
Dereplication and Chemotaxonomical Studies of Marine Algae of the Ochrophyta and Rhodophyta Phyla


Abstract
:
1. Introduction
2. Results and Discussion
2.1. Chemical Profiling (HPLC-NMR & HPLC-MS)

{kind=link}
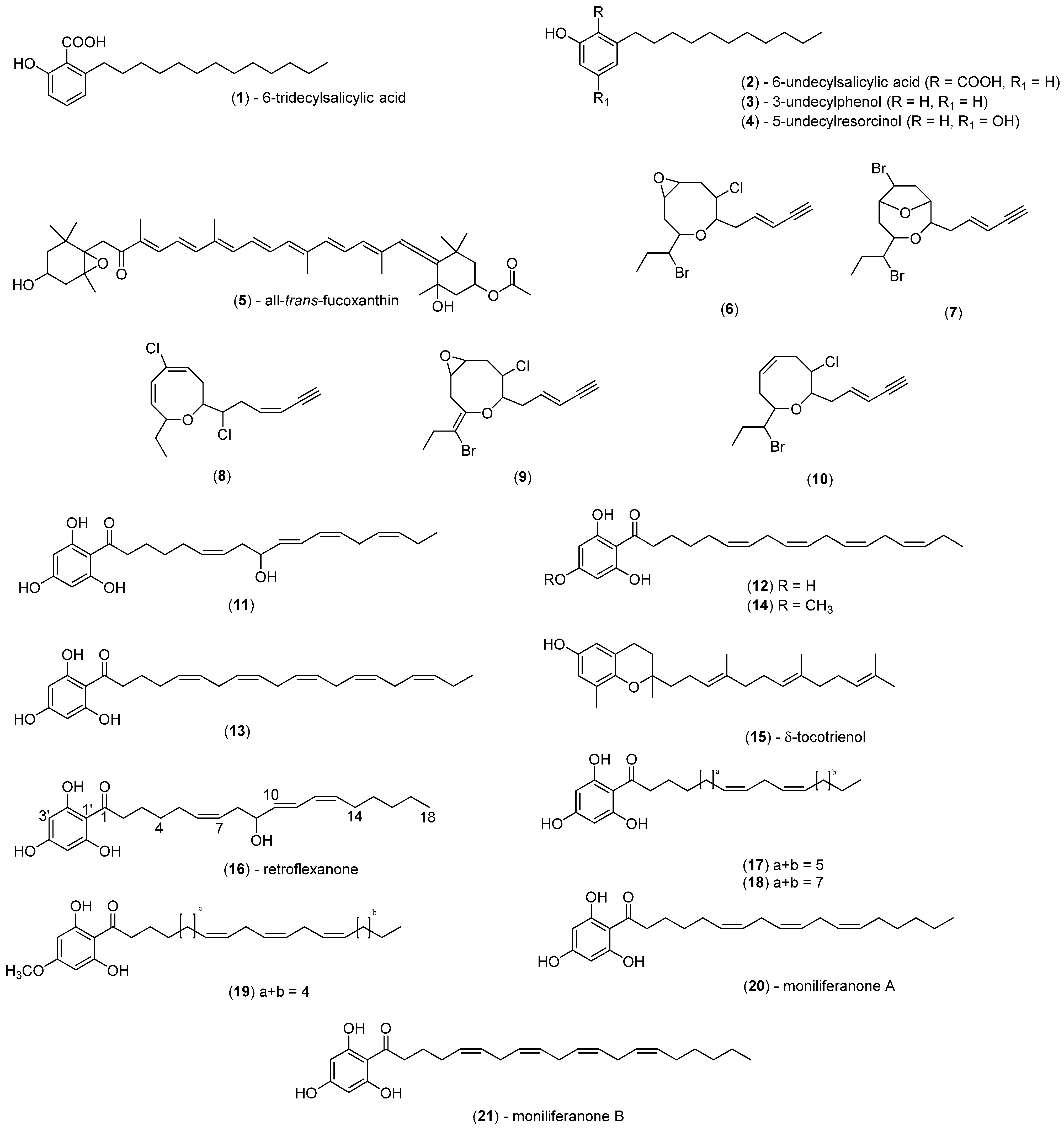{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Alga | Chemical Class(es) Present | Compounds Present * |
|---|---|---|
| C. subfarcinata | Phloroglucinols | 18, 19 |
| C. retroflexa | Phloroglucinols | 11-13, 16, 18, 20, 21 |
| S. cf. fallax | Phlroglucinols, tocotrienols | 12, 14, 15, 17, 20, 21 |
| S. decipiens | Phenols, phenolic acids, resorcinols | 1–4 |
| S. vestitum | Xanthophylls | 5 |
| H. pseudospicata | Xanthophylls | 5 |
| Laurencia sp. | Polyhalogenated C15 acetogenins | - |
| Peak # | Rt (min) | Compound | Structure Class | Marine Alga (~Amount Present in μg) | UV (nm) | MarinLit Search Parameters | New/Known |
|---|---|---|---|---|---|---|---|
| 1 | 2.29 | (2) | Phenolic acid | S. decipiens (500–1000) | 240, 302 | Compound not in MarinLit database | Known |
| 2 | 2.44 | (3) | Phenol | S. decipiens (500–1000) | 236, 301 | Compound not in MarinLit database | Known |
| 3 | 3.42 | (11) | Phloroglucinol | C. retroflexa (750–1000) | 235, 285 | Molecular formula, UV ± 5 | Known |
| 4 | 3.55 | (1) | Phenolic acid | S. decipiens (<100) | 235, 301 | Molecular formula, UV ± 10 | Known |
| 5 | 4.45 | (16) | Phloroglucinol | C. retroflexa (750–1000) | 235, 285 | Compound not in MarinLit database | New |
| 6 | 5.00 | n.a. | C15 acetogenin | Laurencia sp. (<25) | 220, 237 | Genus, UV ± 5, 1 triplet methyl group | Not Identified |
| 7 | 6.05 | n.a. | Unknown | Laurencia sp. (500–1000) | 220, 240, 255 | Unable to dereplicate using any parameters | Not Identified |
| 8 | 6.70 | n.a. | C15 acetogenin | Laurencia sp. (<25) | 220, 237 | Genus, UV ± 5, 1 triplet methyl group | Not Identified |
