Molecular Response to Toxic Diatom-Derived Aldehydes in the Sea Urchin Paracentrotus lividus


 ,
, 

Abstract
:1. Introduction
2. Results
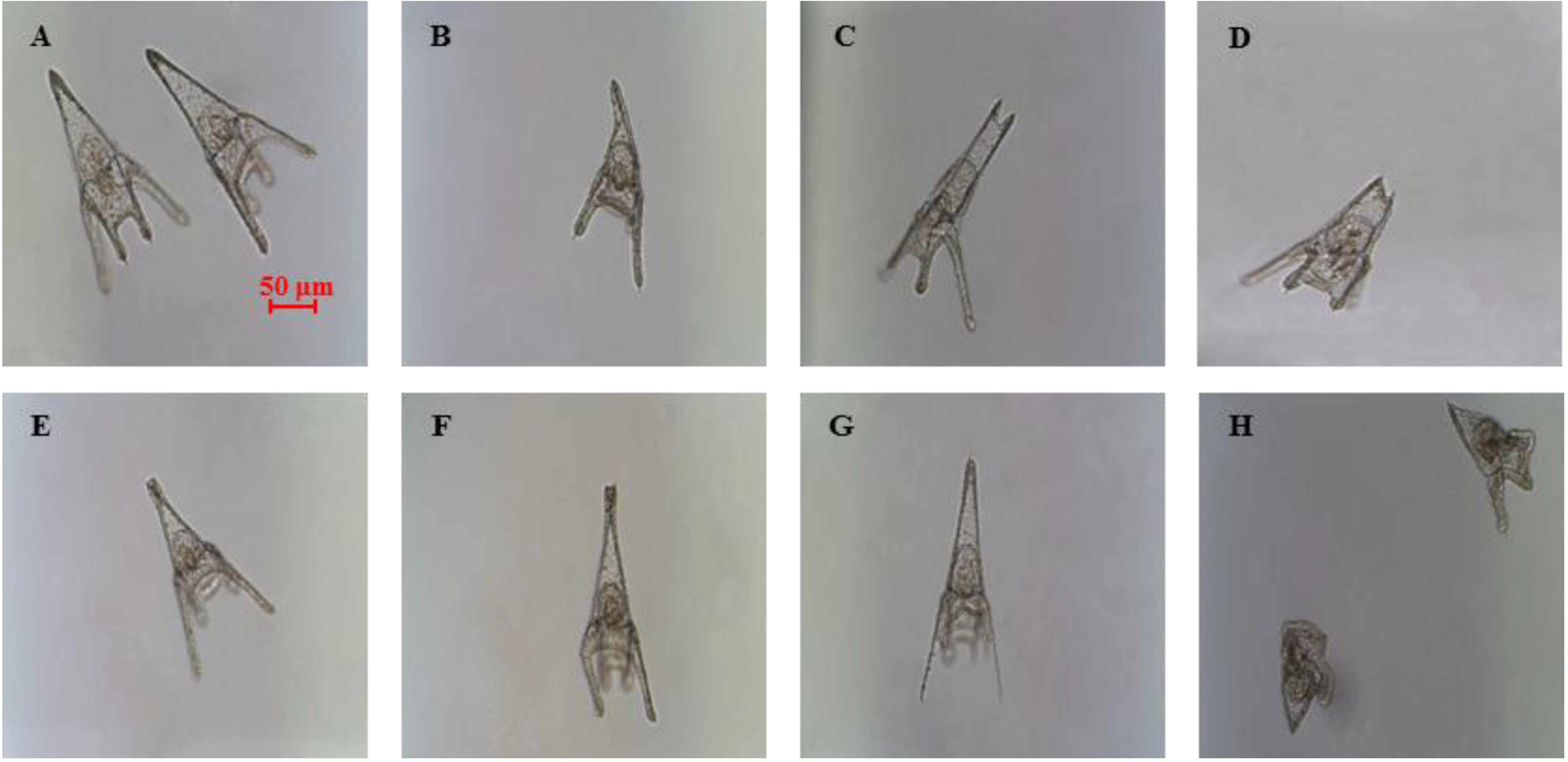
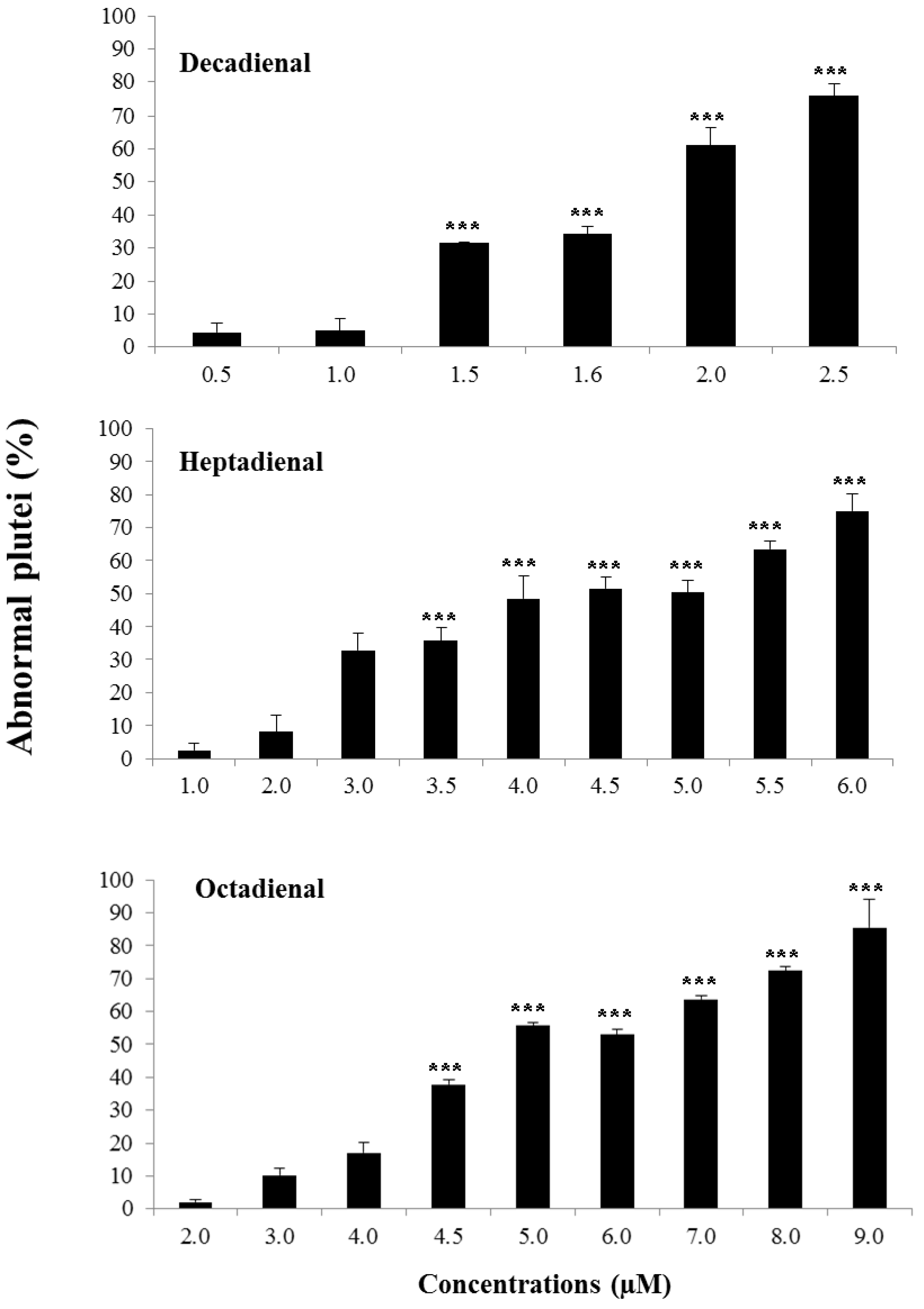
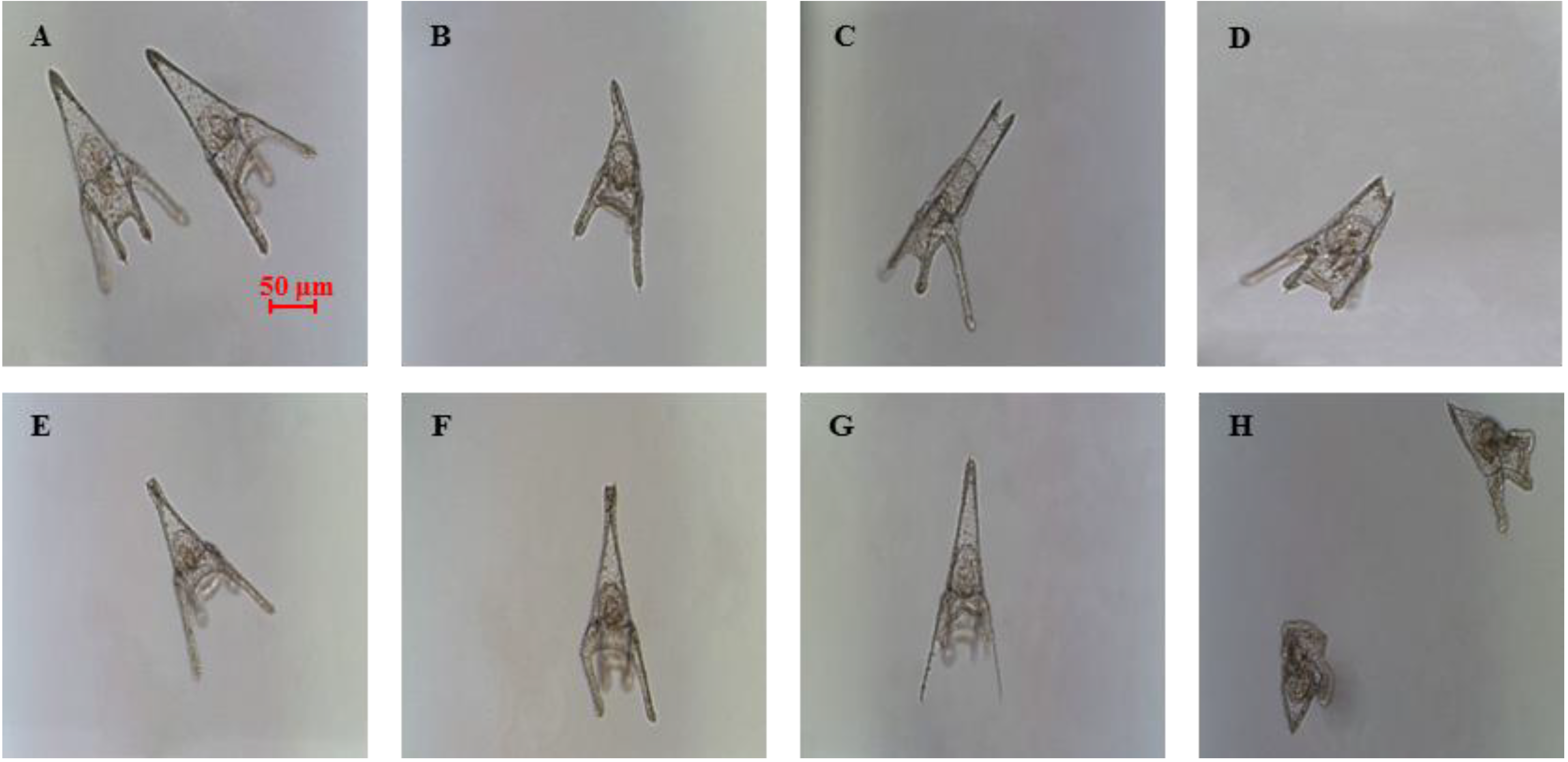
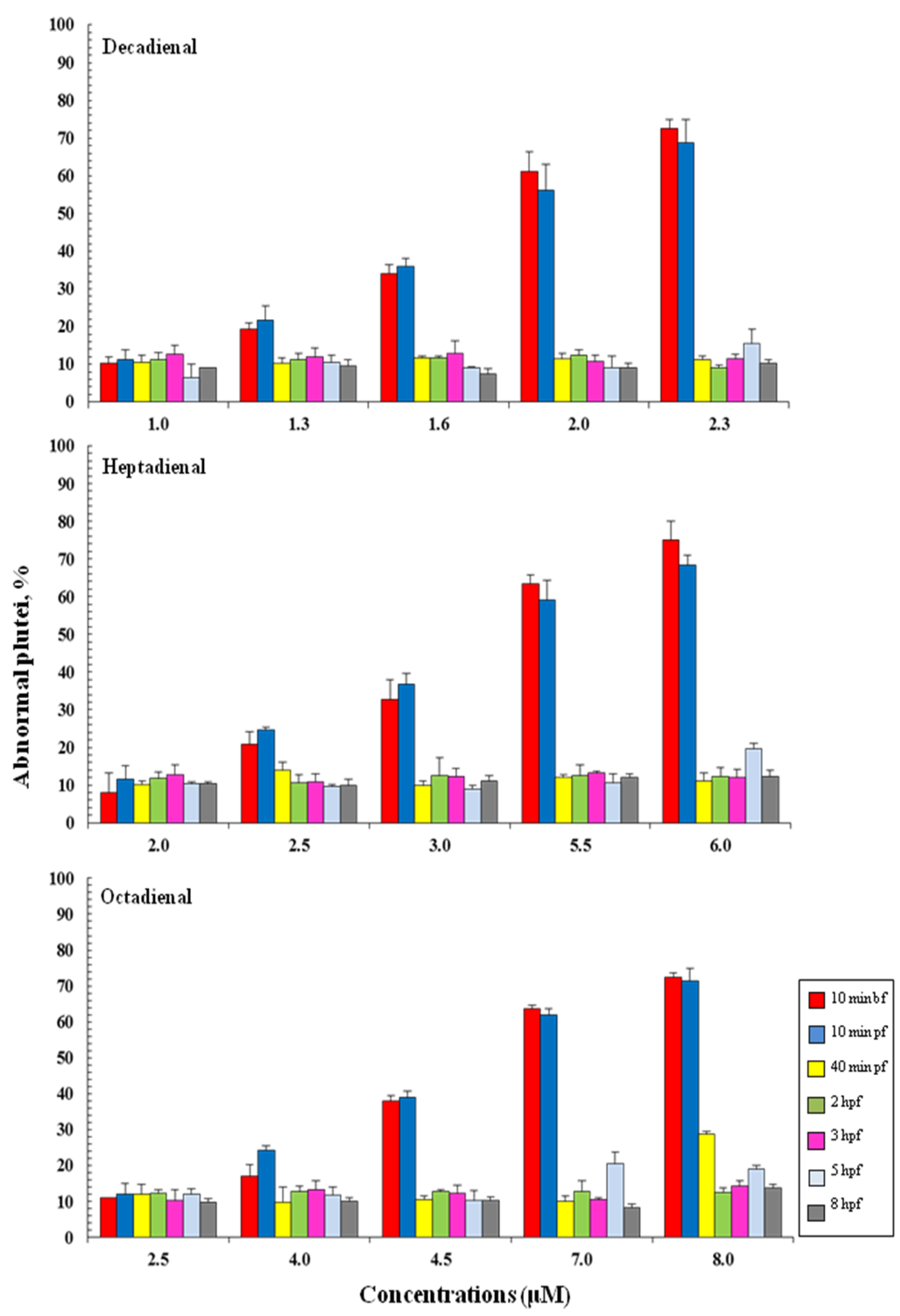
2.1. Determination of Teratogenic Effects and PUA Dose-Dependent Concentrations
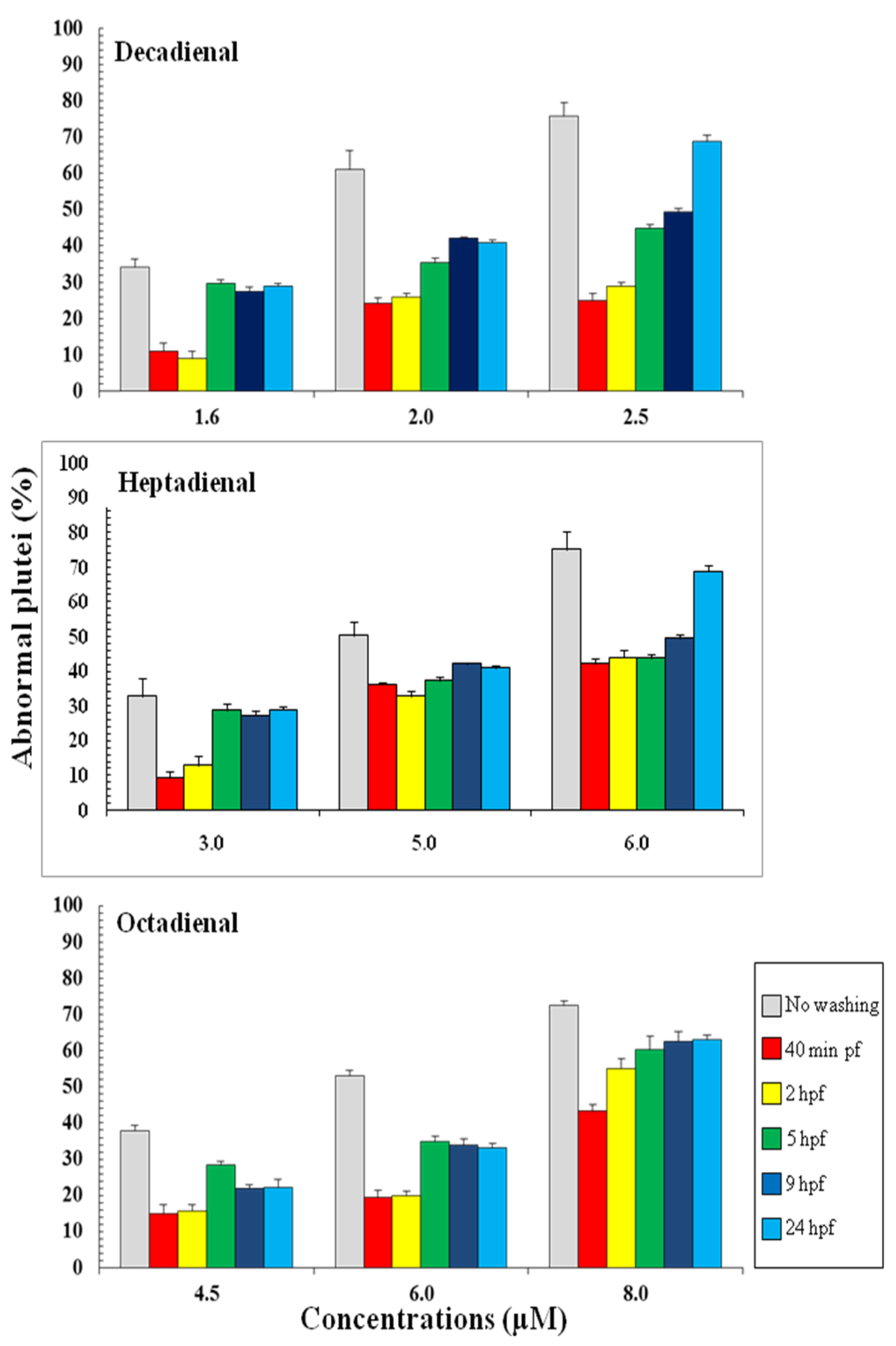
2.2. Post-Recovery Experiments
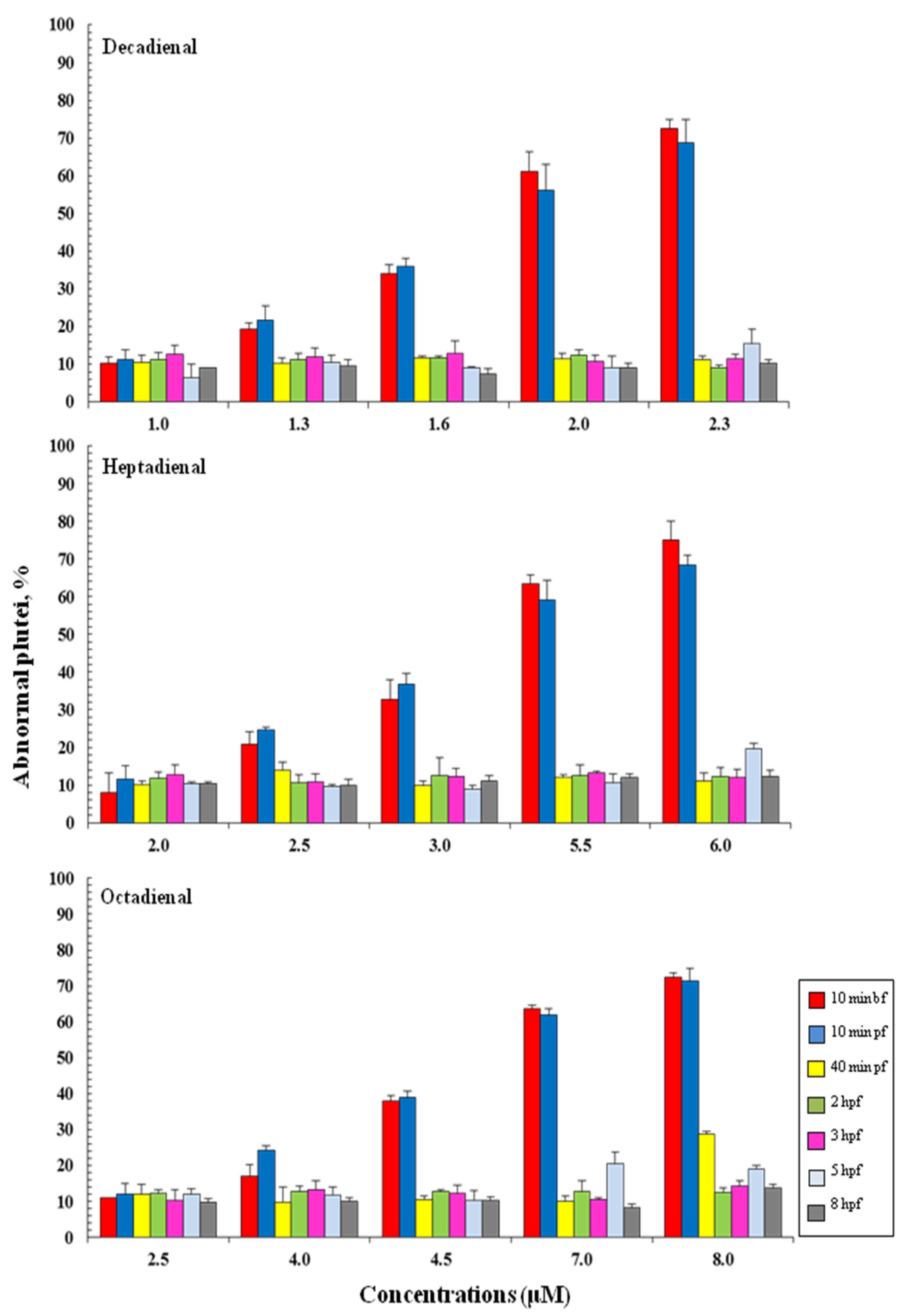
2.3. Addition of PUAs at Different Developmental Stages
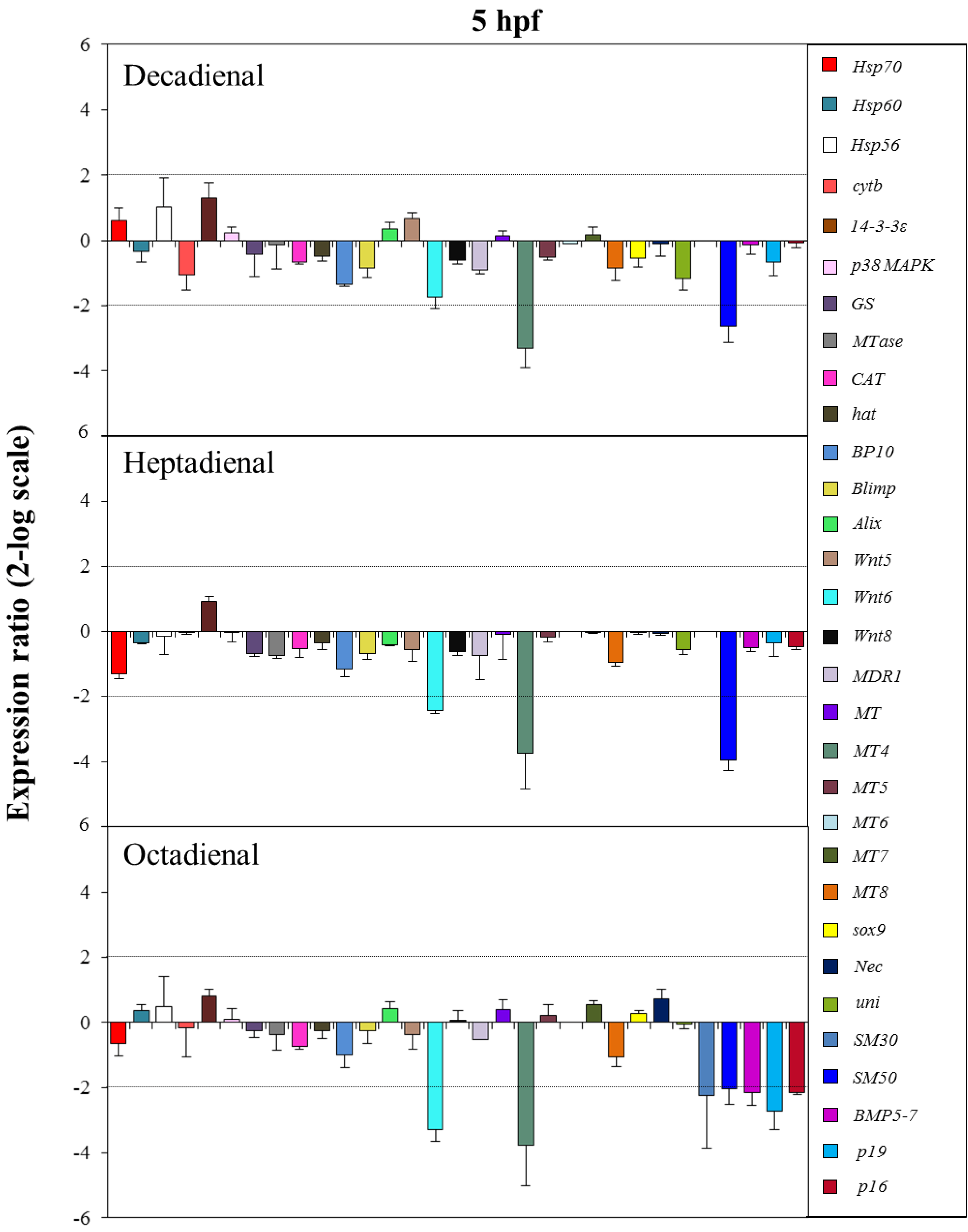
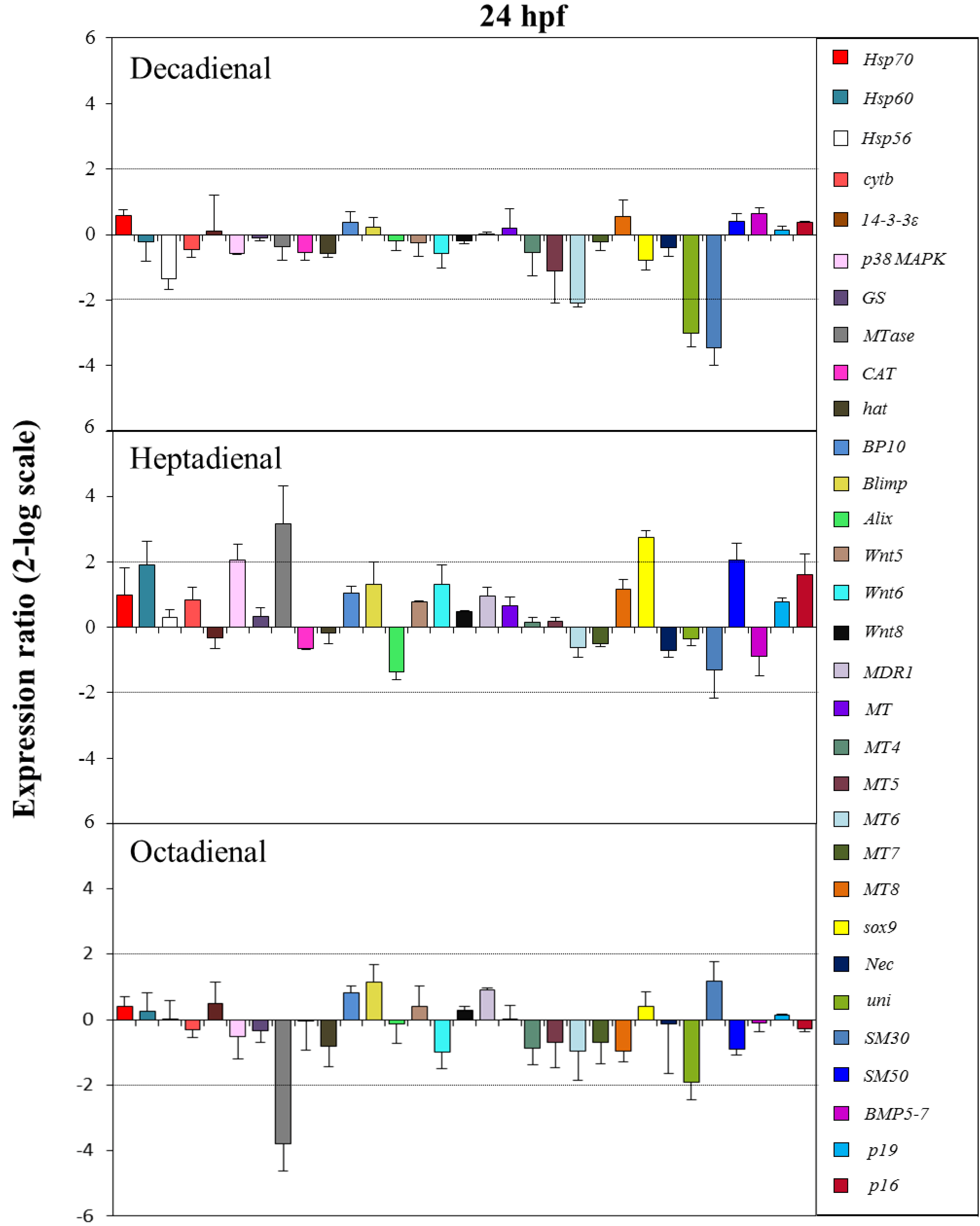
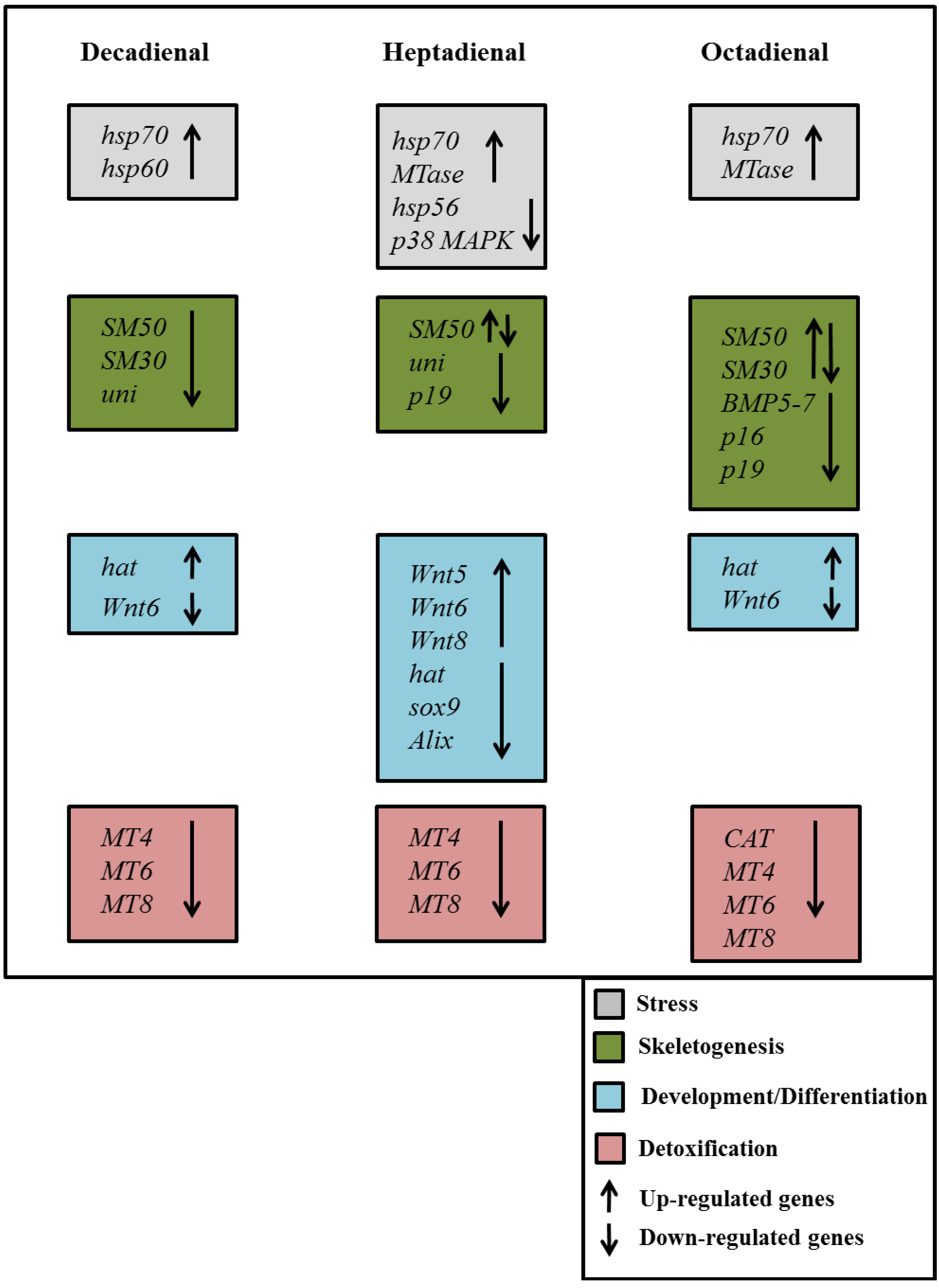
2.4. Gene Stress Response to PUAs

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
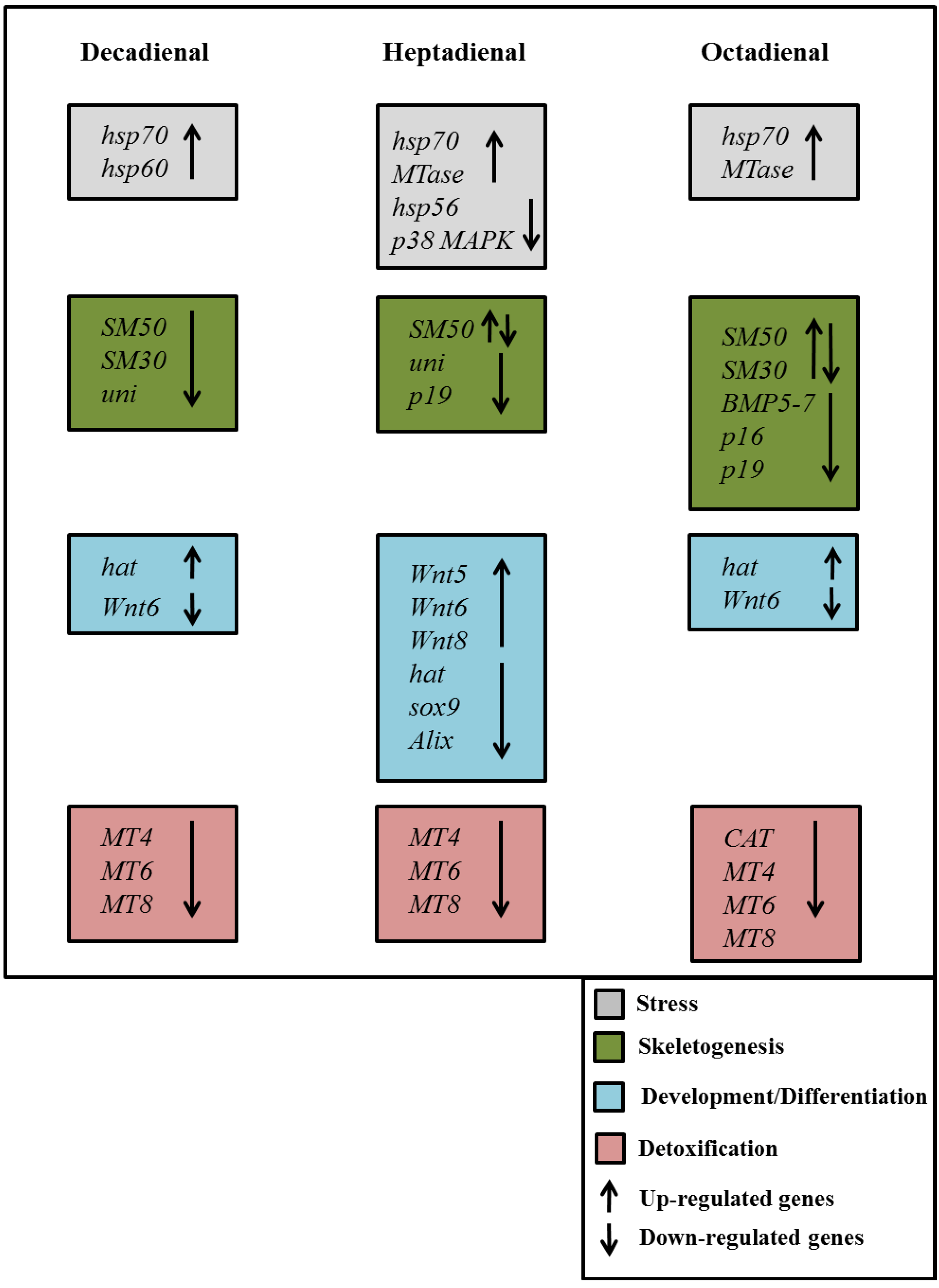{kind=link}
| Gene Name | Acronym | Function | Reference |
|---|---|---|---|
| Pl-p19 | p19 | small acidic proteins, involved in the formation of the | [14] |
| Pl-p16 | p16 | biomineralised skeleton of sea urchin embryos and adults | |
| ALG-2 interacting protein X/1 | Alix | protein involved in endocytic membrane trafficking, | [15] |
| filamentous (F)-actin remodelling and cytokinesis | |||
| Blimp | Blimp | zinc finger transcription factor, which plays a central role | [16] |
| in both early and late endomesoderm specification | |||
| Wnt 5 | Wnt5 | initiates the specification of the sea urchin posterior ectoderm | [17] |
| Wnt 6 | Wnt6 | activates endoderm in the sea urchin gene regulatory network | [18] |
| Wnt 8 | Wnt8 | endomesodermal specification, embryo patterning, early | [19] |
| primary mesenchyme cells-gene regulatory network regulator | |||
| Multi drug resistance protein 1 | MDR1 | ATP-binding cassette protein | [20] |
| Metallothionein 4 | MT4 | ||
| Metallothionein 5 | MT5 | proteins capable of binding to heavy metals, involved in | [21] |
| Metallothionein 6 | MT6 | the transport of heavy metals and cellular detoxification | |
| Metallothionein 7 | MT7 | ||
| Metallothionein 8 | MT8 | ||
| Catalase | Cat | antioxidant defensive protein |

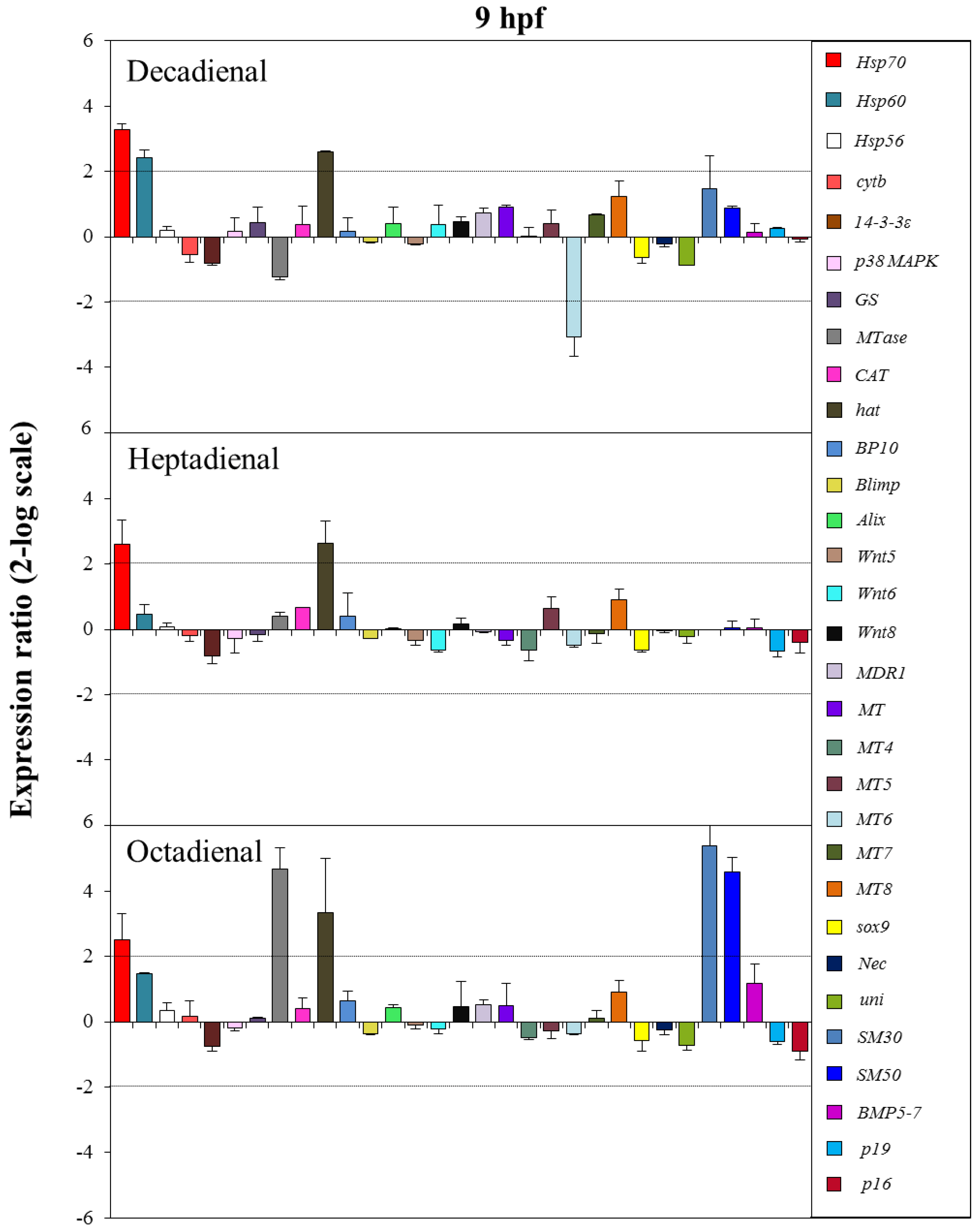
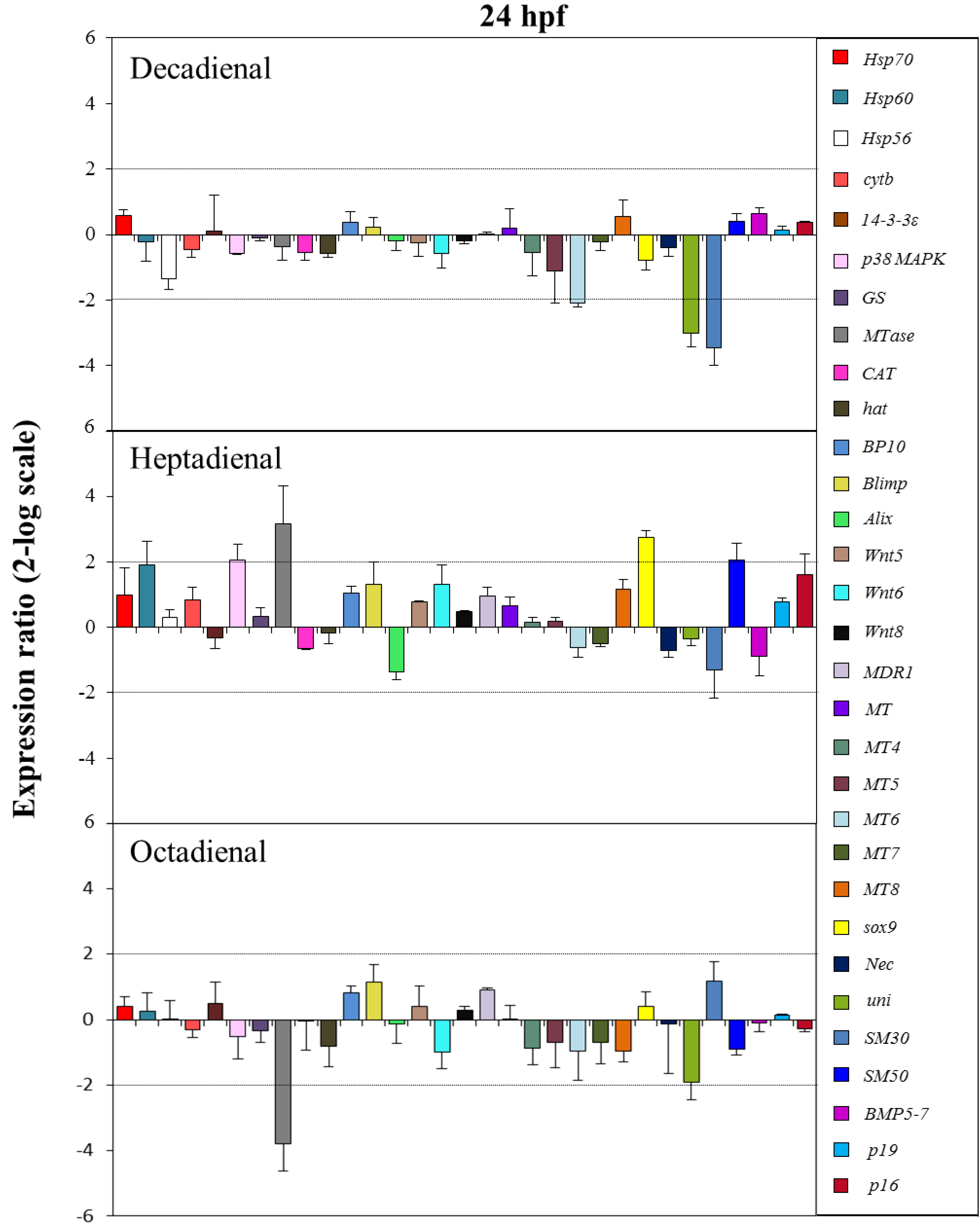
