Quorum Quenching Agents: Resources for Antivirulence Therapy

College of Marine Life Sciences, Ocean University of China, Qingdao 266003, China
*
Author to whom correspondence should be addressed.
Mar. Drugs 2014, 12(6), 3245-3282; https://doi.org/10.3390/md12063245
Submission received: 29 January 2014
/
Revised: 7 May 2014
/
Accepted: 9 May 2014
/
Published: 30 May 2014
(This article belongs to the Special Issue Advances and New Perspectives in Marine Biotechnology)
Abstract
:The continuing emergence of antibiotic-resistant pathogens is a concern to human health and highlights the urgent need for the development of alternative therapeutic strategies. Quorum sensing (QS) regulates virulence in many bacterial pathogens, and thus, is a promising target for antivirulence therapy which may inhibit virulence instead of cell growth and division. This means that there is little selective pressure for the evolution of resistance. Many natural quorum quenching (QQ) agents have been identified. Moreover, it has been shown that many microorganisms are capable of producing small molecular QS inhibitors and/or macromolecular QQ enzymes, which could be regarded as a strategy for bacteria to gain benefits in competitive environments. More than 30 species of marine QQ bacteria have been identified thus far, but only a few of them have been intensively studied. Recent studies indicate that an enormous number of QQ microorganisms are undiscovered in the highly diverse marine environments, and these marine microorganism-derived QQ agents may be valuable resources for antivirulence therapy.
1. Introduction
Antibiotics are recognized as effective antimicrobial agents for curing diseases caused by pathogenic bacteria. Traditional antibiotics are bactericidal or bacteriostatic by targeting essential processes for bacterial growth including cell wall synthesis, DNA replication, RNA transcription and protein synthesis [1]. However, because of the life-or-death selective pressure imposed on the targeted pathogens, antibiotic-resistant strains are constantly emerging. The inappropriate and excessive use of antibiotics accelerates the emergence of antibiotic resistance. Unfortunately, in contrast to the rising levels of antibiotic resistance, the pace of novel antibiotic development has severely slowed in the preceding few decades. This problem leads to the urgent need for the development of new antimicrobial agents targeting virulence (toxin function and delivery, regulation of virulence expression and bacterial adhesion) rather than the essential processes of pathogenic microorganisms [1]. The so-called antivirulence therapy may impose less selective pressure on pathogenic microorganisms, and in theory, decrease resistance [1].
Quorum sensing (QS) is a process for bacteria to communicate, regulate gene expression and synchronize social behaviors, such as biofilm formation, bioluminescence and secretion of virulence factors [2,3]. Hitherto diverse molecular mechanisms of sophisticated QS have been unraveled [2]. In particular, it has been established that QS regulates the secretion of virulence factors in many pathogens, such as Pseudomonas aeruginosa, Erwinia carotovora, Vibrio spp. and Burkholderia spp. [2]. In addition, these pathogens use QS to regulate biofilm formation, which is a critical defense against antibacterial drugs or the immune system of a host. Because QS is not essential for the growth of bacteria, quenching QS (quorum quenching, QQ) in these pathogens would disarm virulence rather than kill the bacteria, which may considerably weaken the selective pressure imposed on the pathogens and delay the evolution of resistance to QQ drugs. All of these features make QS an ideal target for antivirulence therapy [1].
Here, we provide an overall summary of the uniformity and diversity of QS in microorganisms, as well as the QS circuits in some representative species. Two main QQ agents, i.e., small molecular QS inhibitors and macromolecular QQ enzymes, are discussed. The current status and trends of antivirulence therapy utilizing QQ resources from the marine environment are considered. Based on this knowledge, a critical appraisal and perspective of the resource for antivirulence in the environment is presented.
2. The Distribution of QS Systems in Microorganisms
The term QS was first proposed to describe the phenomenon that marine bacteria, i.e., V. fischeri and V. harveyi, use signaling molecules (autoinducers, AIs) as sensors of cell density so that the population as a whole may coordinate the social behavior of bioluminescence [4]. However, in addition to cell density, the concentration of AIs in the natural environment is determined by many other biotic and abiotic factors, such as the spatial distribution of cells and the diffusional characteristics of the environment [5,6,7,8,9]. Redfield [5] proposed a concept of diffusion sensing (DS) to challenge the previously assumed role of QS. She argued that cells employ AIs to assess diffusive properties of the environment, and thereby determine when to produce more costly secreted molecules, such as extracellular protease. This may be less effective in the environment with a high diffusion rate. Another concept of efficiency sensing (ES) was introduced in an attempt to unify both QS and DS [6]. In particular, it has been demonstrated that even a single cell may initiate QS-regulated behaviors by physical confinement [10,11], which provides support for the hypotheses of DS and ES. However, QS and DS are not diametrically opposed, and the utility of ES has been debated [8,12]. In addition, many other hypotheses emphasizing different factors have been suggested to compete or unite with QS [13]. However, Platt et al. [13] argued that the introduction of new hypotheses would lead to confusion rather than clarification. Therefore, it was suggested that the processes of QS could be viewed broadly with full awareness of the effects of environmental factors [13].
Despite the presence of distinct QS systems in different microorganisms, the fundamental processes are similar. Initially, AIs are synthesized by AI synthases and diffuse away. When the concentrations of AIs increase to a threshold, AIs are detected by receptors. Subsequently, the AI-bound receptors activate the expression of relevant genes, including AI synthase-encoding genes. This results in a positive feed-back loop for biosynthesis of AIs, which may be able to promote the synchrony among a population.
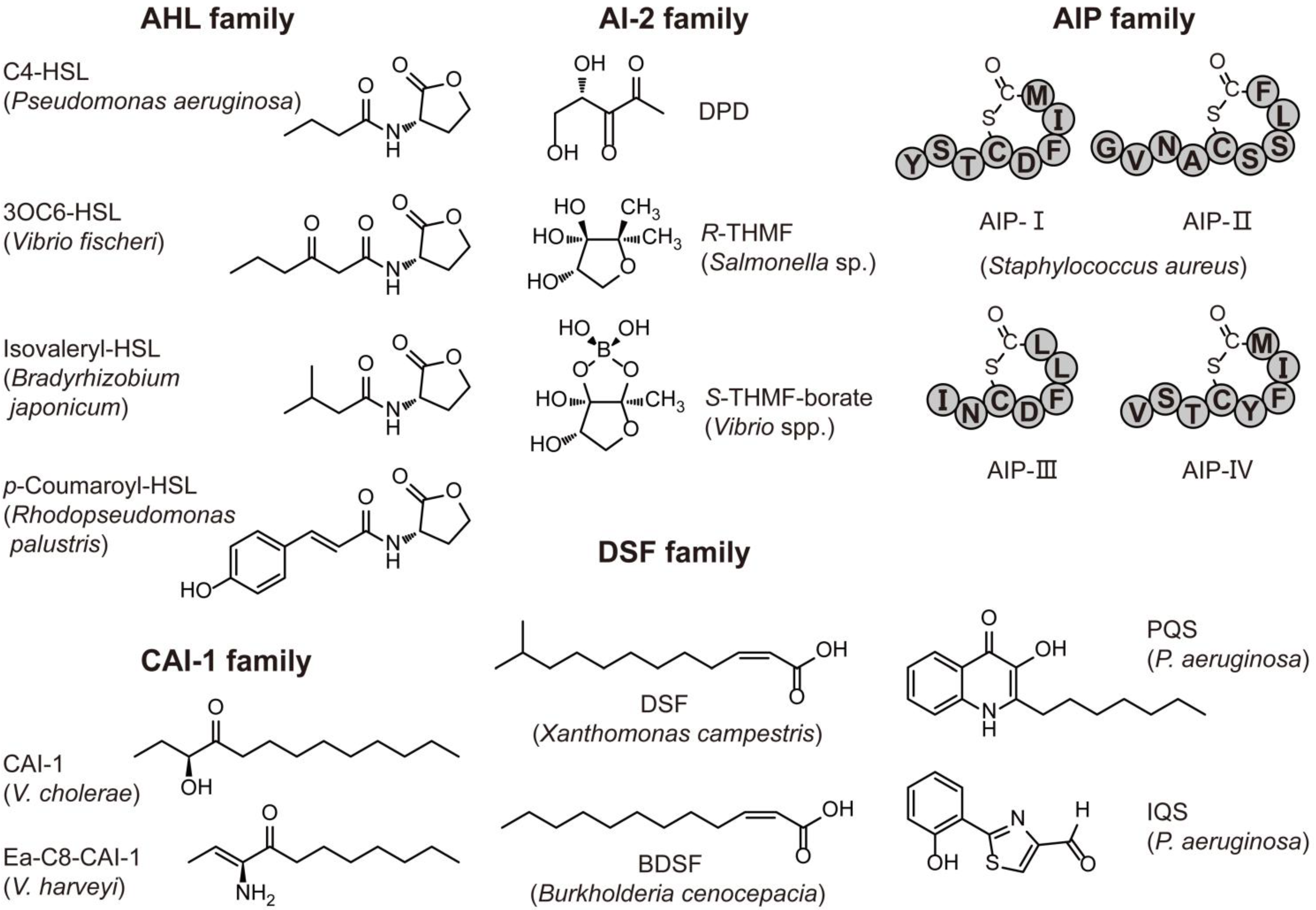
Increasing evidence has revealed that QS is prevalent in bacteria, fungi and archaea (Figure 1, Table 1). For example, it is widely accepted that N-acylhomoserine lactones (AHLs) and autoinducing peptides (AIPs) are mainly used by Gram-negative and Gram-positive bacteria for intraspecies communication, respectively. Autoinducter-2 (AI-2) signals are hypothesized to be used for interspecies communication because AI-2 production and the synthase LuxS homologues are widespread among Gram-negative and Gram-positive bacteria [14,15]. However, many bacteria only produce AI-2, but lack cognate receptors. The two classes of AI-2 receptors, LuxPQ and Lsr-receptor, are restricted to Vibrionales representatives and pathogenic bacteria associated with endotherms, respectively [16,17]. Therefore, the QS role of LuxS protein is arguable. It may be only a metabolic enzyme involved in the activated methyl cycle (AMC) [18] in these bacteria that are devoid of a complete AI-2 signaling pathway [16]. In addition to AI-2, indole has been suggested as an interspecies signal molecule, because it is shared by 85 species of Gram-positive and Gram-negative bacteria [19] (Table 1). However, some exceptions have been discovered. One type of AHL, 3-oxo-octanoyl homoserine lactone (3OC8-HSL), is utilized by a Gram-positive bacterium, Exiguobacterium sp. MPO, as a QS signaling molecule to regulate biofilm formation and extracellular polymeric substance production [20]. In addition, peptide-based QS is also found in a hyperthermophilic Gram-negative bacterium, Thermotoga maritima [21], and additionally in the yeast Cryptococcus neoformans [22]. Furthermore, AHLs are not the only signal type employed by Gram-negative bacteria. The diffusible signal factor (DSF) family, V. cholerae autoinducer-1 (CAI-1) family and other particular signals, such as Pseudomonas quinolone signal (PQS), integrating QS signal (IQS) and pyrone signal are also employed by some Gram-negative bacteria (Table 1). Among these molecules, the CAI-1 family is found mainly in Vibrio spp. [23] whereas the DSF family commonly exists in some plant pathogens, such as Xanthomonas and Burkholderia spp. [24]. Likewise, social behaviors namely filamentation and biofilm formation by the opportunistic fungal pathogen Candida albicans are regulated by farnesol- and tyrosol-based QS [25,26]. Moreover, it is striking that AHLs are present in more microorganisms than originally expected. Recently, a novel type of AHL, N-carboxyl-acyl-homoserine lactones, was found in a methanogenic archaeum Methanosaeta harundinacea to regulate its filamentous growth [27]. Each of these signal families has different structures and is used by different microbial groups (Figure 1, Table 1). These diverse signals may allow microbial populations to differentiate themselves from others, so as to synchronize and coordinate social behaviors.

{kind=link}
{kind=link}
{kind=link}
| QS Signaling Type | Structure | Representative Microorganisms | Associated Phenomena | Reference |
|---|---|---|---|---|
| Intraspecies Communication Signals | ||||
| “Traditional” AHL | C4-C18, 3OC4-3OC18 and 3OHC4-3OHC18 | Various Gram-negative bacteria; only one Gram-positive bacteria: Exiguobacterium sp. MPO | Virulence, biofilm, swarming and bioluminescence | [2,20] |
| “Noncanonical” AHL | p-Coumaroyl-HSL | Rhodopseudomonas palustris CGA009 | Global gene expression | [28] |
| DSF family | Cis-unsaturated fatty acid | Xanthomonas spp., and Burkholderia cenocepacia | Virulence, biofilm and antibiotic tolerance | [24] |
| CAI-1 family | α-Hydroxyketones | Vibrio spp. and Legionella pneumophila | Virulence and biofilm | [23,31] |
| AIP family | Linear or cyclized oligopeptide | Many Gram-positive bacteria; only one Gram-negative bacterium: Thermotoga maritima | Virulence, biofilm, sporulation and exopolysaccharide production | [21,32] |
| PQS or IQS | Quinolone or thiazole compounds | Pseudomonas aeruginosa | Virulence and biofilm formation | [33,34] |
| Pyrones | α-Pyrones | Photorhabdus luminescens | Virulence | [35] |
| Interspecies and Interkingdom Communication Signals | ||||
| AI-2 | S-THMF-borate R-THMF | Many Gram-negative and Gram-positives bacteria | Virulence and biofilm formation | [15] |
| AI-3 | Unknown | Enterohemorrhagic Escherichia coli (EHEC) | Virulence | [36] |
| Indole | 2,3-Benzopyrrole | Many Gram-negative and Gram-positives bacteria | Virulence and biofilm formation | [19] |
| QS Signals in Fungi | ||||
| Farnesol or Tyrosol | Sesquiterpene or phenylethanoid | Candida albicans | Inhibition or stimulation filamentation and biofilm formation | [25,26] |
| Peptide | NH2-NFGAPGGAYPW-COOH | Cryptococcus neoformans | Colony formation in agar media | [22] |
AHL: N-Acyl-homoserine lactonse; AI-2: Autoinducter-2; AI-3: Autoinducter-3; AIP: Autoinducing peptides; DSF: Diffusible signal factor; CAI-1: Cholerae autoinducer-1; PQS: Pseudomonas quinolone signal; IQS: Integrating QS signal.
Figure 1.
Structures of representative quorum sensing (QS) signals.

AHL: N-Acyl-homoserine lactonse; AI-2: Autoinducter-2; AIP: Autoinducing peptides; CAI-1: Cholerae autoinducer-1; Ea-C8-CAI-1: (Z)-3-Aminoundec-2-en-4-one; DSF: Diffusible signal factor; BDSF: Burkholderia cenocepacia diffusible signal factor; PQS: Pseudomonas quinolone signal; IQS: Integrating QS signal; R-THMF: (2R,4S)-2-Methyl-2,3,3,4-tetrahydroxytetrahydrofuran ; S-THMF-borate: (2S,4S)-2-Methyl-2,3,3,4-tetrahydroxytetrahydrofuran-borate; DPD: 4,5-Dihydroxy-2,3-pentanedione.
In most cases, one species usually employs a single type of QS. However, some species may harbor multiple QS systems. For example, V. harveyi utilizes a four-channel integrated QS system (AHL-, CAI-1-, AI-2- and nitric oxide-dependent channels) [37,38], whereas V. fischeri possesses a multichannel system (C8-HSL- and AI-2-dependent channels) and an additionally LuxI/R circuit (3OC6-HSL) [37]. P. aeruginosa employs various QS systems, including two complete AHL-dependent QS circuits (RhlI/R and LasI/R circuits), a 3OC12-HSL-responsive orphan receptor QscR, a Pseudomonas quinolone signal (PQS)-dependent QS, and a recently identified IQS (integrating the QS network) signal, which together compose a hierarchical QS network to regulate virulence expression and respond to environmental stress [33,39].
2.1. Characteristics of AHL Molecules
AHL-dependent QS exists in many pathogenic bacteria. A typical AHL molecule consists of a homoserine lactone and an acyl chain with an even number of carbons (C4-C18) together with an occasional modification at the C3 position (hydroxy or olefinic double bond) [2]. The shortest and longest AHLs found in nature are C4-HSL and C18-HSL, respectively. Recently, some special AHLs with novel structures have been discovered in Gram-negative bacteria and archaea, and include aryl-homoserine lactone (p-coumaroyl-HSL and cinnamoyl-HSL) [28,29], branched-chain fatty acyl-homoserine lactone (isovaleryl-HSL) [30] and N-carboxyl-acyl-homoserine lactone [27] (Table 1).
The solubility, diffusibility and stabilization of AHLs are correlated with their structures. Generally, the solubility and diffusibility will be increased, and the stabilization decreased along with the shorter length of acyl chain of AHLs. For example, 3OC6-HSL used by V. fischeri shows higher solubility and diffusibility which enables diffusion inside and outside of cells [2]. Conversely, 3OC12-HSL in P. aeruginosa can only be transported through efflux pumps [2]. In addition, the modification at the C3 position (also termed the β position) may increase the solubility of AHLs. AHLs are susceptible to pH and temperature of the surrounding environment. In fact, alkaline pH and high temperatures will promote their abiotic hydrolysis [40]. Therefore, it seems appropriate that the hyperthermophilic Gram-negative bacterium Thermotoga maritima utilizes peptides as the QS signal rather than AHLs in its hydrothermal habitat [21]. Another case is that of the photosynthetic bacterium, Rhodopseudomonas palustris, which uses an AHL synthase to produce p-coumaroyl-HSL from environmental p-coumaric acid, which is a plant metabolite, rather than fatty acids from cellular pools. This indicates that R. palustris QS might have evolved to fit its surrounding [28].
