Complex Toxin Profile of French Mediterranean Ostreopsis cf. ovata Strains, Seafood Accumulation and Ovatoxins Prepurification

 , ,
, ,  and
and
Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
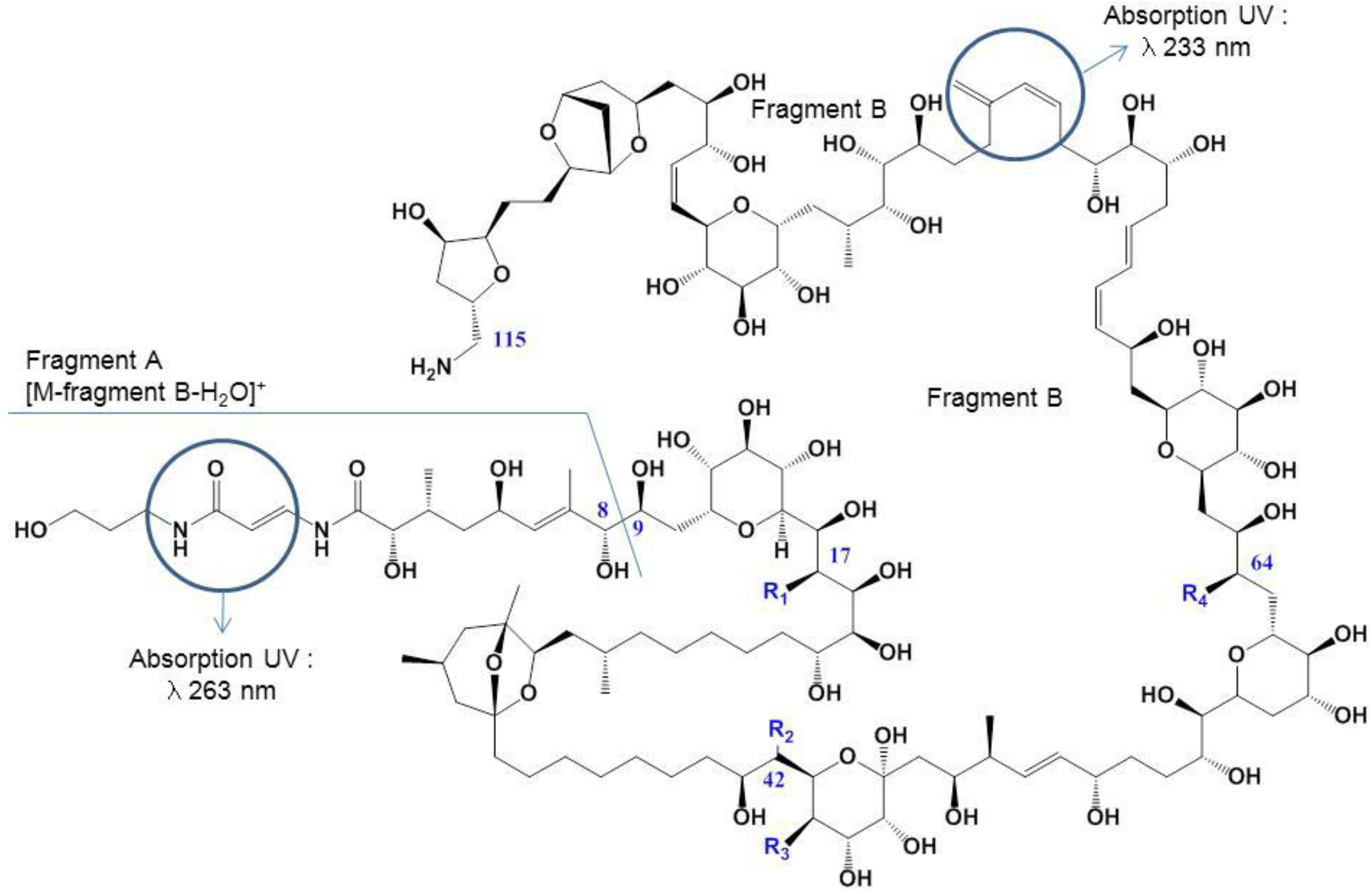
| Toxins | Elementary Formulae | Differences with PLTX | MW Da |
|---|---|---|---|
| Palytoxin | C129H223N3O54 | 2678.48 | |
| Ovatoxin-a | C129H223N3O52 | −2 O | 2646.49 |
| Ovatoxin-b | C131H227N3O53 | +C2H4; −1 O | 2690.52 |
| Ovatoxin-c | C131H227N3O54 | +C2H4 | 2706.51 |
| Ovatoxin-d/-e | C129H223N3O53 | −1 O | 2662.49 |
| Ovatoxin-f | C131H227N3O52 | +C2H4; −2 O | 2674.52 |

2. Results and Discussion
2.1. Ostreopsis cf. ovata Blooms and Accumulation in Marine Organisms
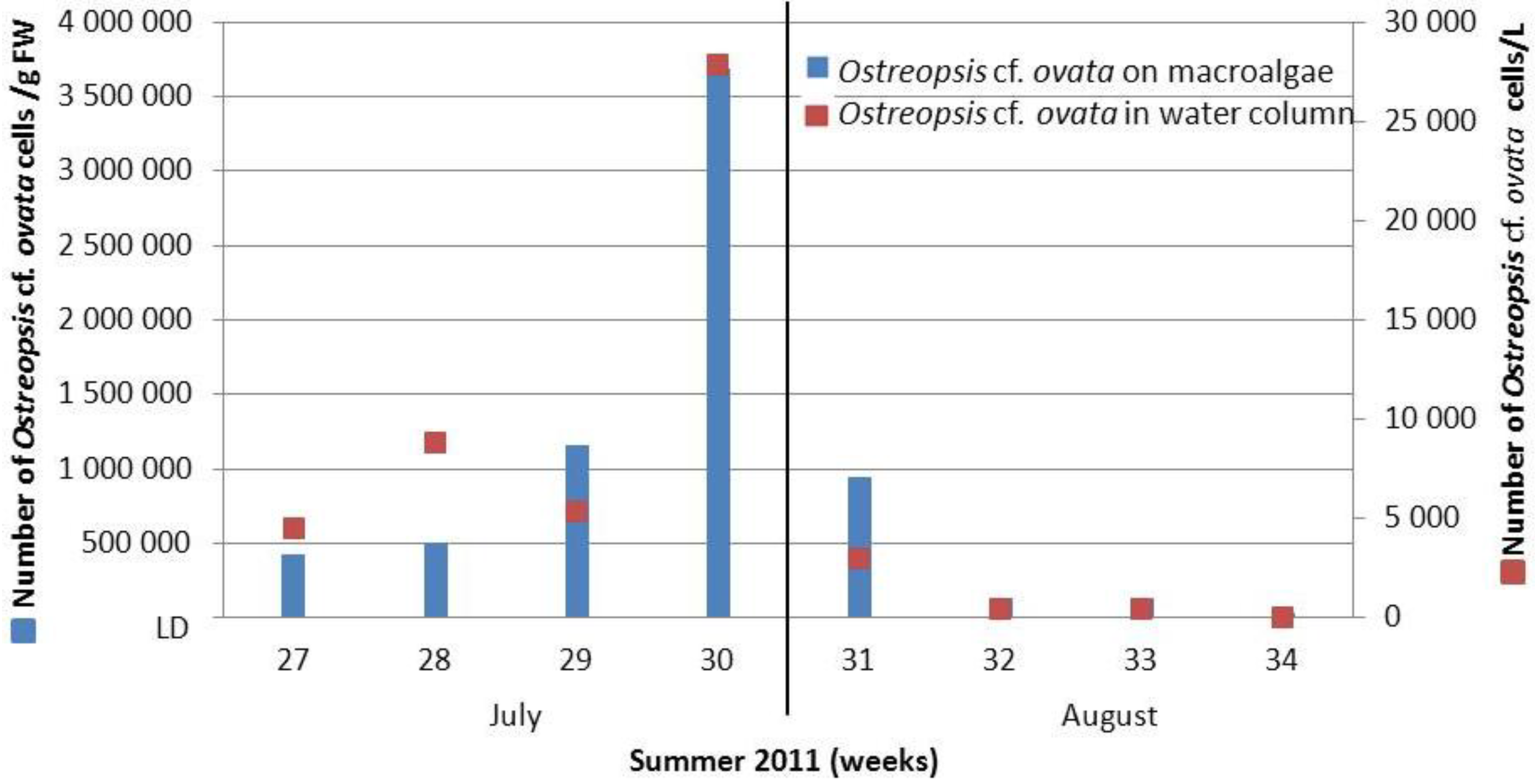
2.1.1. Recurrence of Ostreopsis cf. ovata Bloom on the French Mediterranean Coast


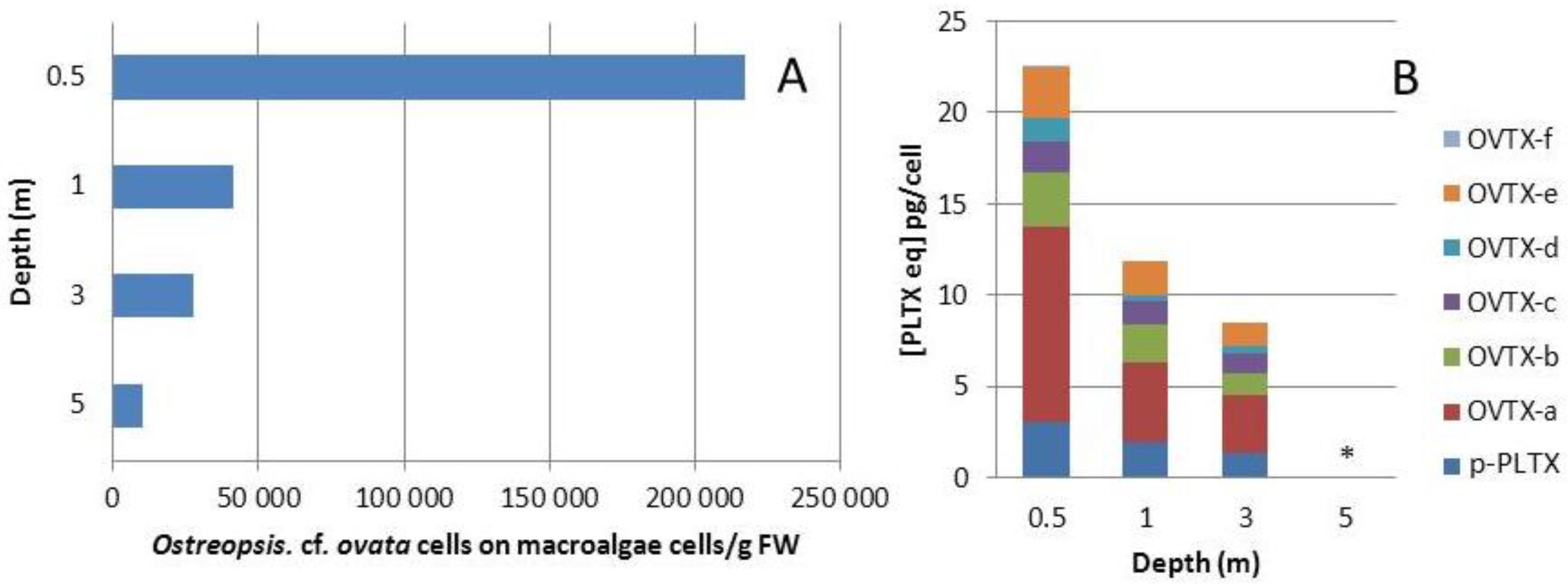
2.1.2. Influence of Depth on Toxin and Cell Concentration in the Field

2.1.3. Accumulation of PLTX-Group in Marine Organisms
| Whole Flesh Sea Urchin Paracentrotus Lividus | By LC-MS/MS μg·kg−1 | By Hemolytic Test μg·kg−1 |
| 231 | 247 | |
| 309 | 270 | |
| 205 | 201 | |
| 215 | 179 |
2.2. Ostreopsis cf. ovata Culture
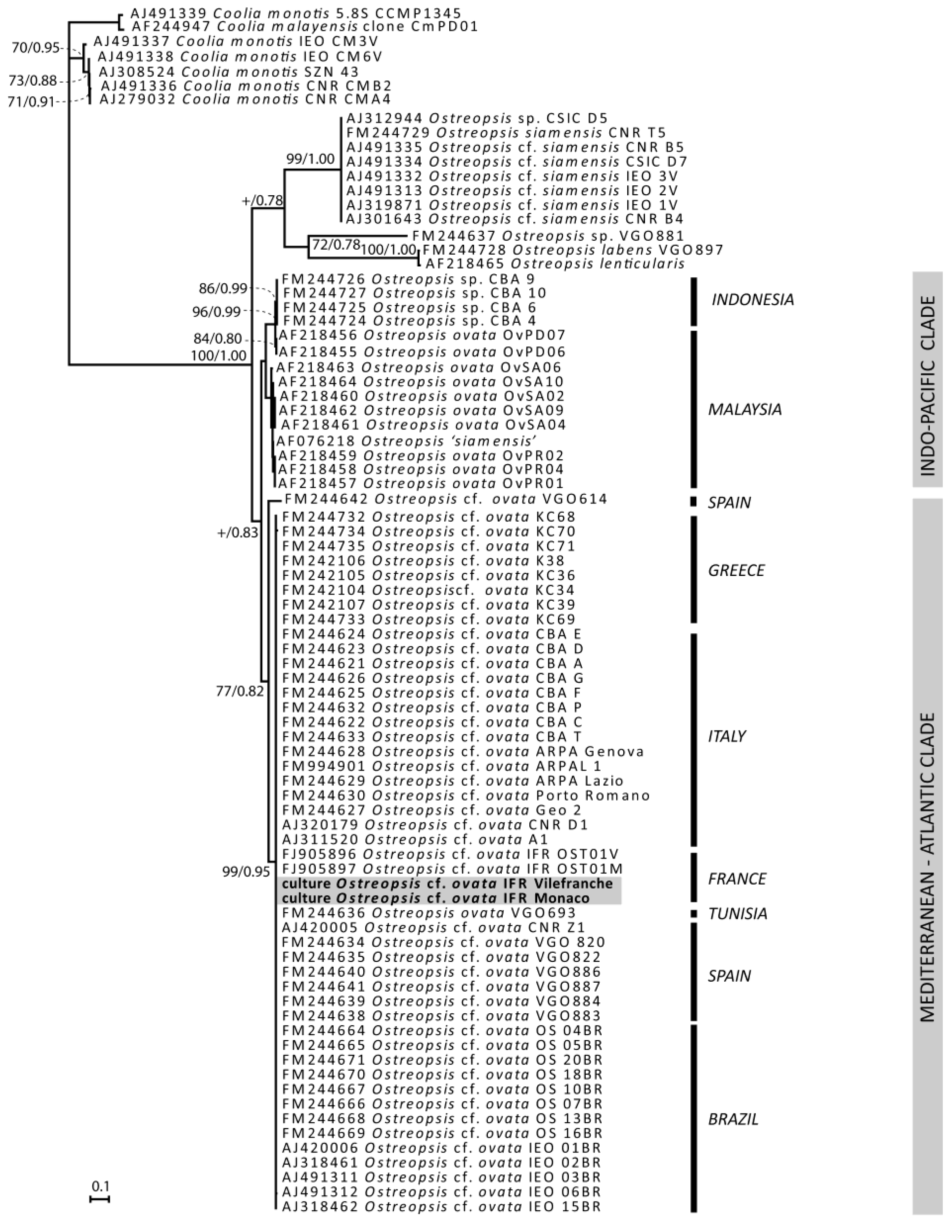
2.2.1. Genetic Characterization of Ostreopsis Strains
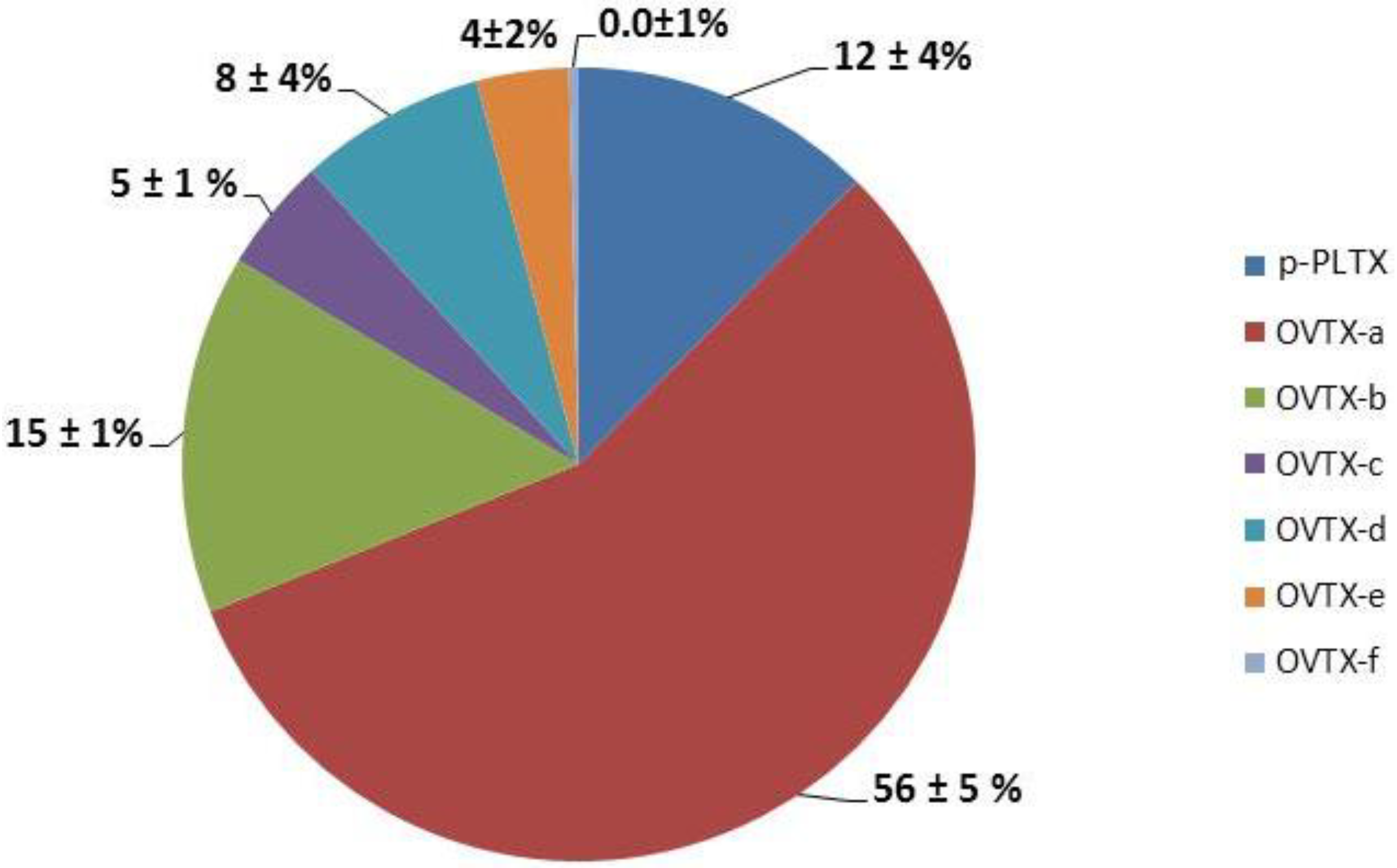
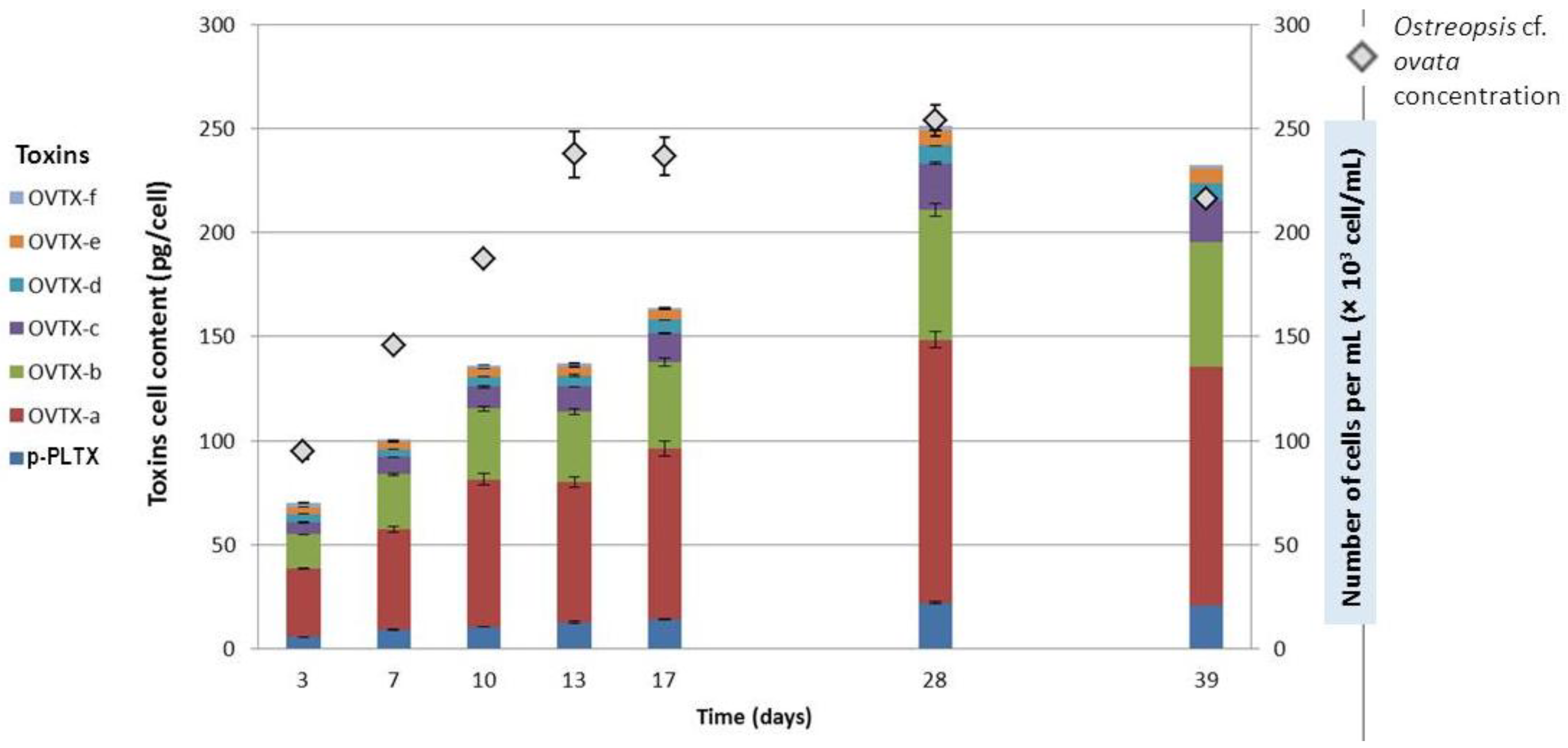
2.2.2. Cell Densities in Sea Water and Toxin Profiles of Ostreopsis cf. ovata Strains

2.2.3. Toxin Content as a Function of Growth Stage

2.3. Purification of Ovatoxins
2.3.1. Preparation of Ostreopsis cf. ovata Extract
2.3.2. Pre-Purification of Ostreopsis cf. ovata Extract
2.3.2.1. Pre-Purification of Ostreopsis cf. ovata Extract by Chemical Interaction Separation
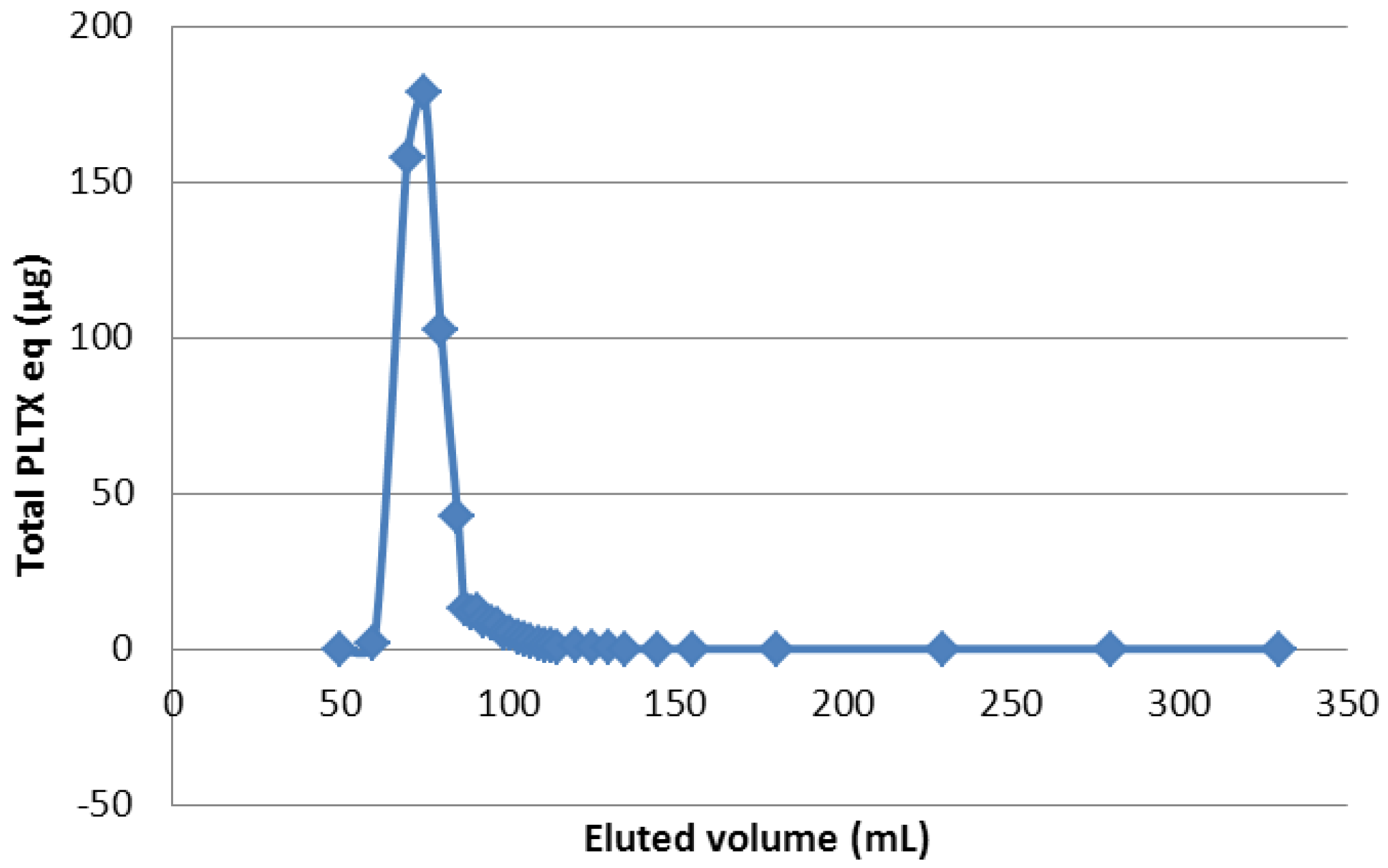
2.3.2.2. Pre-Purification of Ostreopsis cf. ovata Extract by Size Exclusion Separation

| Step | Weight (mg) | Toxin Quantity (mg) | Toxin Percentage (%) |
|---|---|---|---|
| Cell Extract | 770 | 3.10 | 0.4 |
| LH20 (Fraction of Interest) | 54 | 2.76 | 5.1 |
3. Experimental Section
3.1. Sampling Site and Collection of Marine Organisms
3.1.1. Cell Abundance
3.1.2. Isolation and Maintenance of Cultures of Ostreopsis cf. ovata
3.1.3. Hemolytic Assay
3.2. Phylogeny
3.2.1. DNA Amplification and Sequencing
3.2.2. Sequences Alignment and Phylogenetic Analysis
3.3. Evaluation of Cell Densities in Culture Media and Toxin Profile of Ostreopsis Strains
 where N0 and N1 are cell density values at time t0 and t1. (ii) the other one was used for extraction of toxin content without addition of HCl. Samples were harvested in triplicate, centrifuged at 3000× g over 15 min and the supernatant was discarded. Cell pellets were stored at −20 °C until extraction and analyzed by LC–MS/MS. (see Section 3.4.2. Extraction from cells). The rest of the biomass was collected and frozen to use for purification.