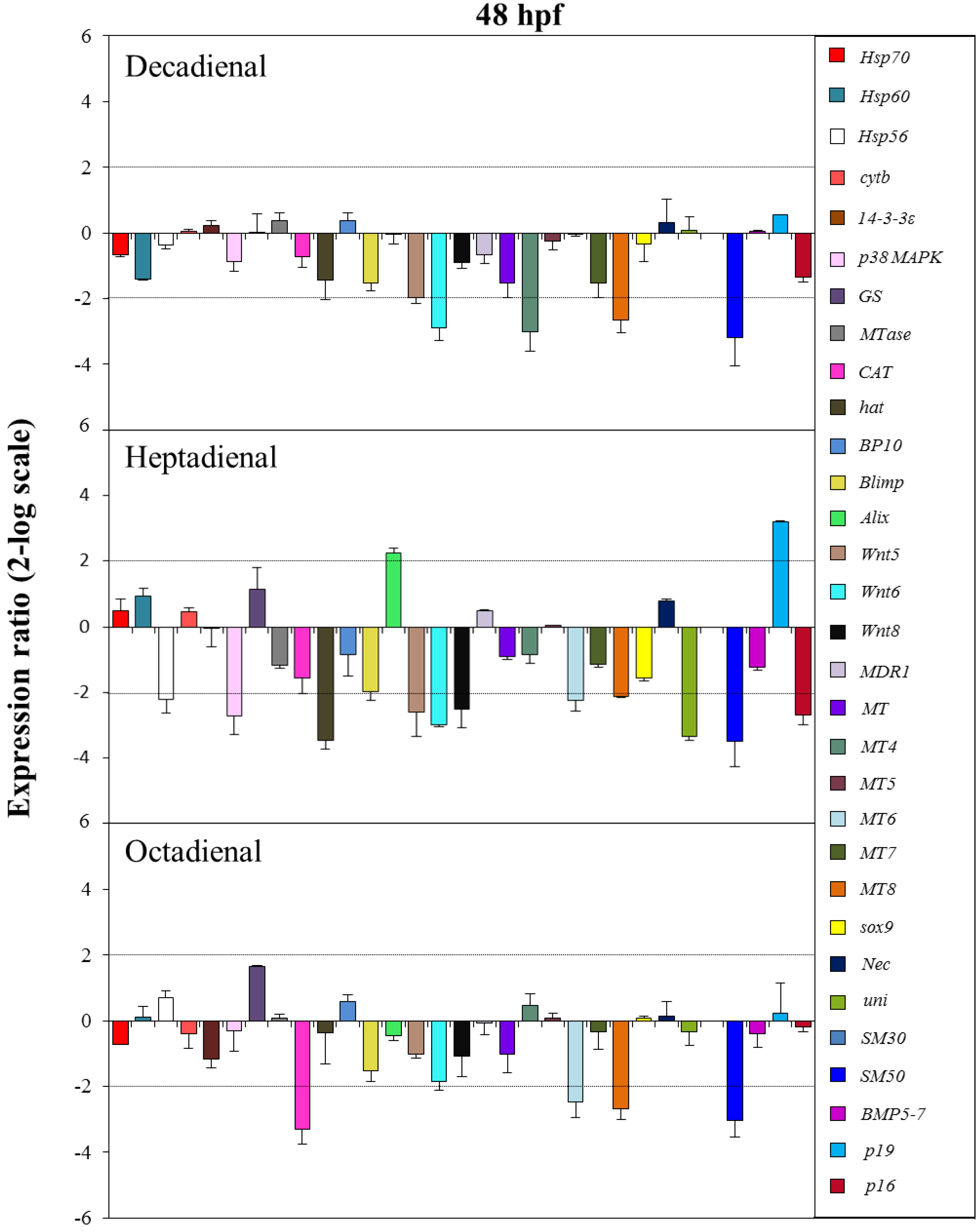
3. Discussion
4. Experimental Section
4.1. Ethics Statement
4.2. Gamete Collection, Embryo Culture, Exposure to Aldehydes and Morphological Analysis
4.3. Post-Recovery Experiments
4.4. Addition of PUAs after Fertilization
4.5. RNA Extraction and cDNA Synthesis
4.6. Gene Expression by Real-Time qPCR
| Gene Name | Acronym | Accession Number | Primer | Sequence (5′→3′) | PCR Fragment (bp) |
|---|---|---|---|---|---|
| Pl-p19 | p19 | FR693764 | Pl_P19_F1 | GACAAGCTCGACATCAACAAG | 205 |
| Pl_P19_R1 | CTGGAGTCGATGCTGCATCATG | ||||
| Pl-p16 | p16 | FR693763 | Pl-p16 For | CGGGCAGCGATGACTCA | 104 |
| [14] | Pl-p16 Rev | AAATGCCATACCGCTCTTCTGT | |||
| Metallothionein 4 | MT4 | MT4 For | GCTCAAAATCTTCAACATGGCTAATGA | ||
| [21] | MT4 Rev | AGCACTTTCCAGTTTCACAACAAGC | |||
| Metallothionein 5 | MT5 | MT5 For | CGACTTTAGCTCAAATTCATCACCATG | ||
| [21] | MT5 Rev | TCCACAGCATTTACCATCCTTGC | |||
| Metallothionein 6 | MT6 | MT6 For | CACGATTTGTGCTCAATCCTTCAT | ||
| [21] | MT6 Rev | TTTGTGCATGATGTTCCACAGC | |||
| Metallothionein 7 | MT7 | MT7 For | CGTCAAGAGATCAAAATCATCAACCA | ||
| [21] | MT7 Rev | ACAGCACTCGCCAGTAATACAGCAC | |||
| Metallothionein 8 | MT8 | MT8 For | GATGGTTGTCGTCGCTCCTAACA | ||
| [21] | MT8 Rev | TCAAGAAAGGCTGGTATCAAATCTGAC | |||
| Blimp | Blimp | HQ322503 | Pl_Blimp1_For | CTGTCTACTCCATGCCGTCC | 161 |
| Pl_Blimp1_Rev | GCCTCCTGCTTCAGATCAGC | ||||
| ALG-2 interacting protein X/1 | Alix | HE646599 | AL1500for | TACCAGACCATTCTCAACAAT | 110 |
| [15] | AL1610rev | TGCTATTTCCGCTTCGCTTTT | |||
| Multi drug resistance protein 1 | MDR1 | JQ793791 | Pl_MDR1_F2 | GTCAAGGTACTCAATGGGGTC | 158 |
| Pl_MDR1_Rev | CGGATGTCAATGCCATCAATC | ||||
| Wnt 5 | Wnt5 | HM449806 | Pl_Wnt5_F2 | CACCCAGCCCCTGTGCAGTG | 135 |
| Pl_Wnt5_Rev | CTGCAGTTCCACCTCCTATTC | ||||
| Wnt 6 | Wnt6 | HQ322504 | Pl_Wnt6_For | CGAATCTGCCGACGATCACG | 164 |
| Pl_Wnt6_For | GCATTGTCGTACAGTTCCACC | ||||
| Wnt 8 | Wnt8 | HM449816 | Pl_Wnt8_For | CTGTAAGTGTCATGGCGTCTC | 197 |
| Pl_Wnt8_For | GAGCGAATCGGAGATGACGG | ||||
| Catalase | CAT | SPU_000281.1 | Sp-CAT_F1 | GACTTCGTCTTCACCGACGAG | 156 |
| Sp-CAT_R1 | GACTCAAAGGGTGCAGCCTTG |
