, Norway. Unpublished work. 2010.
- Tanaka, Y.; Yoshihara, K.; Tsuyuki, M.; Kamiya, T. Apoptosis induced by adenosine in human leukemia HL-60 cells. Exp. Cell Res. 1994, 213, 242–252. [Google Scholar] [CrossRef]
- Fischer, W.J.; Altheimer, S.; Cattori, V.; Meier, P.J.; Dietrich, D.R.; Hagenbuch, B. Organic anion transporting polypeptides expressed in liver and brain mediate uptake of microcystin. Toxicol. Appl. Pharmacol. 2005, 203, 257–263. [Google Scholar] [CrossRef]
- Lu, H.; Choudhuri, S.; Ogura, K.; Csanaky, I.L.; Lei, X.; Cheng, X.; Song, P.Z.; Klaassen, C.D. Characterization of organic anion transporting polypeptide 1b2-null mice: Essential role in hepatic uptake/toxicity of phalloidin and microcystin-LR. Toxicol. Sci. 2008, 103, 35–45. [Google Scholar]
- Herfindal, L.; Myhren, L.; Kleppe, R.; Krakstad, C.; Selheim, F.; Jokela, J.; Sivonen, K.; Døskeland, S.O. Nostocyclopeptide-M1: A potent, nontoxic inhibitor of the hepatocyte drug transporters OATP1B3 and OATP1B1. Mol. Pharm. 2011, 8, 360–367. [Google Scholar] [CrossRef]
- Jokela, J.; Herfindal, L.; Wahlsten, M.; Permi, P.; Selheim, F.; Vasconçelos, V.; Døskeland, S.O.; Sivonen, K. A novel cyanobacterial nostocyclopeptide is a potent antitoxin against microcystins. Chembiochem 2010, 11, 1594–1599. [Google Scholar] [CrossRef]
- Matsuo, Y.; MacLeod, R.A.; Uphoff, C.C.; Drexler, H.G.; Nishizaki, C.; Katayama, Y.; Kimura, G.; Fujii, N.; Omoto, E.; Harada, M.; et al. Two acute monocytic leukemia (AML-M5a) cell lines (MOLM-13 and MOLM-14) with interclonal phenotypic heterogeneity showing MLL-AF9 fusion resulting from an occult chromosome insertion, ins(11;9)(q23;p22p23). Leukemia 1997, 11, 1469–1477. [Google Scholar]
- Quentmeier, H.; Reinhardt, J.; Zaborski, M.; Drexler, H.G. FLT3 mutations in acute myeloid leukemia cell lines. Leukemia 2003, 17, 120–124. [Google Scholar] [CrossRef]
- Youle, R.J.; Strasser, A. The BCL-2 protein family: Opposing activities that mediate cell death. Nat. Rev. Mol. Cell Biol. 2008, 9, 47–59. [Google Scholar] [CrossRef]
- Soussi, T. p53 Zutations and resistance to chemotherapy: A stab in the back for p73. Cancer Cell 2003, 3, 303–305. [Google Scholar] [CrossRef]
- Herfindal, L.; Oftedal, L.; Selheim, F.; Wahlsten, M.; Sivonen, K.; Døskeland, S.O. A high proportion of Baltic Sea benthic cyanobacterial isolates contain apoptogens able to induce rapid death of isolated rat hepatocytes. Toxicon 2005, 46, 252–260. [Google Scholar] [CrossRef]
- Oftedal, L.; Skjærven, K.H.; Coyne, R.T.; Edvardsen, B.; Rohrlack, T.; Skulberg, O.M.; Døskeland, S.O.; Herfindal, L. The apoptosis-inducing activity towards leukemia and lymphoma cells in a cyanobacterial culture collection is not associated with mouse bioassay toxicity. J. Ind. Microbiol. Biotechnol. 2011, 38, 489–501. [Google Scholar] [CrossRef]
- Oftedal, L.; Selheim, F.; Wahlsten, M.; Sivonen, K.; Døskeland, S.O.; Herfindal, L. Marine benthic cyanobacteria contain apoptosis-inducing activity synergizing with daunorubicin to kill leukemia cells, but not cardiomyocytes. Mar. Drugs 2010, 8, 2659–2672. [Google Scholar] [CrossRef]