2.2. The Diversity and Uniformity of QS Systems
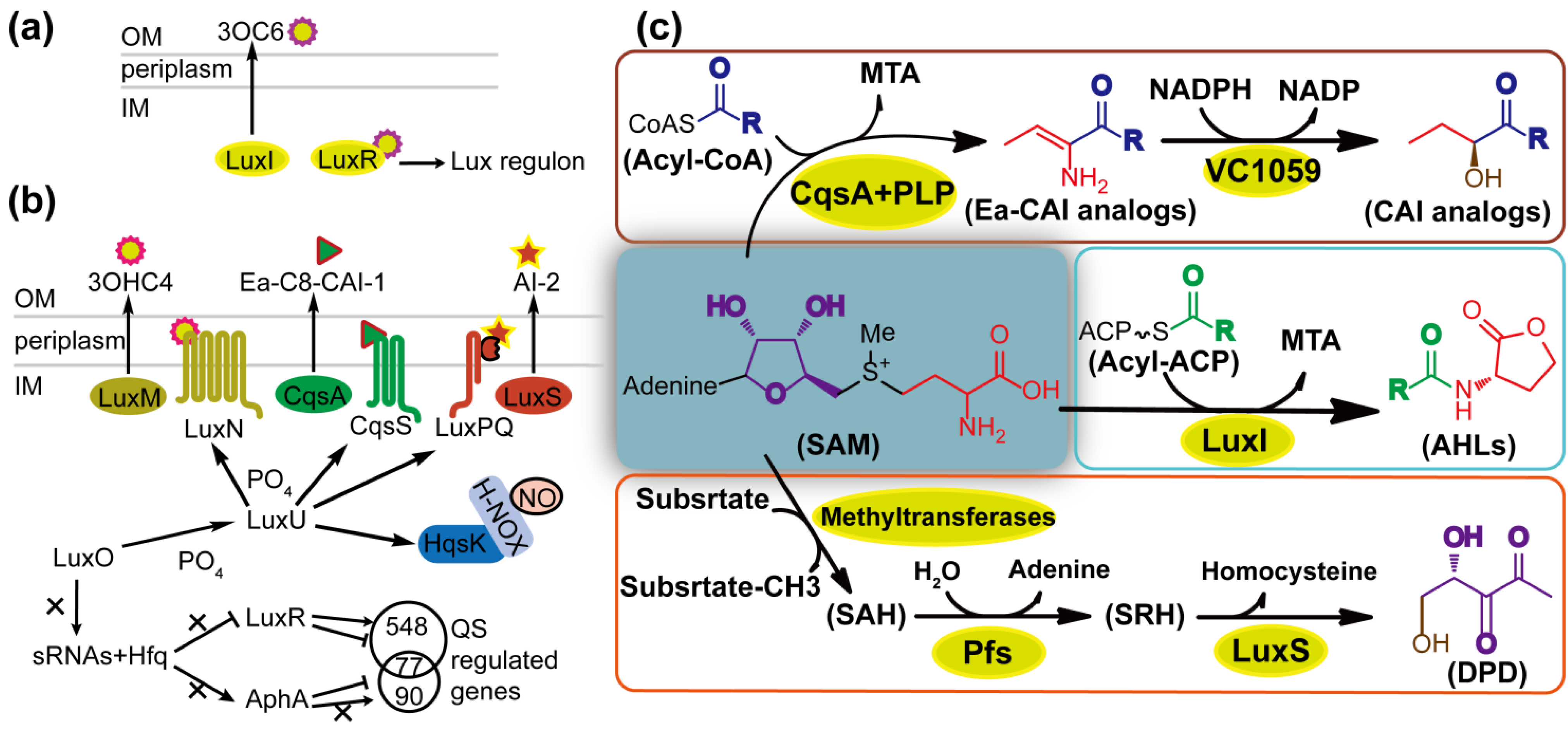
The LuxI/R circuit in marine bioluminescent V. fischeri (Figure 2a) and the integrated QS of V. harveyi are good examples to explain how different QS circuits work in a similar way (Figure 2b). V. harveyi possesses a sophisticated QS system consisting of four parallel channels, i.e., LuxM/N (AHL-dependent), LuxS/PQ (AI-2-dependent), CqsA/S (CAI-1-dependent) [2] and NO/H-NOX/HqsK (nitric oxide-dependent) [38]. In AHL-dependent QS, HAI-1 (3OHC4-HSL) is synthesized by the synthase LuxM. With AI-2-dependent QS, DPD (4,5-dihydroxy-2,3-pentanedione) is synthesized by LuxS, and is spontaneously cyclized and hydrated to form R-THMF and S-THMF (Figure 1). R-THMF may be detected by Salmonella enterica serovar Typhimurium directly [41], whereas, in the presence of boron, S-THMF is further catalyzed to form S-THMF-borate that can be utilized by Vibrio species as the AI-2 signal. In CAI-1-dependent QS, Ea-C8-CAI-1 is synthesized by CqsA [42]. The cognate receptors LuxN, LuxPQ, and CqsS are bi-functional two-component enzymes that possess both kinase and phosphatase activities [2]. LuxP is a periplasmic protein in complex with LuxQ; the former detects signals, and the latter transfers phosphate [2]. In the NO-responsive QS, NO is sensed by H-NOX and the NO/H-NOX complex regulates kinase and phosphatase activities of HqsK [38]. LuxR and AphA are the primary and secondary master regulators of QS in V. harveyi, respectively. At low cell density (LCD) or low NO concentration, phosphates from unliganded receptors are transduced to a single phosphotransfer protein LuxU, and subsequently to LuxO. Phospho-LuxO, together with a sigma factor σ54, activates the transcriptions of five small regulatory RNAs. Small RNAs repress the translation of LuxR and activate the translation of AphA [43,44]. At high cell density (HCD) or high NO concentration, phosphates are conversely transferred to the AI/NO-bound receptors and thereby the production of AphA is depressed whereas LuxR protein is maximally produced to regulate the expression of target genes [2]. Actually, LuxR is also present at LCD, whereas little or no AphA is present at HCD [45]. The two master regulators AphA and LuxR regulate 167 genes and 625 genes, respectively, and coregulate 77 genes, thereby generating a precise temporal pattern of gene expression [45].
Intriguingly, the biosynthesis pathways of AHLs, AI-2 and CAI-1 are related to each other and involved in the AMC, which is an important metabolic pathway responsible for the generation of the cell major methyl donor S-adenosylmethionine (SAM) and the recycling of methionine by detoxification of S-adenosyl-L-homocysteine (SAH) (Figure 2c). AHL is synthesized from SAM and acyl carrier protein (acyl-ACP) by LuxI-type synthase [2]. The structurally conserved homoserine lactone of AHL molecule is derived from SAM whereas the variable acyl tail is assembled from fatty acid. CAI-1 is synthesized from SAM and acyl-coenzyme A via a multistep reaction involving CsqA, pyridoxal phosphatase (PLP) and VC1059 [23]. CAI-1 and Ea-CAI-1 contain single three-carbon units, being derived from SAM and subsequently attached to an acyl tail. In AI-2 biosynthesis, the release of the activated methyl group from SAM to an acceptor molecule gives rise to a toxic intermediate, SAH, which is converted by 5′-methylthioadenosine/sadenosylhomocysteine nucleosidase (MTAN/Pfs) to S-ribosylhomocysteine (SRH) [46]. LuxS catalyzes the cleavage of SRH to homocysteine and DPD, which further undergoes spontaneously cyclization and hydration reactions to be mature AI-2 [46]. Unlike AHLs and CAI-1, all the carbons of DPD are derived from SAM.
All of the discovered QS circuits share three critical steps:
(1) production of signal molecules,
(2) release of signal molecules, and
(3) recognition of signal molecules by receptors.
In most cases, the LuxR-type transcriptional regulators are able to activate target genes, whereas the EsaR in Pantoea stewartii functions as a transcriptional repressor. Unliganded EsaR binds DNA and represses transcription. The binding of AI to EsaR promotes DNA release and thereby promotes gene expression [47]. Also, QS in Gram-positive bacteria contains three steps. Unlike AHL-dependent QS, AIPs are secreted to the environment by specialized transporters and subjected to post-translation modification to become mature linear or cyclized AIPs during transport [32].
Figure 2.
Representative QS circuits and autoinducer biosynthesis. (a) LuxI/R circuit of Vibrio fischeri. OM: outer membrane. IM: inner membrane; (b) QS circuit of V. harveyi; (c) Biosynthesis of N-acylhomoserine lactones (AHLs), V. cholerae autoinducer-1 (CAI-1) analogs and 4,5-dihydroxy-2,3-pentanedione (DPD). Differently colored carbons, nitrogens and oxygens show where they are derived. See details in the text.
Figure 2.
Representative QS circuits and autoinducer biosynthesis. (a) LuxI/R circuit of Vibrio fischeri. OM: outer membrane. IM: inner membrane; (b) QS circuit of V. harveyi; (c) Biosynthesis of N-acylhomoserine lactones (AHLs), V. cholerae autoinducer-1 (CAI-1) analogs and 4,5-dihydroxy-2,3-pentanedione (DPD). Differently colored carbons, nitrogens and oxygens show where they are derived. See details in the text.

3. Natural Quorum Quenching Resources
Quorum quenching is now referred to as the interference with QS, and may reverse the QS regulated phenotypes, such as virulence. Because QS is not essential for the growth of bacteria, quenching QS has been recommended as a promising strategy for antivirulence therapy. In particular, diverse QQ agents have been identified from various sources and organisms. All of the QQ agents may be classified into two groups according to their molecular weight, i.e., small molecular and macromolecular QQ agents, which are also referred to as QS inhibitors and QQ enzymes, respectively.
The words “quorum quenching” and “QS inhibitors” were first used to describe the enzymatic degradation of AHL signals [48] and inhibition of QS by small molecular antagonists [49], respectively. Here, in order to avoid a semantic quagmire, the word “quorum quenching” is used in its most extensive connotation to describe any forms of interference with QS, including QS inhibitors and QQ enzymes.
3.1. Small Molecular QQ Agents
There is abundant literature about the identification of novel and effective QS inhibitors from natural products and synthetic molecules. Multiple methods have been applied to identify QS inhibitors: purification from crude extracts of candidate organisms, random and high-throughput screening of QS inhibitors from compound libraries, and computer-aided screening from 3D structure libraries of known compounds. However, QS biosensors, which are genetically manufactured strains that express reporter genes (e.g., lacZ, gfp or luxCDABEG) in response to specific QS signals, are potent and indispensable tools for identifying QS inhibitors.
Natural QS inhibitors with highly diverse structures possess inhibitory activities against AHL, PQS, AI-2 and AIP-dependent QS (Table 2). It is notable that there are some unusual forms of interference with QS. Because the virulence factors of many pathogens are regulated positively by QS, it is reasonable to control pathogenicity by inhibiting QS. On the contrary, the orphan receptor QscR of P. aeruginosa [50] and CAI-1-dependent QS in V. cholerae [51] are negatively correlated with virulence, which means that these two particular pathways should be activated rather than inhibited for reducing virulence. Various synthetic agonists of each circuit have been identified [52,53,54]. However, natural compounds with this activity have not yet been found except for some corresponding AIs analogs, e.g., 3OC10-HSL for QscR and Ea-C8-CAI-1 for CqsS of V. cholerae. Therefore, specific screening work could be carried out to explore more natural QS inhibitors.
3.1.1. Marine-Derived Inhibitors against AHL-Dependent QS
AHL-dependent QS inhibitors account for the largest proportion of natural QS inhibitors (Table 2). Marine-derived QS inhibitors have been found in marine bacteria, fungi, algae, bryozoan, corals and sponges.
An increasing number of studies indicate that marine cyanobacteria have become one of the richest resources for biologically active and structurally unique natural products. Many AHL-dependent QS inhibitors with divergent structures have been discovered in several species of marine cyanobacteria (Table 2). The tumonoic acids (E, F, G and H), which have been isolated from Blennothrix cantharidosmum, inhibit the bioluminescence of V. harveyi BB120 without affecting bacterial growth. Here, tumonoic acid F is the most active moiety (IC50 of 62 μM) [55]. Moreover, Lyngbya majuscula produces four different compounds, including malyngolide (MAL), 8-epi-malyngamide C, lyngbic acid and lyngbyoic acid, which inhibit LasR of P. aeruginosa responding to exogenous 3OC12-HSL [56]. Interestingly, honaucins A–C [57], isolated from Leptolyngbya crossbyana, possess dual-inhibitory activity against both QS and inflammation, which may represent a new strategy for developing multi-functional drugs. It is noteworthy that marine cyanobacteria produce many different kinds of AHL-dependent QS inhibitors, and most likely have evolved mechanisms to control the associated microbial communities by interfering with their cell-to-cell communication.
| Category | Species | Inhibitor | Target | Reference |
|---|---|---|---|---|
| Marine-Derived Inhibitors against AHL-Dependent QS | ||||
| Actinobacteria | Streptomyces sp. | Piericidin A1 | CviR | [58] |
| Bacteria | Halobacillus salinus | N-(2-Phenylethyl)-isobutyramide and 3-Methyl-N-(2-phenylethyl)-butyramide | LuxR, CviR and Vibrio harveyi | [59,60] |
| Bacteria | Bacillus cereus D28 | Cyclo-l-proline-l-tyrosine | Chromobacterium violaceum | [60] |
| Bacteria | Marinobacter sp. SK-3 | Diketopiperazines (dkps) | CviR and LuxR | [61] |
| Bryozoan | Flustra foliacea | Brominated alkaloids compounds | CepR and LuxR | [62] |
| Coral | Pseudoplexaura flagellosa and Eunicea knighti | Cembranoids | LuxR and V. harveyi | [63,64] |
| Cyanobacteria | Blennothrix cantharidosmum | Tumonoic acid F | V. harveyi | [55] |
| Cyanobacteria | Lyngbya majuscula | 8-Epi-malyngamide C and lyngbic acid | LasR | [65] |
| Cyanobacteria | L. majuscula | Lyngbyoic acid | LasR | [66] |
| Cyanobacteria | L. majuscula | Malyngolide | CviR and LasR | [56] |
| Cyanobacteria | Leptolyngbya crossbyana | Honaucins A–C | LuxR | [57] |
| Cyanobacteria | Lyngbya sp. | Pitinoic acid A | LasR | [67] |
| Cyanobacteria | Lyngbya sp. | Pepitdes (microcolins A and B) | LuxR | [68] |
| Fungi | Penicillium atramentosum | Crude extracts | LasR | [69] |
| Red algae | Ahnfeltiopsis flabelliformes | Floridoside, betonicine and isethionic acid | TraR | [70] |
| Sponge | Luffareilla variabilis | Manoalide, manoalide monoacetate, and secomanoalide | LuxR and LasR | [71] |
| Sponge | Hymeniacidon aldis | Alkaloid (hymenialdisin) | LuxR and LasR | [68] |
| Terrestrial-Derived Inhibitors against AHL-Dependent QS | ||||
| Bacteria | Staphylococcus delphini | Yayurea A and B | LuxN | [72] |
| Bacteria | Stenotrophomonas maltophilia BJ01 | Cis-9-octadecenoic acid | CviR | [73] |
| Bacteria | Pseudomonas spp. | Protoanemonin | LasR | [74] |
| Fungi | Aspergillus spp. | Kojic acid | LuxR | [68] |
| Fungi | Penicillium spp. | Patulin and Penilillic acid | LasR and RhlR | [75] |
| Insect productions | Bee | Honey and propolis | C. violaceum, LasR and RhlR | [76,77] |
| Insect: fire ant | Solenopis invicta | Solenopsin A | rhl circuit | [78] |
| Plant | Baccharis cassinaefolia | Benzopyran | CviR, LuxR and LasR | [68] |
| Plant | Syzygium aromaticum | Eugenol | CviR, LasR and PQS | [79] |
| Plant: alfalfa | Medicago sativa | l-Canavanine | CviR and ExpR | [80] |
| Plant: Combretaceae | Combretum albiflorum | Catachin and naringenin | CviR and RhlR | [81,82] |
| Plant: Compositae | Centratherum punctatum | Sesquiterpene lactones | Pseudomonas aeruginosa | [83] |
| Plant: garlic | Allium sativum | Ajoene | LuxR family | [84] |
| Plant: horseradish | Armoracia rusticana | Iberin | LasR and RhlR | [85] |
| Plant: Myristicaceae | Myristica cinnamomea | Malabaricone C | CviR | [86] |
| Plant: tree | Drimys winteri | A drimane sesquiterpene | C. violaceum | [87] |
| Plant: turmeric | Curcuma longa | Curcumin | CviR | [88] |
| Plants | Various plants | p-Coumaric acid | PpuR, CviR and TraR | [89] |
| Plants and bacteria | Conocarpus erectus | Ellagitannins and urolithins | P. aeruginosa and Yersinia enterocolitica | [90,91] |
| Roundworm | Caenorhabditis elegans | Exudates | LuxR | [92] |
| Soil-freshwater alga | Chlamydomonas reinhardtii | Lumichrome | Sinorhizobium meliloti | [93,94] |
| TCMs * | Rhubarb | Emodin | P. aeruginosa and S. maltophilia | [95] |
| TCMs | Scutellaria baicalensis | Flavonoid (baicalein) | TraR and RhlR | [96] |
| Inhibitors against PQS System | ||||
| Fungi | Candida albicans | Farnesol (sesquiterpene) | PqsA | [97] |
| Inhibitors against AI-2 and AI-3 System | ||||
| Marine alga | Delisea pulchra | Furanone and its derivatives | LuxR and LuxS | [98,99] |
| Plant | Many plants | Cinnamaldehyde and its derivatives | LuxR and AI-2 | [100,101,102] |
| Plant | Broccoli | Quercetin | AI-2 and AI-3 | [103] |
| Plant | Grapefruit | Limonoids (obacunone) | EHEC | [104] |
| Inhibitors against AIP System | ||||
| Bacteria | Lactobacillus reuteri | Cyclic dipeptides: cyclo(l-Phe-l-Pro) and cyclo(l-Tyr-l-Pro) | agr system | [105] |
| Marine bacteria | Photobacterium | Cyclodepsipeptides (solonomide a, b) | agr system | [106] |
| Plant: witch hazel | Hamamelis virginiana | 2,5-di-O-galloyl-d hamamelose | RNAIII | [107] |
* TCMs: traditional Chinese medicines.
Other marine microorganisms also produce QS inhibitors (Table 2). The Gram-positive bacterium, Halobacillus salinus, produces two compounds, namely N-(2-phenylethyl)-isobutyramide and 3-methyl-N-(2-phenylethyl)-butyramide, which are capable of inhibiting violacein biosynthesis of Chromobacterium violaceum CV026 and GFP production of Escherichia coli JB525 in the presence of exogenous AHLs [59]. Both Gram-positive Bacillus cereus and Gram-negative Marinobacter sp. SK-3 produce diketopiperazines (DKPs) which inhibit AHL-dependent QS [60,61]. Additionally, piericidin derived from marine actinobacteria inhibits violacein biosynthesis in C. violaceum CV026 [58,108].
Another source for QS inhibitors is marine algae. The red alga Delisea pulchra produces a number of halogenated furanones with antifouling and antimicrobial properties [98,109]. Furanones are supposed to bind AHL receptors due to the five-membered lactone scaffold structurally similar with AHLs. A series of synthetic derivatives of native furanones have been reported to interfere with AHL-mediated QS in many bacteria [99,110]. It was suggested that furanones may act to destabilize the AHL-dependent transcriptional activator LuxR of V. fischeri [111] and attenuate the DNA-binding activity of LuxR of V. harveyi [112]. Moreover, it has been found that furanones may affect the AI-2 circuit in Gram-negative and Gram-positive bacteria by covalently modifying and inactivating AI-2 synthase LuxS [113]. The protective effects of halogenated furanones in rotifers, brine shrimp and rainbow trout against pathogenic Vibrio species [114,115,116] and in mice against P. aeruginosa lung infection [117] have been demonstrated, though some furanones were found to be toxic to rainbow trout, rotifers and human fibroblasts [115,116]. Therefore, studies have focused on the synthesis of more effective and less toxic furanone derivatives that may promote their commercial or therapeutic use. Except for furanones, a mixture of floridoside, betonicine and isethionic acid, produced by the marine red alga, Ahnfeltiopsis flabelliformes, may also inhibit the reporter strain A. tumefaciens NTL4 (pCF218) (pCF372) responding to 3OC6-HSL [70]. In addition, extracts of several marine micro-algae are capable of inhibiting the QS-dependent responses of reporter strains E. coli JB523 and V. harveyi JMH612 [118].