5. Conclusions
Abbreviations
| FSW | filtered sea water |
| hpf | hours post fertilization |
| µL | microliter |
| µM | micromolar |
| min | minutes |
| mM | millimolar |
| nm | nanometer |
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Caldwell, G.S. The Influence of Bioactive Oxylipins from Marine Diatoms on Invertebrate Reproduction and Development. Mar. Drugs 2009, 7, 367–400. [Google Scholar] [CrossRef]
- Ianora, A.; Miralto, A. Toxigenic effects of diatoms on grazers, phytoplankton and other microbes: A review. Ecotoxicology 2010, 19, 493–511. [Google Scholar] [CrossRef]
- Miralto, A.; Barone, G.; Romano, G.; Poulet, S.A.; Ianora, A.; Russo, G.L.; Buttino, I.; Mazzarella, G.; Laabir, M.; Cabrini, M.; et al. The insidious effect of diatoms on copepod reproduction. Nature 1999, 402, 173–176. [Google Scholar] [CrossRef]
- D’Ippolito, G.; Iadicicco, I.; Romano, G.; Fontana, A.; Iadicicco, O. Detection of short-chain aldehydes in marine organisms: The diatom Thalassiosira rotula. Tetrahedron Lett. 2002, 43, 6137–6140. [Google Scholar] [CrossRef]
- Wichard, T.; Poulet, S.A.; Halsband-Lenk, C.; Albaina, A.; Harris, R.; Liu, D.; Pohnert, G. Survey of the chemical defence potential of diatoms: Screening of fifty species for α,β,γ,δ-unsaturated aldehydes. J. Chem. Ecol. 2005, 31, 949–958. [Google Scholar] [CrossRef]
- Wendel, T.; Juttner, F. Lipoxygenase-mediated formation of hydrocarbons and unsaturated aldehydes in freshwater diatoms. Phytochemistry 1996, 41, 1445–1449. [Google Scholar] [CrossRef]
- Caldwell, G.S.; Olive, P.J.W.; Bentley, M.G. Inhibition of embryonic development and fertilization in broadcast spawning marine invertebrates by water soluble diatom extracts and the diatom toxin 2-trans,4-trans decadienal. Aquat. Toxicol. 2002, 60, 123–137. [Google Scholar] [CrossRef]
- Caldwell, G.S.; Bentley, M.G.; Olive, P.J.W. First evidence of sperm motility inhibition by the diatom aldehyde 2E,4E-decadienal. Mar. Ecol. Prog. Ser. 2004, 273, 97–108. [Google Scholar] [CrossRef]
- Romano, G.; Russo, G.L.; Buttino, I.; Ianora, A.; Miralto, A. A marine diatom-derived aldehyde induces apoptosis in copepod and sea urchin embryos. J. Exp. Biol. 2003, 206, 3487–3494. [Google Scholar] [CrossRef]