| 9 | 7.87 | (4) | Resorcinol | S. decipiens (<10) | 229, 276, 281 | Compound not in MarinLit database | Known |
| 10 | 9.98 | (12) | Phloroglucinol | C. retroflexa (500–1000), S. cf. fallax (<250) | 230, 285 | Molecular formula, UV ± 5 | Known |
| 11 | 12.95 | (13) | Phloroglucinol | C. retroflexa (250–750) | 230, 285 | Molecular formula, UV ± 5 | Known |
| 12 | 13.65 | (17) | Phloroglucinol | S. cf. fallax (<250) | 215, 228, 285 | Compound not in MarinLit database | New |
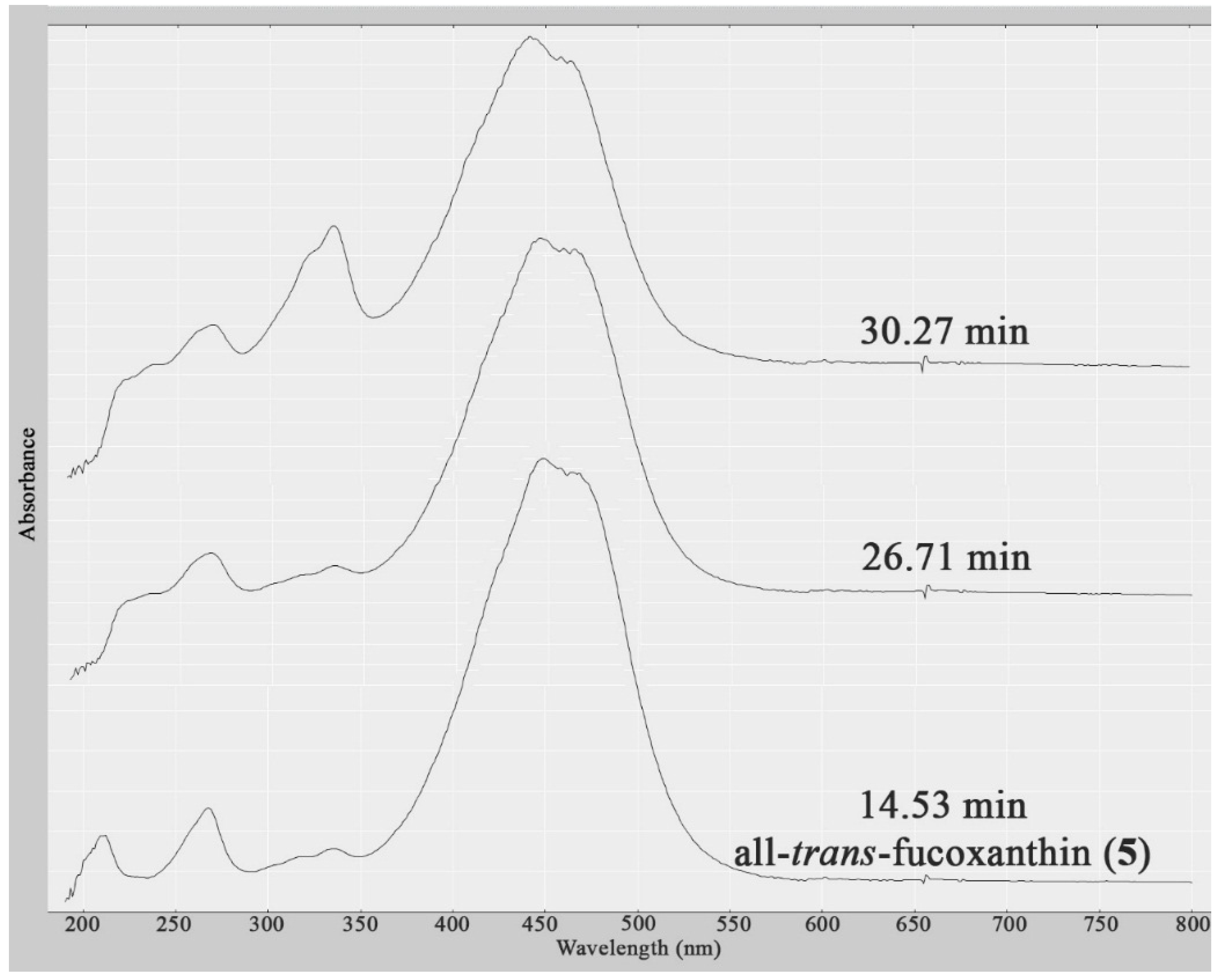
| 13 | 14.53 | (5) | Xanthophyll | S. vestitum (<100), H. pseudospicata (<100) | 450 | Molecular formula | Known |
| 14 | 15.50 | (20) | Phloroglucinol | C. retroflexa (<500), S. cf. fallax (<100) | 212, 228, 285 | Compound not in MarinLit database | Known [13] |
| 15 | 20.15 | (21) | Phloroglucinol | C. retroflexa (<250), S. cf. fallax (<50) | 230, 285 | Compound not in MarinLit database | Known [13] |
| 16 | 21.62 | (14) | Phloroglucinol | S. cf. fallax (<100) | 212, 228, 285 | Molecular formula, UV ± 5 | Known |
| 17 | 22.96 | (18) | Phloroglucinol | C. retroflexa (<100), C. subfarcinata (<50) | 228, 285 | Compound not in MarinLit database | New |
| 18 | 23.16 | n.a. | Phloroglucinol | C. retroflexa (<100) | 238, 288 | Genus, UV ± 5 | Not Identified |
| 19 | 26.71 | n.a. | Xanthophyll | S. vestitum (<5), H. pseudospicata (<5) | 450 | Insufficient data to search MarinLit Database | Not Identified |
| 20 | 30.27 | n.a. | Xanthophyll | S. vestitum (<5), H. pseudospicata (<5) | 450 | Insufficient data to search MarinLit Database | Not Identified |
| 21 | 33.40 | (19) | Phloroglucinol | C. subfarcinata (<50) | 213, 228, 285 | Compound not in MarinLit database | New |
| 22 | 60.80 | (15) | Tocotrienol | S. cf. fallax (<10) | 212, 300 | Class, UV ± 5, contains only singlet aromatic/vinyl CH3 groups, aromatic ring | Known |

2.2. Identification of Phenols, Phenolic Acids and Resorcinols
