
Advances in Microalgae-Derived Phytosterols for Functional Food and Pharmaceutical Applications

Abstract
:1. Phytosterols: Chemistry, Origin and Applications
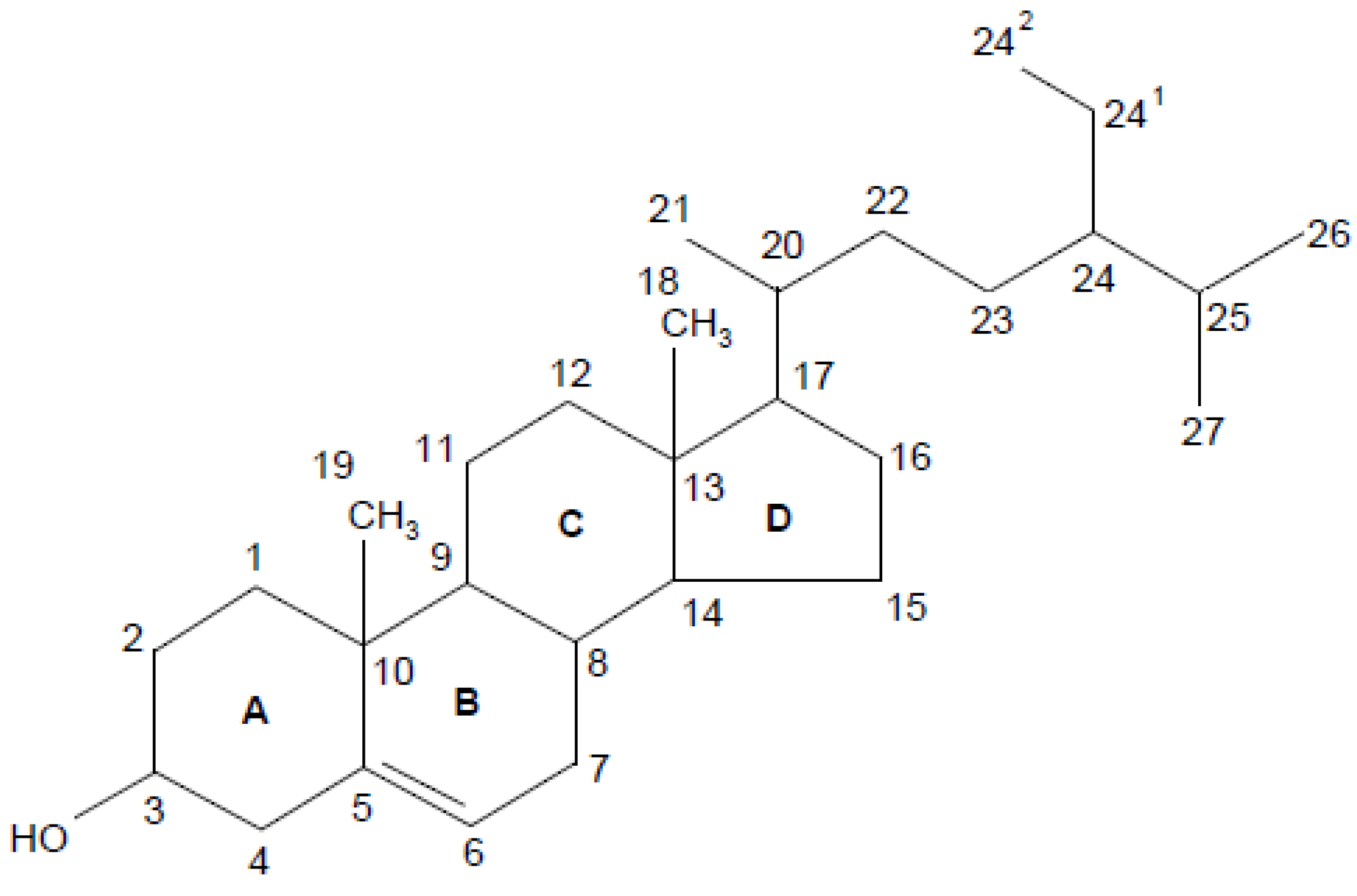
1.1. Chemistry of Phytosterols

1.2. Origin and Applications of Phytosterols
1.3. Health Promoting Effects of Phytosterol and Its Regulations
2. Microalgae as a Potential Source of Phytosterols
2.1. Types of Phytosterols from Microalgae for Human Consumption

{kind=link}
| Species | Identified Phytosterols | References |
|---|---|---|
| Attheya ussurensis sp. nov. | 24-Ethylcholest-5-en-3β-ol | [27] |
| Bigelowiella | Crinosterol, Stigmasterol | [34] |
| Chattonella antique | Isofucosterol | [41] |
| Chattonella marina | Isofucosterol | [41] |
| Chattonella subsalsa | Isofucosterol | [41] |
| Chlorella vulgaris | Ergosterol, 7-Dehydroporiferasterol, Ergosterol peroxide, 7-Oxocholesterol | [42] |
| Chrysoderma sp. | Stigmasterol, Sitosterol, Fucosterol | [38] |
| Chrysomeris | Stigmasterol, Sitosterol, Fucosterol | [38] |
| Chrysowaernella | Stigmasterol, Sitosterol, Fucosterol | [38] |
| Crypthecodinium cohnii | 4α-Methyl sterols, Dinosterols, Dehydrodinosterol 4α,24-Dimethyl-cholestan-3β-ol 4α,24-Dimethylcholest-5-en-3β-ol Cholesta-5,7-dien-3β-ol | [43] |
| Cyanophora paradoxa | Sitosterol, Campesterol and 24-Ethylcholesta-5,22E-dien-3β-ol | [28] |
| Diacronema vlkianum | 24-Ethylcholesta-5,22E-dien-3β-ol 4α-Methyl-24-ethyl-5α-cholest-22E-en-3β-ol | [30] |
| Dunaliella salina | Ergosterol, 7-Dehydroporiferasterol, 7-Dehydroporiferasterol peroxide, Ergosterol peroxide | [44,45] |
| Dunaliella tertiolecta | Ergosterol, 7-Dehydroporiferasterol | [44,46] |
| Fragilaria pinnata | 23,24-Dimethylcholesta-5,22E-dien-3β-ol | [47] |
| Giraudyopsis | Stigmasterol, Sitosterol, Fucosterol | [38] |
| Glaucocystis nostochinearum | Sitosterol, Campesterol, 24-Ethylcholesta-5,22E-dien-3β-ol | [28] |
| Gymnochlora | Crinosterol, Stigmasterol | [34] |
| Isochrysis galbana | 24-Oxocholesterol acetate, Ergost-5-en-3β-ol, Cholest-5-en-24-1,3-(acetyloxy)-,3β-ol | [48] |
| Karenia brevis | 27-Nor-(24R)-4α-methyl-5α-ergosta-8(14),22-dien-3β-ol Brevesterol (24S)-4α-Methyl-5α-ergosta-8(14),22-dien-3β, its 27-Nor derivative | [49,50] |
| Karenia mikimotoi | 27-Nor-(24R)-4α-methyl-5α-ergosta-8(14),22-dien-3β-ol Brevesterol, Gymnodinosterol (24R)-4α-Methyl-5α-ergosta-8(14),22-dien-3β-ol | [49] |
| Karenia papilionacea | 23-Methyl-27-norergosta-8(14),22-dien-3β-ol | [49] |
| Karenia umbella | (24R)-4α-Methyl-5α-ergosta-8(14),22-dien-3β-ol Gymnodinosterol | [49] |
| Karlodinium veneficum | (24R)-4α-Methyl-5α-ergosta-8(14),22-dien-3β-ol Gymnodinosterol | [49] |
| Lotharella | Crinosterol and Stigmasterol | [34] |
| Micromonas aff.pusilla | 24-Methycholesta-5,24(28)-dien-3β-ol 24-Methylcholesta-5-en-3β-ol 28-Isofucosterol and saringosterol | [51] |
| Micromonas pusilla | 24-Methycholesta-5,24(28)-dien-3β-ol 24-Methylcholesta-5-en-3β-ol 28-Isofucosterol | [51] |
| Navicula incerta | Stigmasterol, 5β-Hydroxysitostanol | [52,53] |