- Hansen, E.; Even, Y.; Geneviere, A.M. The alpha,beta,gamma,delta-unsaturated aldehyde 2-trans-4-trans-decadienal disturbs DNA replication and mitotic events in early sea urchin embryos. Toxicol. Sci. 2004, 81, 190–197. [Google Scholar] [CrossRef]
- Romano, G.; Miralto, A.; Ianora, A. Teratogenic Effects of diatom metabolites on sea urchin Paracentrotus lividus embryos. Mar. Drugs 2010, 8, 950–967. [Google Scholar] [CrossRef]
- Romano, G.; Costantini, M.; Buttino, I.; Ianora, A.; Palumbo, A. Nitric oxide mediates the stress response induced by diatom aldehydes in the sea urchin Paracentrotus lividus. PLoS One 2011, 6, e25980. [Google Scholar]
- Marrone, V.; Piscopo, M.; Romano, G.; Ianora, A.; Palumbo, A.; Costantini, M. Defensome against toxic diatom aldehydes in the sea urchin Paracentrotus lividus. PLoS One 2012, 7, e31750. [Google Scholar]
- Costa, C.; Karakostis, K.; Zito, F.; Matranga, V. Phylogenetic analysis and expression patterns of p16 and p19 in Paracentrotus lividus embryos. Dev. Genes Evol. 2012, 222, 245–251. [Google Scholar] [CrossRef]
- Romancino, D.P.; Anello, L.; Morici, G.; d’Azzo, A.; Bongiovanni, A.; Di Bernardo, M. Identification and characterization of PlAlix, the Alix homologue from the Mediterranean sea urchin Paracentrotus lividus. Dev. Growth Differ. 2013, 55, 237–246. [Google Scholar] [CrossRef] [Green Version]
- Lhomond, G.; McClay, D.R.; Gache, C.; Croce, J.C. Frizzled 1/2/7 signaling directs β-catenin nuclearisation and initiates andoderm specification in macromeres during sea urchin embryogenesis. Development 2012, 139, 816–825. [Google Scholar] [CrossRef]
- McIntyre, D.C.; Seay, N.W.; Croce, J.C.; McClay, D.R. Short-range Wnt5 signaling initiates specification of sea urchin posterior ectoderm. Development 2013, 140, 4881–4889. [Google Scholar] [CrossRef]
- Croce, J.; Range, R.; Wu, S.Y.; Miranda, E.; Lhomond, G.; Peng, J.C.; Lepage, T.; McClay, D.R. Wnt6 activates endoderm in the sea urchin gene regulatory network. Development 2011, 138, 3297–3306. [Google Scholar]