2.3. Identification of Xanthophylls


2.4. Identification of Halogenated C15 Acetogenins

2.5. Identification of Phloroglucinols and Tocotrienols
| Retroflexanone (16) | ||||
|---|---|---|---|---|
| Position | δH (J in Hz) | δC a, Type | gCOSY | gHMBCAD |
| 1 | 207.2, s | |||
| 2 | 3.89, t (7.0) | 44.2, t | 3 | 1, 3, 4 |
| 3 | 2.48, p (7.0) | 25.1, t | 2, 4 | 1, 2, 4, 5 |
| 4 | 2.23, m | 29.9, t | 3 | 2, 3, 5, 6 |
| 5 | SS | 27.8, t | ||
| 6 | 6.32, m | 132.6, d | 7 | |
| 7 | 6.23, m | 126.2, d | 6, 8 | |
| 8 | 3.12, m | 35.8, t | 7 w, 9 | 6, 7, 9 |
| 9 | 4.96, dt (6.5, 7.0) | 72.6, d | 8, 10 | 11 |
| 10 | 6.48, dd (7.0, 15.0) | 136.6, d | 9, 11 | 12 |
| 11 | 7.30, dd (15.0, 11.0) | 126.4, d | 10, 12 | |
| 12 | 6.80, t (11.0) | 128.8, d | 11, 13 w | |
| 13 | 6.28, m | 133.3, d | 12, 14 | |
| 14 | 3.00, m b | ND | 12, 13, 15 | |
| 15 | SS | 29.9, t | ||
| 16 | SS | 32.0, t | ||
| 17 | 2.13, m | 23.0, t | 18 | 16 |
| 18 | 1.72, t (6.5) | 14.3, q | 17 | 16, 17 |
| 1′ | 105.2, s | |||
| 2′ | 165.0, s * | |||
| 3′ | 6.72, s | 95.7, d | 1′, 2′, 5′ | |
| 4′ | ND * | |||
| 5′ | 6.72, s | 95.7, d | 1′, 3′, 6′ | |
| 6′ | 165.0, s * | |||
| 2′-OH | ND | |||
| 4′-OH | ND | |||
| 6′-OH | ND | |||
2.6. Anti-Microbial Activity
3. Experimental Section
3.1. Marine Alga Material
3.2. Extraction
| Crude | Microorganism Concentration (mg/mL) | E. coli ATCC 25922 | S. aureus ATCC 25923 | S. aureus MRSA 344/2-32 | P. aeruginosa ATCC 27853 | S. pyogenes 345/1 | B. subtilis ATCC 19659 | C. albicans ATCC 10231 or 14053* | T. mentagrophytes ATCC 28185 | C. resinae |
|---|---|---|---|---|---|---|---|---|---|---|
| S. vestitum (3:1 MeOH/DCM) | 50 | ND a | NT b | NT b | 3 | NT b | ND a | 3 * | 2 | ND a |
| H. pseudospicata (3:1 MeOH/DCM) | 50 | ND a | NT b | NT b | ND a | NT b | ND a | ND a | ND a | ND a |
| S. cf. fallax (3:1 MeOH/DCM) | 50 | ND a | NT b | NT b | ND a | NT b | 2 | ND a* | ND a | ND a |