where N0 and N1 are cell density values at time t0 and t1. (ii) the other one was used for extraction of toxin content without addition of HCl. Samples were harvested in triplicate, centrifuged at 3000× g over 15 min and the supernatant was discarded. Cell pellets were stored at −20 °C until extraction and analyzed by LC–MS/MS. (see Section 3.4.2. Extraction from cells). The rest of the biomass was collected and frozen to use for purification.3.4. Extraction Procedure
3.4.1. Chemicals
3.4.2. Toxins Extraction from Cells
3.4.3. Toxins Extraction from the Marine Organism
3.5. LC-MS/MS and LC-MS Analysis
| Toxins | [M + 2H]2+→Part A | [M + 2H − H2O]2+→Part A | [M + 3H − H2O]3+→Part A |
|---|---|---|---|
| p-PLTX | 1340.3→327.3 | 1331.3→327.3 | 887.8→327.3 |
| OVTX-a | 1324.3→327.3 | 1315.3→327.3 | 877.2→327.3 |
| OVTX-b | 1346.3→371.2 | 1337.3→371.2 | 891.8→371.2 |
| OVTX-c | 1354.3→371.2 | 1345.3→371.2 | 897.2→371.2 |
| OVTX-d | 1332.3→327.3 | 1323.3→327.3 | 882.5→327.3 |
| OVTX-e | 1332.3→343.2 | 1323.3→343.2 | 882.5→343.2 |
| OVTX-f | 1338.3→327.3 | 1329.3→327.3 | 886.5→327.3 |
3.6. Extraction and Pre-Purification of Ovatoxins
3.6.1. Evaluation of PLTX-Group Toxins Released in Culture Media
3.6.2. Pre-Purification of Ostreopsis cf. ovata Extract by Chemical Properties
3.6.3. Pre-Purification of Ostreopsis cf. ovata Extract by Chemical Separation
3.6.3.1. Experiment 1: Filtration on Membranes
3.6.3.2. Experiment 2: LH20 Chromatography
4. Conclusions
Supplementary Files
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Moore, R.E.; Scheuer, P.J. Palytoxin: A new marine toxin from a coelenterate. Science 1971, 172, 495–498. [Google Scholar]
- Moore, R.E.; Bartolini, G. Structure of palytoxin. J. Am. Chem. Soc. 1981, 103, 2491–2494. [Google Scholar] [CrossRef]
- Deeds, J.R.; Schwartz, M.D. Human risk associated with palytoxin exposure. Toxicon 2010, 56, 150–162. [Google Scholar]
- Gleibs, S.; Mebs, D.; Werding, B. Studies on the origin and distribution of palytoxin in a caribeean coral-reef. Toxicon 1995, 33, 1531–1537. [Google Scholar]
- Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; Grauso, L.; Tartaglione, L.; Florio, C.; Lorenzon, P.; de Bortoli, M.; et al. Stereostructure and biological activity of 42-hydroxy-palytoxin: A new palytoxin analogue from Hawaiian Palythoa subspecies. Chem. Res. Toxicol. 2009, 22, 1851–1859. [Google Scholar] [CrossRef]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Forino, M.; Grauso, L.; Tartaglione, L. A 4-decade-long (and still ongoing) hunt for palytoxins chemical architecture. Toxicon 2011, 57, 362–367. [Google Scholar] [CrossRef]