- Hammond, L.M.; Hofmann, G.E. Early developmental gene regulation in Strongylocentrotus purpuratus embryos in response to elevated CO2 seawater conditions. J. Exp. Biol. 2012, 215, 2445–2454. [Google Scholar] [CrossRef]
- Bošnjak, I.; Zaja, R.; Klobučar, R.S.; Sver, L.; Franekić, J.; Smital, T. Identification of ABC transporter genes in gonad tissue of two mediterranean sea urchin species: Black, Arbacia lixula L., and rocky, Paracentrotus lividus L. Bull. Environ. Contam. Toxicol. 2013, 91, 415–419. [Google Scholar] [CrossRef]
- Ragusa, M.A.; Costa, S.; Gianguzza, M.; Roccheri, M.C.; Gianguzza, F. Effects of cadmium exposure on sea urchin development assessed by SSH and RT-qPCR: Metallothionein genes and their differential induction. Mol. Biol. Rep. 2013, 40, 2157–2167. [Google Scholar] [CrossRef]
- Pohnert, G. Wound-activated chemical defense in unicellular planktonic algae. Angew. Chem. Int. Ed. 2000, 39, 4352–4354. [Google Scholar] [CrossRef]
- Wichard, T.; Gerecht, A.; Boersma, M.; Poulet, S.A.; Wiltshire, K.; Pohnert, G. Lipid and fatty acid composition of diatoms revisited: Rapid wound-activated change of food quality parameters influences herbivorous copepod reproductive success. Chembiochem 2007, 8, 1146–1153. [Google Scholar] [CrossRef] [Green Version]
- Vidoudez, C.; Nejstgaard, J.C.; Jakobsen, H.H.; Pohnert, G. Dynamics of dissolved and particulate polyunsaturated aldehydes in mesocosms inoculated with different densities of the diatom Skeletonema marinoi. Mar. Drugs 2011, 9, 345–358. [Google Scholar] [CrossRef]
- Buttino, I.; De Rosa, G.; Carotenuto, Y.; Mazzella, M.; Ianora, A.; Esposito, F.; Vitiello, V.; Quaglia, F.; La Rotonda, M.I.; Miralto, A. Aldehyde-encapsulating liposomes impair marine grazer survivorship. J. Exp. Biol. 2008, 211, 1426–1433. [Google Scholar] [CrossRef]
- Ceballos, S.; Ianora, A. Different diatoms induce contrasting effects on the reproductive success of the copepod Temora stylifera. J. Exp. Mar. Bio. Ecol. 2003, 294, 189–202. [Google Scholar] [CrossRef]
- Taylor, R.L.; Caldwell, G.S.; Dunstan, H.J.; Bentley, M.G. Short-term impacts of polyunsaturated aldehyde-producing diatoms on the harpacticoid copepod, Tisbe holothuriae. J. Exp. Mar. Biol. Ecol. 2007, 341, 60–69. [Google Scholar] [CrossRef]