| C. subfarcinata (3:1 MeOH/DCM) | 50 | ND a | NT b | NT b | 1 | NT b | ND a | 3* | ND a | ND a |
| C. retroflexa (DCM) | 50 | ND a | 2 | 3 | 5 | ND a | NT b | ND a | NT b | NT b |
| C. retroflexa (MeOH) | 50 | 1 | 4 | 6 | 1 | ND a | NT b | ND a | NT b | NT b |
3.3. Biological Evaluation
3.4. Chemical Profiling
3.5. HPLC-NMR & HPLC-MS Conditions
3.6. On-Line (HPLC-NMR & HPLC-MS) Partial Characterization of Compounds
4. Conclusions
Acknowledgments
Author Contributions
Supplementary Information
Conflicts of Interest
References
- Guiry, M.D.; Guiry, G.M. AlgaeBase. Available online: http://www.algaebase.org (accessed on 6 March 2015).
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2015, 32, 116–211. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Pereira, A.R.; Gerwick, W.H. The Chemistry of Marine Algae and Cyanobacteria. In Handbook of Marine Natural Products; Fattorusso, E., Gerwick, W.H., Taglialatela-Scafati, O., Eds.; Springer: London, England, UK, 2012. [Google Scholar]
- Sailler, B.; Glombitza, K.-W. Halogenated phlorethols and fucophlorethols from the brown alga Cystophora retroflexa. Nat. Toxins 1999, 7, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Sailler, B.; Glombitza, K.-W. Phlorethols and fucophlorethols from the brown alga Cystohpora retroflexa. Phytochemistry 1999, 50, 869–881. [Google Scholar] [CrossRef]
- Czeczuga, B.; Taylor, F.J. Carotenoid content in some species of the brown and red algae from the coastal area of New Zealand. Biochem. Syst. Ecol. 1986, 15, 5–8. [Google Scholar] [CrossRef]
- Laird, D.W.; Bennett, S.; Bian, B.; Sauer, B.; Wright, K.; Hughes, V.; van Altena, I.A. Chemical investigation of seven Australasian Cystophora species: New chemistry and taxonomic insights. Biochem. Syst. Ecol. 2010, 38, 187–194. [Google Scholar] [CrossRef]
- MarinLit Database. Available online: http://pubs.rsc.org/marinlit/ (accessed on 6 March 2015).
