
Profiling of the Molecular Weight and Structural Isomer Abundance of Macroalgae-Derived Phlorotannins

Abstract
:1. Introduction
2. Results and Discussion
2.1. Total Phenolic Content (TPC) and in Vitro Antioxidant Activities

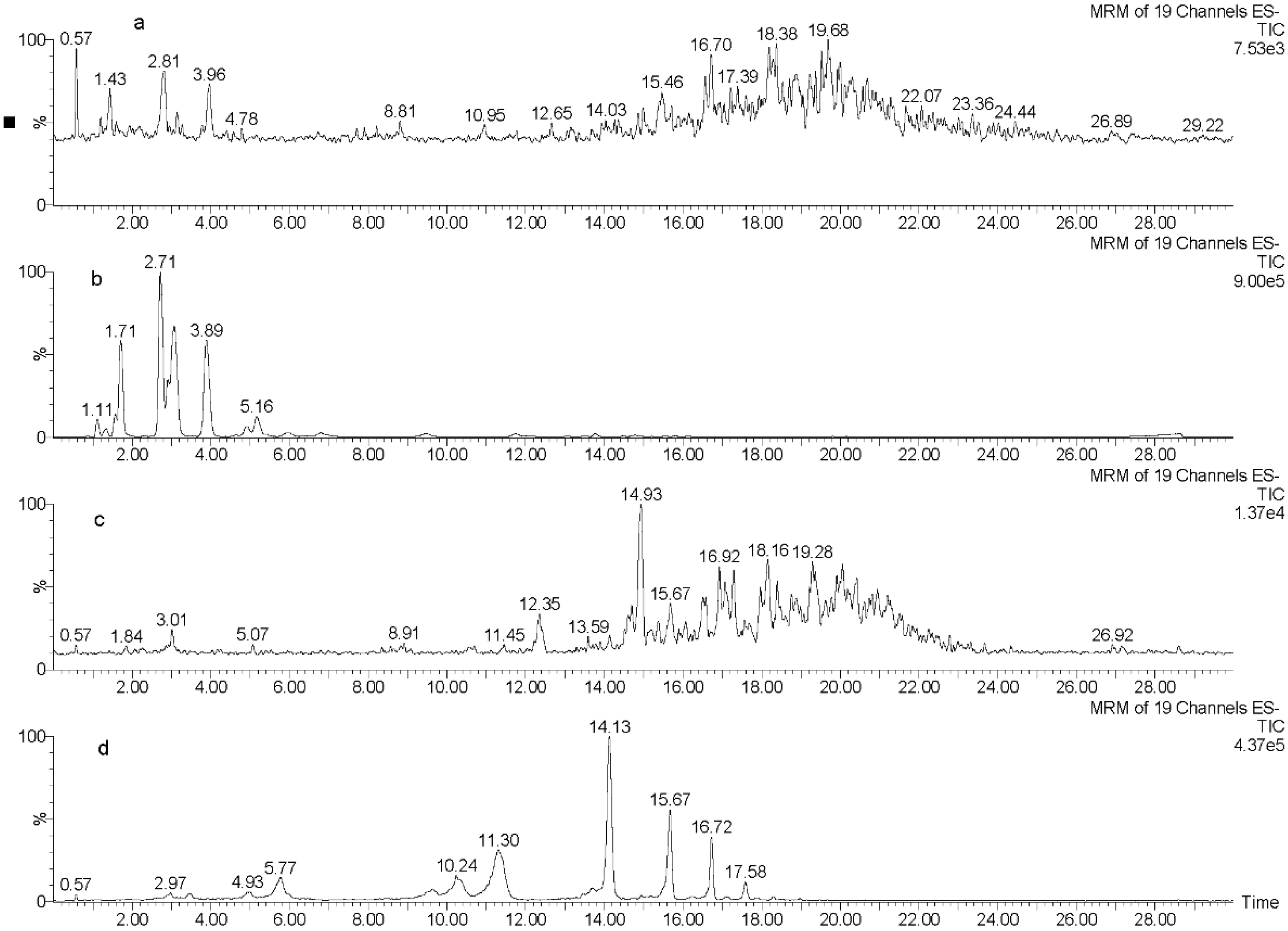{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Seaweed Species | TPC (μg PE/mg Sample) | FRAP (μg TE/mg Sample) | DPPH (IC50 µg/mL) |
|---|---|---|---|
| Fucus vesiculosus | 231.95 ± 8.97 a | 307.27 ± 1.22 a | 4.00 ± 0.01 b |
| Himanthalia elongata | 198.28 ± 9.17 b | 130.89 ± 0.45 b | 14.00 ± 0.04 a |
| Cystoseira nodicaulis | 89.14 ± 2.57 c | 101.35 ± 0.36 b | 28.00 ± 0.01 a |
| Fucus serratus | 180.55 ± 16.98 b | 110.94 ± 0.65 b | 19.00 ± 0.03 a |
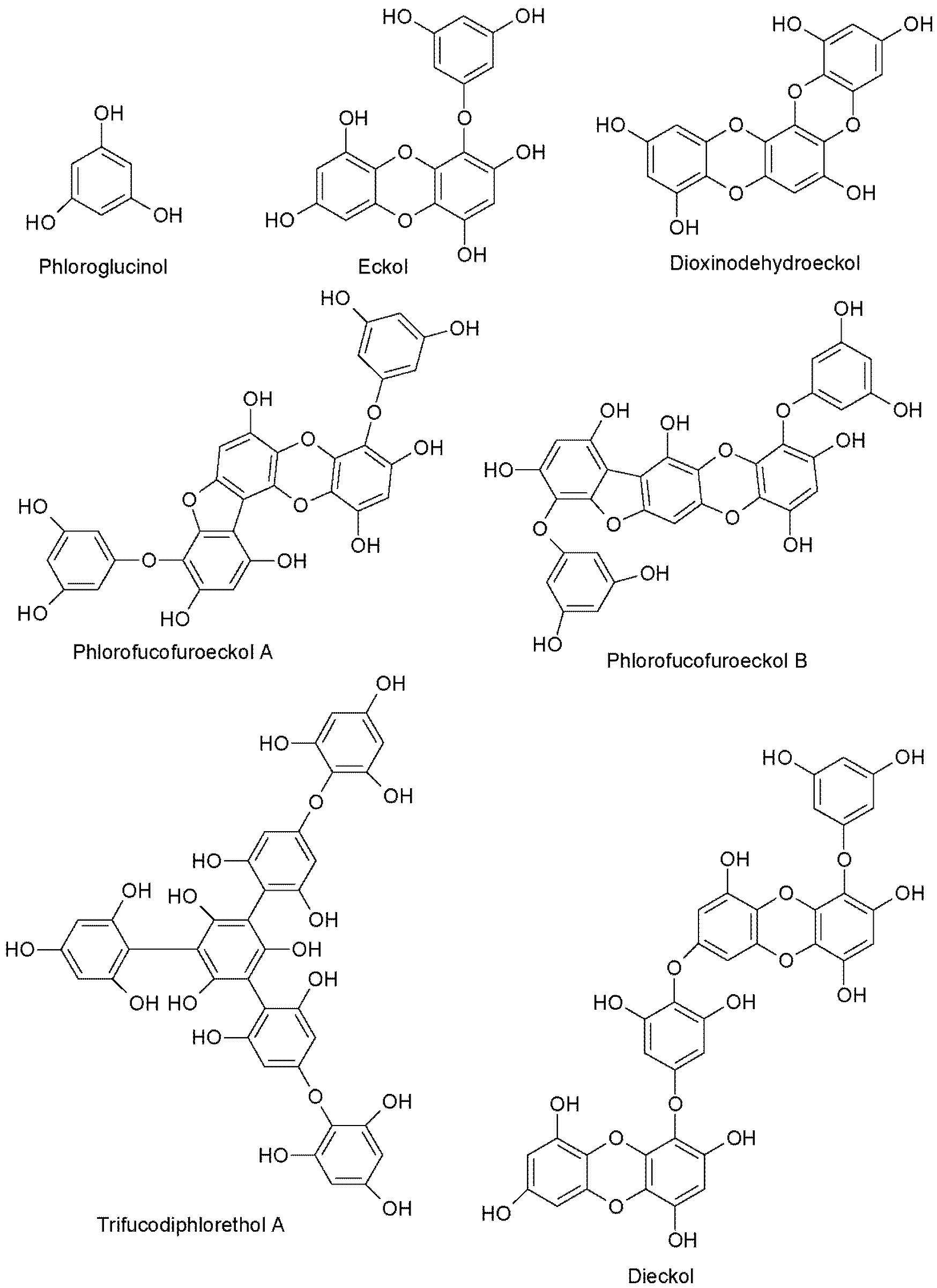
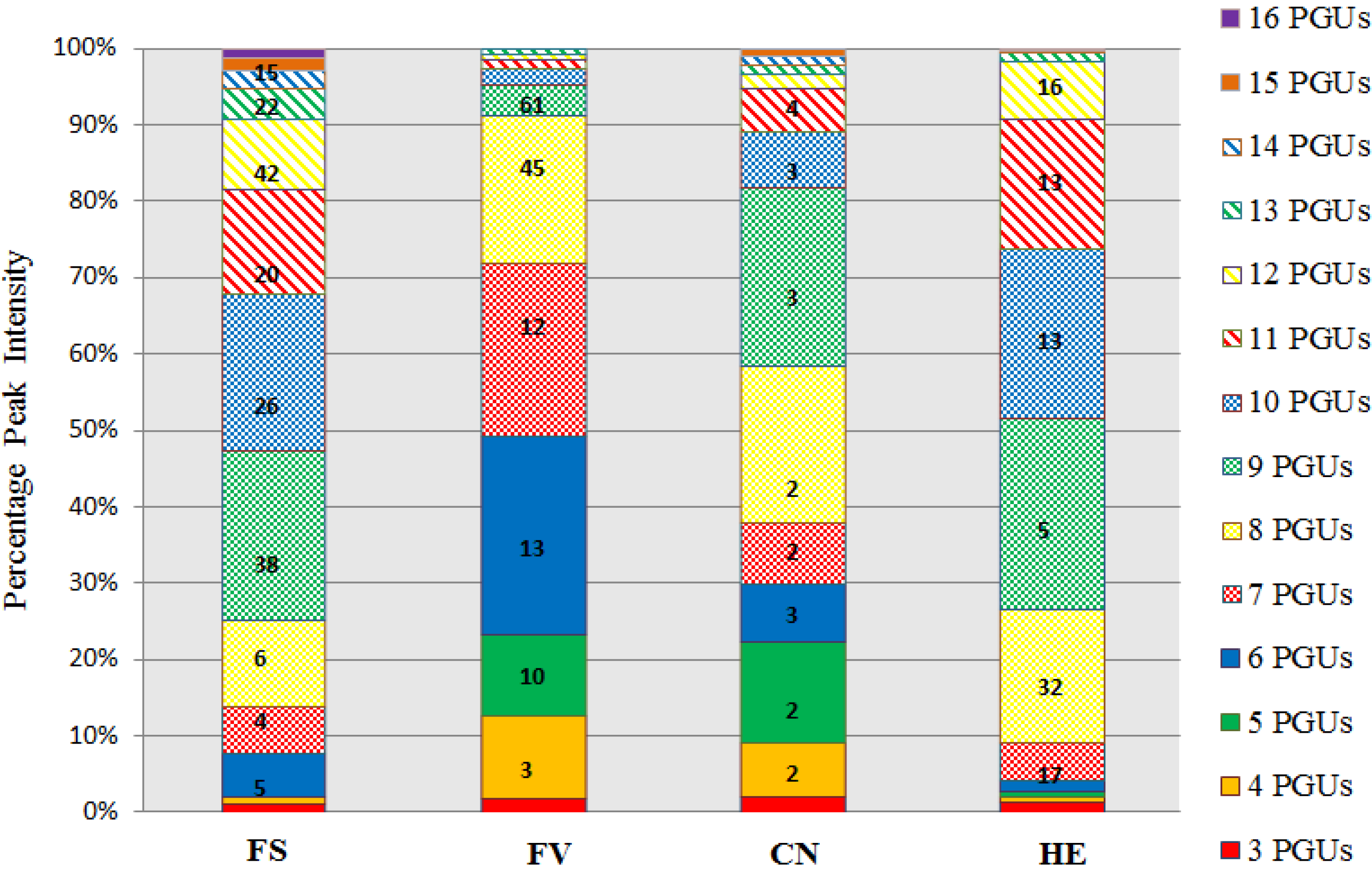
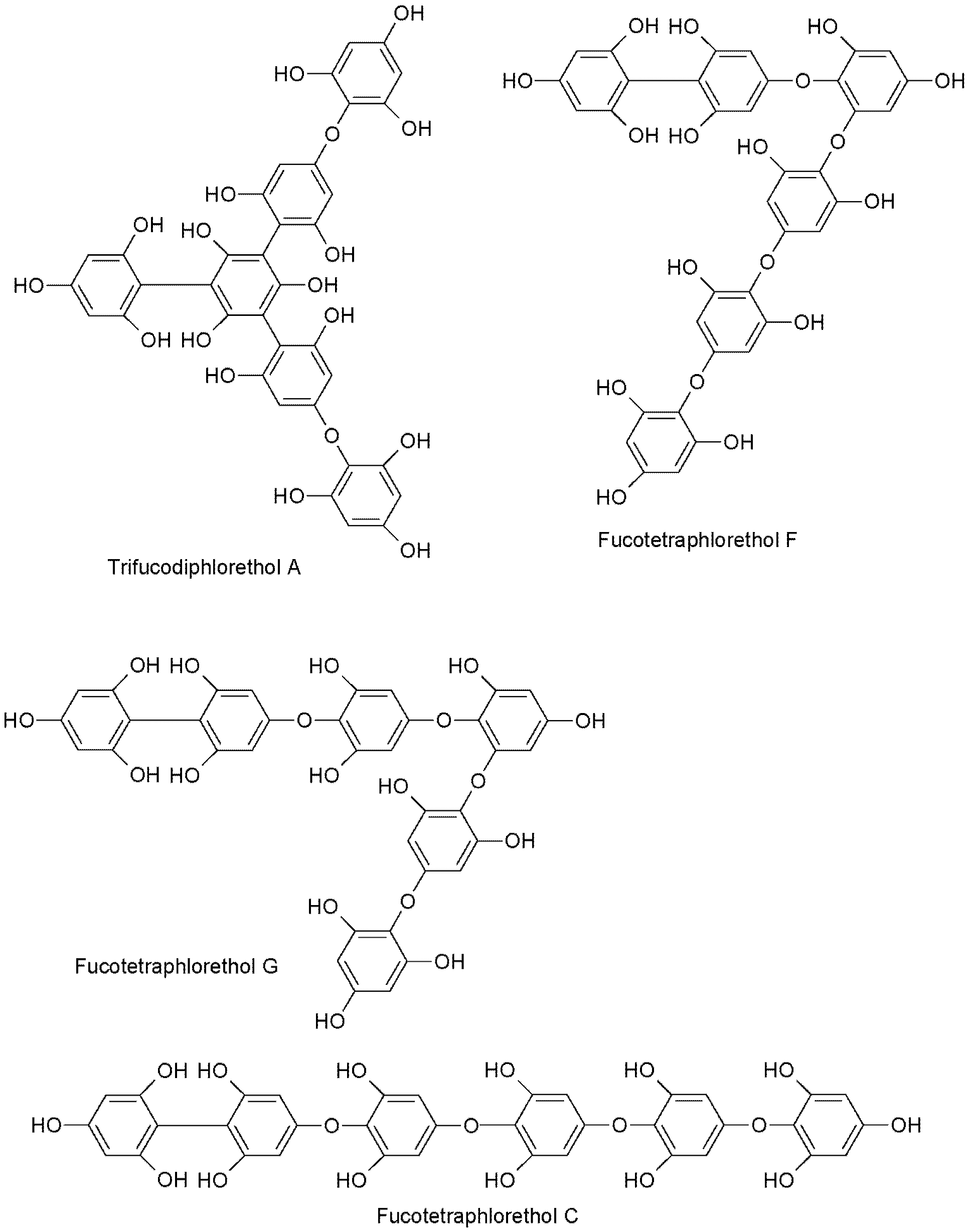
2.2. UPLC-QQQ-MS Profiling of Phlorotannin-Enriched Fractions






3. Experimental Section
3.1. Standards and Reagents
3.2. Samples
3.3. Solid-Liquid Extraction (SLE)
3.4. Partitioning and Molecular Weight Cut-off (MWCO) Dialysis
3.5. Reversed-Phase Flash Chromatography
3.6. Total Phenolic Content (TPC)
3.7. In Vitro Antioxidant Activity
3.7.1. Ferric Reducing Antioxidant Power (FRAP)
3.7.2. DPPH (1,1-diphenyl-2-picryl-hydrazyl) Scavenging Activity
3.8. Statistical Analysis
3.9. UPLC-ESI-MS of Phlorotannin-Enriched Fractions
| PGU’s | Precursor Ion (m/z) | Product Ion (m/z) | Cone Voltage (V) | Collision Voltage (eV) |
|---|---|---|---|---|
| 3 | 373.1 | 139 | 40 | 34 |
| 4 | 497.2 | 139 | 40 | 40 |
| 5 | 621.2 | 139 | 42 | 42 |
| 6 | 745.3 | 139 | 56 | 40 |
| 7 | 869.3 | 139 | 56 | 52 |
| 8 | 993.4 | 229.1 | 60 | 50 |
| 9 | 1117.40 | 229.1 | 70 | 50 |
| 10 | 1241.40 | 229.1 | 70 | 52 |
| 11 | 1365.40 | 229.1 | 70 | 52 |
| 12 | 1489.50 | 229.1 | 70 | 54 |
| 13 | 1614.00 | 229.1 | 70 | 54 |
| 14 | 1738.30 | 229.1 | 70 | 56 |