| Nematochrysopsis sp. | (24E)-24-n-propylidenecholesterol | [38] |
| Nitzschia closterium | Cholesta-5,24-dien-3β-ol 24-Methylcholesta-5,22E-dien-3β-ol | [47] |
| Nostoc commune var. sphaeroides Kützing | Campesterol, Sitosterol, Clionasterol | [54,55] |
| Olisthodiscus luteus | Brassicasterol, Stigmasterol, Fucosterol | [41] |
| Pavlova | 24-Ethylcholesta-5,22E-dien-3β-ol 4α-Methyl-24-ethyl-5α-cholest-22E-en-3β-ol | [30] |
| Phaeodactylum tricornutum | (24S)-24-Methylcholesta-5,22E-dien-3β-ol | [39] |
| Polarella glacialis | 27-Nor-24-Methylcholest-5,22E-dien-3β-ol | [35] |
| Porphyridium cruentum | Stigmasterol, β-Sitosterol | [56] |
| Pycnococcus provasolii | 24-Methycholesta-5,24(28)-dien-3β-ol 24-Methylcholesta-5-en-3β-ol 28-Isofucosterol | [51] |
| Pyramimonas cf. cordata | Stigmasterol | [27] |
| Pyramimonas cordata | 24-Methycholesta-5,24(28)-dien-3β-ol 24-Methylcholesta-5-en-3β-ol 28-Isofucosterol | [51] |
| Rhizosolenia setigera | Cholesta-5,24-dien-3β-ol | [47] |
| Sarcinochrysis sp. | (24E)-24-n-propylidenecholesterol | [38] |
| Schizochytrium aggregatum | Campesterol, 24-Methylene cholesterol, Ergosterol, 24-Methyl-colest-7-en-3β-ol, Stigmasterol and others | [57] |
| Schizochytrium sp. | Lathosterol, Ergosterol, Stigmasterol, 24-Ethylcholesta-5,7,22-trienol, Stigmasta-7,24-(241)-dien-3β-ol, | [58] |
| Stephanodiscus meyerii | 24-Methycholesta-5,24(28)-dien-3β-ol | [27] |
| Takayama helix | 27-Nor-(24R)-4α-methyl-5α-ergosta-8(14),22-dien-3β-ol Brevesterol | [49] |
| Takayama tasmanica | 27-Nor-(24R)-4α-methyl-5α-ergosta-8(14),22-dien-3β-ol Brevesterol | [49] |
| Tetraselmis chui | 24-Methycholesta-5,24(28)-dien-3β-ol 24-Methylcholesta-5-en-3β-ol 28-Isofucosterol | [51] |
| Tetraselmis suecica | 24-Methylcholest-5-en-3β-ol 24-Methylcholest-5,24(28)-dien-3β-ol | [39] |
| Thalassi-onema nitzschioides | 23-Methylcholesta-5,22E-dien-3β-ol 23-Methyl-5α-cholest-22E-en-3β-ol | [47] |
2.2. Biosynthesis of Phytosterols in Microalgae
2.3. Bio-Functionalities of Microalgal Phytosterols and Their Mechanisms of Action
2.3.1. Cholesterol-Lowering Activity
2.3.2. Anti-Inflammatory Activity
2.3.3. Anticancer Activity
2.3.4. Antioxidant
2.3.5. Other Activities
| Microalgae Species | Major Phytosterols | Biological Activity | Function | References |
|---|---|---|---|---|
| Chlorella vulgaris | Ergosterol, 7-Dehydroporiferasterol, Ergosterol peroxide, 7-Dehydroporiferasterol peroxide, 7-oxocholesterol | Anti-inflammatory | 50% inhibitory dose was 0.2–0.7 mg/ear | [42] |
| Chlorella vulgaris | Ergosterol peroxide | Anti-cancer | 2 μmol led to 77% reduction in tumour progression | [42] |
| Dunaliella tertiolecta | Ergosterol, 7-Dehydroporiferasterol | Immunomodulatory Anti-inflammatory | 0.4 mg/mL mixture for the highest production of IL-10, 0.8mg/mL for ergosterol alone | [46] |
| Dunaliella tertiolecta | Ergosterol, 7-Dehydroporiferasterol | Neuromodulatory | Neuromodulatory action was found in selective brain areas of rats | [80] |