- Kazlauskas, R.; Murphy, P.T.; Wells, R.J.; Daly, J.J.; Oberhansli, W.E. Heterocladol, a halogenated selinane sesquiterpene of biosynthetic significance from the red alga Laurencia filiformis: Its isolation, crystal structure and absolute configuration. Aust. J. Chem. 1977, 30, 2679–2687. [Google Scholar] [CrossRef]
- Capon, R.; Ghisalberti, E.L.; Jefferies, P.R.; Skelton, B.W.; White, A.H. Sesquiterpene metabolites from Laurencia filiformis. Tetrahedron 1981, 37, 1613–1621. [Google Scholar] [CrossRef]
- Capon, R.J.; Ghisalberti, E.L.; Mori, T.A.; Jefferies, P.R. Sesquiterpenes from Laurencia spp. J. Nat. Prod. 1988, 51, 1302–1304. [Google Scholar] [CrossRef]
- Dias, D.A.; White, J.M.; Urban, S. Laurencia filiformis: Phytochemical profiling by conventional and HPLC-NMR approaches. Nat. Prod. Commun. 2009, 4, 157–172. [Google Scholar] [PubMed]
- Brkljača, R.; Urban, S. Dereplication (HPLC-NMR & HPLC-MS) and structural identification studies of marine brown algae of the genus Cystophora. Phytochemistry 2015. submitted for publication. [Google Scholar]
- Suzuki, M.; Takahashi, Y.; Matsuo, Y.; Guiry, M.D.; Masuda, M. Scanlonenyne, a novel halogenated C15 acetogenin from the red alga Laurencia obtusa in Irish waters. Tetrahedron 1997, 53, 4271–4278. [Google Scholar] [CrossRef]
- San-Martin, A.; Darias, J.; Soto, H.; Contreras, C.; Herrera, J.S.; Rovirosa, J. A new C15 acetogenin from the marine alga Laurencia claviformis. Nat. Prod. Lett. 1997, 10, 303–311. [Google Scholar] [CrossRef]
- Suzuki, M.; Nakano, S.; Takahashi, Y.; Abe, T.; Masuda, M. Bisezakyne-A and -B, halogenated C15 acetogenins from a Japanese Laurencia species. Phytochemistry 1999, 51, 657–662. [Google Scholar] [CrossRef]
- Aydogmus, Z.; Imre, S.; Ersoy, L.; Wray, V. Halogenated secondary metabolites from Laurencia obtusa. Nat. Prod. Res. 2004, 18, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Takaichi, S. Carotenoids in Algae: Distributions, Biosyntheses and Functions. Mar. Drugs 2011, 9, 1101–1118. [Google Scholar] [CrossRef] [PubMed]
- Brkljača, R.; Urban, S. Limit of detection studies for application to natural product identification using high performance liquid chromatography coupled to nuclear magnetic resonance spectroscopy. J. Chromatogr. A 2015, 1375, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Kazlauskas, R.; Mulder, J.; Murphy, P.T.; Wells, R.J. New metabolites from the brown alga Caulocystis cephalornithos. Aust. J. Chem. 1980, 33, 2097–2101. [Google Scholar] [CrossRef]
- Narkowicz, C.K.; Blackman, A.J. Further acetogenins from Tasmanian collections of Caulocystis cephalornithos demonstrating chemical variability. Biochem. Syst. Ecol. 2006, 34, 635–641. [Google Scholar] [CrossRef]
- Spencer, G.F.; Tjarks, L.W.; Kleiman, R. Alkyl and phenylalkyl anacardic acids from Knema elegans seed oil. J. Nat. Prod. 1980, 43, 724–730. [Google Scholar] [CrossRef] [PubMed]
- Asakawa, Y.; Masuya, T.; Tori, M.; Campbell, E.O. Long chain alkyl phenols from the liverwort Schistochila appendiculata. Phytochemistry 1987, 26, 735–738. [Google Scholar] [CrossRef]
- Englert, G.; Bjornland, T.; Liaaen-Jensen, S. 1D and 2D NMR studies of some allenic carotenoids of the fucoxanthin series. Magn. Reson. Chem. 1990, 28, 519–528. [Google Scholar] [CrossRef]
