Effects of Long Chain Fatty Acid Synthesis and Associated Gene Expression in Microalga Tetraselmis sp.

Abstract
:
1. Introduction

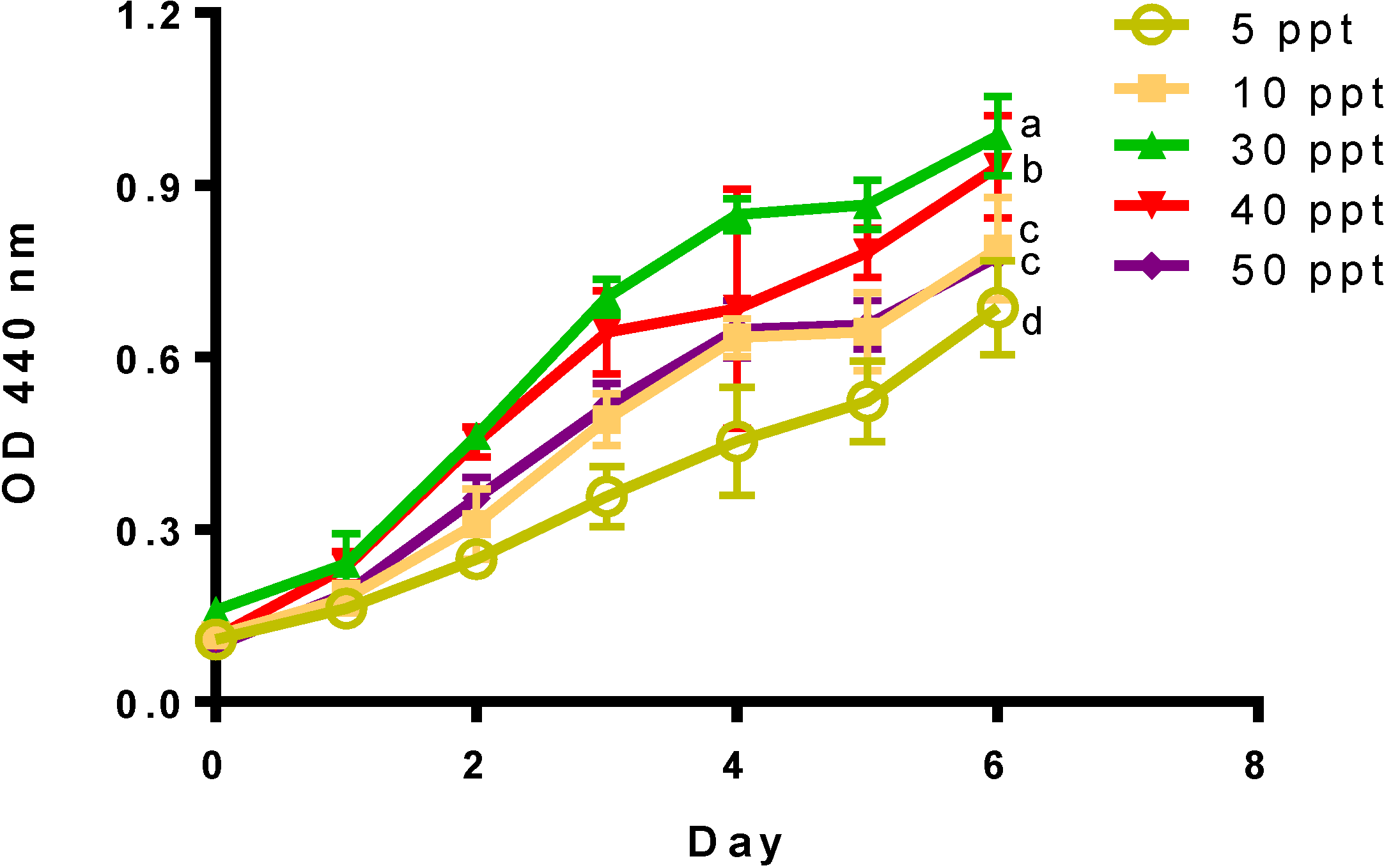
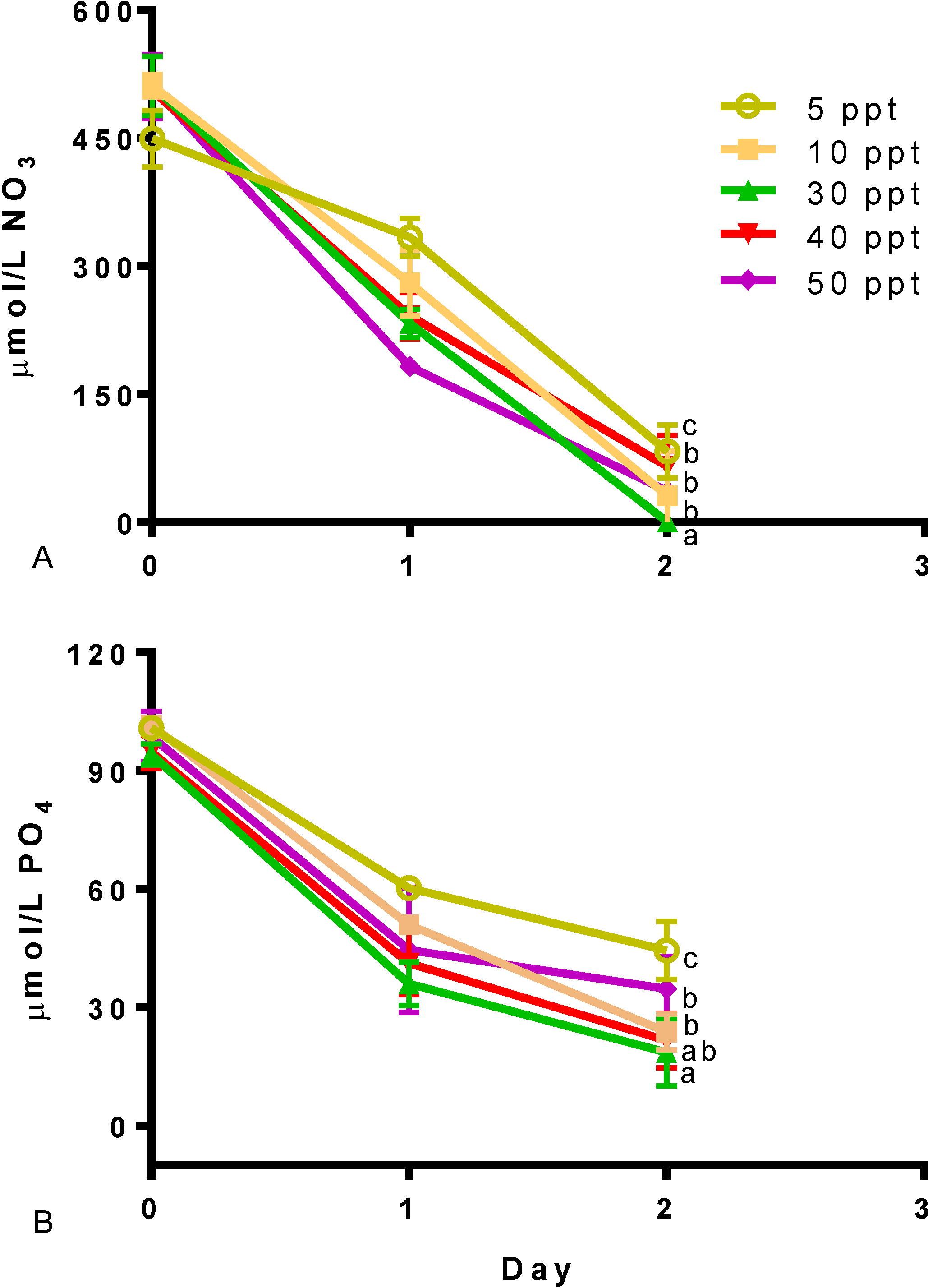
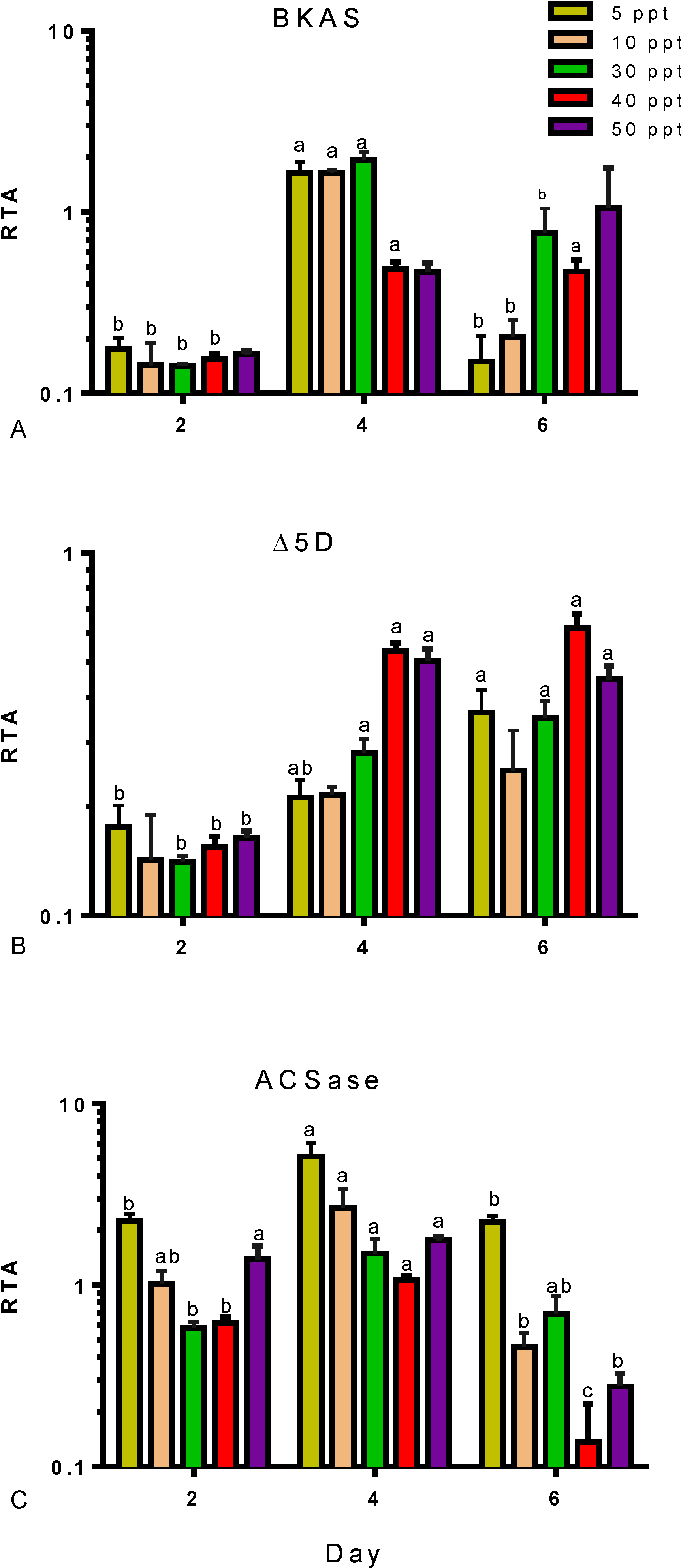
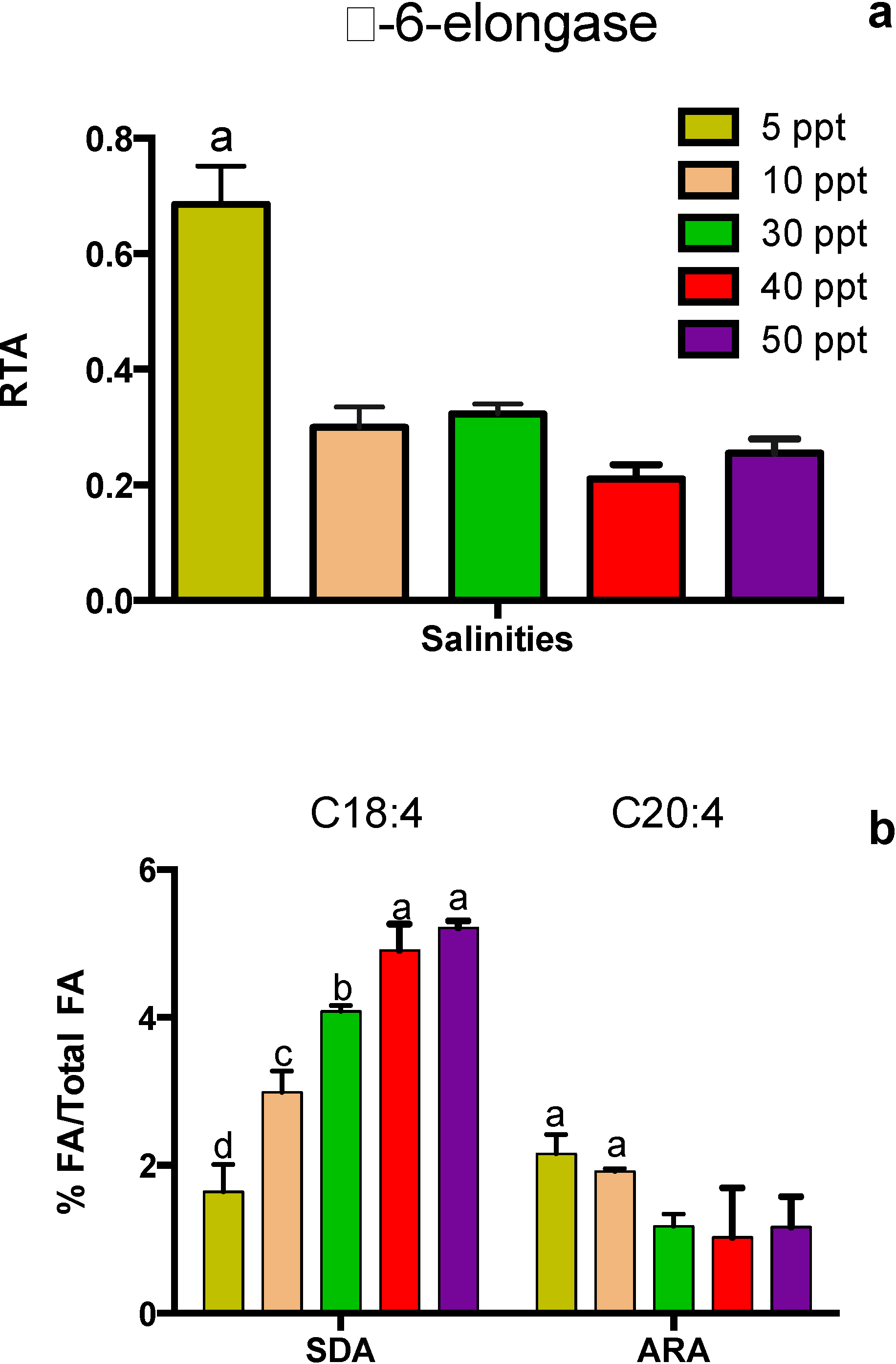
2. Results
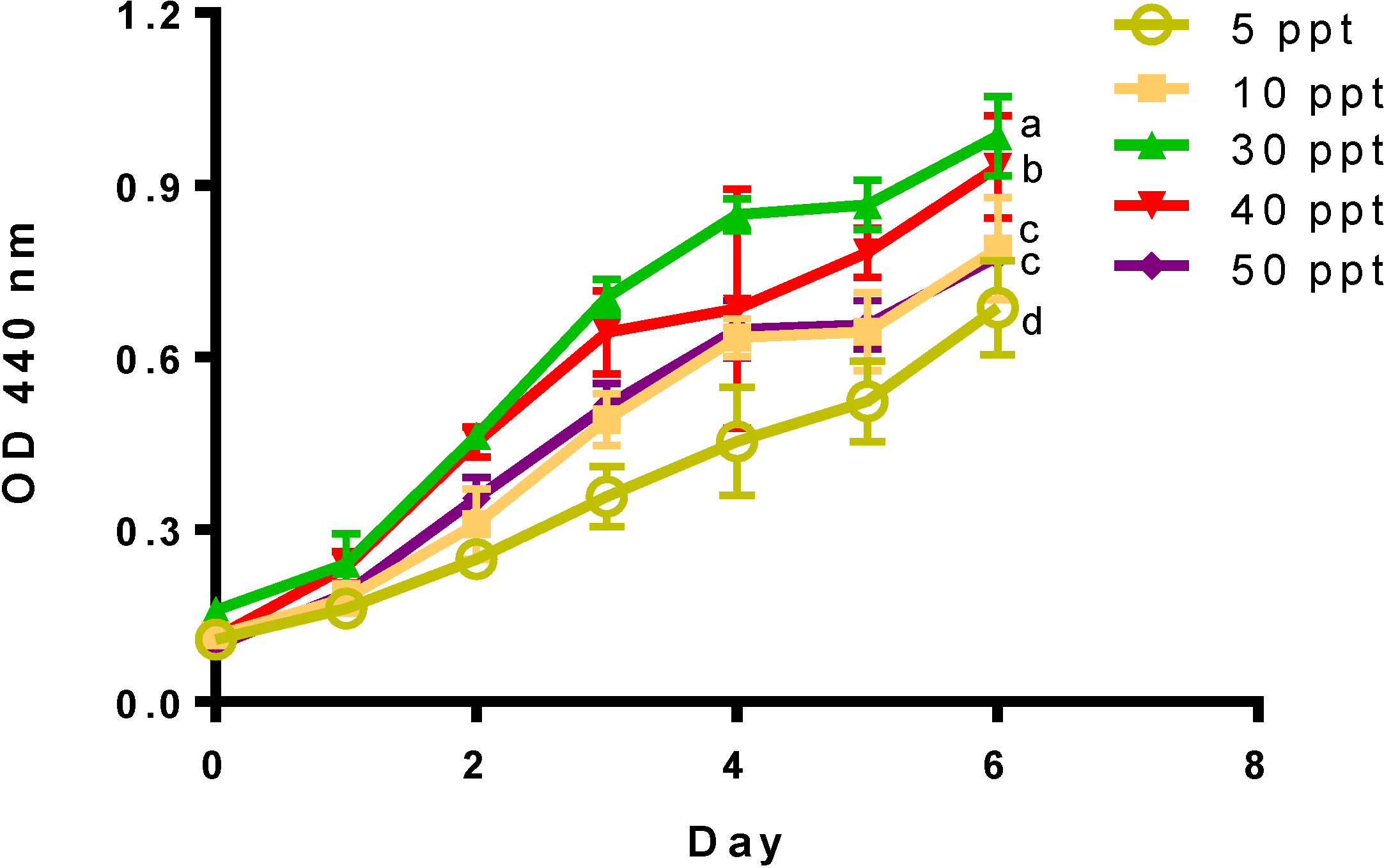

{kind=link}
{kind=link}
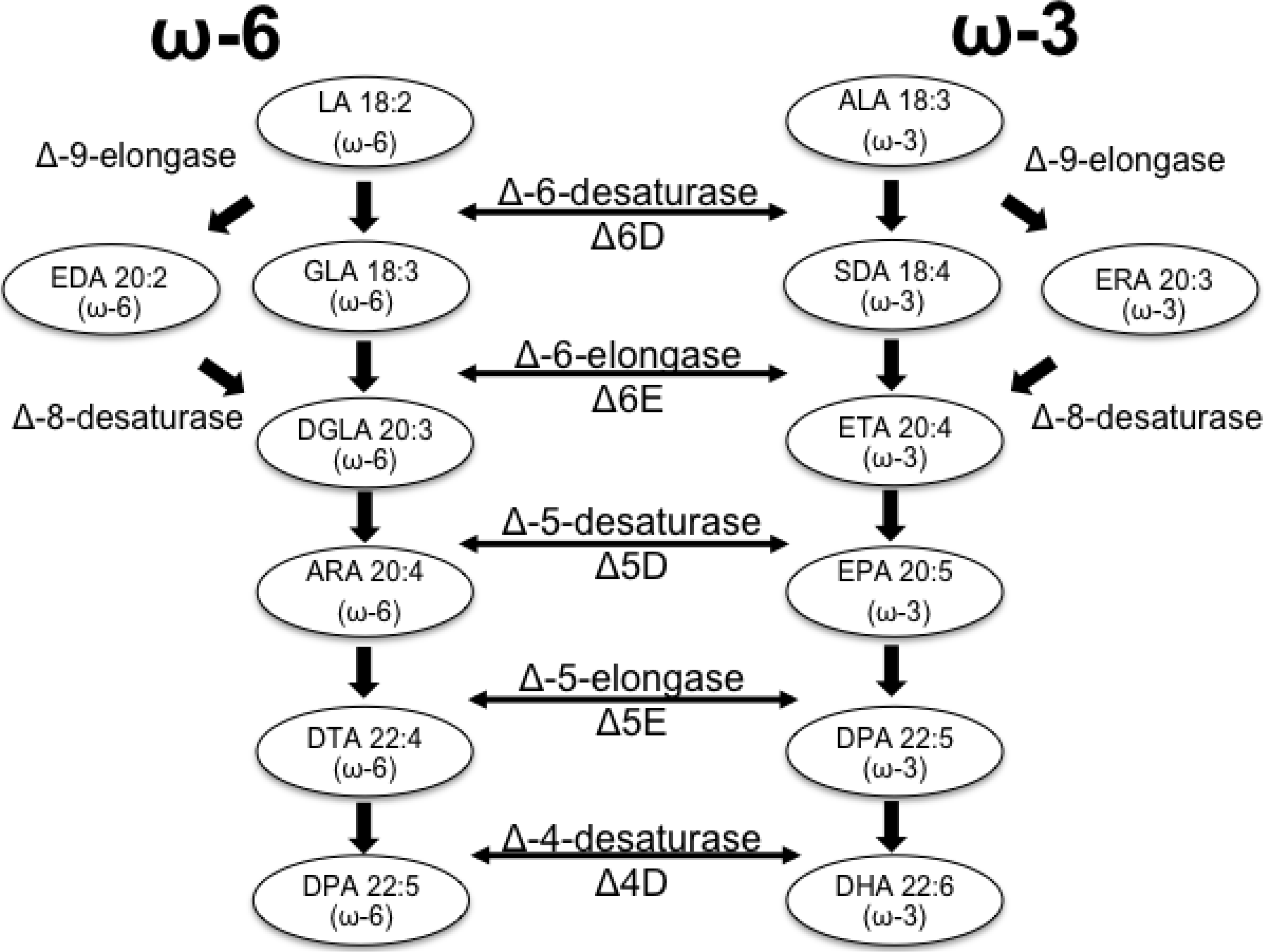{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primers |
|---|---|
| Beta-Keto acyl synthase (BKAS) | 5′-CAGGCCTTCGAGCATTTCTG-3′ 3′-GCGTCATATCAGGCGACAGC-5′ |
| Delta-5-desaturase (Δ5D) | 5′-TGGACGTTGGACATTGTAGGC-3′ 3′-CATTGTCATGCAGATTTGTGTACG-5′ |
| Delta-6-elongase (Δ6E) | 5′-CACCTACTACCTGCTTGCTGCC-3′ 3′-CTGGAACATTGTCAGGTAATGCC-5′ |
| Acyl-CoA-synthase (ACSase) | 5′-CACGTTGCTGTGCTTAATCTGC-3′ 3′-CGAGTGCAACCCTGAGGATATG-5′ |
| Delta-5-elongase (Δ5E) | 5′-TGAGGAAATGGTGCCAGCAG-3′ 3′-ACAAGTTCATCGAGTACCTCGACAC-5′ |
| Glycerol-3-phosphate dehydrogenase (D3PDH) | 5′-TCGTACCGCATCCACAAAGG-3′ 3′-GCTAAGGTGAAAGACAACGAGTCC-5′ |
| Glucose-6-phosphate isomerase (G6Pi) | 5′-GGGACAGCAGGTTATTGTGGAC-3′ 3′-TGCGCACCTTATCGGAGAAG-5′ |
| Sodium ATPase (PyKPA) | 5′-AAGGAAGCTGCGGATATGATTCTC-3′ 3′-TCAAGTTGTCAAAAATCAGACGACC-5′ |
| Phosphate transporter (PHO) | 5′-GACTTGGCACCCTTGAAGATAATG-3′ 3′-CTTACGCTCGCTCTTGGTGG-5′ |
| 3-ketoacyl-ACP reductase (KAR) | 5′-CGGAGGAGATGTTAATGATGCG-3′ 3′-ATCAACCTCACCGGCGTCTT-5′ |