3.1.2. Terrestrial-Derived Inhibitors against AHL-Dependent QS
Various natural products from a wide range of terrestrial organisms have been demonstrated to show inhibitory activity against AHL-dependent QS. Most of these bioactive substances are derived from plants, although some originate from bacteria, fungi and insects (Table 2). Many health-benefit food sources and traditional medicines, such as fruits, herbs and medicinal plants, have received special attention as potential sources of QS inhibitory compounds. However, only a few of the compounds have been isolated or structurally and biochemically identified. A variety of phenolic compounds showed inhibitory activity against AHL-dependent QS. Among them, the flavonoids comprise a well-studied group. Flavonoids are widely produced by plants and also exist in plant-related products, e.g., propolis and honey [119]. Flavonoids display many pharmacological activities and present structural divergence [119]. The flavan-3-ol catechin, isolated from Combretum albiflorum bark, is the first identified flavonoid compound and is capable of reducing the production of virulence factors in P. aeruginosa PAO1 by interfering with RhlR [81]. Several other flavonoids from citrus and traditional Chinese medicines inhibit QS in bacteria [82,106,120]. Additionally, honey and propolis are able to interfere with QS in C. violaceum and P. aeruginosa, respectively, which may be attributed to the high abundance of flavonoids in honey and propolis [119]. Hydrolysable tannins are another group of phenolic compounds; ellagitannins or tannin-rich fraction from various plants showed QS inhibitory activity against C. violaceum or P. aeruginosa [90,91].
3.1.3. Natural Inhibitors against Other QS Systems
Compared with the plentiful natural AHL-dependent QS inhibitors, natural inhibitors against other QS systems are rarely reported (Table 2). Peptide-based compounds and the phenolic compound hamamelitannin are capable of blocking agr-dependent QS of Staphylococcus spp. [105,106,107]. It is striking that farnesol (a common sesquiterpene), applied by the opportunistic pathogen C. albicans as a QS signal, inhibits the PQS circuit of P. aeruginosa by promoting non-productive interaction between PqsR and the pqsA promoter [97]. It was suggested that farnesol may play a role in interkingdom communications [97]. Besides farnesol, two other sesquiterpene derivatives, drimendiol from Drimys winteri and sesquiterpene lactones from Centratherum punctatum (Argentine herb), were identified as AHL-dependent QS inhibitors [83,87]. Referring to the AI-2 system, furanone, cinnamaldehyde and their derivatives may be the most effective inhibitors. Cinnamaldehyde (CA) is widely used as a flavoring substance in food chemistry [100]. Low concentrations of CA were previously found to be effective at inhibiting both AHL and AI-2 dependent QS in V. harveyi [101]. Subsequently, mobility shift assays revealed that CA could decrease the DNA-binding ability of LuxR of V. harveyi. CA and its analogs could increase significantly the survival of the nematode Caenorhabditis elegans, brine shrimp and giant freshwater prawn, Macrobrachium rosenbergii, infected with pathogenic Vibrio spp. [100,102,121].
3.1.4. Evaluation of Natural QS Inhibitors
Over the last decade, many natural substances have been evaluated for their ability to interfere with QS. QS inhibitors isolated from natural products are excellent resources for developing potent antivirulence drugs insofar as they may provide novel scaffolds for drug design. The natural D. pulchra furanone compounds are unable to inhibit the QS of P. aeruginosa, but modified furanone analogs enhance the inhibitory effectiveness against this organism [117]. Honaucin A from the cyanobacterium Leptolyngbya was recently identified as an inhibitor against QS of V. harveyi and V. fischeri [57], whereas two synthesized honaucin A derivates, 4'-bromohonaucin A and 4-iodohonaucin A, showed an increased QS inhibitory activity of nearly 30-fold compared to that of honaucin A [57]. Despite these remarkable discoveries, three factors hinder the development of novel antivirulence therapies based on these bioactive substances.
Firstly, there is an increasing requirement for standardization to verify the true QS inhibitory activity. In some cases, the QS inhibitory activities of bioactive substances have been challenged because of the lack of suitable methodologies. Several QS reporter strains are widely used to identify QS inhibitors. Recently, Defoirdt et al. [122] detailed the inherent drawbacks of these assays. In reporter strains, the QS-regulated phenotypes are often co-dependent on other factors and/or dependent on the metabolic activity of the cells, and may thus be directly interfered with by candidate compounds. Hence, the same biosensor strain with a QS-independent expression of reporter genes should be used for adequate control experiments to verify that there is no effect imposed by candidate compounds on the particular phenotype. Additionally, QS-regulated phenotypes may be affected if the candidate compounds are toxic against reporter strains. In most cases, toxicity is only assessed by evaluating the effect on growth in a complex growth medium, which may cause false positives. For example, pyrogallol was found to be capable of inhibiting bioluminescence in a V. harveyi luxN mutant without affecting growth in a complex medium and therefore claimed as an AI-2 QS inhibitor [123]. However, it was subsequently found that pyrogallol inhibited bioluminescence when expressed constituently in an engineered strain, and the pyrogallol-related effect was abolished by the addition of catalase [124]. The apparent QS inhibitory activity of pyrogallol was demonstrated as a side effect of peroxide production [124]. Hence, more sensitive toxic assays towards bacterial cells are required [122].
Secondly, further studies on natural crude extracts with identified QS inhibitory activity are required. In many cases, the precise compounds of the bioactive molecules have not been elucidated. As mentioned above, natural inhibitors are resources for drug design to develop more potent antivirulence drugs, and purification of individual compounds is necessary to improve the likelihood of understanding the mechanisms of inhibition [39].
Moreover, the active mechanisms of these compounds are poorly understood. Successful drug design relies much on the knowledge of molecular mechanism of the connection between native signals or inhibitors and synthase or receptor, such as binding sites, conformational change and affinity change. Currently, the X-ray crystal structures of some AHL-bound LuxR-type receptors and computational protein docking methods provide powerful tools to determine the molecular mechanisms of interaction. However, the molecular mechanisms of only a few effective natural QS inhibitors have been studied in depth. The lack of information of binding interactions has thwarted the rational design of a more potent QS inhibitor.
3.2. Macromolecular QQ Agents
Various macromolecular agents have been found to possess the capability to quench QS. Unlike small molecular QS inhibitors, macromolecular QQ agents interfere with QS mostly through degrading signals rather than competitively binding to signal receptors. Most of the identified macromolecular QQ agents target the AHL-dependent QS, although enzymatic degradations of DSF, PQS and AI-2 have also been reported. Bacillus, Staphylococcus and Pseudomonas possess DSF inactivation activity [125]. The 2,4-dioxygenase, Hod, involved in quinaldine utilization pathway in Arthrobacter nitroguajacolicus is able to cleave PQS [126]. In the Lsr-type AI-2 system, cytoplasmic enzyme LsrK is responsible for phosphorylation of AI-2 and phospho-AI-2 is unstable [127]. When LsrK is artificially provided in vitro, the extracellular phosphor-AI-2 molecules cannot be transported into cells and are degraded overnight. LsrK-mediated degradation of AI-2 attenuates the QS response in S. enterica serovar Typhimurium and V. harveyi [127].
To date, many macromolecular QQ agents against AHL-dependent QS have been reported. Generally QQ enzymes are a major portion of the macromolecular agents, although a few antibodies have been generated to interfere with AHL-dependent QS through sequestration or hydrolyzation of AHLs [39]. Enzymatic degradation of AHLs has been extensively studied, and found in many organisms including mammals, plants, fungi, achaea and bacteria [39,128], although the genes responsible for AHL-degrading activity in plants and fungi have not been identified. A comprehensive summary of AHL-degrading bacteria with marine or terrestrial origin was provided in our previous publication [129].
AHL-degrading enzymes may be classified into three major types according to their enzymatic mechanisms: AHL lactonase (lactone hydrolysis), AHL acylase (amidohydrolysis) and AHL oxidase and reductase (oxidoreduction). AHL lactonase hydrolyzes the ester bond of AHL yielding the corresponding N-acyl-homoserine. This hydrolyzation may also occur spontaneously at alkaline pH, and may be reversed under acid pH [40]. AHL acylase hydrolyzes the amide bond of AHL to yield a homoserine lactone and the corresponding fatty acid chain, whereas AHL oxidase and reductase usually catalyzes a modification of AHLs. In most cases, AHL lactonases require metal ions (except AiiM and QsdH) and target both short and long acyl chain AHLs. Unlike lactonases, acylases exhibit substrate specificity based on the length of the acyl chain and the substitution on the β position of the AHL chain.
3.2.1. AHL Lactonases
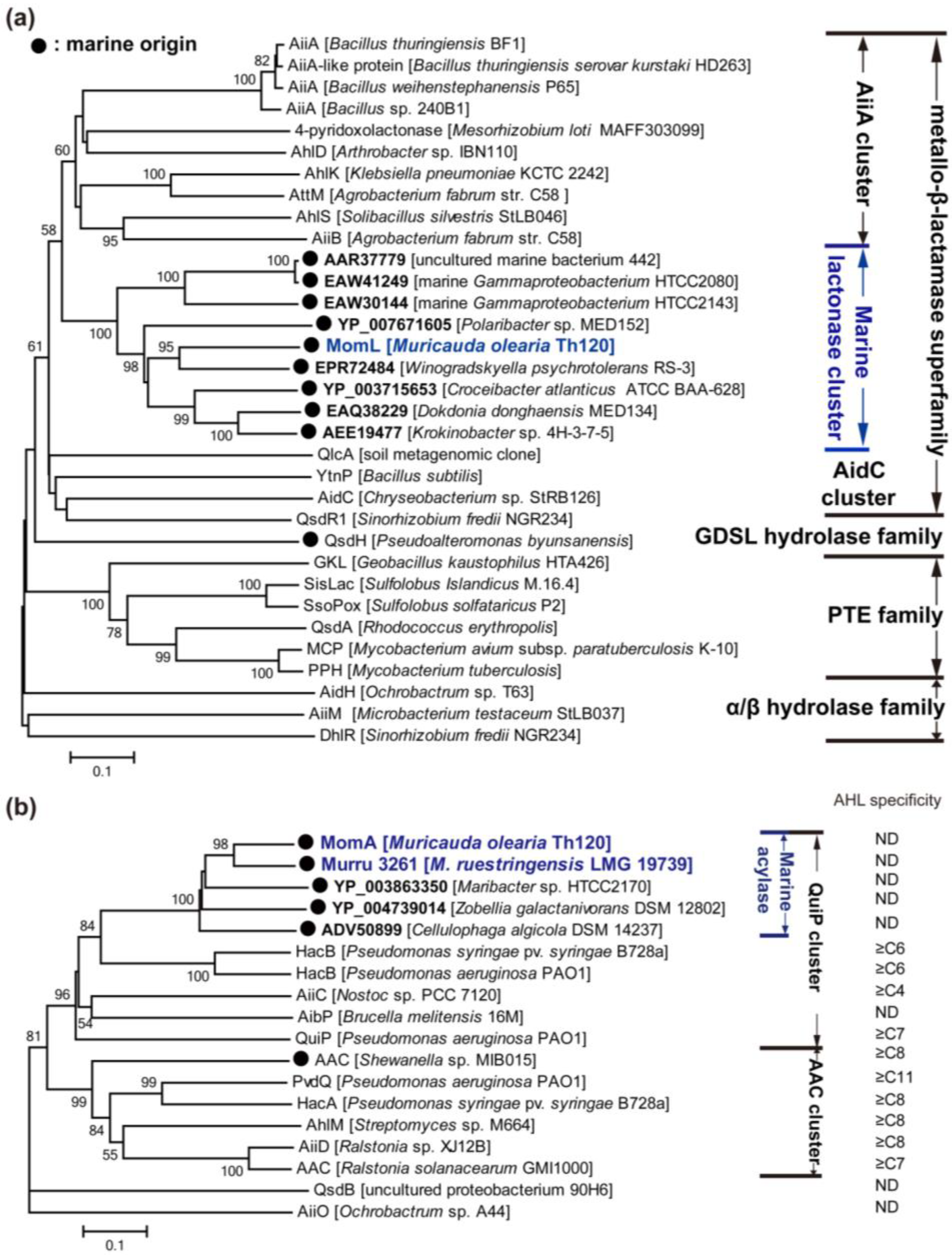
To date, approximately 30 types of AHL lactonases have been identified (except for predicted AHL lactonases or highly similar enzymes in one genus). According to the amino acid sequences, these lactonases belong to the metallo-β-lactamase superfamily, the phosphotriesterase (PTE) family and other particular families (Figure 3a). Among them, the AHL lactonases of the metallo-β-lactamase superfamily have been most extensively studied, and are widespread in various bacterial species. The metallo-β-lactamase group may be further classified into several clusters, i.e., AiiA, AidC and a novel marine AHL lactonase cluster [130] (discussed in depth later on) according to the phylogenetic tree (Figure 3a).
Figure 3.
Neighbour-joining tree of N-acylhomoserine lactone (AHL) enzymes based on amino acid sequences. Each of these AHL lactonases was experimentally identified, except the members named with accession number in Genbank (bold). MomL, MomA and Murru 3261 were identified by us recently (blue colored). The dendrogram was constructed by neighbor-joining method with the MUSCLE program in the MEGA software package (1000 bootstrap replicates). Bootstrap coefficients below 50% were not shown. Scale bar, 0.1 substitutions per amino acid position. Marine clusters were colored in blue. (a) Tree of AHL lactonase; (b) tree of acylase. ND: not determined.
Figure 3.
Neighbour-joining tree of N-acylhomoserine lactone (AHL) enzymes based on amino acid sequences. Each of these AHL lactonases was experimentally identified, except the members named with accession number in Genbank (bold). MomL, MomA and Murru 3261 were identified by us recently (blue colored). The dendrogram was constructed by neighbor-joining method with the MUSCLE program in the MEGA software package (1000 bootstrap replicates). Bootstrap coefficients below 50% were not shown. Scale bar, 0.1 substitutions per amino acid position. Marine clusters were colored in blue. (a) Tree of AHL lactonase; (b) tree of acylase. ND: not determined.
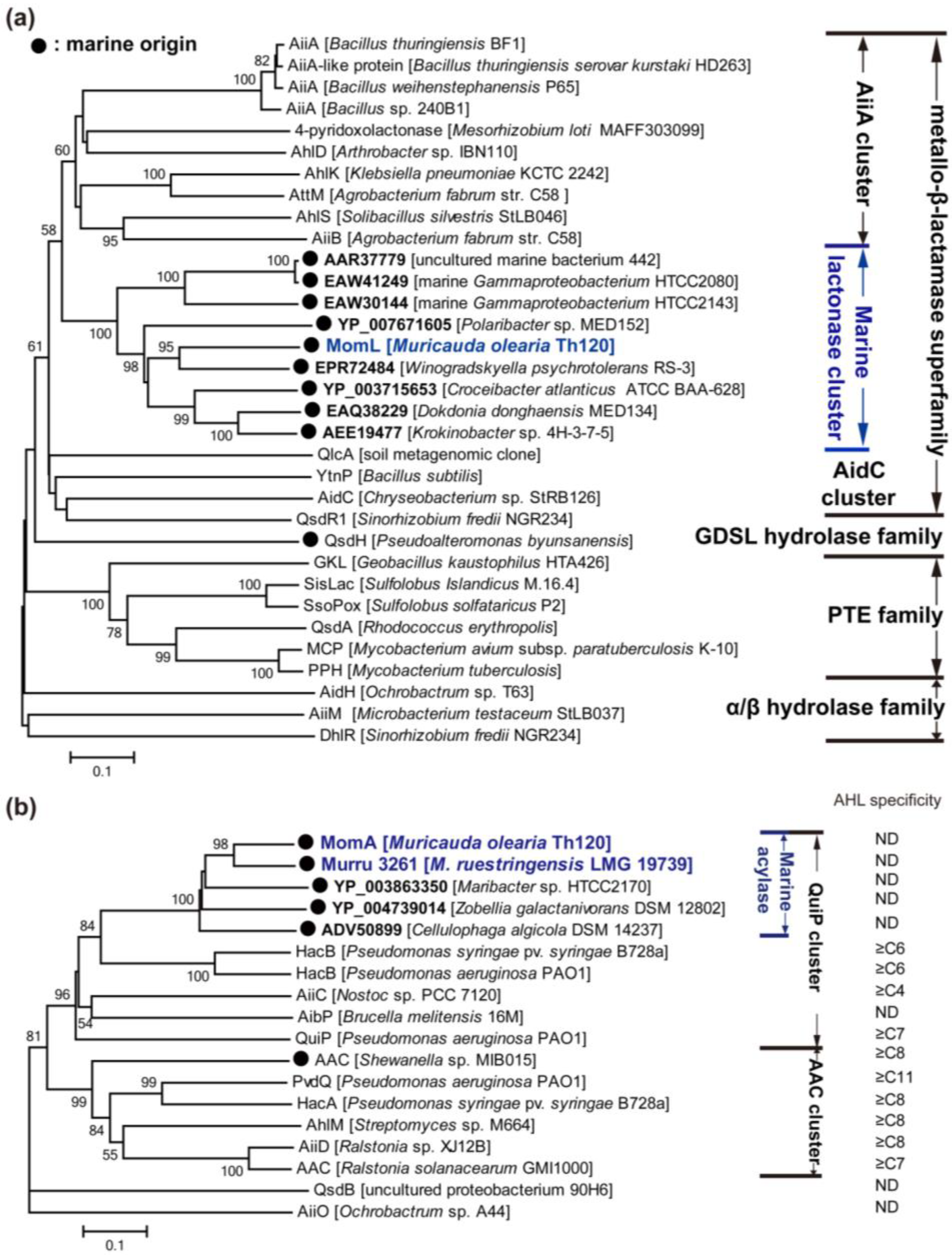
The first identified AHL lactonase AiiA (autoinducer inactivation) from Bacillus sp. 240B1 belongs to the metallo-β-lactamase family. AiiA was initially thought to hydrolyze the amide linkage between homoserine lactone and acyl side chain because metallo-β-lactamase can cleave the amide bond of the penicillin β-lactam ring. However, it was demonstrated later that AiiA degraded the ester rather than amide bond [131]. The aiia homogenous genes are widespread in Bacillus species. The molecular mechanism of AHL degradation has been revealed. AiiA amino acid sequence contains a “104HXHXDH109~60aa~H169” motif, which is common in metallo-β-lactamases. Single-residue mutagenesis has shown that His 106, Asp 108, His 109 and His 169 are necessary for AiiA activity [131]. AiiA is a metalloprotein binding two equivalents of zinc, which is necessary for its activity [132]. Crystal studies of AiiA from B. thuringiensis showed the dinuclear zinc binding sites of AiiA: Zn 1 binds to His 104, His 106 and His 169, whereas Zn 2 binds to Asp 108, His 109 and His 235 [133]. These two Zn2+ ions, which are separated by a distance of 3.4 Å, are bridged by a hydroxide ion and the Oδ2 atom of Asp191 [133]. Additionally, Tyr 194 is essential for activity, and may be able to stabilize the substrate’s carbonyl and/or a tetrahedral intermediate formed subsequent to hydroxide attack [133,134]. AiiA shows a broad substrate specificity and a preference for substrates with long acyl chain AHLs [135], however, C6-HSL is mostly used as substrate in crystal studies of AiiA [133,134]. Recently, another crystal study of a F107W mutation of B. thuringiensis AiiA revealed an unexpected interaction with the ring-opened product. Two aromatic residues, Phe 64 and Phe 68, form a hydrophobic clamp, centered around the seventh carbon of the decanoyl chain of ring-opened C10-HSL, making an interaction that would be available for longer substrates but not for shorter ones [136]. Although zinc was found in native AiiA, dicobalt-, dimanganese- or dicadmium-substituted AiiA exhibits hyperactivity compared with that of dizinc-substituted enzyme [134,137]. Heterologous expression of aiia in numerous pathogenic bacteria, including P. aeruginosa, B. thailandensis and E. carotovora, may reduce AHL accumulation and decrease their virulence expression, which indicates the potential use of AiiA as a strategy for antivirulence therapy [131].