- Haugan, J.A.; Englert, G.; Glinz, E.; Liaaen-Jensen, S. Algal carotenoids. 48. Structural assignments of geometrical isomers of fucoxanthin. Acta Chem. Scand. 1992, 46, 389–395. [Google Scholar] [CrossRef]
- Ohnmacht, S.; West, R.; Simionescu, R.; Atkinson, J. Assignment of the 1H and 13C NMR of tocotrienols. Magn. Reson. Chem. 2008, 46, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Gregson, R.P.; Kazlauskas, R.; Murphy, P.T.; Wells, R.J. New metabolites from the brown alga Cystophora torulosa. Aust. J. Chem. 1977, 30, 2527–2532. [Google Scholar] [CrossRef]
- Kazlauskas, R.; King, L.; Murphy, P.T.; Warren, R.G.; Wells, R.J. New metabolites from the Brown Algal Genus Cystophora. Aust. J. Chem. 1981, 34, 439–447. [Google Scholar] [CrossRef]
- Amico, V.; Currenti, R.; Oriente, G.; Piatelli, M.; Tringali, C. A phloroglucinol derivative from the brown alga Zonaria tournefortii. Phytochemistry 1981, 20, 1451–1453. [Google Scholar] [CrossRef]
- Kim, S.-K.; Pangestuti, R. Biological activities and potential health benefits of fucoxanthin derived from marine brown algae. Adv. Food Nutr. Res. 2011, 64, 111–128. [Google Scholar] [PubMed]
- Tan, C.-P.; Hou, Y.-H. First evidence for the anti-inflammatory activity of fucoxanthin in high-fat-diet-induced obesity in mice and the antioxidant functions in PC12 cells. Inflammation 2014, 37, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-N.; Heo, S.-J.; Kang, S.-M.; Ahn, G.; Jeon, Y.-J. Fucoxanthin induces apoptosis in human leukemia HL-60 cells through a ROS-mediated Bcl-xL pathway. Toxicol. Vitro 2010, 24, 1648–1654. [Google Scholar] [CrossRef]
- Xia, S.; Wang, K.; Wan, L.; Li, A.; Hu, Q.; Zhang, C. Production, charaterization, and antioxidant activity of fucoxanthin from the marine diatom Odontella aurita. Mar. Drugs 2013, 11, 2667–2681. [Google Scholar] [CrossRef] [PubMed]
- Wisespongpand, P.; Kuniyoshi, M. Bioactive phloroglucinols from the brown alga Zonaria diesingiana. J. Appl. Phycol. 2003, 15, 225–228. [Google Scholar] [CrossRef]
- Gerwick, W.; Fenical, W. Phenolic lipids from the related marine algae of the order Dictyotales. Phytochemistry 1982, 21, 633–637. [Google Scholar] [CrossRef]
- Ravi, B.N.; Murphy, P.T.; Lidgard, R.O.; Warren, R.G.; Wells, R.J. C18 terpenoid metabolites of the brown alga Cystophora moniliformis. Aust. J. Chem. 1982, 35, 171–182. [Google Scholar] [CrossRef]
- Brkljača, R.; Urban, S. Chemical profiling (HPLC-NMR & HPLC-MS), isolation, and identification of bioactive meroditerpenoids from the southern Australian marine brown alga Sargassum paradoxum. Mar. Drugs 2015, 13, 102–127. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brkljača, R.; Gӧker, E.S.; Urban, S. Dereplication and Chemotaxonomical Studies of Marine Algae of the Ochrophyta and Rhodophyta Phyla. Mar. Drugs 2015, 13, 2714-2731. https://doi.org/10.3390/md13052714
Brkljača R, Gӧker ES, Urban S. Dereplication and Chemotaxonomical Studies of Marine Algae of the Ochrophyta and Rhodophyta Phyla. Marine Drugs. 2015; 13(5):2714-2731. https://doi.org/10.3390/md13052714
Chicago/Turabian StyleBrkljača, Robert, Emrehan Semih Gӧker, and Sylvia Urban. 2015. "Dereplication and Chemotaxonomical Studies of Marine Algae of the Ochrophyta and Rhodophyta Phyla" Marine Drugs 13, no. 5: 2714-2731. https://doi.org/10.3390/md13052714