| 15 | 1862.30 | 229.1 | 70 | 56 |
| 16 | 1986.40 | 229.1 | 70 | 56 |
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lopes, G.; Sousa, C.; Silva, L.R.; Pinto, E.; Andrade, P.B.; Bernardo, J.; Mouga, T.; Valentão, P. Can phlorotannins purified extracts constitute a novel pharmacological alternative for microbial infections with associated inflammatory conditions? PLoS One 2012, 7, e31145. [Google Scholar] [CrossRef] [PubMed]
- Thomas, N.V.; Kim, S.K. Potential pharmacological applications of polyphenolic derivatives from marine brown algae. Environ. Toxicol. Pharmacol. 2011, 32, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Jónsdóttir, R.; Liu, H.; Gu, L.; Kristinsson, H.G.; Raghavan, S.; Ólafsdóttir, G. Antioxidant capacities of phlorotannins extracted from the brown algae Fucus vesiculosus. J. Agric. Food Chem. 2012, 60, 5874–5883. [Google Scholar] [CrossRef] [PubMed]
- Audibert, L.; Fauchan, M.; Blanc, N.; Hauchard, D.; ArGalla, E. Phenolic compounds in the brown seaweed Ascophyllum nodosum: Distribution and radical scavenging activities. Phytochem. Anal. 2010, 21, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.X.; Wijeseker, I.; Li, Y.; Kim, S.K. Phlorotannins as bioactive agents from brown algae. Process Biochem. 2011, 46, 2219–2224. [Google Scholar] [CrossRef]
- Zubia, M.; Robledo, D.; Freile-Pelegrin, Y. Antioxidant activities in tropical marine macroalgae from the Yucatan Penisula, Mexico. J. Appl. Phycol. 2007, 19, 449–458. [Google Scholar] [CrossRef]
- Sailler, B.; Glombitza, K.W. Phlorethols and fucophlorethols from the brown alga Cystophoroa retroflexa. Phytochemistry 1999, 50, 869–881. [Google Scholar] [CrossRef]
- Boettcher, A.A.; Targett, N.M. Role of polyphenolic molecular-size in reduction of assimilation efficiency in Xiphister mucosus. Ecology 1993, 74, 891–903. [Google Scholar] [CrossRef]