Two members of AiiA cluster, AttM and AiiB, showing 32% and 28% identity to AiiA240B1, were found in the plant pathogen A. tumefaciens C58, and their corresponding coding genes attm and aiib (an attm-paralogous gene) located on the pAt plasmid and the pTi plasmid [138], respectively. Both AttM and AiiB have the “HXHXDH” motif and AiiB appears to be a metal-dependent AHL lactonase with broad substrate specificity [139]. A. tumefaciens infects a broad range of dicotyledonous plants and may transfer an oncogenic DNA fragment, the T-DNA, from its tumor-inducing plasmid to the nuclear genome of the plant hosts. These processes are regulated by TraI/R QS circuit, lactonases AttM and AiiB [140,141]. During the early process of A. tumefaciens infection in wounded plant hosts, the expression of attm is promoted by γ-aminobutyric acid, a molecule synthesized for defense by plants, and thereby the 3OC8-HSL-dependent virulence expression is attenuated. However, in the growing tumor, the high synthesis rate of 3OC8-HSL and the moderated expression of attm caused by plant produced l-proline permit the expression of QS-regulated functions including the transfer of Ti plasmid by conjugation [141].
The AidC cluster contains AidC and QsdR1 encoded by Chryseobacterium sp. StRB126 and Sinorhizobium fredii NGR234, respectively, and additionally includes YtnP, another AHL lactonase from Bacillus species [142,143,144]. Interestingly, the expression of ytnP in B. subtilis may be induced by streptomycin, and YtnP may conversely inhibit the production of streptomycin and the development of aerial mycelium in Streptomyces griseus. B. subtilis may activate ytnP expression in response to the presence of certain antimicrobial compounds as a defensive strategy against threatening bacteria since it would allow B. subtilis to selectively inhibit the QS-regulated behaviors of harmful microbial communities [144]. Furthermore, most of the AHL lactonases (both in the AiiA cluster and the AidC cluster) belonging to metallo-β-lactamase family are soil-derived. In this connection, we recently identified many marine bacteria with AHL-degrading activity [129], and one of these organisms, Muricauda olearia Th120, possesses a gene encoding a novel AHL lactonase which represents a new cluster of AHL lactonase in the metallo-β-lactamase family [130] (Figure 3a).
Apart from members of the metallo-β-lactamase family, another group of AHL lactonases shares key sequence and active site features with phosphotriesterases (PTEs), and these lactonases are termed PTE-like lactonases (PLLs). PLLs exist in both bacteria and archaea. Afriat et al. [145] discovered three PLLs, M. tuberculosis PPH, R. erythropolis AhlA and Sulfolobus solfataricus SsoPox. Although no significant sequence identities are shared with AiiA, PLLs show a wide range of AHL-degrading capability and require metal ions for their activities [145]. Crystal studies have revealed that the hyperthermophilic SsoPox shows a high level of similarity with the structure of AiiA [146]. Significantly, the production of AHLs and virulence factors of P. aeruginosa PAO1 may be greatly reduced in the presence of SsoPox-immobilized membranes [147]. The thermostability of SsoPox [147], as well as GKL of Geobacillus kaustophilus [148] and SisLac of S. islandicus [149], is an advantage for their biomedical applications. However, most PLLs showed relatively lower AHL-degrading activities than AiiA [147,148,149]. Curiously, all the identified PLLs exhibit promiscuous phosphotriesterase activities [147,148,149]. Bacterial PTEs belong to the amidohydrolase superfamily, a highly diverse superfamily with many different hydrolytic activities. They harbor a degrading rate approaching the diffusion limit (kcat/KM ≥ 4 × 107) for their best substrate paraoxon, which is a widely used pesticide introduced in the 20th century. PTEs could have therefore evolved from a member of the PLL family, utilizing its latent promiscuous paraoxonase activity as an essential starting point [145]. Additionally, DhlR, AidH and AiiM can be temporarily sorted into one group because they belong to the alpha/beta hydrolase family (Figure 3a). Both AidH and AiiM are capable of degrading short- and long-chain AHLs in an unknown and metal-independent mechanism [150,151]. There is another non-bacteria-derived group of AHL lactonases, paraoxonases 1, 2, and 3, which are prevalent in mammalian cells [128]. They all catalyze lactone hydrolysis, but differ in their substrate specificity [128].
3.2.2. AHL Acylases
AHL acylases have been found in bacteria including Pseudomonas, Ochrobactrum, Arthrobacter, Streptomyces, Nostoc and Brucella (Figure 3b). The AiiD of Ralstonia sp. XJ12B isolated from a biofilm in an experimental water treatment system is the first identified AHL acylase [152], although an AHL acylase-like activity was previously detected in Variovorax paradoxus VAI-C which degrades and utilizes multiple AHLs as the sole source of carbon, nitrogen and energy [153]. AiiD belongs to the Ntn (N-terminal nucleophile) hydrolase superfamily, and shares 22%–24% identities with several cephalosporin and penicillin acylases [152]. Actually, AiiD degrades several AHLs, rather than penicillin G or ampicillin, indicating that AHLs are its unique substrates. In comparison, AhlM from Streptomyces sp. which shows 35% identity with AiiD in the deduced amino acid sequence, was capable of degrading penicillin G and long-chain AHLs [154]. The gene responsible for AHL-acylase activity in V. paradoxus was not identified until whole genomic sequencing of V. paradoxus revealed recently a putative AHL acylase gene (Vapar_3883 of V. paradoxus S110) [155]. However, biochemical studies are required to confirm its activity. Like AHL lactonase, AHL acylase is considered to have the potential to interfere with QS of bacterial pathogens. Expression of aiiD in P. aeruginosa PAO1 weakened its ability to swarm, to produce elastase and pyocyanin, and to paralyse nematodes [152]. The addition of AhlM to the growth medium for P. aeruginosa also reduced the accumulation of AHLs, and decreased the production of virulence factors including elastase, total protease and LasA [154].
P. aeruginosa PAO1 was previously found to utilize AHLs for growth, so far three proteins (PvdQ, QuiP, and HacB) belonging to the Ntn hydrolase have been characterized biochemically [156]. Among them, PvdQ (PA2385) is the most extensively studied AHL acylase. PvdQ is expressed as a proenzyme that is auto-proteolytically activated by post-translational cleavage resulting in the excision of a 23-residue prosegment and the formation of an 18 kDa α-chain and a 60 kDa β-chain [156]. The mature PvdQ can hydrolyze the amide bond of AHLs, and demonstrates substrate specificity for long-chain AHLs [156]. However, it is the gene quiP (pa1032) rather than pvdQ that is responsible for the ability of P. aeruginosa PAO1 to utilize AHLs as a sole carbon and energy source for growth [156,157]. Additionally, another AHL acylase HacB (PA0305) can degrade AHLs with acyl chains ranging in length from 6 to 14 carbons [158]. The physiological function of each of the three acylases in P. aeruginosa PAO1 is intriguing. PvdQ was found to be expressed only when iron is present at very low concentrations [159]. Mutation of pvdQ did not affect the growth of P. aeruginosa but abrogated pyoverdine production and greatly affected swarming motility and biofilm formation at low iron concentrations [159]. Moreover, the virulence of pvdQ mutant against C. elegans was reduced. All of these data indicate that PvdQ plays an essential role in siderophore biosynthesis, on which P. aeruginosa depends for growth in iron-limited environments [159]. Therefore, PvdQ is a target for antivirulence therapy and different synthetic inhibitors are able to block its activity [160]. Nevertheless, the physiological functions of QuiP and HacB are still unknown.
To date, most of the identified AHL acylases belong to the Ntn hydrolase superfamily, except QsdB and AiiO which belong to the amidase family and α/β hydrolase fold family, respectively [161,162]. AHL acylases belonging to the Ntn hydrolase superfamily may also be classified into two clusters according to the phylogenetic tree (Figure 3c). These are referred to as AAC and QuiP clusters, respectively. The substrate specificity of each acylase cluster was also summarized, and it was speculated that the QuiP cluster might degrade a broader range of AHLs than the AAC cluster because some members of the QuiP cluster degrades C6-HSL even C4-HSL whereas those of AAC cluster could only degrade AHLs longer than C8-HSL. Additionally, most of these AHL acylases are located in the periplasmic space, whereas AhlM and HacB are secretory.
3.2.3. AHL Oxidoreductases
Compared to the abundant data of AHL lactonases and acylases, there are fewer reports about inactivation of AHLs by the modification of chemical structure of AHLs; only a few AHL oxidoreductases have been identified thus far. Because signal receptors usually respond to specific AHLs, the modification might affect the signal recognition, and thereby interfere with QS-regulated functions. Bacteria-derived AHL reduction activity was first discovered in Rhodococcus erythropolis in which AHLs with 3-oxo substituents were rapidly degraded by reduction of the keto group at the β position, yielding the corresponding 3-hydroxy derivative AHLs [163]. However, the gene responsible for this activity has not yet been identified. CYP102A1 from Bacillus megaterium, a widely studied cytochrome P450, is the second identified AHL oxidoreductases that oxidizes AHL at the ω-1, ω-2, and ω-3 carbons of the acyl chain [164]. Furthermore, this oxidation activity is very efficient towards ring-opened AHLs and fatty acid chains which are the corresponding products of AHL lactonase and acylase, respectively [164]. The third oxidoreductase, the NADH-dependent enzyme BpiB09, was identified by metagenomic analysis. Expression of bpiB09 in P. aeruginosa reduced its swimming motilities, pyocyanin production, biofilm formation and thereby the pathogenicity to C. elegans [165]. Moreover, Burkholderia sp. GG4, isolated from ginger rhizosphere, was previously found to possess a unique AHL-modifying activity that reduces 3-oxo-AHLs to 3-hydroxy-AHLs [166] although the responsible gene has not been identified. The complete genome of this strain reported recently might reveal its responsible gene [167]. AHL can also be enzymatically inactivated by haloperoxidases from D. pulchra [168], Laminaria digitata [169] and Nitzschia cf pellucida [170] via a H2O2-dependent mechanism.
4. Microorganisms May Produce QQ Agents to Gain Benefits in a Competitive Environment
Microorganisms exist in a multi-species and competitive environment, and have developed many survival strategies to gain benefits and compete for space, nutrition and ecological niches. QS is possibly one critical strategy used for competition by microorganisms to synchronize and coordinate social behaviors. Many of these behaviors (e.g., the production of antimicrobial compounds) are primarily advantageous when expressed by a group of bacteria but seemingly futile if performed by a single bacterium [171], though QS-regulated processes can also be induced in single cells in a confined environment [10,11]. Whereas, for other microorganisms exposed to QS-regulated competitive determinants, the selective pressure may drive the evolution of defensive mechanisms of fighting with competing species. It may be assumed that one microbial species may evolve two possible strategies to fight with another species that produces an antimicrobial compound in a QS-regulated mechanism. One strategy is to develop antimicrobial compound-degrading enzymes or other antimicrobial compound-resistant mechanisms. Another conceivable strategy is to interrupt the QS of competing species [171]. The hypothesis of QS interruption is straightforward because QQ-agent-producing bacteria can inhibit the QS-regulated behaviors of competing species and therefore gain benefits or avoid being killed. In this situation, small molecular QS inhibitors should be secreted outside of cells since their targets (e.g., signal synthases or receptors) are located in the membranes or cytoplasm of competing cells, whereas QQ enzymes could be either secreted or cytoplasmic because the signal molecules are diffusible. However, the cellular localization of QQ compounds has been little investigated.
The discovery of co-existence of QS and QQ bacteria in various environments might provide supports for this hypothesis [166,172,173]. However, conclusive empirical evidence has not been demonstrated regarding the relationship between QQ agents and the benefits gained in the natural environment. Additionally, some laboratory co-cultures of QS and QQ bacteria may provide evidence. The pyocyanin, which is a QS-regulated product of P. aeruginosa, is toxic to S. delphini [174] that produces two AHL-dependent QS inhibitors (yayurea A and B) and protects itself from killing by P. aeruginosa via suppressing the production of pyocyanin [72]. Likewise, pyocyanin is toxic to C. albicans [175]. With the production of farnesol, C. albicans blocks the PQS circuit and thus the pyocyanin biosynthesis of P. aeruginosa [97]. Farnesol-like molecules are ubiquitous in the natural environment, and are able to interrupt the PQS circuit, which suggests that other organisms may have the potential to moderate P. aeruginosa virulence [97]. It seems likely that the benefits outweigh the costs for these microorganisms to produce compounds to prevent harmful QS-regulated activities of other bacteria and thereby gain space and other resources within microbial communities.
The interference of QS exists not only between different species but also between different strains of the same species.In Gram-negative bacteria, the native AHL utilized by C. violaceum ATCC 31532 is C6-HSL but the QS-regulated violacein production can be inhibited by long-chain AHLs produced by C. violaceum ATCC 12472 or other bacteria [176,177]. Gram-positive S. aureus utilizes four different groups of AIPs. Each can specifically activate its cognate AgrC receptor, but inhibit all others by competitive binding to the non-cognate receptors [178,179,180,181]. This form of QS inhibition was suggested as a strategy for microorganisms to occupy specific niches during infection.
The physiological function of QQ enzyme has been discussed repeatedly but is still unclear [182,183,184]. PvdQ of P. aeruginosa participates in siderophore biosynthesis [159]. The major role of AttM in A. tumefaciens is for the degradation of γ-butyrolactone rather than regulation of AHL accumulation, and the AHL-degrading activity might be only a side effect [185]. V. paradoxus and Arthrobacter sp. utilize AHLs as a source of nitrogen or/and carbon for growth depending on their AHL-degrading enzymes [153,186]. AiiA is essential for rhizosphere colonization of B. thuringensis [187]. However, these results are insufficient to explain the physiological functions of QQ enzymes in these QQ bacteria that do not harbor the AHL-dependent QS and cannot utilize AHL to grow. Recently, Schneider et al. [144] reported that the expression of YtnP in B. subtilis was induced by streptomycin, an antibiotic produced by the Streptomyces species. Conversely, YtnP inhibits the production of streptomycin in S. griseus probably by degrading its QS signaling molecule γ-butyrolactone [144]. The streptomycin-induced expression of YtnP may allow B. subtilis to response to certain antimicrobial compounds and selectively inhibit the QS of harmful microorganisms before being killed. It seems likely that QQ enzyme could allow its producers to obtain competitive advantages over competitors in natural ecosystems.
The hypothesis that microorganisms produce QQ agents in order to gain competitive advantage is not sufficient to explain all of these discoveries. Although several examples showed that the QQ agent producers could survive or even gain benefit through inhibiting QS-regulated harmful behaviors of their competitors, the possibility of “accidental” QQ activity of some QQ agents still exists.
5. QQ in the Marine Environment: A Tremendous Resource to Be Developed
QQ may be a strategy used by microorganisms to gain benefit in a competitive environment. Also, it is believed that in the highly diverse marine ecosystem, microorganisms with capabilities of producing small QS inhibitors and QQ enzymes remain to be discovered. Romero et al. [184] proposed that QQ is likely to be a common activity in marine bacteria because a high abundance of QQ bacteria was found among marine cultivable bacteria [188] and a high frequency of QQ genes was discovered in marine metagenomes. In our previous study, 25 marine QQ strains belonging to 14 bacterial species were obtained and it is noteworthy that the QQ activities in 12 species had not been reported previously [129]. Although only a few studies have been carried out to assay the AHL-degrading activity of marine bacteria, more than 30 species of QQ bacteria belonging to Alphaproteobacteria, Gammaproteobacteria Actinobacteria, Flavobacteriia and Firmicutes have been identified thus far (Table 3). Additionally, some QQ strains have revealed degradative activity only against long-chain AHLs. Because AHL lactonases normally present broad AHL inactivating activities while many acylases are specific to long-chain AHLs, we assumed that AHL acylases might be more common than lactonases in the ocean. This is consistent with the distribution of acylase and lactonase coding sequences in metagenome collections [184]. Therefore, many marine QQ bacteria may be still undiscovered, and the prevalence of QQ enzymes in marine bacteria may be higher than expected.