| Isochrysis galbana | 24-Oxocholesterol acetate, Ergost-5-en-3β-ol, Cholest-5-en-24-1,3-(acetyloxy)-, 3β-ol and others | Antituberculosis | Minimum inhibitory concentration of 50–60 μg/mL against M. tuberculosis | [48] |
| Navicula incerta | Stigmasterol, 5β-Hydroxysitostanol | Anti-cancer | 40%, 43% and 54% toxicity at 5, 10 and 20 μM, respectively | [52,53] |
| Nostoc commune var. sphaeroides Kützing | Lipid extract | Cholesterol-lowering activity | Reduced HMGR activity by 90% and reduced SREBP-1 mature protein by 30% | [54] |
| Schizochytrium aggregatum | Campesterol, 24-Methylene cholesterol, 24-Methyl-colest-7-en-3β-ol, Ergosterol, Stigmasterol and other lipids | Antioxidant | IC50 in DPPH radical scavenging study was 5.76 mg/mL. Digested microalgae oil had an α-tocopherol equivalent antioxidant capacity of 42.071 μg/mg At 10 mg/mL, reducing power was 0.874 | [57] |
| Schizochytrium sp. | Lathosterol, Ergosterol, Stigmasterol, 24-Ethylcholesta-5,7,22-trienol, Stigmasta-7,24-(241)-dien-3β-ol and others | Cholesterol-lowering activity | 0.06–0.3 g/kg diet decreased blood cholesterol by 19.5%–34% | [58] |
| Chemical Structure | Nomenclatures | Species of Origin | Bioactivity | Same Sterol(s) Observed in Microalgae | |
|---|---|---|---|---|---|
| Campesterol |  | Campesterin Cholest5-en-3-ol (24R)-24-Methylcholest-5-en-3β-ol Campest-5-en-3β-ol Δ5-24α-Methyl-cholesten-3β-ol (24R)-Ergost-5-en-3β-ol | Flower Chrysanthemum coronarium L. [66] Red algae Porphyra dentata [81] Shorea singkawang [82] | Cholesterol-lowering Anticancer Antiangiogenic | Tetraselmis [33] Porphyridium cruentum [83] Schizochytrium aggregatum [57] |
| 7-Dehydroporiferasterol | 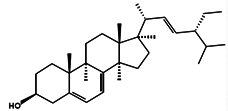 | (22E,24R)-Ethylcholesta-5,7,22-trien-3β-ol 24R-Stigmasta-5,7,22-trien-3β-ol Porifersta-5,7,22E-trienol | Rarely found in other organisms | - | Chlorella vulgaris [84] Chlamydomonas reinhardtii [85] Dictyonella incisa [86] |
| Ergosterol |  | (22E)-Ergosta-5,7,22-trien-3β-ol (22E,24R)-Methylcholesta-5,7,22-trien-3β-ol | Mushroom Sarcodon aspratus [72] Mushroom Inonotus obliquus [87] Ganoderma lucidum [88] Agaricus bisporus [89] | Anticancer Anti-inflammatory Cholesterol-lowering | Chlorella pyranoidosa [90] Dunaliella tertiolecta [80] Schizochytrium aggregatum [57] |
| Fucosterol |  | (24(28)E)-Stigmasta-5,24(28)-dien-3β-ol (24(24′)E)-Stigmasta-5,24(24’)-dien-3β-ol (24E)-Ethylidenecholesta-5,24(28)-dien-3β-ol | Macroalgae Pelvetia siliquosa [64,67] Brown alga Turbinaria conoides [65] Macroalgae Himanthalia elongate, Undaria pinnatifida, Phorphyra sp., Chondus crispus, Cystoseira sp. and Ulva sp. [91] | Antioxidant Antidiabetic Anticancer Cholesterol-lowering | Chrysoderma sp. Chrysomeris Chrysowaernella Giraudyopsis [38] Olisthodiscus luteus [41] |
| Saringosterol | 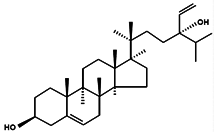 | 24(S)-Saringosterol Sargasso sterol | Brown algae Sargassum ringgoldianum [79] Sargassum thunbergii [92] Lessonia nigrescens [93] Seaweed Sargassum fusiforme [94] | Antitubercular Antiatherosclerotic Lipase-inhibitory | Micromonas aff.pusilla [51] |