- Surakka, A.; Sihvonen, L.M.; Lehtimäki, J.M.; Wahlsten, M.; Vuorela, P.; Sivonen, K. Benthic cyanobacteria from the Baltic Sea contain cytotoxic Anabaena, Nodularia, and Nostoc strains and an apoptosis-inducing Phormidium strain. Environ. Toxicol. 2005, 20, 285–292. [Google Scholar] [CrossRef]
- Jokela, J.; Oftedal, L.; Herfindal, L.; Permi, P.; Wahlsten, M.; Døskeland, S.O.; Sivonen, K. Anabaenolysins, novel cytolytic lipopeptides from benthic Anabaena cyanobacteria. PLoS One 2012, 7, e41222. [Google Scholar]
- Fewer, D.P.; Jokela, J.; Paukku, E.; Osterholm, J.; Wahlsten, M.; Permi, P.; Aitio, O.; Rouhiainen, L.; Gomez-Saez, G.V.; Sivonen, K. New structural variants of aeruginosin produced by the toxic bloom forming cyanobacterium Nodularia spumigena. PLoS One 2013, 8, e73618. [Google Scholar] [CrossRef]
- Leikoski, N.; Fewer, D.P.; Jokela, J.; Wahlsten, M.; Rouhiainen, L.; Sivonen, K. Highly diverse cyanobactins in strains of the genus Anabaena. Appl. Environ. Microbiol. 2010, 76, 701–709. [Google Scholar] [CrossRef]
- Leikoski, N.; Fewer, D.P.; Jokela, J.; Alakoski, P.; Wahlsten, M.; Sivonen, K. Analysis of an inactive cyanobactin biosynthetic gene cluster leads to discovery of new natural products from strains of the genus Microcystis. PLoS One 2012, 7, e43002. [Google Scholar]
- Leikoski, N.; Liu, L.; Jokela, J.; Wahlsten, M.; Gugger, M.; Calteau, A.; Permi, P.; Kerfeld, C.A.; Sivonen, K.; Fewer, D.P. Genome mining expands the chemical diversity of the cyanobactin family to include highly modified linear peptides. Chem. Biol. 2013, 20, 1033–1043. [Google Scholar] [CrossRef]
- Wang, H.; Sivonen, K.; Rouhiainen, L.; Fewer, D.P.; Lyra, C.; Rantala-Ylinen, A.; Vestola, J.; Jokela, J.; Rantasärkkä, K.; Li, Z.; et al. Genome-derived insights into the biology of the hepatotoxic bloom-forming cyanobacterium Anabaena sp. strain 90. BMC Genomics 2012, 13, 613. [Google Scholar] [CrossRef]
- Shih, P.M.; Wu, D.; Latifi, A.; Axen, S.D.; Fewer, D.P.; Talla, E.; Calteau, A.; Cai, F.; Tandeau de Marsac, N.; Rippka, R.; et al. Improving the coverage of the cyanobacterial phylum using diversity-driven genome sequencing. Proc. Natl. Acad. Sci. USA 2013, 110, 1053–1058. [Google Scholar] [CrossRef]
- Welker, M.; von Döhren, H. Cyanobacterial peptides––Nature’s own combinatorial biosynthesis. FEMS Microbiol. Rev. 2006, 30, 530–563. [Google Scholar] [CrossRef]
- Kaasalainen, U.; Fewer, D.P.; Jokela, J.; Wahlsten, M.; Sivonen, K.; Rikkinen, J. Cyanobacteria produce a high variety of hepatotoxic peptides in lichen symbiosis. Proc. Natl. Acad. Sci. USA 2012, 109, 5886–5891. [Google Scholar]
- Carmichael, W.W.; Azevedo, S.M.; An, J.S.; Molica, R.J.; Jochimsen, E.M.; Lau, S.; Rinehart, K.L.; Shaw, G.R.; Eaglesham, G.K. Human fatalities from cyanobacteria: chemical and biological evidence for cyanotoxins. Environ. Health Perspect. 2001, 109, 663–668. [Google Scholar] [CrossRef]
- Hockenbery, D.; Nuñez, G.; Milliman, C.; Schreiber, R.D.; Korsmeyer, S.J. Bcl-2 is an inner mitochondrial membrane protein that blocks programmed cell death. Nature 1990, 348, 334–336. [Google Scholar] [CrossRef]
- Fujita, N.; Tsuruo, T. In vivo veritas: Bcl-2 and Bcl-XL mediate tumor cell resistance to chemotherapy. Drug Resist. Updates 2000, 3, 149–154. [Google Scholar] [CrossRef]