| Delta-8-desaturase (Δ8D) | 5′-GTCCGTAAAGGCTCCACTTCG-3′ 3′-GTATTTGACAAGACCACGCAGTTG-5′ |
| Enoyl-ACP reductase (ENR) | 5′-CTCCTTGACCTCAGTTGGGACA-3′ 3′-CTCAAACGGGTCCTTAATGGAGT-5′ |
| Phosphatidic acid phosphatase (PP) | 5′-TGTGGTCGGAGATCACATACGATA-3′ 3′-CAGTAGAGCGAGAACGACACCAG-5′ |
| Delta-9-desaturase (Δ9D) | 5′-GATATGAAAGCGTATGCCGAG-3′ 3′-GTAGCTCTAGCCGCCCCCTT-5′ |
| Diacyl glycerol acyl transferase (DGAT) | 5′-ATCAGAGGAACCTGTCCCATCA-3′ 3′-CTGCCATTTTTCACGAGCTAATG-5′ |
| Beta-actin | 5′-GCCTCAGAATCCCAAGACCAA-3′ 3′-GGCCTGGATCTGAACGTACATG-5′ |
| Salinity (ppt) | Growth Rate (μ) | Doubling Time (dt) |
|---|---|---|
| 5 | 0.414 ± 0.049 | 1.689 ± 0.207 |
| 10 | 0.487 ± 0.087 | 1.457 ± 0.287 |
| 30 | 0.532 ± 0.016 | 1.303 ± 0.038 |
| 40 | 0.695 ± 0.064 * | 1.003 ± 0.088 |
| 50 | 0.644 ± 0.063 * | 1.083 ± 0.105 |



| Salinity | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fatty Acids | 5 ppt | 10 ppt | 30 ppt | 40 ppt | 50 ppt | ||||||||||
| 2 | 4 | 6 | 2 | 4 | 6 | 2 | 4 | 6 | 2 | 4 | 6 | 2 | 4 | 6 | |
| C12:0 | 0.19 ± 0.07 | 0.23 ± 0.39 | 0.26 ±0.40 | 0.11 ± 0.05 | 0.11 ± 0.09 | 0.08 ± 0.09 | 0.15 ± 0.10 | - | 0.01 ± 0.01 | 0.06 ± 0.02 | - | 0.04 ± 0.07 | 0.09 ± 0.06 | 0.22 ± 0.21 | 0.05 ± 0.06 |
| C14:0 | 0.38 ± 0.02 | 0.15 ± 0.27 | 0.32 ± 0.30 | 0.30 ± 0.01 | 0.23 ± 0.01 | 0.19 ± 0.09 | 0.23 ± 0.17 | 0.14 ± 0.04 | 0.17 ± 0.05 | 0.19 ± 0.06 | 0.11 ± 0.1 | 0.08 ± 0.07 | 0.24 ± 0.07 | 0.25 ± 0.22 | 0.17 ± 0.15 |