| β-Sitosterol | 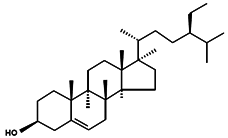 | Sitosterol Stigmast-5-en-3β-ol 24α-Ethylcholest-5-en-3β-ol | Peanuts [95] Coral subergorgia reticulate [96] Plant Verbena officinalis [97] Leaves of Mentha cordifolia Opiz [98] | Anticancer Anti-inflammatory Analgesic activity Anthelminthic Antimutagenic | Bigelowiella natans Gymnochlora stellata Lotharella amoeboformis [34] Porphyridium cruentum [56] |
| Stigmasterol | 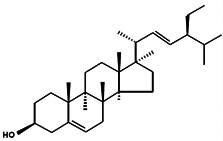 | (Δ5,22E) (24α=24S) Poriferasterol (22E)-Stigmasta-5,22-dien-3β-ol 24α-Ethylcholesta-5,22E-dien-3β-ol | Butea monosperma [76] Parkia speciosa seeds [99] | Thyroid-inhibitory Antioxidant Hypoglycaemic | Porphyridium cruentum [56] |
| Δ5-Avenasterol | 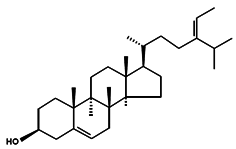 | (5-Avenasterol) (Δ5,24Z) Isofucosterol 28-Isofucosterol 29-Iso-fucosterol 24Z-Ethylidenecholesta-5,24(28)-dien-3β-ol [24(28)Z]-Stigmasta-5,24(28)-dien-3β-ol [24(24′)Z]-Stigmasta-5,24(28’)-dien-3β-ol | Brown algae Fucus vesiculosus Green algae Ulva lactuca [100] Wheat germ oil [101] Tomato seed oil [102] Sargassum thunbergii [92] Rape bee pollen [103] Marine sponge Petrosia weinbergi [104] | Antioxidant Lipase-inhibitory Precursor of antiviral orthoesterol | Myxophyceae
Chlorophyceae [55] Chattonella marina [41] |
| Brassicasterol | 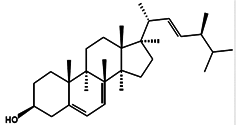 | 24-Methyl cholest-5,22-dien-3β-ol (3β,22E)-Ergosta-5,22-dien-3-ol Ergosta-5,22-dien-3β-ol | Rapeseed oil [105] | Cholesterol-lowering | Isochrysis galbana and Chaetoceros calcitrans [106] Rhodomonas salina [107] |
2.4. Advanced Green Extraction and Purification Technology of Phytosterols
3. Future Prospects of Microalgae-Derived Phytosterols
4. Conclusions
Acknowledgments
Author Contribution
Conflicts of Interest
References
- Moss, G.P. Nomenclature of steroids (Recommendations 1989). Pure Appl. Chem. 1989, 61, 1783–1822. [Google Scholar] [CrossRef]
- Trautwein, E.A.; Demonty, I. Phytosterols: natural compounds with established and emerging health benefits. Ol. Corps Gras Lipides 2007, 14, 259–266. [Google Scholar] [CrossRef]
- Volkman, J. Sterols in microorganisms. Appl. Microbiol. Biotechnol. 2003, 60, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Moreau, R.A.; Whitaker, B.D.; Hicks, K.B. Phytosterols, phytostanols, and their conjugates in foods: Structural diversity, quantitative analysis, and health-promoting uses. Prog. Lipid Res. 2002, 41, 457–500. [Google Scholar] [CrossRef]
- Goad, J.L. Methods in Plant Biochemistry; Academic Press: London, UK, 1991; pp. 369–434. [Google Scholar]
- Hernandez-Ledesma, B.; Herrero, M. Bioactive Compounds from Marine Foods: Plant and Animal Sources, 1st ed.; John Wiley & Sons Ltd: Chichester, UK, 2014; pp. 173–187. [Google Scholar]
- Piironen, V.; Lindsay, D.G.; Miettinen, T.A.; Toivo, J.; Lampi, A.M. Plant sterols: Biosynthesis, biological function and their importance to human nutrition. J.Sci. Food Agric. 2000, 80, 939–966. [Google Scholar] [CrossRef]
- Geçgel, M.T.B.B.Ü.; Demirci, A. Phytosterols as functional food ingredients. J. Tekirdag Agric. Fac. 2006, 3, 153–159. [Google Scholar]