- Hirata, Y.; Uemura, D. Toxic compounds of Palythoa tuberculosa (Coelenterata), palytoxin and its analogs. Yakugaku ZasshiJ. Pharm. Soc. Jpn. 1985, 105, 1–10. [Google Scholar]
- Usami, M.; Satake, M.; Ishida, S.; Inoue, A.; Kan, Y.; Yasumoto, T. Palytoxin analogs from the dinoflagellate Ostreopsis siamensis. J. Am. Chem. Soc. 1995, 117, 5389–5390. [Google Scholar] [CrossRef]
- Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; Grauso, L.; Tartaglione, L.; Guerrini, F.; Pistocchi, R. Complex palytoxin-like profile of Ostreopsis ovata. Identification of four new ovatoxins by high-resolution liquid chromatography/mass spectrometry. Rapid Commun. Mass Spectrom. 2010, 24, 2735–2744. [Google Scholar] [CrossRef]
- Lenoir, S.; Ten-Hage, L.; Turquet, J.; Quod, J.P.; Bernard, C.; Hennion, M.C. First evidence of palytoxin analogues from an Ostreopsis mascarenensis (Dinophyceae) benthic bloom in Southwestern Indian Ocean. J. Phycol. 2004, 40, 1042–1051. [Google Scholar] [CrossRef]
- Vila, M.; Garces, E.; Maso, M. Potentially toxic epiphytic dinoflagellate assemblages on macroalgae in the NW Mediterranean. Aquat. Microbl Ecol. 2001, 26, 51–60. [Google Scholar] [CrossRef]
- Penna, A.; Vila, M.; Fraga, S.; Giacobbe, M.G.; Andreoni, F.; Riobo, P.; Vernesi, C. Characterization of Ostreopsis and Coolia (Dinophyceae) isolates in the western Mediterranean Sea based on morphology, toxicity and internal transcribed spacer 5.8s rDNA sequences. J. Phycol. 2005, 41, 212–225. [Google Scholar] [CrossRef]
- Mangialajo, L.; Bertolotto, R.; Cattaneo-Vietti, R.; Chiantore, M.; Grillo, C.; Lemee, R.; Melchiorre, N.; Moretto, P.; Povero, P.; Ruggieri, N. The toxic benthic dinoflagellate Ostreopsis ovata: Quantification of proliferation along the coastline of Genoa, Italy. Mar. Pollut. Bull. 2008, 56, 1209–1214. [Google Scholar] [CrossRef]
- Totti, C.; Accoroni, S.; Cerino, F.; Cucchiari, E.; Romagnoli, T. Ostreopsis ovata bloom along the Conero Riviera (northern Adriatic Sea): Relationships with environmental conditions and substrata. Harmful Algae 2010, 9, 233–239. [Google Scholar] [CrossRef]
- Taylor, F.J.R. A description of the benthic dinoflagellate associated with maitotoxin and ciguatoxin, including observations on Hawaiian material. In Toxic Dinoflagellate Blooms; Taylor, D.L., Seliger, H.H., Eds.; Elsevier North Holland: New York, NY, USA, 1979; pp. 71–76. [Google Scholar]
- Pagliara, P.; Caroppo, C. Toxicity assessment of Amphidinium carterae, Coolia cfr. monotis and Ostreopsis cfr. ovata (Dinophyta) isolated from the northern Ionian Sea (Mediterranean Sea). Toxicon 2012, 60, 1203–1214. [Google Scholar] [CrossRef]