- McClintock, J.B.; Baker, B.J. Marine Chemical Ecology; CRC Press: Boca Ratopm, FA, USA, 2001. [Google Scholar]
- Schoenwaelder, M.E.A.; Clayton, M.N. Secretion of phenolic substances into the zygote wall and cellplate in embryos of Hormosira and Acrocarpia (fucales, phaeophyceae). J. Phycol. 1998, 34, 969–980. [Google Scholar] [CrossRef]
- Mazid, M.; Khan, T.A.; Mohammad, F. Role of secondary metabolites in defense mechanisms of plants. Biol. Med. 2011, 3, 232–249. [Google Scholar]
- Steinberg, P.D. Algal chemical defense against herbivores: Allocation of phenolic compounds in the kelp Alaria marginata. Science 1984, 223, 405–407. [Google Scholar] [CrossRef] [PubMed]
- Targett, N.M.; Arnold, T.M. Effects on secondary metabolites on digestion in marine herbivores. In Marine Chemical Ecology; McClinlock, J.B., Baker, B.J., Eds.; CRC Press: Boca Ratopm, FL, USA, 2001; pp. 391–412. [Google Scholar]
- Waterman, P.G.; Mole, S. Analysis of Phenolic Plant Metabolities; Blackwell Scientific Publication: Oxford, UK, 1994; pp. 85–87. [Google Scholar]
- Pavia, H.; Toth, G.B. Inducible chemical resistance to herbivory in the brown seaweed Acophyllum nodosum. Ecology 2000, 81, 3212–3225. [Google Scholar] [CrossRef]
- Pavia, H.; Cervin, G.; Lindgren, A.; Åberg, P. Effects of UV-B radiation and simulated herbivory on phlorotannins in the brown alga Ascophyllum nodosum. Mar. Ecol. Prog. Ser. 1997, 157, 139–146. [Google Scholar] [CrossRef]
- Davis, A.R.; Tagett, N.M.; McConnell, O.J.; Young, C.M. Epibiosis of marine algae and benthic invertebrates: Natural products chemistry and other mechanisms inhibiting settlement and growth. In Biorganic Marine Chemistry; Schever, P.J., Ed.; Springer Verlag: Berlin/Heidelberg, Germany, 1989; Volume 3, pp. 85–114. [Google Scholar]
- Schoenwaelder, M.E.A. The occurrence and cellular significance of physodes in brown algae. Phycologia 2002, 41, 125–139. [Google Scholar] [CrossRef]