- Hicks, K.B.; Moreau, R.A. Phytosterols and phytostanols: Functional food cholesterol busters. Food Technol. 2001, 55, 63–67. [Google Scholar]
- Cerqueira, M.T.; Fry, M.M.; Connor, W.E. The food and nutrient intakes of the Tarahumara Indians of Mexico. Am. J. Clin. Nutr. 1979, 32, 905–915. [Google Scholar] [PubMed]
- Fernandes, P.; Cabral, J.M.S. Phytosterols: Applications and recovery methods. Bioresour. Technol. 2007, 98, 2335–2350. [Google Scholar] [CrossRef] [PubMed]
- Pollak, O.J. Reduction of blood cholesterol in man. Circulation 1953, 7, 702–706. [Google Scholar] [CrossRef] [PubMed]
- Miettinen, T.; Vanhanen, H.; Wester, I. Use of a Stanol Fatty Acid Ester for Reducing Serum Cholesterol Level. U.S. Patent 5,502,045, 26 March 1996. [Google Scholar]
- Tan, T.; Zhang, M.; Gao, H. Ergosterol production by fed-batch fermentation of Saccharomyces cerevisiae. Enzym. Microb. Technol. 2003, 33, 366–370. [Google Scholar] [CrossRef]
- Global Phytosterols Market By Application (Pharmaceuticals, Cosmetics, Food Ingredients), By Product (Beta-sitosterol, Campesterol, Stigmasterol) is Expected to Reach USD 989.8 Million by 2020: Grand View Research, Inc. Available online: http://www.grandviewresearch.com/press-release/global-phytosterols-market (accessed on 6 July 2015).
- FDA. 65 FR 54686—Food Labeling: Health Claims; Plant Sterols/Stanol Esters and Coronary Heart Disease; Federal FR Doc No: 00-22892; Office of the Federal Register, National Archives and Records Administration: Washington, DC, USA, 2000.
- Srigley, C.T.; Haile, E.A. Quantification of plant sterols/stanols in foods and dietary supplements containing added phytosterols. J. Food Compos. Anal. 2015, 40, 163–176. [Google Scholar] [CrossRef]
- Gylling, H.; Plat, J.; Turley, S.; Ginsberg, H.N.; Ellegård, L.; Jessup, W.; Jones, P.J.; Lütjohann, D.; Maerz, W.; Masana, L. Plant sterols and plant stanols in the management of dyslipidaemia and prevention of cardiovascular disease. Atherosclerosis 2014, 232, 346–360. [Google Scholar] [CrossRef] [PubMed]
- Piironen, V.; Toivo, J.; Lampi, A.M. Natural sources of dietary plant sterols. J. Food Compos. Anal. 2000, 13, 619–624. [Google Scholar] [CrossRef]
- Ryckebosch, E.; Bruneel, C.; Termote-Verhalle, R.; Muylaert, K.; Foubert, I. Influence of extraction solvent system on extractability of lipid components from different microalgae species. Algal Res. 2014, 3, 36–43. [Google Scholar] [CrossRef]
- Ahmed, F.; Zhou, W.; Schenk, P.M. Pavlova lutheri is a high-level producer of phytosterols. Algal Res. 2015, 10, 210–217. [Google Scholar] [CrossRef]
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Demirbas, A.M.; Fatih, D.M. Importance of algae oil as a source of biodiesel. Energy Convers. Manag. 2011, 52, 163–170. [Google Scholar] [CrossRef]
- Brown, M.R. Nutritional Value and Use of Microalgae in Aquaculture. In Proceedings of the Avances en Nutrición Acuícola VI. Memorias del VI Simposium Internacional de Nutrición Acuícola, Cancún, Quintana Roo, México, 3–6 September, 2002; pp. 281–292.