| C14:1 | 1.22 ± 0.01 | 0.76 ± 0.10 | 0.72 ± 0.09 | 1.45 ± 0.05 | 1.11 ± 0.10 | 0.77± 0.16 | 1.67 ± 0.08 | 1.27 ±0.23 | 1.01 ± 0.16 | 1.66 ± 0.06 | 1.00 ± 0.48 | 0.65 ± 0.56 | 1.69 ± 0.05 | 1.27 ± 0.38 | 0.98 ± 0.28 |
| C16:0 | 19.38 ± 0.01 | 21.22 ± 1.95 | 22.79 ± 1.36 | 18.14 ± 0.10 | 21.81 ± 0.58 | 22.73 ± 0.52 | 19.37 ± 0.90 | 22.15 ± 1.24 | 23.29 ± 1.00 | 18.38 ± 0.30 | 19.44 ± 1.88 | 20.99 ± 1.24 | 17.76 ± 1.21 | 18.70 ± 0.20 | 20.51 ± 1.83 |
| C16:1 | 1.56 ± 0.11 | 1.31 ± 0.13 | 2.20 ± 1.29 | 3.63 ± 0.27 | 1.90 ± 0.07 | 1.627 ± 0.22 | 4.71 ± 0.76 | 2.36 ± 0.30 | 1.96 ± 0.32 | 4.26 ± 0.80 | 2.06 ± 0.80 | 1.72 ± 0.58 | 4.64 ± 0.55 | 2.72 ± 0.60 | 2.43 ±0.35 |
| C16:3 | 6.51 ± 0.55 | 6.80 ± 0.77 | 5.89 ± 0.76 | 5.69 ± 0.18 | 5.58 ± 0.12 | 4.73 ± 0.33 | 4.33 ± 0.37 | 4.25 ± 0.09 | 3.44 ± 0.18 | 4.24 ± 0.25 | 4.70 ± 0.16 | 3.70 ± 0.12 | 3.94 ± 0.16 | 5.38 ± 0.14 | 4.68 ± 0.64 |
| C16:4 | 16.54 ± 0.53 | 17.37 ± 4.48 | 13.80 ± 3.49 | 17.82 ± 0.01 | 15.02 ± 0.50 | 16.42 ± 1.61 | 19.01 ± 1.51 | 17.51 ± 1.36 | 17.16 ± 1.57 | 19.37 ± 0.17 | 22.26 ± 9.65 | 20.66 ± 4.87 | 19.47 ± 0.98 | 18.81 ± 4.11 | 18.37 ± 5.18 |