- Faust, M.A. Observation of sand-dwelling toxic dinoflagellates (Dinophyceae) from widely differing sites, including two new species. J. Phycol. 1995, 31, 996–1003. [Google Scholar]
- Honsell, G.; Bonifacio, A.; de Bortoli, M.; Penna, A.; Battocchi, C.; Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Sosa, S.; Yasumoto, T.; et al. New Insights on Cytological and Metabolic Features of Ostreopsis cf. ovata Fukuyo (Dinophyceae): A Multidisciplinary Approach. PLoS One 2013, 8. [Google Scholar] [CrossRef]
- Pistocchi, R.; Pezzolesi, L.; Guerrini, F.; Vanucci, S.; Dell’aversano, C.; Fattorusso, E. A review on the effects of environmental conditions on growth and toxin production of Ostreopsis ovata. Toxicon 2011, 57, 421–428. [Google Scholar] [CrossRef]
- Vidyarathna, N.K.; Graneli, E. Physiological responses of Ostreopsis ovata to changes in N and P availability and temperature increase. Harmful Algae 2013, 21–22, 54–63. [Google Scholar] [CrossRef]
- Durando, P.; Ansaldi, F.; Oreste, P.; Moscatelli, P.; Marensi, L.; Grillo, C.; Gasparini, R.; Icardi, G.; Collaborative Group for the Ligurian Syndromic Algal Surveillance. Ostreopsis ovata and human health: Epidemiological and clinical features of respiratory syndrome outbreaks from a two-year syndromic surveillance, 2005–2006, in north-west Italy. Eurosurveillance 2007, 12, E070607.1. [Google Scholar]
- Rhodes, L.; Towers, N.; Briggs, L.; Munday, R.; Adamson, J. Uptake of palytoxin-like compounds by shellfish fed Ostreopsis siamensis (Dinophyceae). N. Z. J. Mar. Freshw. Res. 2002, 36, 631–636. [Google Scholar] [CrossRef]
- Mebs, D. Occurrence and sequestration of toxins in food chains. Toxicon 1998, 36, 1519–1522. [Google Scholar] [CrossRef]
- Aligizaki, K.; Katikou, P.; Milandri, A.; Diogene, J. Occurrence of palytoxin-group toxins in seafood and future strategies to complement the present state of the art. Toxicon 2011, 57, 390–399. [Google Scholar] [CrossRef]
- Amzil, Z.; Sibat, M.; Chomerat, N.; Grossel, H.; Marco-Miralles, F.; Lemee, R.; Nezan, E.; Sechet, V. Ovatoxin-a and Palytoxin Accumulation in Seafood in Relation to Ostreopsis cf. ovata Blooms on the French Mediterranean Coast. Mar. Drugs 2012, 10, 477–496. [Google Scholar] [CrossRef]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Grillo, C.; Melchiorre, N. Putative palytoxin and its new analogue, ovatoxin-a, in Ostreopsis ovata collected along the Ligurian coasts during the 2006 toxic outbreak. J. Am. Soc. Mass Spectrom. 2008, 19, 111–120. [Google Scholar] [CrossRef]