- Arnold, T.M.; Targett, N.M. To grow and defend: Lack of trade-offs for brown algal phlorotannins. Oikas 2003, 100, 406–408. [Google Scholar] [CrossRef]
- Shibata, T.; Ishimaru, K.; Kawaguchi, S.; Yoshikawa, H.; Hama, Y. Antioxidant activities of phlorotannins isolated from Japanese Laminariceae. J. Appl. Phycol. 2008, 20, 705–711. [Google Scholar] [CrossRef]
- Tierney, M.S.; Soler-Vila, A.; Croft, A.K.; Hayes, M. Antioxidant Activity of the Brown Macroalgae Fucus spiralis Linnaeus Harvested from the West Coast of Ireland. Curr. Res. J. Biol. Sci. 2013, 5, 81–90. [Google Scholar]
- Tierney, M.S.; Smyth, T.J.; Hayes, M.; Soler-Vila, A.; Croft, A.K.; Brunton, N. Influence of pressurized liquid extraction and solid liquid extraction methods on the phenolic content and antioxidant activities of Irish macroalgae. Int. J. Food Technol. 2013, 48, 860–869. [Google Scholar] [CrossRef]
- Li, Y.; Lee, S.H.; Le, Q.T.; Kim, M.M.; Kim, S.K. Anti-allergic effects of phlorotannins on histamine release via binding inhibition between IgE and Fc epsilon RI. J. Agric. Food Chem. 2008, 56, 12073–12080. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.R.; Shin, T.S.; Lee, M.S.; Park, J.Y.; Park, K.E.; Yoon, N.Y.; Kim, J.S.; Choi, J.S.; Jang, B.C.; Byun, D.S.; et al. Isolation and identification of phlorotannins from Ecklonia stolonifera with antioxidant and anti-inflammatory properties. J. Agric. Food Chem. 2009, 57, 3483–3489. [Google Scholar] [CrossRef] [PubMed]
- Artan, M.; Li, Y.; Karadeniz, F.; Lee, S.H.; Kim, M.M.; Kim, S.K. Anti-HIV-1 activity of phloroglucinol derivative 6,6′-bieckol, from Ecklonia cava. Bio-organ Med. Chem. 2008, 16, 7921–7926. [Google Scholar] [CrossRef]
- Kong, C.S.; Kim, J.A.; Yoon, N.Y.; Kim, S.K. Induction of apoptosis by phloroglucinol derivative from Ecklonia cava in MCF-7 human breast cancer cells. Food Chem. Toxicol. 2009, 47, 1653–1658. [Google Scholar] [CrossRef] [PubMed]