- Tang, G.; Suter, P.M. Vitamin A, nutrition, and health values of algae: Spirulina, Chlorella, and Dunaliella. J. Pharm. Nutr. Sci. 2011, 1, 111–118. [Google Scholar] [CrossRef] [Green Version]
- QuÍlez, J.; GarcÍa-Lorda, P.; Salas-Salvadó, J. Potential uses and benefits of phytosterols in diet: Present situation and future directions. Clin. Nutr. 2003, 22, 343–351. [Google Scholar] [CrossRef]
- Ponomarenko, L.P.; Stonik, I.V.; Aizdaicher, N.A.; Orlova, T.Y.; Popovskaya, G.I.; Pomazkina, G.V.; Stonik, V.A. Sterols of marine microalgae Pyramimonas cf. cordata (Prasinophyta), Attheya ussurensis sp. nov. (Bacillariophyta) and a spring diatom bloom from Lake Baikal. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2004, 138, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Leblond, J.D.; Timofte, H.I.; Roche, S.A.; Porter, N.M. Sterols of glaucocystophytes. Phycol. Res. 2011, 59, 129–134. [Google Scholar] [CrossRef]
- Goad, L.J.; Holz, G.G., Jr.; Beach, D.H. Identification of (24S)-24-methylcholesta-5,22-dien-3β-ol as the major sterol of a marine cryptophyte and a marine prymnesiophyte. Phytochemistry 1983, 22, 475–476. [Google Scholar] [CrossRef]
- Volkman, J.K.; Farmen, C.L.; Barrett, S.M.; Sikes, E.L. Unusual dihydroxysterols as chemotaxonoic markers for microalgae from the order Pavlovales (Haptophyceae). J. Phycol. 1997, 33, 1016–1023. [Google Scholar] [CrossRef]
- Giner, J.L.; Zhao, H.; Boyer, G.L.; Satchwell, M.F.; Andersen, R.A. Sterol chemotaxonomy of marine Pelagophyte algae. Chem. Biodivers. 2009, 6, 1111–1130. [Google Scholar] [CrossRef] [PubMed]
- Rampen, S.W.; Schouten, S.; Hopmans, E.C.; Abbas, B.; Noordeloos, A.A.M.; van Bleijswijk, J.D.L.; Geenevasen, J.A.J.; Sinninghe, D.J.S. Diatoms as a source for 4-desmethyl-23,24-dimethyl steroids in sediments and petroleum. Geochim. Cosmochim. Acta 2009, 73, 377–387. [Google Scholar] [CrossRef]
- Patterson, G.W.; Tsitsa-Tzardis, E.; Wikfors, G.H.; Gladu, P.K.; Chitwood, D.J.; Harrison, D. Sterols of Tetraselmis (Prasinophyceae). Comp. Biochem. Physiol. B Comp. Biochem. 1993, 105, 253–256. [Google Scholar] [CrossRef]
- Leblond, J.D.; Dahmen, J.L.; Seipelt, R.L.; Elrod-Erickson, M.J.; Kincaid, R.; Howard, J.C.; Evens, T.J.; Chapman, P.J. Lipid composition of chlorarachniophytes (Chlorarachniophyceae) from the Genera Bigelowiella, Gymnochlora, and Lotharella. J. Phycol. 2005, 41, 311–321. [Google Scholar] [CrossRef]
- Thomson, P.G.; Wright, S.W.; Bolch, C.J.S.; Nichols, P.D.; Skerratt, J.H.; McMinn, A. Antarctic distribution, pigment and lipid composition, and molecular identification of the brine dinoflagellate Polarella glacialis (Dinophyceae). J. Phycol. 2004, 40, 867–873. [Google Scholar] [CrossRef]
- Kumari, P.; Kumar, M.; Reddy, C.R.K.; Jha, B. Algal Lipids Fat. Acids Sterols; Woodhead Publishing Limited: Cambridge, UK, 2013. [Google Scholar]
- Volkman, J.; Rijpstra, W.I.C.; de Leeuw, J.W.; Mansour, M.P.; Jackson, A.E.; Blackburn, S.I. Sterols of four dinoflagellates from the genus Prorocentrum. Phytochemistry 1999, 52, 659–668. [Google Scholar] [CrossRef]
- Billard, G.; Dauguet, J.C.; Maume, D.; Bert, M. Sterols and chemotaxonomy of marine Chrysophyceae. Bot. Mar. 1990, 33, 225–228. [Google Scholar] [CrossRef]
- Fábregas, J.; Arán, J.; Morales, E.D.; Lamela, T.; Otero, A. Modification of sterol concentration in marine microalgae. Phytochemistry 1997, 46, 1189–1191. [Google Scholar] [CrossRef]
- Piretti, M.V.; Pagliuca, G.; Boni, L.; Pistocchi, R.; Diamante, M.; Gazzotti, T. Investigating of 4-methyl sterols from cultured dinoflagellate algal strains. J. Phycol. 1997, 33, 61–67. [Google Scholar] [CrossRef]
- Marshall, J.A.; Nichols, P.; Hallegraeff, G. Chemotaxonomic survey of sterols and fatty acids in six marine raphidophyte algae. J. Appl. Phycol. 2002, 14, 255–265. [Google Scholar] [CrossRef]