| C18:0 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| C18:1 | 10.29 ± 0.38 | 11.94 ± 3.63 | 16.25 ± 2.80 | 7.54 ± 0.12 | 11.87 ± 0.78 | 13.30 ± 0.65 | 7.15 ± 0.79 | 10.49 ± 0.38 | 12.11 ± 0.32 | 6.36± 0.18 | 7.92 ± 3.44 | 7.12 ± 6.18 | 6.22 ± 0.36 | 8.65 ± 1.71 | 10.50 ± 1.72 |
| C18:2 | 13.86 ± 1.40 | 15.15 ±0.88 | 14.39 ± 0.62 | 13.07 ± 0.27 | 15.55 ± 0.18 | 14.34 ± 0.19 | 11.37 ± 0.43 | 13.43 ± 0.46 | 12.81 ± 0.57 | 10.90 ± 0.45 | 13.20 ± 1.37 | 13.39 ± 0.34 | 9.81 ± 0.80 | 13.15 ± 0.16 | 13.01 ± 1.10 |
| C18:3 (GLA) | 0.61 ± 0.03 | 0.39 ± 0.10 | 0.79 ± 0.09 | 0.51 ± 0.01 | 0.631±0.040 | 0.81 ± 0.08 | 0.30 ± 0.18 | 0.47 ± 0.05 | 0.74 ± 0.06 | 0.32 ± 0.05 | 0.39 ± 0.34 | 0.74 ± 0.17 | 0.35 ± 0.05 | 0.49 ± 0.33 | 0.75 ± 0.14 |
| C18:3 (ALA) | 17.39 ± 1.31 | 14.35 ± 1.37 | 11.25 ± 0.79 | 18.76 ± 0.12 | 14.219±0.572 | 12.44 ± 0.37 | 18.77 ± 0.10 | 15.15 ± 0.52 | 13.99 ± 0.40 | 20.53 ± 0.30 | 16.19 ± 0.71 | 16.00 ± 2.21 | 20.51 ± 0.16 | 16.34 ± 0.60 | 14.69 ± 0.44 |
| C18:4 | 2.26 ± 0.31 | 1.64 ± 0.37 | 1.41 ± 0.21 | 4.04 ± 0.18 | 2.99 ± 0.29 | 2.58 ± 0.21 | 4.84 ± 0.17 | 4.08 ± 0.08 | 3.62 ± 0.05 | 5.41 ± 0.33 | 4.90 ± 0.35 | 4.61 ± 0.84 | 5.95 ± 0.80 | 5.21 ± 0.09 | 4.33 ± 0.54 |
| C20:0 | - | - | - | 0.46 ± 0.65 | - | - | - | - | - | - | - | - | - | - | - |
| C20:1 | 1.67 ± 0.18 | 1.12 ± 0.21 | 1.38 ± 0.27 | 1.30 ± 0.03 | 1.21 ± 0.03 | 1.12 ±0.12 | 2.02 ± 0.26 | 1.64 ± 0.12 | 1.50 ± 0.05 | 2.08 ± 0.26 | 1.36 ± 0.60 | 1.12 ± 0.52 | 2.55 ± 0.36 | 1.90 ± 0.24 | 1.59 ± 0.10 |