- Sechet, V.; Sibat, M.; Chomerat, N.; Nezan, E.; Grossel, H.; Lehebel-Peron, J.-B.; Jauffrais, T.; Ganzin, N.; Marco-Miralles, F.; Lemee, R.; et al. Ostreopsis cf. ovata in the French Mediterranean coast: Molecular characterisation and toxin profile. Cryptogam. Algol. 2012, 33, 89–98. [Google Scholar] [CrossRef]
- EFSA. Scientific Opinion on marine biotoxins in shellfish—Palytoxin group. EFSA J. 2009, 7, 1393–1430. [Google Scholar]
- Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Battocchi, C.; Crinelli, R.; Carloni, E.; Magnani, M.; et al. Unique toxin profile of a Mediterranean Ostreopsis cf. ovata strain: HR LC-MSn characterization of ovatoxin-f, a new palytoxin congener. Chem. Res. Toxicol. 2012, 25, 1243–1252. [Google Scholar] [CrossRef]
- Moore, R.E.; Woolard, F.X.; Bartolini, G. Periodate-oxidation of N-(para-bromobenzoyl)palytoxin. J. Am. Chem. Soc. 1980, 102, 7370–7372. [Google Scholar] [CrossRef]
- Ramos, V.; Vasconcelos, V. Palytoxin and Analogs: Biological and Ecological Effects. Mar. Drugs 2010, 8, 2021–2037. [Google Scholar] [CrossRef]
- Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; Grauso, L.; Tartaglione, L.; Guerrini, F.; Pezzolesi, L.; Pistocchi, R.; et al. Isolation and Structure Elucidation of Ovatoxin-a, the Major Toxin Produced by Ostreopsis ovata. J. Am. Chem. Soc. 2012, 134, 1869–1875. [Google Scholar] [CrossRef]
- Riobo, P.; Franco, J.M. Palytoxins: Biological and chemical determination. Toxicon 2011, 57, 368–375. [Google Scholar] [CrossRef]
- Artigas, P.; Gadsby, D.C. Ion occlusion/deocclusion partial reactions in individual palytoxin-modified Na/K pumps. Ann. N. Y. Acad. Sci. 2003, 986, 116–126. [Google Scholar] [CrossRef]
- Hilgemann, D.W. From a pump to a pore: How palytoxin opens the gates. Proc. Natl. Acad. Sci. USA 2003, 100, 386–388. [Google Scholar] [CrossRef]
- Habermann, E.; Ahnerthilger, G.; Chhatwal, G.S.; Beress, L. Delayed hemolytic action of palytoxin—general characteristics. Biochim. Biophys. Acta 1981, 649, 481–486. [Google Scholar] [CrossRef]
- Habermann, E.; Chhatwal, G.S. Ouabain inhibits the increase dur to palytoxin of cation permeability of erythrocytes. Naunyn-Schmiedebergs Arch. Pharmacol. 1982, 319, 101–107. [Google Scholar] [CrossRef]
- Tichadou, L.; Glaizal, M.; Armengaud, A.; Grossel, H.; Lemee, R.; Kantin, R.; Lasalle, J.-L.; Drouet, G.; Rambaud, L.; Malfait, P.; et al. Health impact of unicellular algae of the Ostreopsis genus blooms in the Mediterranean Sea: Experience of the French Mediterranean coast surveillance network from 2006 to 2009. Clin. Toxicol. 2010, 48, 839–844. [Google Scholar] [CrossRef]
- Cohu, S.; Mangialajo, L.; Thibaut, T.; Blanfune, A.; Lemee, R. Development of the benthic toxic dinoflagellate Ostreopsis cf. ovata in the NW Mediterranean Sea. Eur. J. Phycol. 2011, 46, 55–55. [Google Scholar]