- Yasukawa, K.; Akihisa, T.; Kanno, H.; Kaminaga, T.; Izumida, M.; Sakoh, T.; Tamura, T.; Takido, M. Inhibitory effects of sterols isolated from Chlorella vulgaris on 12-O-tetradecanoylphorbol-13-acetate-Induced inflammation and tumor promotion in mouse skin. Biol. Pharm. Bull. 1996, 19, 573–576. [Google Scholar] [CrossRef] [PubMed]
- Mendes, A.; Reis, A.; Vasconcelos, R.; Guerra, P.; da Silva, T.L. Crypthecodinium cohnii with emphasis on DHA production: A review. J. Appl. Phycol. 2009, 21, 199–214. [Google Scholar] [CrossRef]
- Francavilla, M.; Trotta, P.; Luque, R. Phytosterols from Dunaliella tertiolecta and Dunaliella salina: A potentially novel industrial application. Bioresour. Technol. 2010, 101, 4144–4150. [Google Scholar] [CrossRef]
- Sheffer, M.; Fried, A.; Gottlieb, H.E.; Tietz, A.; Avron, M. Lipid composition of the plasma-membrane of the halotolerant alga, Dunaliella salina. Biochim. Biophys. Acta 1986, 857, 165–172. [Google Scholar] [CrossRef]
- Caroprese, M.; Albenzio, M.; Ciliberti, M.G.; Francavilla, M.; Sevi, A. A mixture of phytosterols from Dunaliella tertiolecta affects proliferation of peripheral blood mononuclear cells and cytokine production in sheep. Vet. Immunol. Immunopathol. 2012, 150, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Barrett, S.M.; Volkman, J.K.; Dunstan, G.A.; LeRoi, J.M. Sterols of 14 species of marine diatoms (bacillariophyta). J. Phycol. 1995, 31, 360–369. [Google Scholar] [CrossRef]
- Prakash, S.; Sasikala, S.; Aldous, V.H.J. Isolation and identification of MDR-Mycobacterium tuberculosis and screening of partially characterised antimycobacterial compounds from chosen marine micro algae. Asian Pac. J. Trop. Med. 2010, 3, 655–661. [Google Scholar] [CrossRef]
- Mooney, B.D.; Nichols, P.D.; de Salas, M.F.; Hallegraeff, G.M. Lipid, fatty acid, and sterol composition of eight species of kareniaceae (Dinophyta): Chemotaxonomy and putative lipid phycotoxin. J. Phycol. 2007, 43, 101–111. [Google Scholar] [CrossRef]
- Leblond, J.D.; Evans, T.J.; Chapman, P.J. The biochemistry of dinoflagellate lipids, with particular reference to the fatty acid and sterol composition of a Karenia brevis bloom. Phycologia 2003, 42, 324–331. [Google Scholar] [CrossRef]
- Volkman, J.K.; Barrett, S.M.; Dunstan, G.A.; Jeffrey, S.W. Sterol biomarkers for microalgae from the green algal class Prasinophyceae. Org. Geochem. 1994, 21, 1211–1218. [Google Scholar] [CrossRef]
- Kim, Y.S.; Li, X.F.; Kang, K.H.; Ryu, B.; Kim, S.K. Stigmasterol isolated from marine microalgae Navicula incerta induces apoptosis in human hepatoma HepG2 cells. BMB Rep. 2014, 47, 433. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; KANG, K.H.; Kim, Y.S. Pharmaceutical Composition for Preventing or Treating Liver Cancer Comprising Stigmasterol and 5 beta-hydroxysitostanol Isolated from Navicula incerta. U.S. Patent 2014/0057884 A1, 27 February 2014. [Google Scholar]
- Rasmussen, H.E.; Blobaum, K.R.; Park, Y.K.; Ehlers, S.J.; Lu, F.; Lee, J.Y. Lipid extract of Nostoc commune var. sphaeroides Kützing, a blue-green alga, inhibits the activation of sterol regulatory element binding proteins in HepG2 Cells. J. Nutr. 2008, 138, 476–481. [Google Scholar]
- Paoletti, C.; Pushparaj, B.; Florenzano, G.; Capella, P.; Lercker, G. Unsaponifiable matter of green and blue-green algal lipids as a factor of biochemical differentiation of their biomasses: II. Terpenic alcohol and sterol fractions. Lipids 1976, 11, 266–271. [Google Scholar] [CrossRef]