| C20:2 | 0.19 ± 0.26 | 0.33 ± 0.07 | 0.27 ± 0.24 | 0.18 ± 0.25 | 0.24 ± 0.03 | 0.31 ± 0.05 | 0.15 ± 0.14 | 0.29 ± 0.03 | 0.21 ± 0.18 | 0.21 ± 0.06 | 0.15 ± 0.13 | 0.18 ± 0.09 | 0.15 ± 0.14 | 0.33 ± 0.10 | 0.29 ± 0.02 |
| C20:3 | 0.14 ± 0.09 | - | - | 0.10 ± 0.02 | - | - | 0.06 ± 0.10 | - | - | - | - | - | 0.05 ± 0.04 | 0.03 ± 0.04 | - |
| C20:4 | 2.12 ± 0.16 | 2.15 ± 0.26 | 2.75 ± 0.24 | 1.63 ± 0.02 | 1.92 ± 0.04 | 2.20 ± 0.06 | 0.94 ± 0.30 | 1.18 ± 0.16 | 1.65 ± 0.12 | 1.06 ± 0.04 | 1.03 ± 0.67 | 1.73 ± 0.10 | 1.08 ± 0.07 | 1.17 ± 0.41 | 1.46 ± 0.29 |
| C20:5 | 5.04 ± 0.11 | 5.06 ± 0.36 | 5.49 ± 0.32 | 4.91 ± 0.04 | 5.61 ± 0.10 | 6.13 ± 0.09 | 4.02 ± 0.43 | 5.01 ± 0.69 | 6.10 ± 0.49 | 4.44 ± 0.15 | 5.12 ± 0.91 | 7.18 ± 1.46 | 4.72 ± 0.44 | 5.29 ± 0.38 | 5.96 ± 0.30 |
| SFA | 19.96 ± 0.06 | 21.60 ± 2.59 | 23.37 ± 2.03 | 19.01 ± 0.79 | 22.15 ± 0.48 | 23.00 ± 0.53 | 19.76 ± 0.70 | 22.29 ± 1.26 | 23.46 ± 1.01 | 16.64 ± 0.24 | 19.55 ± 1.98 | 21.11 ± 1.34 | 18.09 ± 1.12 | 19.17 ± 0.36 | 20.74 ± 2.03 |
| MUFA | 14.74 ± 0.31 | 15.12 ± 3.98 | 20.55 ± 3.62 | 13.93 ± 0.23 | 16.10 ± 0.66 | 16.82 ± 0.96 | 15.56 ± 0.44 | 15.77 ± 0.61 | 16.58 ± 0.71 | 14.35 ± 0.56 | 12.34 ± 5.32 | 10.60 ± 7.83 | 15.11 ± 0.57 | 14.54 ± 2.88 | 15.50 ± 2.38 |
| PUFA | 64.66 ± 0.32 | 63.25 ± 6.52 | 56.04 ± 5.09 | 66.71 ± 0.07 | 61.76 ± 1.10 | 59.97 ± 1.48 | 63.68 ± 0.24 | 61.38 ± 2.00 | 59.74 ± 1.53 | 66.49 ± 0.41 | 67.95 ± 7.44 | 68.18 ± 9.24 | 66.02 ± 1.35 | 66.21 ± 3.18 | 63.55 ± 4.46 |