- Okada, Y.; Ishimaru, A.; Suzuki, R.; Okuyama, T. A new phloroglucinol derivative from the brown algae Eisenia bicyclis: Potential for the effective treatment of diabetic complications. J. Nat. Prod. 2004, 67, 103–105. [Google Scholar] [CrossRef] [PubMed]
- Lordan, S.; Smyth, T.J.; Soler-Vila, A.; Stanton, C.; Ross, R.P. The α-amylase and α-glucosidase inhibitory effects of Irish seaweed extracts. Food Chem. 2013, 141, 2170–2176. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.; Chen, T.; Nines, R.G.; Shin, H.C.; Stoner, G.D. Photochemoprevention of UVBinduced skin carcinogenesis in SKH-1 mice by brown algae polyphenols. Int. J. Cancer 2006, 119, 2742–2749. [Google Scholar] [CrossRef] [PubMed]
- Fukuyama, Y.; Kodama, M.; Miura, I.; Kinzyo, Z.; Mori, H.; Nakayama, Y.; Takahashi, M. Anti-plasmin Inhibitor V. Structures of novel dimeric eckols isolated from the brown alga Ecklonia kurome OKAMURA. Chem. Pharm. Bull. 1989, 37, 2438–2440. [Google Scholar] [CrossRef] [PubMed]
- Shibata, T.; Fujimoto, K.; Nagayama, K.; Yamaguchi, K.; Nakamura, T. Inhibitory activity of brown algal phlorotannins against hyaluronidase. Int. J. Food Sci. Technol. 2002, 37, 703–709. [Google Scholar] [CrossRef]
- Fujii, Y.; Tanaka, R.; Miyake, H.; Tamaru, Y.; Ueda, M.; Shibata, T. Evaluation for antioxidative properties of phlorotannins isolated from the brown alga Eisenia bicyclis, by the HORAC method. Food Nutr. Sci. 2013, 4, 478–482. [Google Scholar] [CrossRef]
- Ferreres, F.; Lopes, G.; Gil-Izquierdo, A.; Andrade, P.B.; Sousa, C.; Mouga, T.; Valentao, P. Phlorotannin extracts from Fucales characterized by HPLC-DAD-ESI-MS; Approaches to hyaluronidase inhibitory capacity and antioxidant properties. Mar. Drugs 2012, 10, 2766–2781. [Google Scholar] [CrossRef] [PubMed]