The high diversity and abundance of marine QQ bacteria may lead to the discovery of new QQ enzymes and AHL-degrading mechanisms. However, few responsible genes in these bacteria have been identified (Table 3). One of our identified QQ bacteria, M. olearia Th120, showed strong AHL-degrading activity, and further studies revealed a novel AHL lactonase and a novel AHL acylase [130] (Figure 3). The identities of the amino acid sequence of AHL lactonase MomL (Muricauda olearia marine AHL lactonase) to known lactonases are below 30%, and the top eight strains (with identity higher than 39%) using BLASTP against the NR protein database are also typical marine bacteria. Therefore, MomL was believed to represent a new class of AHL lactonase, which may be widespread in the marine environment. Likewise, AHL acylase MomA (Muricauda olearia marine AHL acylase) may represent a marine-derived AHL acylase. It is even more astounding that the ethyl acetate extracts of Th120 culture showed inhibitory activity in A. tumefaciens A136 plate assay [130]. All of these findings indicate that marine microorganisms may be important resources for the discovery of new antivirulence strategies. Therefore, an increasing effort is needed in the discovery of new natural QQ agents from marine microorganisms.
| Strain | AHL-Degrading Ability * | Activity ** | Origin | Reference |
|---|---|---|---|---|
| Actinobacteria | ||||
| Rhodococcus erythropolis strains | C4, C6, C10 and 3OC12 | Lactonase | Fucus vesiculosus and sediment | [188] |
| Alphaproteobacteria | ||||
| Hyphomonas sp. USC2 | C4, C6, C10 and 3OC12 | Lactonase | Fucus vesiculosus | [188] |
| Marivita sp. Th30 | C6, C12 and C14 | ND | Flounder | [129] |
| Novosphingobium sp. Th20 | C6-C14 and 3OC6-3OC14 | ND | Flounder | [129] |
| Paracoccus sp. PP2-663 | C4-C12 | ND | Manila clam | [189] |
| Phaeobacter sp. USC177 | C4, C6, C10 and 3OC12 | ND | Fucus vesiculosus | [188] |
| Rhodobacter sp. Th15 | C8-C14 and 3OC14 | ND | Flounder | [129] |
| Roseovarius aestuarii USC61 | C4, C6, C10 and 3OC12 | Lactonase | Water tank | [188] |
| Sphingopyxis flavimaris T51 | C6-C14 and 3OC10-3OC14 | ND | Flounder | [129] |
| Sphingopyxis litoris th8 | C6-C14 and 3OC6-3OC14 | ND | Flounder | [129] |
| Stappia sp. USC176 | C4, C6, C10 and 3OC12 | Lactonase | Fucus vesiculosus | [188] |
| Stappia sp. USC5 | C4, C6, C10 and 3OC12 | Lactonase | Fucus vesiculosus | [188] |
| Firmicutes | ||||
| Bacillus circulans USC24 | C4, C6, C10 and 3OC12 | Lactonase | Sediment | [188] |
| Bacillus sp. KT7 | C6-C14, 3OC8-3OC12 and 3OHC8-3OHC12 | ND | Intertidal rocks colonized by Ulva | [190] |
| Bacillus spp. | C6 | ND | Shrimp and bass | [191] |
| Oceanobacillus spp. | C4, C6, C10 and 3OC12 | Lactonase | Fucus vesiculosus | [188] |
| Flavobacteria | ||||
| Flaviramulus ichthyoenteri Th78 | C6-C14 and 3OC6-3OC14 | Lactonase | Flounder | [129] |
| Maribacter sp. 139 | C4, C6, C10 and 3OC12 | Lactonase | Ocean water | [184] |
| Muricauda olearia Th120 | C6-C14 and 3OC6-3OC14 | Latonase and acylase | Flounder | [129] |
| Olleya marilimosa 138E | C4, C6, C10 and 3OC12 | Lactonase | Ocean water | [184] |
| O. marilimosa t168 | C6-C14 and 3OC6-3OC14 | Lactonase | Marine | [129] |
| Tenacibaculum discolor 20J | C4, C6, C10 and 3OC12 | Lactonase | Sediment | [188] |
| T. discolor t84 | C6-C14 and 3OC6-3OC14 | ND | Gill of flounder | [129] |
| T. maritimum 2154t | C10 | Acylase | Fish farm disease | [192] |
| T. soleae strains | C6-C14 and 3OC6-3OC14 | Lactonase | Gill of flounder | [129] |
| Gammaproteobacteria | ||||
| Alteromonas marina PP2-67 | C4-C12 | ND | Pod razor clam | [189] |
| Alteromonas sp. USC168 | C4, C6, C10 and 3OC12 | ND | Fucus vesiculosus | [188] |
| A. stellipolaris pp2-644 | C4-C12 | ND | Carpet-shell clam | [189] |
| Colwellia aestuarii T171 | C8-C14 and 3OC10-3OC14 | ND | Gill of flounder | [129] |
| Glaciecola sp. B20 | C10-C14, 3OC10-, 3OHC10, 3OC12 and 3OHC12 | ND | Intertidal rocks | [190] |
| Halomonas taeanensis USC33 | C4, C6, C10 and 3OC12 | Lactonase | Sediment | [188] |
| Marinobacterium sp. B2 | 3OC10, C12, 3OC12, 3OHC12 and C14 | ND | Intertidal rocks | [190] |
| Pseudoalteromonas byunsanensis 1A01261 | C4-C14 and 3OC4-3OC12 | Lactonase | Marine | [193] |
| P. rydzensis Th125 | C10-C14 and 3OC10-3OC14 | ND | Flounder | [129] |
| Salinimonas sp. T194 | C8-C14 and 3OC10-3OC14 | ND | Gill of flounder | [129] |
| Salinicola salarius 131 | C4, C6, C10 and 3OC12 | Lactonase | Ocean water | [184] |
| Shewanella sp. B21 | C8-C14, 3OC8-3OC12 and 3OHC8-3OHC12 | ND | Intertidal rocks | [190] |
| Thalassomonas sp. PP2-459 | C4-C12 | ND | Carpet-shell clam | [189] |
| Thalassomonas sp. T202 | C8-C14 and 3OC10-3OC14 | ND | Gill of flounder | [129] |
* All of AHLs contain even number of carbons; ** AHL-degrading activities were identified in bacterial cultures but not purified enzymes, except for P. byunsanensis 1A01261 and M. olearia Th120; ND: not determined.
6. Further Issues of Concern for the Application of QQ Agents
The antivirulence activities of small molecular QS inhibitors and QQ enzymes have been demonstrated in vitro and in vivo. However, both have advantages and drawbacks due to their entirely distinct molecular structures and functional mechanisms.
QS inhibitors may target one specific signal receptor or some homogenous receptors, such as the LuxR-like family. For example, compounds 4606–4327, CTL, CL and mBTL are synthetic AHL analogs with similar structures [194]. Each is the antagonist of CviR of C. violaceum and LuxN of V. harveyi. However, only mBTL is capable of inhibiting the pyocyanin production of P. aeruginosa PA14 whereas CL and CTL show non-inhibitory activity [195]. This may contribute to developing drugs that are capable of preventing virulence expression in specific pathogens. However, microbial infection is often caused by multiple pathogenic species, and one drug may be insufficient in this situation. In contrast, QQ enzymes, especially AHL lactonase, are capable of degrading a wide range of AHLs, and are likely to be more efficient for antivirulence by treating multi-microbial infection. Nevertheless, this capability of AHL-degrading enzymes could cause unintended consequences if a beneficial activity of a probiotic in the intestine of animals or human is positively regulated by its AHL-dependent QS [196].
Compared with QQ enzymes, the structures of QS inhibitors are relatively simple and can be easily modified through synthetic methods. Moreover, small QS inhibitors allow for temporal control of a biological system, and this control is often rapid, depending on the diffusibility of compounds. For example, the low molecular weight of QS inhibitors facilitates their absorption by animals; they may be administered orally or intravenously like other drugs. Furthermore, the nonproteinaceous nature as well as low molecular weight of QS inhibitors can effectively prevent an antibody-based immune response unlike that of QQ enzymes.
Stability is another important issue that should be taken into account for both of these two agents. QQ enzymes could be easily proteolyzed and most of them are sensitive to heat. Likewise, QS inhibitors may be degraded by abiotic or biotic elements. This is of concern especially for some QS inhibitors possessing similar structures with native AHLs. This type of QS inhibitor is suspected to be degraded by AHL-degrading enzymes because AiiA can degrade AHL analogs [135,137], and SisLac harbors AHL lactonase activity, esterase activity and phosphotriesterase activity [149]. Many of the identified QS inhibitors have similar structures to AHLs, especially of the synthetic AHL analogs. If this type of QS inhibitor was used to inhibit QS-dependent virulence in a multi-microbial community, it may be degraded by QQ enzymes produced by other microorganisms to obtain a competitive benefit in the environment. However, the QQ enzyme-mediated degradation of QS inhibitors has been ignored to date. It is advised that whether QS inhibitors can be degraded by QQ enzymes should be determined in the criteria proposed by Defoirdt et al. [122] for the scientific evaluation of QS inhibitory activity.
The emergence of resistance to QQ compounds has raised doubts about whether QS is an ideal target for antivirulence therapy. Defoirdt et al. [197] proposed that bacteria might evolve resistance to QQ compounds because QS disruption could indeed affect bacterial growth under certain conditions (e.g., during infection of a host). Subsequently, growth inhibition was observed by cultivating P. aeruginosa PA14 on minimal medium using adenosine as the sole carbon source and with the simultaneous exposure to the synthetic QS inhibitor furanone C-30 [198]. Further studies revealed a QQ resistance mechanism through an increased efflux of C-30 from the cells by mutations in the mexR and nalC genes, both of which encode negative regulators of the MexAB-OprM multidrug resistance efflux pump [198]. Moreover, mexR and nalC mutations were found in several clinical P. aeruginosa isolates. However, unlike QS inhibitors, application of QQ lactonase would be less likely to induce QQ resistance [197,199]. The degradation activity of QQ enzyme targeting signals is extracellular rather than entering cells and targeting receptors, which would be hardly influenced by an increased efflux of compounds from cells. Despite the possible ways in which bacteria develop resistance to AHL-degrading enzymes, such as increasing production of autoinducers, synthesis of modified autoinducers and evolution of mutations with higher-affinity receptors [199], the high AHL-degrading activity and broad range of substrate specificity of QQ enzymes would reduce the possibility of evolving QQ resistance.
The potential for the development of antivirulence drugs has been emphasized for years, and a number of QQ agents have been discovered or synthesized, but none have been marketed. Certainly, the lack of appropriate delivery systems is one challenge. The corresponding QQ agents should be introduced into the hosts against different pathogens by controlling the rate, time, and place of release. The novel biological nanofactories engineered by Fernandes and colleagues would provide a promising specific delivery of QQ agents [200]. Another self-regulating system designed to release QQ agents dependent on the titer of bacteria surrounding medical devices has potential [201]. It is likely to be more difficult to develop formulations for delivering macromolecular QQ agents due to the extremely low bioavailability of protein drugs [202]. Although oral administration of AiiA homogenous protein and preparation dry powder of PvdQ have been attempted, there are concerns about the stability of QQ enzymes. Clatworthy et al. [1] argued that the greatest challenge for commercialization of antivirulence drugs is not technological but economic. Since antivirulence drugs are narrow-spectrum, their effectiveness is dependent on the precise diagnosis of the pathogens to achieve an appropriate choice of compounds. Therefore, appropriate tools need to be developed to allow decisions to be made [1]. Thus, the technological and economic obstacles for the commercialization of QQ drugs to be overcome in the future become more urgent than the discovery of novel QQ agents [1].
7. Concluding Remarks
In summary, the utilization of quorum quenching as a promising strategy of antivirulence therapy has been demonstrated in vitro and in vivo. The natural QQ agents, especially those derived from marine microorganisms, are great resources for developing antivirulence therapy. Recently, several studies have revealed a wide spread of QQ activities in marine microorganisms, however, these QQ resources need to be explored more deeply. Therefore, further research on QQ resources and mechanisms would provide more alternatives for developing antivirulence therapy.
Abbreviations
| AHL | N-Acyl-homoserine lactonse |
| AI-2 | Autoinducter-2 |
| AI-3 | Autoinducter-3 |
| AIP | Autoinducing peptides |
| DSF | Diffusible signal factor |
| BDSF | Burkholderia cenocepacia diffusible signal factor |
| CAI-1 | Cholerae autoinducer-1 |
| Ea-C8-CAI-1 | (Z)-3-Aminoundec-2-en-4-one |
| PQS | Pseudomonas quinolone signal |
| IQS | Integrating QS signal |
| R-THMF | (2R,4S)-2-Methyl-2,3,3,4-tetrahydroxytetrahydrofuran |
| S-THMF-borate | (2S,4S)-2-Methyl-2,3,3,4-tetrahydroxytetrahydrofuran-borate |
| DPD | 4,5-Dihydroxy-2,3-pentanedione |
Acknowledgments
We are very grateful to Brian Austin (University of Stirling, Scotland, UK) for careful English editing and critical reading of the manuscript. This work was supported by the International Science and Technology Cooperation Programme of China (no. 2012DFG31990).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Clatworthy, A.E.; Pierson, E.; Hung, D.T. Targeting virulence: A new paradigm for antimicrobial therapy. Nat. Chem. Biol. 2007, 3, 541–548. [Google Scholar] [CrossRef]
- Ng, W.L.; Bassler, B.L. Bacterial quorum-sensing network architectures. Annu. Rev. Genet. 2009, 43, 197–222. [Google Scholar] [CrossRef]
- Waters, C.M.; Bassler, B.L. Quorum sensing: Cell-to-cell communication in bacteria. Annu. Rev. Cell. Dev. Biol. 2005, 21, 319–346. [Google Scholar] [CrossRef]
- Fuqua, W.C.; Winans, S.C.; Greenberg, E.P. Quorum sensing in bacteria: The LuxR-LuxI family of cell density-responsive transcriptional regulators. J. Bacteriol. 1994, 176, 269–275. [Google Scholar]
- Redfield, R.J. Is quorum sensing a side effect of diffusion sensing? Trends Microbiol. 2002, 10, 365–370. [Google Scholar] [CrossRef]
- Hense, B.A.; Kuttler, C.; Muller, J.; Rothballer, M.; Hartmann, A.; Kreft, J.U. Does efficiency sensing unify diffusion and quorum sensing? Nat. Rev. Microbiol. 2007, 5, 230–239. [Google Scholar] [CrossRef]
- Boyer, M.; Wisniewski-Dye, F. Cell-cell signalling in bacteria: Not simply a matter of quorum. FEMS Microbiol. Ecol. 2009, 70, 1–19. [Google Scholar] [CrossRef]
- West, S.A.; Winzer, K.; Gardner, A.; Diggle, S.P. Quorum sensing and the confusion about diffusion. Trends Microbiol. 2012, 20, 586–594. [Google Scholar] [CrossRef]
- Decho, A.W.; Norman, R.S.; Visscher, P.T. Quorum sensing in natural environments: Emerging views from microbial mats. Trends Microbiol. 2010, 18, 73–80. [Google Scholar] [CrossRef]
- Boedicker, J.Q.; Vincent, M.E.; Ismagilov, R.F. Microfluidic confinement of single cells of bacteria in small volumes initiates high-density behavior of quorum sensing and growth and reveals its variability. Angew. Chem. Int. Ed. 2009, 48, 5908–5911. [Google Scholar] [CrossRef]
- Carnes, E.C.; Lopez, D.M.; Donegan, N.P.; Cheung, A.; Gresham, H.; Timmins, G.S.; Brinker, C.J. Confinement-induced quorum sensing of individual Staphylococcus aureus bacteria. Nat. Chem. Biol. 2010, 6, 41–45. [Google Scholar] [CrossRef]
- Schuster, M.; Sexton, D.J.; Diggle, S.P.; Greenberg, E.P. Acyl-homoserine lactone quorum sensing: From evolution to application. Annu. Rev. Microbiol. 2013, 67, 43–63. [Google Scholar] [CrossRef]
- Platt, T.G.; Fuqua, C. What’s in a name? The semantics of quorum sensing. Trends Microbiol. 2010, 18, 383–387. [Google Scholar] [CrossRef]
- Surette, M.G.; Miller, M.B.; Bassler, B.L. Quorum sensing in Escherichia coli, Salmonella typhimurium, and Vibrio harveyi: A new family of genes responsible for autoinducer production. Proc. Natl. Acad. Sci. USA 1999, 96, 1639–1644. [Google Scholar] [CrossRef]
- Pereira, C.S.; Thompson, J.A.; Xavier, K.B. AI-2-mediated signalling in bacteria. FEMS Microbiol. Rev. 2013, 37, 156–181. [Google Scholar]
- Rezzonico, F.; Duffy, B. Lack of genomic evidence of AI-2 receptors suggests a non-quorum sensing role for luxS in most bacteria. BMC Microbiol. 2008, 8, 154. [Google Scholar] [CrossRef]