- Cohu, S.; Mangialajo, L.; Thibaut, T.; Blanfune, A.; Marro, S.; Lemee, R. Proliferation of the toxic dinoflagellate Ostreopsis cf. ovata in relation to depth, biotic substrate and environmental factors in the North West Mediterranean Sea. Harmful Algae 2013, 24, 32–44. [Google Scholar] [CrossRef]
- Mangialajo, L.; Ganzin, N.; Accoroni, S.; Asnaghi, V.; Blanfune, A.; Cabrini, M.; Cattaneo-Vietti, R.; Chavanon, F.; Chiantore, M.; Cohu, S.; et al. Trends in Ostreopsis proliferation along the Northern Mediterranean coasts. Toxicon 2011, 57, 408–420. [Google Scholar]
- Bellocci, M.; Ronzitti, G.; Milandri, A.; Melchiorre, N.; Grillo, C.; Poletti, R.; Yasumoto, T.; Rossini, G.P. A cytolytic assay for the measurement of palytoxin based on a cultured monolayer cell line. Anal. Biochem. 2008, 374, 48–55. [Google Scholar] [CrossRef]
- Guerrini, F.; Pezzolesi, L.; Feller, A.; Riccardi, M.; Ciminiello, P.; Dell’Aversano, C.; Tartaglione, L.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; et al. Comparative growth and toxin profile of cultured Ostreopsis ovata from the Tyrrhenian and Adriatic Seas. Toxicon 2010, 55, 211–220. [Google Scholar] [CrossRef]
- Rossi, R.; Castellano, V.; Scalco, E.; Serpe, L.; Zingone, A.; Soprano, V. New palytoxin-like molecules in Mediterranean Ostreopsis cf. ovata (dinoflagellates) and in Palythoa tuberculosa detected by liquid chromatography-electrospray ionization time-of-flight mass spectrometry. Toxicon 2010, 56, 1381–1387. [Google Scholar] [CrossRef]
- Accoroni, S.; Colombo, F.; Pichierri, S.; Romagnoli, T.; Marini, M.; Battocchi, C.; Penna, A.; Totti, C. Ecology of Ostreopsis cf. ovata blooms in the northwestern Adriatic Sea. Cryptogam. Algol. 2012, 33, 191–198. [Google Scholar] [CrossRef]
- Cohu, S.; Lemee, R. Vertical distribution of the toxic epibenthic dinoflagellates Ostreopsis cf. ovata, Prorocentrum lima and Coolia monotis in the NW Mediterranean Sea. Cahiers Biol. Mar. 2012, 53, 373–380. [Google Scholar]
- Fraga, S.; Rodriguez, F.; Bravo, I.; Zapata, M.; Maranon, E. Review of the main ecological features affecting benthic dinoflagellate blooms. Cryptogam. Algol. 2012, 33, 171–179. [Google Scholar] [CrossRef]
- Bire, R.; Trotereau, S.; Lemee, R.; Delpont, C.; Chabot, B.; Aumond, Y.; Krys, S. Occurrence of palytoxins in marine organisms from different trophic levels of the French Mediterranean coast harvested in 2009. Harmful Algae 2013, 28, 10–22. [Google Scholar] [CrossRef]
- Pezzolesi, L.; Guerrini, F.; Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Pistocchi, R. Influence of temperature and salinity on Ostreopsis cf. ovata growth and evaluation of toxin content through HR LC-MS and biological assays. Water Res. 2012, 46, 82–92. [Google Scholar] [CrossRef]