- Montero, L.; Herrero, M.; Ibáňez, E.; Cifuentes, A. Separation and characterization of phlorotannins from brown algae Cystoseira abies-marina by comprehensive two-dimensional liquid chromatography. Electrophoresis 2014, 35, 1644–1651. [Google Scholar] [CrossRef] [PubMed]
- Steevensz, A.J.; MacKinnon, S.L.; Hankinson, R.; Craft, C.; Connan, S.; Stengel, D.B.; Melansona, J.E. Profiling phlorotannina in brown macroalgae by Liquid Chromatography-High Resolution Mass Spectrometry. Phytochem. Anal. 2012, 23, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Tierney, M.S.; Soler-Vila, A.; Rai, D.K.; Croft, A.K.; Brunton, N.P.; Smyth, T.J. UPLC-MS profiling of low molecular weight phlorotannin polymers in Ascophyllum nodosum, Pelvetia canaliculata and Fucus spiralis. Metabolomics 2014, 10, 524–535. [Google Scholar]
- Wang, T.; Jónsdóttir, R.; Ólafsdóttir, G. Total phenolic compounds, radical scavenging and metal chelation of extracts from Icelandic seaweeds. Food Chem. 2009, 116, 240–248. [Google Scholar] [CrossRef]
- Heffernan, N.; Smyth, T.; Soler-Villa, A.; FitzGerald, R.J.; Brunton, N.P. Phenolic content and antioxidant activity of fractions obtained from selected Irish macroalgae species (Laminaria digitata, Fucusserratus, Gracilaria gracilis and Codium fragile). J. Appl. Phycol. 2014, 26. [Google Scholar] [CrossRef]
- Koivikko, R.; Loponen, J.; Pihlaja, K.; Jormalainen, V. High-performance liquid chromatographic analysis of phlorotannins from the brown alga Fucus vesiculosus. Phytochem. Anal. 2007, 18, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Hagerman, A.E.; Riedl, K.M.; Jones, G.A.; Sovik, K.N.; Ritchard, N.T.; Hartzfeld, P.W.; Riechel, T.L. High molecular weight plant polyphenolics (tannins) as biological antioxidants. J. Agric. Food Chem. 1998, 46, 1887–1892. [Google Scholar] [CrossRef]