- Lauritano, C.; Borra, M.; Carotenuto, Y.; Biffali, E.; Miralto, A.; Procaccini, G.; Ianora, A. First molecular evidence of diatom effects in the copepod Calanus helgolandicus. J. Exp. Mar. Biol. Ecol. 2011, 404, 79–86. [Google Scholar] [CrossRef]
- Lauritano, C.; Carotenuto, Y.; Miralto, A.; Procaccini, G.; Ianora, A. Copepod population-specific response to a toxic diatom diet. PLoS One 2012, 7, e47262. [Google Scholar]
- Aluigi, M.G.; Angelini, C.; Corte, G.; Falugi, C. The sea urchin, Paracentrotus lividus, embryo as a “bioethical” model for neurodevelopmental toxicity testing: Effects of diazinon on the intracellular distribution of OTX2-like proteins. Cell Biol. Toxicol. 2008, 24, 587–601. [Google Scholar] [CrossRef]
- Pinsino, A.; Matranga, V.; Trinchella, F.; Roccheri, M.C. Sea urchin embryos as an in vivo model for the assessment of manganese toxicity: Developmental and stress response effects. Ecotoxicology 2010, 19, 555–562. [Google Scholar] [CrossRef]
- Russo, R.; Bonaventura, R.; Zito, F.; Schroder, H.C.; Muller, I.; Muller, W.E.G.; Matranga, V. Stress to cadmium monitored by metallothionein gene induction in Paracentrotus lividus embryos. Cell Stress Chaperones 2003, 8, 232–241. [Google Scholar] [CrossRef]
- Bonaventura, R.; Poma, V.; Costa, C.; Matranga, V. UVB radiation prevents skeleton growth and stimulates the expression of stress markers in sea urchin embryos. Biochem. Biophys. Res. Commun. 2005, 328, 150–157. [Google Scholar] [CrossRef]
- Bonaventura, R.; Zito, F.; Costa, C.; Giarrusso, S.; Celi, F.; Matranga, V. Stress response gene activation protects sea urchin embryos exposed to X-rays. Cell Stress Chaperones 2011, 16, 681–687. [Google Scholar] [CrossRef]
- Lister, K.N.; Lamare, M.D.; Burritt, D.J. Sea ice protects the embryos of the Antarctic sea urchin Sterechinus neumayeri from oxidative damage due to naturally enhanced levels of UV-B radiation. J. Exp. Biol. 2010, 213, 1967–1975. [Google Scholar] [CrossRef]