- McCormack, E.; Haaland, I.; Venås, G.; Forthun, R.B.; Huseby, S.; Gausdal, G.; Knappskog, S.; Micklem, D.R.; Lorens, J.B.; Bruserud, O.; et al. Synergistic induction of p53 mediated apoptosis by valproic acid and nutlin-3 in acute myeloid leukemia. Leukemia 2012, 26, 910–917. [Google Scholar] [CrossRef]
- Soussi, T.; Lozano, G. p53 Mutation heterogeneity in cancer. Biochem. Biophys. Res. Commun. 2005, 331, 834–842. [Google Scholar] [CrossRef]
- Blalock, W.L.; Navolanic, P.M.; Steelman, L.S.; Shelton, J.G.; Moye, P.W.; Lee, J.T.; Franklin, R.A.; Mirza, A.; McMahon, M.; White, M.K.; et al. Requirement for the PI3K/Akt pathway in MEK1-mediated growth and prevention of apoptosis: identification of an achilles heel in leukemia. Leukemia 2003, 17, 1058–1067. [Google Scholar] [CrossRef]
- Castenholz, R.W. Cyanobacteria, Oxygenic Photosynthetic Bacteria. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Boone, D.R., Castenholz, R.W., Eds.; Springer: New York, NY, USA, 2001; pp. 473–599. [Google Scholar]
- Wilmotte, A.; van der Auwera, G.; de Wachter, R. Structure of the 16S ribosomal RNA of the thermophilic cyanobacterium Chlorogloeopsis HTF (‘Mastigocladus laminosus HTF’) strain PCC7518, and phylogenic analysis. FEMS Lett. 1993, 317, 96–100. [Google Scholar]
- Neilan, B.A.; Jacobs, D.; del Dot, T.; Blackall, L.L.; Hawkins, P.R.; Cox, P.T.; Goodman, A.E. rRNA sequences and evolutionary relationships among toxic and nontoxic cyanobacteria of the genus Microcystis. Int. J. Syst. Bacteriol. 1997, 47, 693–697. [Google Scholar] [CrossRef]
- Séité, P.; Ruchaud, S.; Hillion, J.; Gendron, M.C.; Bruland, O.; Ségal-Bendirdjian, E.; Døskeland, S.O.; Lillehaug, J.R.; Lanotte, M. Ectopic expression of Bcl-2 switches over nuclear signalling for cAMP-induced apoptosis to granulocytic differentiation. Cell Death Differ. 2000, 7, 1081–1089. [Google Scholar] [CrossRef]
- König, J.; Cui, Y.; Nies, A.T.; Keppler, D. Localization and genomic organization of a new hepatocellular organic anion transporting polypeptide. J. Biol. Chem. 2000, 275, 23161–23168. [Google Scholar] [CrossRef]
- Kaasalainen, U.; Jokela, J.; Fewer, D.P.; Sivonen, K.; Rikkinen, J. Microcystin production in the tripartite cyanolichen Peltigera leucophlebia. Mol. Plant Microbe Interact. 2009, 20, 695–702. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, L.; Herfindal, L.; Jokela, J.; Shishido, T.K.; Wahlsten, M.; Døskeland, S.O.; Sivonen, K. Cyanobacteria from Terrestrial and Marine Sources Contain Apoptogens Able to Overcome Chemoresistance in Acute Myeloid Leukemia Cells. Mar. Drugs 2014, 12, 2036-2053. https://doi.org/10.3390/md12042036
Liu L, Herfindal L, Jokela J, Shishido TK, Wahlsten M, Døskeland SO, Sivonen K. Cyanobacteria from Terrestrial and Marine Sources Contain Apoptogens Able to Overcome Chemoresistance in Acute Myeloid Leukemia Cells. Marine Drugs. 2014; 12(4):2036-2053. https://doi.org/10.3390/md12042036
Chicago/Turabian StyleLiu, Liwei, Lars Herfindal, Jouni Jokela, Tania Keiko Shishido, Matti Wahlsten, Stein Ove Døskeland, and Kaarina Sivonen. 2014. "Cyanobacteria from Terrestrial and Marine Sources Contain Apoptogens Able to Overcome Chemoresistance in Acute Myeloid Leukemia Cells" Marine Drugs 12, no. 4: 2036-2053. https://doi.org/10.3390/md12042036