- Fan, X.; Yan, X.; Fang, G.; Chen, Y.; Lou, G. Antioxidative properties of high molecular weight polyphenols from brown seaweed. Acta Hydrobiol. Sinica 1999, 5, 494–499. [Google Scholar]
- Wei, Y.X.; Xu, Z.H. Studies on antioxidative activity of high molecular weight polyphenols from two kinds of brown algae. Chin. Tradit. Herb. Drugs 2003, 34, 317–319. [Google Scholar]
- Glombitza, K.W.; Pauli, K. Fucols and Phloretols from the Brown alga Scytothaminus australis Hook. Et Harv (Chnoosporaceae). Bot. Mar. 2003, 46, 315–320. [Google Scholar] [CrossRef]
- Grosse-Damhues, J.; Glombitza, K.W.; Schulter, H.R. An eight-ring phlorotannin from the brown alga Himanthalia elongata. Phytochemistry 1983, 22, 2043–2046. [Google Scholar] [CrossRef]
- Glombitza, K.W.; Grobe-Damhues, J. Antibiotics from algae XXXIII. Phlorotannins of the brown alga Himanthalia elongata. Planta Medica 1985, 1, 42–46. [Google Scholar] [CrossRef]
- Pary, S.; Kehraus, S.; Krick, A.; Glombitza, K.W.; Carmeli, S.; Klimo, K.; Gerhäuser, C.; König, G.M. In vitro chemopreventative potential of fucophlorethols from the brown alga Fucus vesiculosus L. by antioxidant activity and inhibition of selected cytochrome P450 enzymes. Phytochemistry 2010, 71, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Koivikko, R. Brown Algal Phlorotannins: Improving and Applying Chemical Methods (Annales Universitatis Turkvensis A1 381). Ph.D. Thesis, University of Turku, Turku, Finland, February 2008. [Google Scholar]
- Liu, H.; Cu, L. Phlorotannins from brown algae (Fucus vesiculosus) inhibited the formation of advanced glycation end products by scavenging reactive carbonyls. J. Agric. Food Chem. 2012, 60, 1326–1334. [Google Scholar] [CrossRef] [PubMed]
- Heffernan, N.; Smyth, T.J.; FitzGerald, R.J.; Soler-Vila, A.; Brunton, N. Antioxidant activity and phenolic content of pressurized liquid and solid liquid extracts from four Irish origin macroalgae. Int. J. Food Sci. Technol. 2014, 49, 1765–1771. [Google Scholar] [CrossRef]