- Lister, K.N.; Lamare, M.D.; Burritt, D.J. Oxidative damage in response to natural levels of UV-B radiation in larvae of the tropical sea urchin Tripneustes gratilla. Photochem. Photobiol. 2010, 86, 1091–1098. [Google Scholar] [CrossRef]
- Adams, N.L.; Campanale, J.P.; Foltz, K.R. Proteomic responses of sea urchin embryos to stressful ultraviolet radiation. Integr. Comp. Biol. 2012, 52, 665–680. [Google Scholar] [CrossRef]
- Byrne, M. Global change ecotoxicology: Identification of early life history bottlenecks in marine invertebrates, variable species responses and variable experimental approaches. Mar. Environ. Res. 2012, 76, 3–15. [Google Scholar] [CrossRef]
- Falugi, C.; Aluigi, M.G.; Chiantore, M.C.; Privitera, D.; Ramoino, P.; Gatti, M.A.; Fabrizi, A.; Pinsino, A.; Matranga, V. Toxicity of metal oxide nanoparticles in immune cells of the sea urchin. Mar. Environ. Res. 2012, 76, 114–121. [Google Scholar] [CrossRef]
- Roccheri, M.C.; Agnello, M.; Bonaventura, R.; Matranga, V. Cadmium induces the expression of specific stress proteins in sea urchin embryos. Biochem. Biophys. Res. Commun. 2004, 321, 80–87. [Google Scholar] [CrossRef] [Green Version]
- Diller, K.R. Stress protein expression kinetics. Annu. Rev. Biomed. Eng. 2006, 8, 403–424. [Google Scholar] [CrossRef]
- Rhee, J.-S.; Raisuddin, S.; Lee, K.-W.; Seo, J.S.; Ki, J.-S.; Kim, I.-C.; Park, H.G.; Lee, J.-S. Heat shock protein (Hsp) gene responses of the intertidal copepod Tigriopus japonicus to environmental toxicants. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2009, 149, 104–112. [Google Scholar] [CrossRef]
- Campos, A.C.E.; Molognoni, F.; Melo, F.H.M.; Galdieri, L.C.; Carneiro, C.R.W.; D’Almeida, V.; Correa, M.; Jasiulionis, M.G. Oxidative stress modulates DNA methylation during melanocyte anchorage blockade associated with malignant transformation. Neoplasia 2007, 9, 1111–1121. [Google Scholar] [CrossRef]
- Pinsino, A.; Roccheri, M.C.; Matranga, V. Manganese overload affects p38 MAPK phosphorylation and metalloproteinase activity during sea urchin embryonic development. Mar. Environ. Res. 2014, 93, 64–69. [Google Scholar] [CrossRef]
- Lepage, T.; Sardet, C.; Gache, C. Spatial expression of the hatching enzyme gene in the sea urchin embryo. Dev. Biol. 1992, 150, 23–32. [Google Scholar] [CrossRef]