- Pereira, C.S.; de Regt, A.K.; Brito, P.H.; Miller, S.T.; Xavier, K.B. Identification of functional LsrB-like autoinducer-2 receptors. J. Bacteriol. 2009, 191, 6975–6987. [Google Scholar] [CrossRef]
- Winzer, K.; Hardie, K.R.; Burgess, N.; Doherty, N.; Kirke, D.; Holden, M.T.; Linforth, R.; Cornell, K.A.; Taylor, A.J.; Hill, P.J.; et al. LuxS: Its role in central metabolism and the in vitro synthesis of 4-hydroxy-5-methyl-3(2H)-furanone. Microbiology 2002, 148, 909–922. [Google Scholar]
- Lee, J.H.; Lee, J. Indole as an intercellular signal in microbial communities. FEMS Microbiol. Rev. 2010, 34, 426–444. [Google Scholar]
- Biswa, P.; Doble, M. Production of acylated homoserine lactone by Gram-positive bacteria isolated from marine water. FEMS Microbiol. Lett. 2013, 343, 34–41. [Google Scholar] [CrossRef]
- Johnson, M.R.; Montero, C.I.; Conners, S.B.; Shockley, K.R.; Bridger, S.L.; Kelly, R.M. Population density-dependent regulation of exopolysaccharide formation in the hyperthermophilic bacterium Thermotoga maritima. Mol. Microbiol. 2005, 55, 664–674. [Google Scholar]
- Lee, H.; Chang, Y.C.; Nardone, G.; Kwon-Chung, K.J. TUP1 disruption in Cryptococcus neoformans uncovers a peptide-mediated density-dependent growth phenomenon that mimics quorum sensing. Mol. Microbiol. 2007, 64, 591–601. [Google Scholar] [CrossRef]
- Kelly, R.C.; Bolitho, M.E.; Higgins, D.A.; Lu, W.; Ng, W.-L.; Jeffrey, P.D.; Rabinowitz, J.D.; Semmelhack, M.F.; Hughson, F.M.; Bassler, B.L. The Vibrio cholerae quorum-sensing autoinducer CAI-1: Analysis of the biosynthetic enzyme CqsA. Nat. Chem. Biol. 2009, 5, 891–895. [Google Scholar] [CrossRef]
- Deng, Y.; Wu, J.; Tao, F.; Zhang, L.H. Listening to a new language: DSF-based quorum sensing in Gram-negative bacteria. Chem. Rev. 2011, 111, 160–173. [Google Scholar] [CrossRef]
- Chen, H.; Fujita, M.; Feng, Q.; Clardy, J.; Fink, G.R. Tyrosol is a quorum-sensing molecule in Candida albicans. Proc. Natl. Acad. Sci. USA 2004, 101, 5048–5052. [Google Scholar] [CrossRef]
- Hornby, J.M.; Jensen, E.C.; Lisec, A.D.; Tasto, J.J.; Jahnke, B.; Shoemaker, R.; Dussault, P.; Nickerson, K.W. Quorum sensing in the dimorphic fungus Candida albicans is mediated by farnesol. Appl. Environ. Microbiol. 2001, 67, 2982–2992. [Google Scholar] [CrossRef]
- Zhang, G.; Zhang, F.; Ding, G.; Li, J.; Guo, X.; Zhu, J.; Zhou, L.; Cai, S.; Liu, X.; Luo, Y.; et al. Acyl homoserine lactone-based quorum sensing in a methanogenic archaeon. ISME J. 2012, 6, 1336–1344. [Google Scholar] [CrossRef]
- Schaefer, A.L.; Greenberg, E.P.; Oliver, C.M.; Oda, Y.; Huang, J.J.; Bittan-Banin, G.; Peres, C.M.; Schmidt, S.; Juhaszova, K.; Sufrin, J.R.; et al. A new class of homoserine lactone quorum-sensing signals. Nature 2008, 454, 595–599. [Google Scholar] [CrossRef]
- Ahlgren, N.A.; Harwood, C.S.; Schaefer, A.L.; Giraud, E.; Greenberg, E.P. Aryl-homoserine lactone quorum sensing in stem-nodulating photosynthetic bradyrhizobia. Proc. Natl. Acad. Sci. USA 2011, 108, 7183–7188. [Google Scholar] [CrossRef]
- Lindemann, A.; Pessi, G.; Schaefer, A.L.; Mattmann, M.E.; Christensen, Q.H.; Kessler, A.; Hennecke, H.; Blackwell, H.E.; Greenberg, E.P.; Harwood, C.S. Isovaleryl-homoserine lactone, an unusual branched-chain quorum-sensing signal from the soybean symbiont Bradyrhizobium japonicum. Proc. Natl. Acad. Sci. USA 2011, 108, 16765–16770. [Google Scholar] [CrossRef]
- Gooding, J.R.; May, A.L.; Hilliard, K.R.; Campagna, S.R. Establishing a quantitative definition of quorum sensing provides insight into the information content of the autoinducer signals in Vibrio harveyi and Escherichia coli. Biochemistry 2010, 49, 5621–5623. [Google Scholar] [CrossRef]
- Saenz, H.L.; Augsburger, V.; Vuong, C.; Jack, R.W.; Gotz, F.; Otto, M. Inducible expression and cellular location of AgrB, a protein involved in the maturation of the staphylococcal quorum-sensing pheromone. Arch. Microbiol. 2000, 174, 452–455. [Google Scholar] [CrossRef]
- Lee, J.; Wu, J.; Deng, Y.; Wang, J.; Wang, C.; Wang, J.; Chang, C.; Dong, Y.; Williams, P.; Zhang, L.H. A cell-cell communication signal integrates quorum sensing and stress response. Nat. Chem. Biol. 2013, 9, 339–343. [Google Scholar] [CrossRef]
- Pesci, E.C.; Milbank, J.B.; Pearson, J.P.; McKnight, S.; Kende, A.S.; Greenberg, E.P.; Iglewski, B.H. Quinolone signaling in the cell-to-cell communication system of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 1999, 96, 11229–11234. [Google Scholar] [CrossRef]
- Brachmann, A.O.; Brameyer, S.; Kresovic, D.; Hitkova, I.; Kopp, Y.; Manske, C.; Schubert, K.; Bode, H.B.; Heermann, R. Pyrones as bacterial signaling molecules. Nat. Chem. Biol. 2013, 9, 573–578. [Google Scholar] [CrossRef]
- Walters, M.; Sperandio, V. Autoinducer 3 and epinephrine signaling in the kinetics of locus of enterocyte effacement gene expression in enterohemorrhagic Escherichia coli. Infect. Immun. 2006, 74, 5445–5455. [Google Scholar] [CrossRef]
- Milton, D.L. Quorum sensing in vibrios: Complexity for diversification. Int. J. Med. Microbiol. 2006, 296, 61–71. [Google Scholar] [CrossRef]
- Henares, B.M.; Higgins, K.E.; Boon, E.M. Discovery of a nitric oxide responsive quorum sensing circuit in Vibrio harveyi. ACS Chem. Biol. 2012, 7, 1331–1336. [Google Scholar] [CrossRef]
- Lasarre, B.; Federle, M.J. Exploiting quorum sensing to confuse bacterial pathogens. Microbiol. Mol.Biol. Rev. 2013, 77, 73–111. [Google Scholar] [CrossRef]
- Yates, E.A.; Philipp, B.; Buckley, C.; Atkinson, S.; Chhabra, S.R.; Sockett, R.E.; Goldner, M.; Dessaux, Y.; Camara, M.; Smith, H.; et al. N-Acylhomoserine lactones undergo lactonolysis in a pH-, temperature-, and acyl chain length-dependent manner during growth of Yersinia pseudotuberculosis and Pseudomonas aeruginosa. Infect. Immun. 2002, 70, 5635–5646. [Google Scholar] [CrossRef]
- Taga, M.E.; Miller, S.T.; Bassler, B.L. Lsr-mediated transport and processing of AI-2 in Salmonella typhimurium. Mol. Microbiol. 2003, 50, 1411–1427. [Google Scholar] [CrossRef]
- Ng, W.L.; Perez, L.J.; Wei, Y.; Kraml, C.; Semmelhack, M.F.; Bassler, B.L. Signal production and detection specificity in Vibrio CqsA/CqsS quorum-sensing systems. Mol. Microbiol. 2011, 79, 1407–1417. [Google Scholar] [CrossRef]
- Tu, K.C.; Bassler, B.L. Multiple small RNAs act additively to integrate sensory information and control quorum sensing in Vibrio harveyi. Genes Dev. 2007, 21, 221–233. [Google Scholar] [CrossRef]
- Rutherford, S.T.; van Kessel, J.C.; Shao, Y.; Bassler, B.L. AphA and LuxR/HapR reciprocally control quorum sensing in vibrios. Genes Dev. 2011, 25, 397–408. [Google Scholar] [CrossRef]
- Van Kessel, J.C.; Rutherford, S.T.; Shao, Y.; Utria, A.F.; Bassler, B.L. Individual and combined roles of the master regulators AphA and LuxR in control of the Vibrio harveyi quorum-sensing regulon. J. Bacteriol. 2013, 195, 436–443. [Google Scholar] [CrossRef]
- Schauder, S.; Shokat, K.; Surette, M.G.; Bassler, B.L. The LuxS family of bacterial autoinducers: Biosynthesis of a novel quorum-sensing signal molecule. Mol. Microbiol. 2001, 41, 463–476. [Google Scholar] [CrossRef]
- Minogue, T.D.; Wehland-von Trebra, M.; Bernhard, F.; von Bodman, S.B. The autoregulatory role of EsaR, a quorum-sensing regulator in Pantoea stewartii ssp. stewartii: Evidence for a repressor function. Mol. Microbiol. 2002, 44, 1625–1635. [Google Scholar] [CrossRef]
- Dong, Y.H.; Wang, L.H.; Xu, J.L.; Zhang, H.B.; Zhang, X.F.; Zhang, L.H. Quenching quorum-sensing-dependent bacterial infection by an N-acyl homoserine lactonase. Nature 2001, 411, 813–817. [Google Scholar] [CrossRef]
- Eberhard, A.; Widrig, C.A.; McBath, P.; Schineller, J.B. Analogs of the autoinducer of bioluminescence in Vibrio fischeri. Arch. Microbiol. 1986, 146, 35–40. [Google Scholar] [CrossRef]
- Fuqua, C. The QscR quorum-sensing regulon of Pseudomonas aeruginosa: An orphan claims its identity. J. Bacteriol. 2006, 188, 3169–3171. [Google Scholar] [CrossRef]
- Kovacikova, G.; Skorupski, K. Regulation of virulence gene expression in Vibrio cholerae by quorum sensing: HapR functions at the aphA promoter. Mol. Microbiol. 2002, 46, 1135–1147. [Google Scholar] [CrossRef]
- Mattmann, M.E.; Shipway, P.M.; Heth, N.J.; Blackwell, H.E. Potent and selective synthetic modulators of a quorum sensing repressor in Pseudomonas aeruginosa identified from second-generation libraries of N-acylated L-homoserine lactones. ChemBioChem 2011, 12, 942–949. [Google Scholar] [CrossRef]
- Ng, W.L.; Perez, L.; Cong, J.; Semmelhack, M.F.; Bassler, B.L. Broad spectrum pro-quorum-sensing molecules as inhibitors of virulence in vibrios. PLoS Pathog. 2012, 8, e1002767. [Google Scholar] [CrossRef]
- Perez, L.J.; Karagounis, T.K.; Hurley, A.; Bassler, B.L.; Semmelhack, M.F. Highly potent, chemically stable quorum sensing agonists for Vibrio cholerae. Chem. Sci. 2014, 5, 151–155. [Google Scholar] [CrossRef]
- Clark, B.R.; Engene, N.; Teasdale, M.E.; Rowley, D.C.; Matainaho, T.; Valeriote, F.A.; Gerwick, W.H. Natural products chemistry and taxonomy of the marine cyanobacterium Blennothrix cantharidosmum. J. Nat. Prod. 2008, 71, 1530–1537. [Google Scholar] [CrossRef]
- Dobretsov, S.; Teplitski, M.; Alagely, A.; Gunasekera, S.P.; Paul, V.J. Malyngolide from the cyanobacterium Lyngbya majuscula interferes with quorum sensing circuitry. Environ. Microbiol. Rep. 2010, 2, 739–744. [Google Scholar] [CrossRef]
- Choi, H.; Mascuch, S.J.; Villa, F.A.; Byrum, T.; Teasdale, M.E.; Smith, J.E.; Preskitt, L.B.; Rowley, D.C.; Gerwick, L.; Gerwick, W.H. Honaucins A–C, potent inhibitors of inflammation and bacterial quorum sensing: Synthetic derivatives and structure-activity relationships. Chem. Biol. 2012, 19, 589–598. [Google Scholar] [CrossRef]
- Ooka, K.; Fukumoto, A.; Yamanaka, T.; Shimada, K.; Ishihara, R.; Anzai, Y.; Kato, F. Piericidins, novel quorum-sensing inhibitors against Chromobacterium violaceum CV026, from Streptomyces sp. TOHO-Y209 and TOHO-O348. Open J. Med. Chem. 2013, 3, 93–99. [Google Scholar]
- Teasdale, M.E.; Liu, J.Y.; Wallace, J.; Akhlaghi, F.; Rowley, D.C. Secondary metabolites produced by the marine bacterium Halobacillus salinus that inhibit quorum sensing-controlled phenotypes in Gram-negative bacteria. Appl. Environ. Microbiol. 2009, 75, 567–572. [Google Scholar]
- Teasdale, M.E.; Donovan, K.A.; Forschner-Dancause, S.R.; Rowley, D.C. Gram-positive marine bacteria as a potential resource for the discovery of quorum sensing inhibitors. Mar. Biotechnol. 2011, 13, 722–732. [Google Scholar] [CrossRef]
- Abed, R.M.; Dobretsov, S.; Al-Fori, M.; Gunasekera, S.P.; Sudesh, K.; Paul, V.J. Quorum-sensing inhibitory compounds from extremophilic microorganisms isolated from a hypersaline cyanobacterial mat. J. Ind. Microbiol. Biotechnol. 2013, 40, 759–772. [Google Scholar] [CrossRef]
- Peters, L.; König, G.M.; Wright, A.D.; Pukall, R.; Stackebrandt, E.; Eberl, L.; Riedel, K. Secondary metabolites of Flustra foliacea and their influence on bacteria. Appl. Environ. Microbiol. 2003, 69, 3469–3475. [Google Scholar] [CrossRef]
- Tello, E.; Castellanos, L.; Arevalo-Ferro, C.; Rodríguez, J.; Jiménez, C.; Duque, C. Absolute stereochemistry of antifouling cembranoid epimers at C-8 from the Caribbean octocoral Pseudoplexaura flagellosa. Revised structures of plexaurolones. Tetrahedron 2011, 67, 9112–9121. [Google Scholar] [CrossRef]
- Tello, E.; Castellanos, L.; Arevalo-Ferro, C.; Duque, C. Disruption in quorum-sensing systems and bacterial biofilm inhibition by cembranoid diterpenes isolated from the octocoral Eunicea knighti. J. Nat. Prod. 2012, 75, 1637–1642. [Google Scholar] [CrossRef]
- Kwan, J.C.; Teplitski, M.; Gunasekera, S.P.; Paul, V.J.; Luesch, H. Isolation and biological evaluation of 8-epi-malyngamide C from the Floridian marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2010, 73, 463–466. [Google Scholar] [CrossRef]
- Kwan, J.C.; Meickle, T.; Ladwa, D.; Teplitski, M.; Paul, V.; Luesch, H. Lyngbyoic acid, a “tagged” fatty acid from a marine cyanobacterium, disrupts quorum sensing in Pseudomonas aeruginos. Mol. Biosyst. 2011, 7, 1205–1216. [Google Scholar] [CrossRef]
- Montaser, R.; Paul, V.J.; Luesch, H. Modular strategies for structure and function employed by marine cyanobacteria: Characterization and synthesis of pitinoic acids. Org. Lett. 2013, 15, 4050–4053. [Google Scholar] [CrossRef]
- Dobretsov, S.; Teplitski, M.; Bayer, M.; Gunasekera, S.; Proksch, P.; Paul, V.J. Inhibition of marine biofouling by bacterial quorum sensing inhibitors. Biofouling 2011, 27, 893–905. [Google Scholar] [CrossRef]
- Wang, L.; Zou, S.; Yin, S.; Liu, H.; Yu, W.; Gong, Q. Construction of an effective screening system for detection of Pseudomonas aeruginosa quorum sensing inhibitors and its application in bioautographic thin-layer chromatography. Biotechnol. Lett. 2011, 33, 1381–1387. [Google Scholar] [CrossRef]
- Liu, H.B.; Koh, K.P.; Kim, J.S.; Seo, Y.; Park, S. The effects of betonicine, floridoside, and isethionic acid from the red alga Ahnfeltiopsis flabelliformis on quorum-sensing activity. Biotechnol. Bioprocess Eng. 2008, 13, 458–463. [Google Scholar] [CrossRef]
- Skindersoe, M.E.; Ettinger-Epstein, P.; Rasmussen, T.B.; Bjarnsholt, T.; de Nys, R.; Givskov, M. Quorum sensing antagonism from marine organisms. Mar. Biotechnol. (N. Y.) 2008, 10, 56–63. [Google Scholar] [CrossRef]
- Chu, Y.Y.; Nega, M.; Wolfle, M.; Plener, L.; Grond, S.; Jung, K.; Gotz, F. A new class of quorum quenching molecules from Staphylococcus species affects communication and growth of Gram-negative bacteria. PLoS Pathog. 2013, 9, e1003654. [Google Scholar] [CrossRef]
- Singh, V.K.; Kavita, K.; Prabhakaran, R.; Jha, B. Cis-9-octadecenoic acid from the rhizospheric bacterium Stenotrophomonas maltophilia BJ01 shows quorum quenching and anti-biofilm activities. Biofouling 2013, 29, 855–867. [Google Scholar] [CrossRef]
- Bobadilla Fazzini, R.A.; Skindersoe, M.E.; Bielecki, P.; Puchalka, J.; Givskov, M.; Martins sos Santos, V.A. Protoanemonin: A natural quorum sensing inhibitor that selectively activates iron starvation response. Environ. Microbiol. 2013, 15, 111–120. [Google Scholar] [CrossRef]
- HjelmgaardRasmussen, T.B.; Skindersoe, M.E.; Bjarnsholt, T.; Phipps, R.K.; Christensen, K.B.; Jensen, P.O.; Andersen, J.B.; Koch, B.; Larsen, T.O.; Hentzer, M.; et al. Identity and effects of quorum-sensing inhibitors produced by Penicillium species. Microbiology 2005, 151, 1325–1340. [Google Scholar] [CrossRef]
- Truchado, P.; Lopez-Galvez, F.; Gil, M.I.; Tomas-Barberan, F.A.; Allende, A. Quorum sensing inhibitory and antimicrobial activities of honeys and the relationship with individual phenolics. Food Chem. 2009, 115, 1337–1344. [Google Scholar] [CrossRef]
- Bulman, Z.; Le, P.; Hudson, A.O.; Savka, M.A. A novel property of propolis (bee glue): Anti-pathogenic activity by inhibition of N-acyl-homoserine lactone mediated signaling in bacteria. J. Ethnopharmacol. 2011, 138, 788–797. [Google Scholar] [CrossRef]
- Park, J.; Kaufmann, G.F.; Bowen, J.P.; Arbiser, J.L.; Janda, K.D. Solenopsin A, a venom alkaloid from the fire ant Solenopsis invicta, inhibits quorum-sensing signaling in Pseudomonas aeruginosa. J. Infect. Dis. 2008, 198, 1198–1201. [Google Scholar] [CrossRef]
- Zhou, L.; Zheng, H.; Tang, Y.; Yu, W.; Gong, Q. Eugenol inhibits quorum sensing at sub-inhibitory concentrations. Biotechnol. Lett. 2013, 35, 631–637. [Google Scholar] [CrossRef]
- Keshavan, N.D.; Chowdhary, P.K.; Haines, D.C.; Gonzalez, J.E. l-canavanine made by Medicago sativa interferes with quorum sensing in Sinorhizobium meliloti. J. Bacteriol. 2005, 187, 8427–8436. [Google Scholar] [CrossRef]
- Vandeputte, O.M.; Kiendrebeogo, M.; Rajaonson, S.; Diallo, B.; Mol, A.; El Jaziri, M.; Baucher, M. Identification of catechin as one of the flavonoids from Combretum albiflorum bark extract that reduces the production of quorum-sensing-controlled virulence factors in Pseudomonas aeruginosa PAO1. Appl. Environ. Microbiol. 2010, 76, 243–253. [Google Scholar] [CrossRef]