- Kang, M.C.; Kim, E.A.; Kang, S.M.; Wijesinghe, W.A.J.P.; Yang, X.; Kang, N.; Jeon, Y.J. Thermostability of a marine polyphenolic antioxidant dieckol, derived from the brown seaweed Ecklonia cava. Algae 2012, 27, 205–213. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Stratil, P.; Klejdus, B.; Kuban, V. Determination of total content of phenolic compounds and their antioxidant activity in vegetables evaluation of spectrophotometric methods. J. Agric. Food Chem. 2006, 54, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Goupy, P.; Hughes, M.; Boivin, P.; Amiot, M.J. Antioxidant composition and activity of barley (Hordeum vulgare) and malt extracts and of isolated phenolic compounds. J. Sci. Food Agric. 1999, 79, 1625–1634. [Google Scholar] [CrossRef]
- Ollanketo, M.; Peltoketo, A.; Hartonen, K.; Hiltunen, R.; Riekkola, M.L. Extraction of sage (Salvia officinalis) by pressurized hot water and conventional methods: Antioxidant activity of the extracts. Eur. Food Res. Technol. 2002, 215, 158–163. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Culvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heffernan, N.; Brunton, N.P.; FitzGerald, R.J.; Smyth, T.J. Profiling of the Molecular Weight and Structural Isomer Abundance of Macroalgae-Derived Phlorotannins. Mar. Drugs 2015, 13, 509-528. https://doi.org/10.3390/md13010509
Heffernan N, Brunton NP, FitzGerald RJ, Smyth TJ. Profiling of the Molecular Weight and Structural Isomer Abundance of Macroalgae-Derived Phlorotannins. Marine Drugs. 2015; 13(1):509-528. https://doi.org/10.3390/md13010509
Chicago/Turabian StyleHeffernan, Natalie, Nigel P. Brunton, Richard J. FitzGerald, and Thomas J. Smyth. 2015. "Profiling of the Molecular Weight and Structural Isomer Abundance of Macroalgae-Derived Phlorotannins" Marine Drugs 13, no. 1: 509-528. https://doi.org/10.3390/md13010509