- Penna, A.; Fraga, S.; Battocchi, C.; Casabianca, S.; Giacobbe, M.G.; Riobo, P.; Vernesi, C. A phylogeographical study of the toxic benthic dinoflagellate genus Ostreopsis Schmidt. J. Biogeogr. 2010, 37, 830–841. [Google Scholar] [CrossRef] [Green Version]
- Pistocchi, R.; Guerrini, F.; Pezzolesi, L.; Riccardi, M.; Vanucci, S.; Ciminiello, P.; Dell’Aversano, C.; Forino, M.; Fattorusso, E.; Tartaglione, L.; et al. Toxin levels and profiles in microalgae from the North-Western Adriatic Sea-15 years of studies on cultured species. Mar. Drugs 2012, 10, 140–162. [Google Scholar] [CrossRef]
- Nascimento, S.M.; Correa, E.V.; Menezes, M.; Varela, D.; Paredes, J.; Morris, S. Growth and toxin profile of Ostreopsis cf. ovata (Dinophyta) from Rio de Janeiro, Brazil. Harmful Algae 2012, 13, 1–9. [Google Scholar]
- Jauffrais, T.; Kilcoyne, J.; Sechet, V.; Herrenknecht, C.; Truquet, P.; Herve, F.; Berard, J.B.; Nulty, C.; Taylor, S.; Tillmann, U.; et al. Production and Isolation of Azaspiracid-1 and-2 from Azadinium spinosum Culture in Pilot Scale Photobioreactors. Mar. Drugs 2012, 10, 1360–1382. [Google Scholar] [CrossRef]
- Uchida, H.; Taira, Y.; Yasumoto, T. Structural elucidation of palytoxin analogs produced by the dinoflagellate Ostreopsis ovata IK2 strain by complementary use of positive and negative ion liquid chromatography/quadrupole time-of-flight mass spectrometry. Rapid Commun. Mass Spectrom. 2013, 27, 1999–2008. [Google Scholar] [CrossRef]
- Hwang, B.S.; Yoon, E.Y.; Kim, H.S.; Yih, W.; Park, J.Y.; Jeong, H.J.; Rho, J.-R.; Ostreol, A. A new cytotoxic compound isolated from the epiphytic dinollagellate Ostreopsis cf. ovata from the coastal waters of Jeju Island, Korea. Bioorgan. Med. Chem. Lett. 2003, 23, 3023–3027. [Google Scholar]
- Inuzuka, T.; Uemura, D.; Arimoto, H. The conformational features of palytoxin in aqueous solution. Tetrahedron 2008, 64, 7718–7723. [Google Scholar] [CrossRef]
- Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Yasumoto, T.; Battocchi, C.; Giacobbe, M.; Amorim, A.; et al. Investigation of toxin profile of Mediterranean and Atlantic strains of Ostreopsis cf. siamensis (Dinophyceae) by liquid chromatography-high resolution mass spectrometry. Harmful Algae 2013, 23, 19–27. [Google Scholar]
- Crinelli, R.; Carloni, E.; Giacomini, E.; Penna, A.; Dominici, S.; Battocchi, C.; Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Forino, M.; et al. Palytoxin and an Ostreopsis Toxin Extract Increase the Levels of mRNAs Encoding Inflammation-Related Proteins in Human Macrophages via p38 MAPK and NF-kappa B. PLoS One 2012, 7, e38139. [Google Scholar] [CrossRef]