| ω-3 | 26.81 ± 1.35 | 23.21 ± 1.66 | 20.89 ± 0.79 | 29.34 ± 0.25 | 24.73 ± 0.87 | 23.35 ± 0.65 | 28.57 ± 0.79 | 25.43 ± 1.14 | 25.37 ± 0.73 | 31.44 ± 0.22 | 27.24 ± 0.53 | 29.51 ± 4.40 | 32.25 ± 1.13 | 28.02 ± 0.75 | 26.44 ± 0.08 |
 ) 5 ppt, Day 2; (
) 5 ppt, Day 2; (  ) 5 ppt, Day 4; (
) 5 ppt, Day 4; (  ) 5 ppt, Day 6; (
) 5 ppt, Day 6; (  ) 10 ppt, Day 2; (
) 10 ppt, Day 2; (  ) 10 ppt, Day 4; (
) 10 ppt, Day 4; (  ) 10 ppt, Day 6; (
) 10 ppt, Day 6; (  ) 30 ppt, Day 2; (
) 30 ppt, Day 2; (  ) 30 ppt, Day 4; (
) 30 ppt, Day 4; (  ) 30 ppt, Day 6; (
) 30 ppt, Day 6; (  ) 40 ppt, Day 2; (
) 40 ppt, Day 2; (  ) 40 ppt, Day 4; (
) 40 ppt, Day 4; (  ) 40 ppt, Day 6; (
) 40 ppt, Day 6; (  ) 50 ppt, Day 2; (
) 50 ppt, Day 2; (  ) 50 ppt, Day 4, (
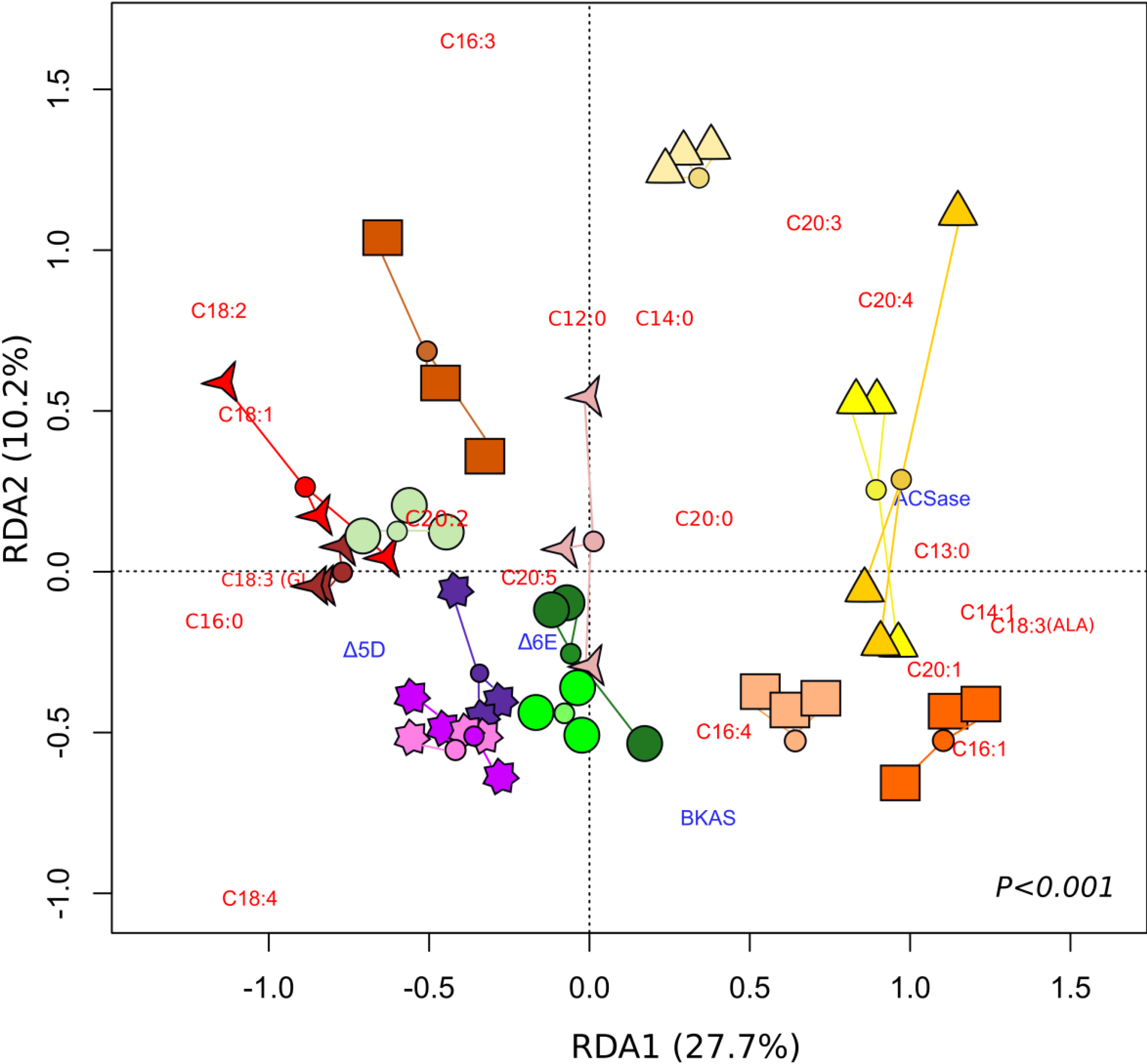
) 50 ppt, Day 4, (  ) 50 ppt, Day 6. The small coloured circles represent the centroid of the treatment replicates.
) 5 ppt, Day 2; ( ) 5 ppt, Day 4; ( ) 5 ppt, Day 6; ( ) 10 ppt, Day 2; ( ) 10 ppt, Day 4; ( ) 10 ppt, Day 6; ( ) 30 ppt, Day 2; ( ) 30 ppt, Day 4; ( ) 30 ppt, Day 6; ( ) 40 ppt, Day 2; ( ) 40 ppt, Day 4; ( ) 40 ppt, Day 6; ( ) 50 ppt, Day 2; ( ) 50 ppt, Day 4, ( ) 50 ppt, Day 6. The small coloured circles represent the centroid of the treatment replicates.