- Vandeputte, O.M.; Kiendrebeogo, M.; Rasamiravaka, T.; Stevigny, C.; Duez, P.; Rajaonson, S.; Diallo, B.; Mol, A.; Baucher, M.; El Jaziri, M. The flavanone naringenin reduces the production of quorum sensing-controlled virulence factors in Pseudomonas aeruginosa PAO1. Microbiology 2011, 157, 2120–2132. [Google Scholar] [CrossRef]
- Amaya, S.; Pereira, J.A.; Borkosky, S.A.; Valdez, J.C.; Bardon, A.; Arena, M.E. Inhibition of quorum sensing in Pseudomonas aeruginosa by sesquiterpene lactones. Phytomedicine 2012, 19, 1173–1177. [Google Scholar] [CrossRef]
- Jakobsen, T.H.; van Gennip, M.; Phipps, R.K.; Shanmugham, M.S.; Christensen, L.D.; Alhede, M.; Skindersoe, M.E.; Rasmussen, T.B.; Friedrich, K.; Uthe, F.; et al. Ajoene, a sulfur-rich molecule from garlic, inhibits genes controlled by quorum sensing. Antimicrob. Agents Chemother. 2012, 56, 2314–2325. [Google Scholar] [CrossRef] [Green Version]
- Jakobsen, T.H.; Bragason, S.K.; Phipps, R.K.; Christensen, L.D.; van Gennip, M.; Alhede, M.; Skindersoe, M.; Larsen, T.O.; Hoiby, N.; Bjarnsholt, T.; et al. Food as a source for quorum sensing inhibitors: Iberin from horseradish revealed as a quorum sensing inhibitor of Pseudomonas aeruginosa. Appl. Environ. Microbiol. 2012, 78, 2410–2421. [Google Scholar] [CrossRef]
- Chong, Y.M.; Yin, W.F.; Ho, C.Y.; Mustafa, M.R.; Hadi, A.H.A.; Awang, K.; Narrima, P.; Koh, C.L.; Appleton, D.R.; Chan, K.G. Malabaricone C from Myristica cinnamomea exhibits anti-quorum sensing activity. J. Nat. Prod. 2011, 74, 2261–2264. [Google Scholar]
- Paza, C.; Carcamo, G.; Silva, M.; Becerra, J.; Urrutia, H.; Sossa, K. Drimendiol, a drimane sesquiterpene with quorum sensing inhibition activity. Nat. Prod. Commun. 2013, 8, 147–148. [Google Scholar]
- Packiavathy, I.A.; Priya, S.; Pandian, S.K.; Ravi, A.V. Inhibition of biofilm development of uropathogens by curcumin-an anti-quorum sensing agent from Curcuma longa. Food Chem. 2014, 148, 453–460. [Google Scholar] [CrossRef]
- Bodini, S.F.; Manfredini, S.; Epp, M.; Valentini, S.; Santori, F. Quorum sensing inhibition activity of garlic extract and p-coumaric acid. Lett. Appl. Microbiol. 2009, 49, 551–555. [Google Scholar] [CrossRef]
- Giménez-Bastida, J.A.; Truchado, P.; Larrosa, M.; Espín, J.C.; Tomás-Barberán, F.A.; Allende, A.; García-Conesa, M.T. Urolithins, ellagitannin metabolites produced by colon microbiota, inhibit quorum sensing in Yersinia enterocolitica: Phenotypic response and associated molecular changes. Food Chem. 2012, 132, 1465–1474. [Google Scholar]
- Adonizio, A.; Dawlaty, J.; Ausubel, F.M.; Clardy, J.; Mathee, K. Ellagitannins from Conocarpus erectus exhibit anti-quorum sensing activity against Pseudomonas aeruginosa. Planta Med. 2008, 74, 1035–1035. [Google Scholar]
- Kaplan, F.; Badri, D.V.; Zachariah, C.; Ajredini, R.; Sandoval, F.J.; Roje, S.; Levine, L.H.; Zhang, F.L.; Robinette, S.L.; Alborn, H.T.; et al. Bacterial attraction and quorum sensing inhibition in Caenorhabditis elegans exudates. J. Chem. Ecol. 2009, 35, 878–892. [Google Scholar] [CrossRef]
- Teplitski, M.; Chen, H.; Rajamani, S.; Gao, M.; Merighi, M.; Sayre, R.T.; Robinson, J.B.; Rolfe, B.G.; Bauer, W.D. Chlamydomonas reinhardtii secretes compounds that mimic bacterial signals and interfere with quorum sensing regulation in bacteria. Plant Physiol. 2004, 134, 137–146. [Google Scholar] [CrossRef]
- Rajamani, S.; Bauer, W.D.; Robinson, J.B.; Farrow, J.M.; Pesci, E.C.; Teplitski, M.; Gao, M.S.; Sayre, R.T.; Phillips, D.A. The vitamin riboflavin and its derivative lumichrome activate the LasR bacterial quorum-sensing receptor. Mol. Plant-Microbe Interact. 2008, 21, 1184–1192. [Google Scholar] [CrossRef]
- Ding, X.; Yin, B.; Qian, L.; Zeng, Z.R.; Yang, Z.L.; Li, H.X.; Lu, Y.J.; Zhou, S.N. Screening for novel quorum-sensing inhibitors to interfere with the formation of Pseudomonas aeruginosa biofilm. J. Med. Microbiol. 2011, 60, 1827–1834. [Google Scholar] [CrossRef]
- Zeng, Z.; Qian, L.; Cao, L.; Tan, H.; Huang, Y.; Xue, X.; Shen, Y.; Zhou, S. Virtual screening for novel quorum sensing inhibitors to eradicate biofilm formation of Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 2008, 79, 119–126. [Google Scholar] [CrossRef]
- Cugini, C.; Calfee, M.W.; Farrow, J.M., 3rd.; Morales, D.K.; Pesci, E.C.; Hogan, D.A. Farnesol, a common sesquiterpene, inhibits PQS production in Pseudomonas aeruginosa. Mol. Microbiol. 2007, 65, 896–906. [Google Scholar] [CrossRef]
- De Nys, R.; Wright, A.D.; König, G.M.; Sticher, O. New halogenated furanones from the marine alga Delisea pulchra (cf. fimbriata). Tetrahedron 1993, 49, 11213–11220. [Google Scholar] [CrossRef]
- Hentzer, M.; Riedel, K.; Rasmussen, T.B.; Heydorn, A.; Andersen, J.B.; Parsek, M.R.; Rice, S.A.; Eberl, L.; Molin, S.; Hoiby, N.; et al. Inhibition of quorum sensing in Pseudomonas aeruginosa biofilm bacteria by a halogenated furanone compound. Microbiology 2002, 148, 87–102. [Google Scholar]
- Brackman, G.; Celen, S.; Hillaert, U.; van Calenbergh, S.; Cos, P.; Maes, L.; Nelis, H.J.; Coenye, T. Structure-activity relationship of cinnamaldehyde analogs as inhibitors of AI-2 based quorum sensing and their effect on virulence of Vibrio spp. PLoS One 2011, 6, e16084. [Google Scholar] [CrossRef] [Green Version]
- Niu, C.; Afre, S.; Gilbert, E.S. Subinhibitory concentrations of cinnamaldehyde interfere with quorum sensing. Lett. Appl. Microbiol. 2006, 43, 489–494. [Google Scholar] [CrossRef]
- Brackman, G.; Defoirdt, T.; Miyamoto, C.; Bossier, P.; van Calenbergh, S.; Nelis, H.; Coenye, T. Cinnamaldehyde and cinnamaldehyde derivatives reduce virulence in Vibrio spp. by decreasing the DNA-binding activity of the quorum sensing response regulator LuxR. BMC Microbiol. 2008, 8, 149. [Google Scholar] [CrossRef]
- Lee, K.M.; Lim, J.; Nam, S.; Yoon, M.Y.; Kwon, Y.K.; Jung, B.Y.; Park, Y.; Park, S.; Yoon, S.S. Inhibitory effects of broccoli extract on Escherichia coli O157:H7 quorum sensing and in vivo virulence. FEMS Microbiol. Lett. 2011, 321, 67–74. [Google Scholar] [CrossRef]
- Vikram, A.; Jesudhasan, P.R.; Jayaprakasha, G.K.; Pillai, B.S.; Patil, B.S. Grapefruit bioactive limonoids modulate E. coli O157:H7 TTSS and biofilm. Int. J. Food Microbiol. 2010, 140, 109–116. [Google Scholar] [CrossRef]
- Li, J.; Wang, W.; Xu, S.X.; Magarvey, N.A.; McCormick, J.K. Lactobacillus reuteri-produced cyclic dipeptides quench agr-mediated expression of toxic shock syndrome toxin-1 in staphylococci. Proc. Natl. Acad. Sci. USA 2011, 108, 3360–3365. [Google Scholar]
- Mansson, M.; Nielsen, A.; Kjaerulff, L.; Gotfredsen, C.H.; Wietz, M.; Ingmer, H.; Gram, L.; Larsen, T.O. Inhibition of virulence gene expression in Staphylococcus aureus by novel depsipeptides from a marine Photobacterium. Mar. Drugs 2011, 9, 2537–2552. [Google Scholar] [CrossRef]
- Kiran, M.D.; Adikesavan, N.V.; Cirioni, O.; Giacometti, A.; Silvestri, C.; Scalise, G.; Ghiselli, R.; Saba, V.; Orlando, F.; Shoham, M.; et al. Discovery of a quorum-sensing inhibitor of drug-resistant staphylococcal infections by structure-based virtual screening. Mol. Pharmacol. 2008, 73, 1578–1586. [Google Scholar] [CrossRef]
- Duncan, M.C.; Wong, W.R.; Dupzyk, A.J.; Bray, W.M.; Linington, R.G.; Auerbuch, V. An NF-kappaB-based high-throughput screen identifies piericidins as inhibitors of the Yersinia pseudotuberculosis type III secretion system. Antimicrob. Agents Chemother. 2014, 58, 1118–1126. [Google Scholar] [CrossRef]
- De Nys, R.; Steinberg, P.; Willemsen, P.; Dworjanyn, S.; Gabelish, C.; King, R. Broad spectrum effects of secondary metabolites from the red alga Delisea pulchra in antifouling assays. Biofouling 1995, 8, 259–271. [Google Scholar] [CrossRef]
- Martinelli, D.; Grossmann, G.; Sequin, U.; Brandl, H.; Bachofen, R. Effects of natural and chemically synthesized furanones on quorum sensing in Chromobacterium violaceum. BMC Microbiol. 2004, 4, 25. [Google Scholar] [CrossRef] [Green Version]
- Manefield, M.; Rasmussen, T.B.; Henzter, M.; Andersen, J.B.; Steinberg, P.; Kjelleberg, S.; Givskov, M. Halogenated furanones inhibit quorum sensing through accelerated LuxR turnover. Microbiology 2002, 148, 1119–1127. [Google Scholar]
- Defoirdt, T.; Miyamoto, C.M.; Wood, T.K.; Meighen, E.A.; Sorgeloos, P.; Verstraete, W.; Bossier, P. The natural furanone (5Z)-4-bromo-5-(bromomethylene)-3-butyl-2(5H)-furanone disrupts quorum sensing-regulated gene expression in Vibrio harveyi by decreasing the DNA-binding activity of the transcriptional regulator protein luxR. Environ. Microbiol. 2007, 9, 2486–2495. [Google Scholar] [CrossRef]
- Zang, T.; Lee, B.W.; Cannon, L.M.; Ritter, K.A.; Dai, S.; Ren, D.; Wood, T.K.; Zhou, Z.S. A naturally occurring brominated furanone covalently modifies and inactivates LuxS. Bioorg. Med. Chem. Lett. 2009, 19, 6200–6204. [Google Scholar] [CrossRef]
- Defoirdt, T.; Crab, R.; Wood, T.K.; Sorgeloos, P.; Verstraete, W.; Bossier, P. Quorum sensing-disrupting brominated furanones protect the gnotobiotic brine shrimp Artemia franciscana from pathogenic Vibrio harveyi, Vibrio campbellii, and Vibrio parahaemolyticus isolates. Appl. Environ. Microbiol. 2006, 72, 6419–6423. [Google Scholar] [CrossRef] [Green Version]
- Rasch, M.; Buch, C.; Austin, B.; Slierendrecht, W.J.; Ekmann, K.S.; Larsen, J.L.; Johansen, C.; Riedel, K.; Eberl, L.; Givskov, M.; et al. An inhibitor of bacterial quorum sensing reduces mortalities caused by vibriosis in rainbow trout (Oncorhynchus mykiss, Walbaum). Syst. Appl. Microbiol. 2004, 27, 350–359. [Google Scholar] [CrossRef]
- Tinh, N.T.N.; Linh, N.D.; Wood, T.K.; Dierckens, K.; Sorgeloos, P.; Bossier, P. Interference with the quorum sensing systems in a Vibrio harveyi strain alters the growth rate of gnotobiotically cultured rotifer Brachionus plicatilis. J. Appl. Microbiol. 2007, 103, 194–203. [Google Scholar] [CrossRef]
- Wu, H.; Song, Z.; Hentzer, M.; Andersen, J.B.; Molin, S.; Givskov, M.; Hoiby, N. Synthetic furanones inhibit quorum-sensing and enhance bacterial clearance in Pseudomonas aeruginosa lung infection in mice. J. Antimicrob. Chemother. 2004, 53, 1054–1061. [Google Scholar] [CrossRef]
- Natrah, F.M.I.; Kenmegne, M.M.; Wiyoto, W.; Sorgeloos, P.; Bossier, P.; Defoirdt, T. Effects of micro-algae commonly used in aquaculture on acyl-homoserine lactone quorum sensing. Aquaculture 2011, 317, 53–57. [Google Scholar] [CrossRef]
- Havsteen, B.H. The biochemistry and medical significance of the flavonoids. Pharmacol. Ther. 2002, 96, 67–202. [Google Scholar] [CrossRef]
- Vikram, A.; Jayaprakasha, G.K.; Jesudhasan, P.R.; Pillai, S.D.; Patil, B.S. Suppression of bacterial cell-cell signalling, biofilm formation and type III secretion system by citrus flavonoids. J. Appl. Microbiol. 2010, 109, 515–527. [Google Scholar]
- Pande, G.S.J.; Scheie, A.A.; Benneche, T.; Wille, M.; Sorgeloos, P.; Bossier, P.; Defoirdt, T. Quorum sensing-disrupting compounds protect larvae of the giant freshwater prawn Macrobrachium rosenbergii from Vibrio harveyi infection. Aquaculture 2013, 406–407, 121–124. [Google Scholar] [CrossRef]
- Defoirdt, T.; Brackman, G.; Coenye, T. Quorum sensing inhibitors: How strong is the evidence? Trends Microbiol. 2013, 21, 619–624. [Google Scholar] [CrossRef]
- Ni, N.; Choudhary, G.; Li, M.; Wang, B. Pyrogallol and its analogs can antagonize bacterial quorum sensing in Vibrio harveyi. Bioorg. Med. Chem. Lett. 2008, 18, 1567–1572. [Google Scholar] [CrossRef]
- Defoirdt, T.; Pande, G.S.; Baruah, K.; Bossier, P. The apparent quorum-sensing inhibitory activity of pyrogallol is a side effect of peroxide production. Antimicrob. Agents Chemother. 2013, 57, 2870–2873. [Google Scholar] [CrossRef]
- Newman, K.L.; Chatterjee, S.; Ho, K.A.; Lindow, S.E. Virulence of plant pathogenic bacteria attenuated by degradation of fatty acid cell-to-cell signaling factors. Mol. Plant Microbe Interact. 2008, 21, 326–334. [Google Scholar] [CrossRef]
- Pustelny, C.; Albers, A.; Buldt-Karentzopoulos, K.; Parschat, K.; Chhabra, S.R.; Camara, M.; Williams, P.; Fetzner, S. Dioxygenase-mediated quenching of quinolone-dependent quorum sensing in Pseudomonas aeruginosa. Chem. Biol. 2009, 16, 1259–1267. [Google Scholar] [CrossRef]
- Roy, V.; Fernandes, R.; Tsao, C.Y.; Bentley, W.E. Cross species quorum quenching using a native AI-2 processing enzyme. ACS Chem. Biol. 2010, 5, 223–232. [Google Scholar] [CrossRef]
- Draganov, D.I.; Teiber, J.F.; Speelman, A.; Osawa, Y.; Sunahara, R.; La Du, B.N. Human paraoxonases (PON1, PON2, and PON3) are lactonases with overlapping and distinct substrate specificities. J. Lipid Res. 2005, 46, 1239–1247. [Google Scholar] [CrossRef]
- Tang, K.; Zhang, Y.; Yu, M.; Shi, X.; Coenye, T.; Bossier, P.; Zhang, X.H. Evaluation of a new high-throughput method for identifying quorum quenching bacteria. Sci. Rep. 2013, 3, 2935. [Google Scholar] [Green Version]
- Zhang, X.H.; Ocean University of China, Qingdao, China. MomL, a novel marine-derived N-acyl homeserine lactonase in Muricauda olearia. 2014; Unpublished work. [Google Scholar]
- Dong, Y.H.; Xu, J.L.; Li, X.Z.; Zhang, L.H. AiiA, an enzyme that inactivates the acylhomoserine lactone quorum-sensing signal and attenuates the virulence of Erwinia carotovora. Proc. Natl. Acad. Sci. USA 2000, 97, 3526–3531. [Google Scholar] [CrossRef]
- Thomas, P.W.; Stone, E.M.; Costello, A.L.; Tierney, D.L.; Fast, W. The quorum-quenching lactonase from Bacillus thuringiensis is a metalloprotein. Biochemistry 2005, 44, 7559–7569. [Google Scholar] [CrossRef]
- Kim, M.H.; Choi, W.C.; Kang, H.O.; Lee, J.S.; Kang, B.S.; Kim, K.J.; Derewenda, Z.S.; Oh, T.K.; Lee, C.H.; Lee, J.K. The molecular structure and catalytic mechanism of a quorum-quenching N-acyl-L-homoserine lactone hydrolase. Proc. Natl. Acad. Sci. USA 2005, 102, 17606–17611. [Google Scholar] [CrossRef]
- Liu, D.; Momb, J.; Thomas, P.W.; Moulin, A.; Petsko, G.A.; Fast, W.; Ringe, D. Mechanism of the quorum-quenching lactonase (AiiA) from Bacillus thuringiensis. 1. product-bound structures. Biochemistry 2008, 47, 7706–7714. [Google Scholar] [CrossRef]
- Momb, J.; Wang, C.; Liu, D.; Thomas, P.W.; Petsko, G.A.; Guo, H.; Ringe, D.; Fast, W. Mechanism of the quorum-quenching lactonase (AiiA) from Bacillus thuringiensis. 2. substrate modeling and active site mutations. Biochemistry 2008, 47, 7715–7725. [Google Scholar] [CrossRef]
- Liu, C.F.; Liu, D.; Momb, J.; Thomas, P.W.; Lajoie, A.; Petsko, G.A.; Fast, W.; Ringe, D. A phenylalanine clamp controls substrate specificity in the quorum-quenching metallo-gamma-lactonase from Bacillus thuringiensis. Biochemistry 2013, 52, 1603–1610. [Google Scholar] [CrossRef]
- Momb, J.; Thomas, P.W.; Breece, R.M.; Tierney, D.L.; Fast, W. The quorum-quenching metallo-gamma-lactonase from Bacillus thuringiensis exhibits a leaving group thio effect. Biochemistry 2006, 45, 13385–13393. [Google Scholar] [CrossRef]
- Carlier, A.; Uroz, S.; Smadja, B.; Fray, R.; Latour, X.; Dessaux, Y.; Faure, D. The Ti plasmid of Agrobacterium tumefaciens harbors an attM-paralogous gene, aiiB, also encoding N-acyl homoserine lactonase activity. Appl. Environ. Microbiol. 2003, 69, 4989–4993. [Google Scholar] [CrossRef]
- Liu, D.; Thomas, P.W.; Momb, J.; Hoang, Q.Q.; Petsko, G.A.; Ringe, D.; Fast, W. Structure and specificity of a quorum-quenching lactonase (AiiB) from Agrobacterium tumefaciens. Biochemistry 2007, 46, 11789–11799. [Google Scholar] [CrossRef]
- Piper, K.R.; Beck von Bodman, S.; Farrand, S.K. Conjugation factor of Agrobacterium tumefaciens regulates Ti plasmid transfer by autoinduction. Nature 1993, 362, 448–450. [Google Scholar] [CrossRef]
- Haudecoeur, E.; Planamente, S.; Cirou, A.; Tannieres, M.; Shelp, B.J.; Morera, S.; Faure, D. Proline antagonizes GABA-induced quenching of quorum-sensing in Agrobacterium tumefaciens. Proc. Natl. Acad. Sci. USA 2009, 106, 14587–14592. [Google Scholar] [CrossRef]