- Selwood, A.I.; van Ginkel, R.; Harwood, D.T.; McNabb, P.S.; Rhodes, L.R.; Holland, P.T. A sensitive assay for palytoxins, ovatoxins and ostreocins using LC-MS/MS analysis of cleavage fragments from micro-scale oxidation. Toxicon 2012, 60, 810–820. [Google Scholar] [CrossRef]
- Guillard, R.R.L.; Smith, W.L.; Chanley, M.H. Culture of phytoplankton for feeding marine invertebrates. In Culture of Marine Invertebrates Animals; Plenum Press: New York, NY, USA, 1975; pp. 26–60. [Google Scholar]
- Bignami, G.S. A rapid and sensitive hemolysis neutralization assay for palytoxin. Toxicon 1993, 31, 817–820. [Google Scholar] [CrossRef]
- Riobo, P.; Paz, B.; Franco, J.M.; Vazquez, J.A.; Murado, M.A. Proposal for a simple and sensitive haemolytic assay for palytoxin Toxicological dynamics, kinetics, ouabain inhibition and thermal stability. Harmful Algae 2008, 7, 415–429. [Google Scholar] [CrossRef] [Green Version]
- Aligizaki, K.; Katikou, P.; Nikolaidis, G.; Panou, A. First episode of shellfish contamination by palytoxin-like compounds from Ostreopsis species (Aegean Sea, Greece). Toxicon 2008, 51, 418–427. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procodure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Edgar, R.C. Muscle: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5. [Google Scholar] [CrossRef] [Green Version]
- Hall, T. BioEdit; Ibis Biosciences: Carlsbad, CA, USA, 2011. [Google Scholar]
- Hall, T.A.B. A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Posada, D. jModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar]
- Felsenstein, J.J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Brissard, C.; Herrenknecht, C.; Séchet, V.; Hervé, F.; Pisapia, F.; Harcouet, J.; Lémée, R.; Chomérat, N.; Hess, P.; Amzil, Z. Complex Toxin Profile of French Mediterranean Ostreopsis cf. ovata Strains, Seafood Accumulation and Ovatoxins Prepurification. Mar. Drugs 2014, 12, 2851-2876. https://doi.org/10.3390/md12052851
Brissard C, Herrenknecht C, Séchet V, Hervé F, Pisapia F, Harcouet J, Lémée R, Chomérat N, Hess P, Amzil Z. Complex Toxin Profile of French Mediterranean Ostreopsis cf. ovata Strains, Seafood Accumulation and Ovatoxins Prepurification. Marine Drugs. 2014; 12(5):2851-2876. https://doi.org/10.3390/md12052851
Chicago/Turabian StyleBrissard, Charline, Christine Herrenknecht, Véronique Séchet, Fabienne Hervé, Francesco Pisapia, Jocelyn Harcouet, Rodolphe Lémée, Nicolas Chomérat, Philipp Hess, and Zouher Amzil. 2014. "Complex Toxin Profile of French Mediterranean Ostreopsis cf. ovata Strains, Seafood Accumulation and Ovatoxins Prepurification" Marine Drugs 12, no. 5: 2851-2876. https://doi.org/10.3390/md12052851