) 50 ppt, Day 6. The small coloured circles represent the centroid of the treatment replicates.
) 5 ppt, Day 2; ( ) 5 ppt, Day 4; ( ) 5 ppt, Day 6; ( ) 10 ppt, Day 2; ( ) 10 ppt, Day 4; ( ) 10 ppt, Day 6; ( ) 30 ppt, Day 2; ( ) 30 ppt, Day 4; ( ) 30 ppt, Day 6; ( ) 40 ppt, Day 2; ( ) 40 ppt, Day 4; ( ) 40 ppt, Day 6; ( ) 50 ppt, Day 2; ( ) 50 ppt, Day 4, ( ) 50 ppt, Day 6. The small coloured circles represent the centroid of the treatment replicates.
3. Discussion
4. Methods
4.1. Algae Culture and Cultivation Conditions
4.2. Culture Media Nutrient Analysis
4.3. Fatty Acid Analysis
4.4. Total RNA Extraction and cDNA Synthesis
4.5. Real-Time Quantitative Reverse Transcriptase PCR
4.6. Statistical Analyses
5. Conclusion
Supplementary Files
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lee, J.H.; O’Keefe, J.H.; Lavie, C.J.; Harris, W.S. Omega-3 fatty acids: Cardiovascular benefits, sources and sustainability. Nat. Rev. Cardiol. 2009, 6, 753–758. [Google Scholar] [CrossRef]
- Simopoulos, A.P.; Bazán, N.G.; Karger, S. Omega-3 Fatty Acids, the Brain and Retina; Karger: Basel, Switzerland, 2009. [Google Scholar]
- Damude, H.G.; Kinney, A.J. Enhancing plant seed oils for human nutrition. Plant Physiol. 2008, 147, 962–968. [Google Scholar] [CrossRef]
- Ruiz-López, N.; Haslam, R.P.; Venegas-Calerón, M.; Li, T.; Bauer, J.; Napier, J.A.; Sayanova, O. Enhancing the accumulation of omega-3 long chain polyunsaturated fatty acids in transgenic Arabidopsis thaliana via iterative metabolic engineering and genetic crossing. Transgenic Res. 2012, 21, 1233–1243. [Google Scholar]
- Adarme-Vega, T.C.; Lim, D.; Timmins, M.; Vernen, F.; Li, Y.; Schenk, P.M. Microalgal biofactories: A promising approach towards sustainable omega-3 fatty acid production. Microb. Cell Fact. 2012, 11, 96. [Google Scholar] [CrossRef]
- Rubio-Rodríguez, N.; Beltrán, S.; Jaime, I.; de Diego, S.M.; Sanz, M.T.; Carballido, J.R. Production of omega-3 polyunsaturated fatty acid concentrates: A review. Innov. Food Sci. Emerg. Technol. 2010, 11, 1–12. [Google Scholar] [CrossRef]
- Schenk, P.M.; Thomas-Hall, S.R.; Stephens, E.; Marx, U.C.; Mussgnug, J.H.; Posten, C.; Kruse, O.; Hankamer, B. Second generation biofuels: High-efficiency microalgae for biodiesel production. Bioenerg. Res. 2008, 1, 20–43. [Google Scholar] [CrossRef]
- Fabregas, J.; Abalde, J.; Herrero, C.; Cabezas, B.; Veiga, M. Growth of the marine microalga Tetraselmis suecica in batch cultures with different salinities and nutrient concentrations. Aquaculture 1984, 42, 207–215. [Google Scholar] [CrossRef]
- Roessler, P.G. Environmental control of glycerolipid metabolism in microalgae: Commercial implications and future research directions. J. Phycol. 1990, 26, 393–399. [Google Scholar]
- de Castro Araujo, S.; Garcia, V.M.T. Growth and biochemical composition of the diatom Chaetoceros cf. wighamii Brightwell under different temperature, salinity and carbon dioxide levels. I. Protein, carbohydrates and lipids. Aquaculture 2005, 246, 405–412. [Google Scholar] [CrossRef]
- Brown, M.R.; Jeffrey, S.W.; Garland, C.D. Nutritional Aspects of Microalgae Used in Mariculture: A literature Review; CSIRO: Hobart, Austrilia, 1989. [Google Scholar]
- Richmond, A. CRC Hhandbook of Microalgal Mass Culture; Richmond, A., Ed.; CRC Press: Boca Raton, FL, USA, 1986; pp. 69–99. [Google Scholar]
- Hu, H.; Gao, K. Response of growth and fatty acid compositions of Nannochloropsis sp. to environmental factors under elevated CO2 concentration. Biotechnol. Lett. 2006, 28, 987–992. [Google Scholar] [CrossRef]
- Lee, Y.K.; Tan, H.M.; Low, C.S. Effect of salinity of medium on cellular fatty acid composition of marine alga Porphyridium cruentum (Rhodophyceae). J. Appl. Phycol. 1989, 1, 19–23. [Google Scholar] [CrossRef]
- Hu, Q.; Sommerfeld, M.; Jarvis, E.; Ghirardi, M.; Posewitz, M.; Seibert, M.; Darzins, A. Microalgal triacylglycerols as feedstocks for biofuel production: Perspectives and advances. Plant J. 2008, 54, 621–639. [Google Scholar] [CrossRef]
- Nuutila, A.M.; Aura, A.M.; Kiesvaara, M.; Kauppinen, V. The effect of salinity, nitrate concentration, pH and temperature on eicosapentaenoic acid (EPA) production by the red unicellular alga Porphyridium purpureum. J. Biotechnol. 1997, 55, 55–63. [Google Scholar] [CrossRef]
- Yongmanitchai, W.; Ward, O.P. Growth of and omega-3 fatty acid production by Phaeodactylum tricornutum under different culture conditions. Appl. Environ. Microbiol. 1991, 57, 419–425. [Google Scholar]
- Pal, D.; Khozin-Goldberg, I.; Cohen, Z.; Boussiba, S. The effect of light, salinity, and nitrogen availability on lipid production by Nannochloropsis sp. Appl. Microbiol. Biotechnol. 2011, 90, 1429–1441. [Google Scholar] [CrossRef]
- Takagi, M. Effect of salt concentration on intracellular accumulation of lipids and triacylglyceride in marine microalgae Dunaliella cells. J. Biosci. Bioeng. 2006, 101, 223–226. [Google Scholar]
- Sukenik, A. Ecophysiological considerations in the optimization of eicosapentaenoic acid production by Nannochloropsis sp. (Eustigmatophyceae). Bioresour. Technol. 1991, 35, 263–269. [Google Scholar] [CrossRef]
- Venegas-Calerón, M.; Sayanova, O.; Napier, J.A. An alternative to fish oils: Metabolic engineering of oil-seed crops to produce omega-3 long chain polyunsaturated fatty acids. Progr. Lipid Res. 2010, 49, 108–119. [Google Scholar] [CrossRef]
- Meesapyodsuk, D.; Qiu, X. The front-end desaturase: Structure, function, evolution and biotechnological use. Lipids 2012, 47, 227–237. [Google Scholar] [CrossRef]