- Ghiglione, C.; Lhomond, G.; Lepage, T.; Gache, C. Structure of the sea urchin hatching enzyme gene. Eur. J. Biochem. 1994, 219, 845–854. [Google Scholar] [CrossRef]
- Duboc, V.; Röttinger, E.; Lapraz, F.; Besnardeau, L.; Lepage, T. Left-right asymmetry in the sea urchin embryo is regulated by nodal signaling on the right side. Dev. Cell 2005, 9, 147–158. [Google Scholar] [CrossRef]
- Shipp, L.E.; Hamdoun, A. ATP-binding cassette (ABC) transporter expression and localization in sea urchin development. Dev. Dyn. 2012, 241, 1111–1124. [Google Scholar] [CrossRef]
- Nemer, M.; Rondinelli, E.; Infante, D.; Infante, A.A. Polyubiquitin RNA characteristics and conditional induction in sea urchin embryos. Dev. Biol. 1991, 145, 255–265. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. Available online: http://www.ncbi.nlm.nih.gov (accessed on 20 January 2013).
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef]
- Vanaverbeke, J.; Franco, M.A.; van Oevelen, D.; Moodley, L.; Provoost, P.; Steyaert, M.; Soetaert, K.; Vincx, M. Benthic responses to sedimentation of phytoplankton on the Belgian Continental Shelf. In Current Status of Eutrophication in the Belgian Coastal Zone; Presses Universitaires de Bruxelles: Bruxelles, Belgium, 2006; pp. 73–90. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Varrella, S.; Romano, G.; Ianora, A.; Bentley, M.G.; Ruocco, N.; Costantini, M. Molecular Response to Toxic Diatom-Derived Aldehydes in the Sea Urchin Paracentrotus lividus. Mar. Drugs 2014, 12, 2089-2113. https://doi.org/10.3390/md12042089
Varrella S, Romano G, Ianora A, Bentley MG, Ruocco N, Costantini M. Molecular Response to Toxic Diatom-Derived Aldehydes in the Sea Urchin Paracentrotus lividus. Marine Drugs. 2014; 12(4):2089-2113. https://doi.org/10.3390/md12042089
Chicago/Turabian StyleVarrella, Stefano, Giovanna Romano, Adrianna Ianora, Matt G. Bentley, Nadia Ruocco, and Maria Costantini. 2014. "Molecular Response to Toxic Diatom-Derived Aldehydes in the Sea Urchin Paracentrotus lividus" Marine Drugs 12, no. 4: 2089-2113. https://doi.org/10.3390/md12042089