- Wang, W.Z.; Morohoshi, T.; Someya, N.; Ikeda, T. AidC, a novel N-acylhomoserine lactonase from the potato root-associated cytophaga-flavobacteria-bacteroides (CFB) group bacterium Chryseobacterium sp. strain StRB126. Appl. Environ. Microbiol. 2012, 78, 7985–7992. [Google Scholar] [CrossRef]
- Krysciak, D.; Schmeisser, C.; Preuss, S.; Riethausen, J.; Quitschau, M.; Grond, S.; Streit, W.R. Involvement of multiple loci in quorum quenching of autoinducer I molecules in the nitrogen-fixing symbiont Rhizobium (Sinorhizobium) sp. strain StRB126. Appl. Environ. Microbiol. 2011, 77, 5089–5099. [Google Scholar]
- Schneider, J.; Yepes, A.; Garcia-Betancur, J.C.; Westedt, I.; Mielich, B.; Lopez, D. Streptomycin-induced expression in Bacillus subtilis of YtnP, a lactonase-homologous protein that inhibits development and streptomycin production in Streptomyces griseus. Appl. Environ. Microbiol. 2012, 78, 599–603. [Google Scholar] [CrossRef]
- Afriat, L.; Roodveldt, C.; Manco, G.; Tawfik, D.S. The latent promiscuity of newly identified microbial lactonases is linked to a recently diverged phosphotriesterase. Biochemistry 2006, 45, 13677–13686. [Google Scholar] [CrossRef]
- Elias, M.; Dupuy, J.; Merone, L.; Mandrich, L.; Porzio, E.; Moniot, S.; Rochu, D.; Lecomte, C.; Rossi, M.; Masson, P.; et al. Structural basis for natural lactonase and promiscuous phosphotriesterase activities. J. Mol. Biol. 2008, 379, 1017–1028. [Google Scholar]
- Ng, F.S.; Wright, D.M.; Seah, S.Y. Characterization of a phosphotriesterase-like lactonase from Sulfolobus solfataricus and its immobilization for disruption of quorum sensing. Appl. Environ. Microbiol. 2011, 77, 1181–1186. [Google Scholar] [CrossRef]
- Xue, B.; Chow, J.Y.; Baldansuren, A.; Yap, L.L.; Gan, Y.H.; Dikanov, S.A.; Robinson, R.C.; Yew, W.S. Structural evidence of a productive active site architecture for an evolved quorum-quenching GKL lactonase. Biochemistry 2013, 52, 2359–2370. [Google Scholar] [CrossRef]
- Hiblot, J.; Gotthard, G.; Chabriere, E.; Elias, M. Structural and enzymatic characterization of the lactonase SisLac from Sulfolobus islandicus. PLoS One 2012, 7, e47028. [Google Scholar]
- Mei, G.Y.; Yan, X.X.; Turak, A.; Luo, Z.Q.; Zhang, L.Q. AidH, an alpha/beta-hydrolase fold family member from an Ochrobactrum sp. strain, is a novel N-acylhomoserine lactonase. Appl. Environ. Microbiol. 2010, 76, 4933–4942. [Google Scholar] [CrossRef]
- Wang, W.Z.; Morohoshi, T.; Ikenoya, M.; Someya, N.; Ikeda, T. AiiM, a novel class of N-acylhomoserine lactonase from the leaf-associated bacterium Microbacterium testaceum. Appl. Environ. Microbiol. 2010, 76, 2524–2530. [Google Scholar] [CrossRef]
- Lin, Y.H.; Xu, J.L.; Hu, J.; Wang, L.H.; Ong, S.L.; Leadbetter, J.R.; Zhang, L.H. Acyl-homoserine lactone acylase from Ralstonia strain XJ12B represents a novel and potent class of quorum-quenching enzymes. Mol. Microbiol. 2003, 47, 849–860. [Google Scholar] [CrossRef]
- Leadbetter, J.R.; Greenberg, E.P. Metabolism of acyl-homoserine lactone quorum-sensing signals by Variovorax paradoxus. J. Bacteriol. 2000, 182, 6921–6926. [Google Scholar]
- Park, S.Y.; Kang, H.O.; Jang, H.S.; Lee, J.K.; Koo, B.T.; Yum, D.Y. Identification of extracellular N-acylhomoserine lactone acylase from a Streptomyces sp. and its application to quorum quenching. Appl. Environ. Microbiol. 2005, 71, 2632–2641. [Google Scholar] [CrossRef]
- Han, J.I.; Choi, H.K.; Lee, S.W.; Orwin, P.M.; Kim, J.; Laroe, S.L.; Kim, T.G.; O’Neil, J.; Leadbetter, J.R.; Lee, S.Y.; et al. Complete genome sequence of the metabolically versatile plant growth-promoting endophyte Variovorax paradoxus S110. J. Bacteriol. 2011, 193, 1183–1190. [Google Scholar] [CrossRef]
- Sio, C.F.; Otten, L.G.; Cool, R.H.; Diggle, S.P.; Braun, P.G.; Bos, R.; Daykin, M.; Cámara, M.; Williams, P.; Quax, W.J. Quorum quenching by an N-acyl-homoserine lactone acylase from Pseudomonas aeruginosa PAO1. Infect. Immun. 2006, 74, 1673–1682. [Google Scholar] [CrossRef]
- Huang, J.J.; Petersen, A.; Whiteley, M.; Leadbetter, J.R. Identification of QuiP, the product of gene PA1032, as the second acyl-homoserine lactone acylase of Pseudomonas aeruginosa PAO1. Appl. Environ. Microbiol. 2006, 72, 1190–1197. [Google Scholar] [CrossRef]
- Wahjudi, M.; Papaioannou, E.; Hendrawati, O.; van Assen, A.H.G.; van Merkerk, R.; Cool, R.H.; Poelarends, G.J.; Quax, W.J. PA0305 of Pseudomonas aeruginosa is a quorum quenching acylhomoserine lactone acylase belonging to the Ntn hydrolase superfamily. Microbiology 2011, 157, 2042–2055. [Google Scholar] [CrossRef]
- Nadal Jimenez, P.; Koch, G.; Papaioannou, E.; Wahjudi, M.; Krzeslak, J.; Coenye, T.; Cool, R.H.; Quax, W.J. Role of PvdQ in Pseudomonas aeruginosa virulence under iron-limiting conditions. Microbiology 2010, 156, 49–59. [Google Scholar] [CrossRef]
- Clevenger, K.D.; Wu, R.; Er, J.A.; Liu, D.; Fast, W. Rational design of a transition state analogue with picomolar affinity for Pseudomonas aeruginosa PvdQ, a siderophore biosynthetic enzyme. ACS Chem. Biol. 2013, 8, 2192–2200. [Google Scholar] [CrossRef]
- Czajkowski, R.; Krzyżanowska, D.; Karczewska, J.; Atkinson, S.; Przysowa, J.; Lojkowska, E.; Williams, P.; Jafra, S. Inactivation of AHLs by Ochrobactrum sp. A44 depends on the activity of a novel class of AHL acylase. Environ. Microbiol. Rep. 2011, 3, 59–68. [Google Scholar] [CrossRef]
- Tannieres, M.; Beury-Cirou, A.; Vigouroux, A.; Mondy, S.; Pellissier, F.; Dessaux, Y.; Faure, D. A metagenomic study highlights phylogenetic proximity of quorum-quenching and xenobiotic-degrading amidases of the AS-family. PLoS One 2013, 8, e65473. [Google Scholar]
- Uroz, S.; Chhabra, S.R.; Camara, M.; Williams, P.; Oger, P.; Dessaux, Y. N-Acylhomoserine lactone quorum-sensing molecules are modified and degraded by Rhodococcus erythropolis W2 by both amidolytic and novel oxidoreductase activities. Microbiology 2005, 151, 3313–3322. [Google Scholar] [CrossRef]
- Chowdhary, P.K.; Keshavan, N.; Nguyen, H.Q.; Peterson, J.A.; Gonzalez, J.E.; Haines, D.C. Bacillus megaterium CYP102A1 oxidation of acyl homoserine lactones and acyl homoserines. Biochemistry 2007, 46, 14429–14437. [Google Scholar] [CrossRef]
- Bijtenhoorn, P.; Mayerhofer, H.; Muller-Dieckmann, J.; Utpatel, C.; Schipper, C.; Hornung, C.; Szesny, M.; Grond, S.; Thurmer, A.; Brzuszkiewicz, E.; et al. A novel metagenomic short-chain dehydrogenase/reductase attenuates Pseudomonas aeruginosa biofilm formation and virulence on Caenorhabditis elegans. PLoS One 2011, 6, e26278. [Google Scholar] [CrossRef]
- Chan, K.G.; Atkinson, S.; Mathee, K.; Sam, C.K.; Chhabra, S.R.; Camara, M.; Koh, C.L.; Williams, P. Characterization of N-acylhomoserine lactone-degrading bacteria associated with the Zingiber officinale (ginger) rhizosphere: Co-existence of quorum quenching and quorum sensing in Acinetobacter and Burkholderia. BMC Microbiol. 2011, 11, 51. [Google Scholar] [CrossRef]
- Hong, K.-W.; Koh, C.-L.; Sam, C.-K.; Yin, W.-F.; Chan, K.-G. Complete genome sequence of Burkholderia sp. strain GG4, a Betaproteobacterium that reduces 3-oxo-N-acylhomoserine lactones and produces different N-acylhomoserine lactones. J. Bacteriol. 2012, 194, 6317. [Google Scholar] [CrossRef]
- Sandy, M.; Carter-Franklin, J.N.; Martin, J.D.; Butler, A. Vanadium bromoperoxidase from Delisea pulchra: Enzyme-catalyzed formation of bromofuranone and attendant disruption of quorum sensing. Chem. Commun. 2011, 47, 12086–12088. [Google Scholar] [CrossRef]
- Borchardt, S.A.; Allain, E.J.; Michels, J.J.; Stearns, G.W.; Kelly, R.F.; McCoy, W.F. Reaction of acylated homoserine lactone bacterial signaling molecules with oxidized halogen antimicrobials. Appl. Environ. Microbiol. 2001, 67, 3174–3179. [Google Scholar] [CrossRef]
- Syrpas, M.; Ruysbergh, E.; Blommaert, L.; Vanelslander, B.; Sabbe, K.; Vyverman, W.; de Kimpe, N.; Mangelinckx, S. Haloperoxidase mediated quorum quenching by Nitzschia cf pellucida: Study of the metabolization of N-acyl homoserine lactones by a benthic diatom. Mar. Drugs 2014, 12, 352–367. [Google Scholar] [CrossRef] [Green Version]
- Hibbing, M.E.; Fuqua, C.; Parsek, M.R.; Peterson, S.B. Bacterial competition: Surviving and thriving in the microbial jungle. Nat. Rev. Microbiol. 2010, 8, 15–25. [Google Scholar] [CrossRef]
- D’Angelo-Picard, C.; Faure, D.; Penot, I.; Dessaux, Y. Diversity of N-acyl homoserine lactone-producing and -degrading bacteria in soil and tobacco rhizosphere. Environ. Microbiol. 2005, 7, 1796–1808. [Google Scholar] [CrossRef]
- Chemistry, A.; July, R.; Sepetember, A.; Shewanella, I. N-acyl homoserine lactone-producing or -degrading bacteria Isolated from the intestinal microbial flora of Ayu fish (Plecoglossus altivelis). ASM News 2005, 20, 264–268. [Google Scholar]
- Biswas, L.; Biswas, R.; Schlag, M.; Bertram, R.; Götz, F. Small-colony variant selection as a survival strategy for Staphylococcus aureus in the presence of Pseudomonas aeruginosa. Appl. Environ. Microbiol. 2009, 75, 6910–6912. [Google Scholar] [CrossRef]
- Kerr, J.; Taylor, G.; Rutman, A.; Høiby, N.; Cole, P.; Wilson, R. Pseudomonas aeruginosa pyocyanin and 1-hydroxyphenazine inhibit fungal growth. J. Clin. Pathol. 1999, 52, 385–387. [Google Scholar] [CrossRef]
- McClean, K.H.; Winson, M.K.; Fish, L.; Taylor, A.; Chhabra, S.R.; Camara, M.; Daykin, M.; Lamb, J.H.; Swift, S.; Bycroft, B.W.; et al. Quorum sensing and Chromobacterium violaceum: Exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiology 1997, 143, 3703–3711. [Google Scholar] [CrossRef]
- Morohoshi, T.; Kato, M.; Fukamachi, K.; Kato, N.; Ikeda, T. N-acylhomoserine lactone regulates violacein production in Chromobacterium violaceum type strain ATCC 12472. FEMS Microbiol. Lett. 2008, 279, 124–130. [Google Scholar] [CrossRef]
- Lyon, G.J.; Wright, J.S.; Muir, T.W.; Novick, R.P. Key determinants of receptor activation in the agr autoinducing peptides of Staphylococcus aureus. Biochemistry 2002, 41, 10095–10104. [Google Scholar] [CrossRef]
- Dufour, P.; Jarraud, S.; Vandenesch, F.; Greenland, T.; Novick, R.P.; Bes, M.; Etienne, J.; Lina, G. High genetic variability of the agr locus in Staphylococcus species. J. Bacteriol. 2002, 184, 1180–1186. [Google Scholar] [CrossRef]
- Jarraud, S.; Lyon, G.J.; Figueiredo, A.M.; Lina, G.; Vandenesch, F.; Etienne, J.; Muir, T.W.; Novick, R.P. Exfoliatin-producing strains define a fourth agr specificity group in Staphylococcus aureus. J. Bacteriol. 2000, 182, 6517–6522. [Google Scholar]
- Ji, G.; Beavis, R.; Novick, R.P. Bacterial interference caused by autoinducing peptide variants. Science 1997, 276, 2027–2030. [Google Scholar] [CrossRef]
- Hong, K.W.; Koh, C.L.; Sam, C.K.; Yin, W.F.; Chan, K.G. Quorum quenching revisited-from signal decays to signalling confusion. Sensors (Basel) 2012, 12, 4661–4696. [Google Scholar] [CrossRef]
- Czajkowski, R.; Jafra, S. Quenching of acyl-homoserine lactone-dependent quorum sensing by enzymatic disruption of signal molecules. Acta Biochim. Pol. 2009, 56, 1–16. [Google Scholar]
- Romero, M.; Martin-Cuadrado, A.B.; Otero, A. Determination of whether quorum quenching is a common activity in marine bacteria by analysis of cultivable bacteria and metagenomic sequences. Appl. Environ. Microbiol. 2012, 78, 6345–6348. [Google Scholar] [CrossRef]
- Khan, S.R.; Farrand, S.K. The BlcC (AttM) lactonase of Agrobacterium tumefaciens does not quench the quorum-sensing system that regulates Ti plasmid conjugative transfer. J. Bacteriol. 2009, 191, 1320–1329. [Google Scholar] [CrossRef]
- Flagan, S.; Ching, W.K.; Leadbetter, J.R. Arthrobacter strain VAI-A utilizes acyl-homoserine lactone inactivation products and stimulates quorum signal biodegradation by Variovorax paradoxus. Appl. Environ. Microbiol. 2003, 69, 909–916. [Google Scholar] [CrossRef]
- Park, S.J.; Park, S.Y.; Ryu, C.M.; Park, S.H.; Lee, J.K. The role of AiiA, a quorum-quenching enzyme from Bacillus thuringiensis, on the rhizosphere competence. J. Microbiol. Biotechnol. 2008, 18, 1518–1521. [Google Scholar]
- Romero, M.; Martin-Cuadrado, A.B.; Roca-Rivada, A.; Cabello, A.M.; Otero, A. Quorum quenching in cultivable bacteria from dense marine coastal microbial communities. FEMS Microbiol. Ecol. 2011, 75, 205–217. [Google Scholar] [CrossRef]
- Torres, M.; Romero, M.; Prado, S.; Dubert, J.; Tahrioui, A.; Otero, A.; Llamas, I. N-acylhomoserine lactone-degrading bacteria isolated from hatchery bivalve larval cultures. Microbiol. Res. 2013, 168, 547–554. [Google Scholar] [CrossRef]
- Tait, K.; Williamson, H.; Atkinson, S.; Williams, P.; Camara, M.; Joint, I. Turnover of quorum sensing signal molecules modulates cross-kingdom signalling. Environ. Microbiol. 2009, 11, 1792–1802. [Google Scholar] [CrossRef]
- Defoirdt, T.; Thanh, L.D.; Van Delsen, B.; De Schryver, P.; Sorgeloos, P.; Boon, N.; Bossier, P. N-acylhomoserine lactone-degrading Bacillus strains isolated from aquaculture animals. Aquaculture 2011, 311, 258–260. [Google Scholar] [CrossRef]
- Romero, M.; Avendaño-Herrera, R.; Magariños, B.; Cámara, M.; Otero, A. Acylhomoserine lactone production and degradation by the fish pathogen Tenacibaculum maritimum, a member of the Cytophaga–Flavobacterium–Bacteroides (CFB) group. FEMS Microbiol. Lett. 2010, 304, 131–139. [Google Scholar] [CrossRef]
- Huang, W.; Lin, Y.; Yi, S.; Liu, P.; Shen, J.; Shao, Z.; Liu, Z. QsdH, a novel AHL lactonase in the RND-type inner membrane of marine Pseudoalteromonas byunsanensis strain 1A01261. PLoS One 2012, 7, e46587. [Google Scholar]
- Swem, L.R.; Swem, D.L.; O’Loughlin, C.T.; Gatmaitan, R.; Zhao, B.X.; Ulrich, S.M.; Bassler, B.L. A quorum-sensing antagonist targets both membrane-bound and cytoplasmic receptors and controls bacterial pathogenicity. Mol. Cell 2009, 35, 143–153. [Google Scholar] [CrossRef]
- O’Loughlin, C.T.; Miller, L.C.; Siryaporn, A.; Drescher, K.; Semmelhack, M.F.; Bassler, B.L. A quorum-sensing inhibitor blocks Pseudomonas aeruginosa virulence and biofilm formation. Proc. Natl. Acad. Sci. USA 2013, 110, 17981–17986. [Google Scholar]
- Whitehead, N.A.; Welch, M.; Salmond, G.P. Silencing the majority. Nat. Biotechnol. 2001, 19, 735–736. [Google Scholar] [CrossRef]
- Defoirdt, T.; Boon, N.; Bossier, P. Can bacteria evolve resistance to quorum sensing disruption? PLoS Pathog. 2010, 6, e1000989. [Google Scholar] [CrossRef] [Green Version]
- Maeda, T.; Garcia-Contreras, R.; Pu, M.; Sheng, L.; Garcia, L.R.; Tomas, M.; Wood, T.K. Quorum quenching quandary: Resistance to antivirulence compounds. ISME J. 2012, 6, 493–501. [Google Scholar] [CrossRef]
- Garcia-Contreras, R.; Maeda, T.; Wood, T.K. Resistance to quorum-quenching compounds. Appl. Environ. Microbiol. 2013, 79, 6840–6846. [Google Scholar] [CrossRef]
- Fernandes, R.; Roy, V.; Wu, H.-C.; Bentley, W.E. Engineered biological nanofactories trigger quorum sensing response in targeted bacteria. Nat. Nano 2010, 5, 213–217. [Google Scholar] [CrossRef]
- Komnatnyy, V.V.; Chiang, W.C.; Tolker-Nielsen, T.; Givskov, M.; Nielsen, T.E. Bacteria-triggered release of antimicrobial agents. Angew. Chem. Int. Ed. Engl. 2014, 53, 439–441. [Google Scholar] [CrossRef]
- Park, K.; Kwon, I.C.; Park, K. Oral protein delivery: Current status and future prospect. React. Funct. Polym. 2011, 71, 280–287. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Tang, K.; Zhang, X.-H. Quorum Quenching Agents: Resources for Antivirulence Therapy. Mar. Drugs 2014, 12, 3245-3282. https://doi.org/10.3390/md12063245
AMA Style
Tang K, Zhang X-H. Quorum Quenching Agents: Resources for Antivirulence Therapy. Marine Drugs. 2014; 12(6):3245-3282. https://doi.org/10.3390/md12063245
Chicago/Turabian StyleTang, Kaihao, and Xiao-Hua Zhang. 2014. "Quorum Quenching Agents: Resources for Antivirulence Therapy" Marine Drugs 12, no. 6: 3245-3282. https://doi.org/10.3390/md12063245