- Khozin-Goldberg, I.; Cohen, Z. Unraveling algal lipid metabolism: Recent advances in gene identification. Biochimie 2011, 93, 91–100. [Google Scholar] [CrossRef]
- Strizh, I.; Popova, L.; Balnokin, Y.V. Physiological aspects of adaptation of the marine microalga Tetraselmis (Platymonas) viridis to various medium salinity. Russ. J. Plant Physiol. 2004, 51, 176–182. [Google Scholar] [CrossRef]
- Pagis, L.Y.; Popova, L.G.; Andreev, I.M.; Balnokin, Y.V. Comparative characterization of the two primary pumps, H+ATPase and Na+ATPase, in the plasma membrane of the marine alga Tetraselmis viridis. Physiol. Plant. 2003, 118, 514–522. [Google Scholar] [CrossRef]
- Azachi, M.; Sadka, A.; Fisher, M.; Goldshlag, P.; Gokhman, I.; Zamir, A. Salt induction of fatty acid elongase and membrane lipid modifications in the extreme halotolerant alga Dunaliella salina. Plant Physiol. 2002, 129, 1320–1329. [Google Scholar] [CrossRef]
- Iwamoto, K.; Shiraiwa, Y. Salt-regulated mannitol metabolism in algae. Mar. Biotechnol. 2005, 7, 407–415. [Google Scholar] [CrossRef]
- Li, S.H.; Xia, B.B.; Zhang, C.; Cao, J.; Bai, L.H. Cloning and characterization of a phosphate transporter gene in Dunaliella salina. J. Basic Microbiol. 2012, 52, 429–436. [Google Scholar] [CrossRef]
- Arisz, S.A.; Munnik, T. The salt stress-induced LPA response in Chlamydomonas is produced via PLA2 hydrolysis of DGK-generated phosphatidic acid. J. Lipid Res. 2011, 52, 2012–2020. [Google Scholar] [CrossRef]
- Petrie, J.R.; Shrestha, P.; Mansour, M.P.; Nichols, P.D.; Liu, Q.; Singh, S.P. Metabolic engineering of omega-3 long-chain polyunsaturated fatty acids in plants using an acyl-coa δ6-desaturase with ω3-preference from the marine microalga Micromonas pusilla. Metab. Eng. 2010, 12, 233–240. [Google Scholar] [CrossRef]
- Qiu, X.; Hong, H.; MacKenzie, S.L. Identification of a δ4 fatty acid desaturase from thraustochytrium sp. Involved in the biosynthesis of docosahexanoic acid by heterologous expression in Saccharomyces cerevisiae and Brassica juncea. J. Biol. Chem. 2001, 276, 31561–31566. [Google Scholar] [CrossRef]
- Cheng, B.; Wu, G.; Vrinten, P.; Falk, K.; Bauer, J.; Qiu, X. Towards the production of high levels of eicosapentaenoic acid in transgenic plants: The effects of different host species, genes and promoters. Transgenic Res. 2010, 19, 221–229. [Google Scholar] [CrossRef]
- Serdar, S.; Lök, A.; Acarli, S.; Köse, A. The effect of two different culture media and five different salinities on growth of Tetraselmis suecica. Rapp. Comm. Int. Mer. Médit. 2007, 38, 394. [Google Scholar]
- Kumar, M.; Bijo, A.; Baghel, R.S.; Reddy, C.; Jha, B. Selenium and spermine alleviate cadmium induced toxicity in the red seaweed Gracilaria dura by regulating antioxidants and DNA methylation. Plant Physiol. Biochem. 2012, 51, 129–138. [Google Scholar] [CrossRef]
- Sang, M.; Wang, M.; Liu, J.; Zhang, C.; Li, A. Effects of temperature, salinity, light intensity, and pH on the eicosapentaenoic acid production of Pinguiococcus pyrenoidosus. J. Ocean Univ. China 2012, 11, 181–186. [Google Scholar] [CrossRef]
- Zhu, L.; Zhang, X.; Ji, L.; Song, X.; Kuang, C. Changes of lipid content and fatty acid composition of Schizochytrium limacinum in response to different temperatures and salinities. Process. Biochem. 2007, 42, 210–214. [Google Scholar] [CrossRef]
- Msanne, J.; Xu, D.; Konda, A.R.; Casas-Mollano, J.A.; Awada, T.; Cahoon, E.B.; Cerutti, H. Metabolic and gene expression changes triggered by nitrogen deprivation in the photoautotrophically grown microalgae Chlamydomonas reinhardtii and Coccomyxa sp. C-169. Phytochemistry 2012, 75, 50–59. [Google Scholar] [CrossRef]
- Hernández, M.L.; Padilla, M.N.; Sicardo, M.D.; Mancha, M.; Martínez-Rivas, J.M. Effect of different environmental stresses on the expression of oleate desaturase genes and fatty acid composition in olive fruit. Phytochemistry 2011, 72, 178–187. [Google Scholar] [CrossRef]
- Schenk, P.M. Algae Biotechnology. Available online: http://www.algaebiotech.org (accessed on 1 September 2010).
- Guillard, R.R.; Ryther, J.H. Studies of Marine Planktonic Diatoms: I. Cyclotella Nana Hustedt, and Detonula Confervacea (Cleve) Gran. Canadian J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef]
- Timmins, M.; Zhou, W.; Rupprecht, J.; Lim, L.; Thomas-Hall, S.R.; Doebbe, A.; Kruse, O.; Hankamer, B.; Schenk, P.M. The metabolome of Chlamydomonas reinhardtii following induction of anaerobic H2 production by sulfur depletion. J. Biol. Chem. 2009, 284, 23415–23425. [Google Scholar] [CrossRef]
- Lim, D.Y.K. Understanding Tetraselmis sp. lipid production: A new transcriptional profiling approach. 2014; Unpublished work. [Google Scholar]
- Team, R.D.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2008. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Adarme-Vega, T.C.; Thomas-Hall, S.R.; Lim, D.K.Y.; Schenk, P.M. Effects of Long Chain Fatty Acid Synthesis and Associated Gene Expression in Microalga Tetraselmis sp. Mar. Drugs 2014, 12, 3381-3398. https://doi.org/10.3390/md12063381
Adarme-Vega TC, Thomas-Hall SR, Lim DKY, Schenk PM. Effects of Long Chain Fatty Acid Synthesis and Associated Gene Expression in Microalga Tetraselmis sp. Marine Drugs. 2014; 12(6):3381-3398. https://doi.org/10.3390/md12063381
Chicago/Turabian StyleAdarme-Vega, T. Catalina, Skye R. Thomas-Hall, David K. Y. Lim, and Peer M. Schenk. 2014. "Effects of Long Chain Fatty Acid Synthesis and Associated Gene Expression in Microalga Tetraselmis sp." Marine Drugs 12, no. 6: 3381-3398. https://doi.org/10.3390/md12063381