- Durmaz, Y.; Monteiro, M.; Koru, E.; Bandarra, N. Concentration of sterols of Porphyridium cruentum biomass at stationary phase. Pak. J. Biol. Sci. 2007, 10, 1144–1146. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Yang, X.; Ma, H.; Hu, X.; Wei, Y.; Zhou, W.; Li, L. The oxidative stability of microalgae oil (Schizochytrium aggregatum) and its antioxidant ability after gastrointestinal digestion: relationship with constituents. Eur. J. Lipid Sci. Technol. 2015. [Google Scholar] [CrossRef]
- Chen, J.; Jiao, R.; Jiang, Y.; Bi, Y.; Chen, Z.Y. Algal Sterols are as effective as β-sitosterol in reducing plasma cholesterol concentration. J. Agric. Food Chem. 2014, 62, 675–681. [Google Scholar] [CrossRef] [PubMed]
- Spanova, M.; Daum, G. Squalene-biochemistry, molecular biology, process biotechnology, and applications. Eur. J. Lipid Sci. Technol. 2011, 113, 1299–1320. [Google Scholar] [CrossRef]
- Lohr, M.; Schwender, J.; Polle, J.E.W. Isoprenoid biosynthesis in eukaryotic phototrophs: A spotlight on algae. Plant Sci. 2012, 185–186, 9–22. [Google Scholar]
- Kim, D.; Filtz, M.R.; Proteau, P.J. The Methylerythritol phosphate pathway contributes to carotenoid but not phytol biosynthesis in Euglena gracilis. J. Nat. Prod. 2004, 67, 1067–1069. [Google Scholar] [CrossRef] [PubMed]
- Grauvogel, C.; Petersen, J. Isoprenoid biosynthesis authenticates the classification of the green alga Mesostigma viride as an ancient streptophyte. Gene 2007, 396, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Disch, A.; Schwender, J.; Müller, C.; Lichtenthaler, H.K.; Rohmer, M. Distribution of the mevalonate and glyceraldehyde phosphate/pyruvate pathways for isoprenoid biosynthesis in unicellular algae and the cyanobacterium Synechocystis PCC 6714. Biochem. J. 1998, 333, 381–388. [Google Scholar] [PubMed]
- Lee, S.; Lee, Y.S.; Jung, S.H.; Kang, S.S.; Shin, K.H. Anti-oxidant activities of fucosterol from the marine algae Pelvetia siliquosa. Arch. Pharm. Res. 2003, 26, 719–722. [Google Scholar] [CrossRef] [PubMed]
- Sheu, J.H.; Wang, G.H.; Sung, P.J.; Duh, C.Y. New Cytotoxic oxygenated fucosterols from the brown alga Turbinaria conoides. J. Nat. Prod. 1999, 62, 224–227. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.M.; Lee, E.O.; Lee, H.J.; Kim, K.H.; Ahn, K.S.; Shim, B.S.; Kim, N.I.; Song, M.C.; Baek, N.I.; Kim, S.H. Identification of campesterol from Chrysanthemum coronarium L. and its antiangiogenic activities. Phytother. Res. 2007, 21, 954–959. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Shin, K.H.; Kim, B.K.; Lee, S. Anti-Diabetic activities of fucosterol from Pelvetia siliquosa. Arch. Pharm. Res. 2004, 27, 1120–1122. [Google Scholar] [CrossRef] [PubMed]
- Neil, H.; Meijer, G.; Roe, L. Randomised controlled trial of use by hypercholesterolaemic patients of a vegetable oil sterol-enriched fat spread. Atherosclerosis 2001, 156, 329–337. [Google Scholar] [CrossRef]
- Nigon, F.; Serfaty-Lacrosnière, C.; Beucler, I.; Chauvois, D.; Neveu, C.; Giral, P.; Chapman, M.J.; Bruckert, E. Plant sterol-enriched margarine lowers plasma LDL in hyperlipidemic subjects with low cholesterol intake: Effect of fibrate treatment. Clin. Chem. Lab. Med. 2001, 39, 634–640. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, A.K.; Lopez, L.A. Absorbability of plant sterols and their distribution in rabbit tissues. Biochim. Biophys. Acta 1979, 574, 146–153. [Google Scholar] [CrossRef]
- Paoletti, C.; Pushparaj, B.; Florenzano, G.; Capella, P.; Lercker, G. Unsaponifiable matter of green and blue-green algal lipids as a factor of biochemical differentiation of their biomasses: I. Total unsaponifiable and hydrocarbon fraction. Lipids 1976, 11, 258–265. [Google Scholar] [CrossRef]
- Kobori, M.; Yoshida, M.; Ohnishi-Kameyama, M.; Shinmoto, H. Ergosterol peroxide from an edible mushroom suppresses inflammatory responses in RAW264.7 macrophages and growth of HT29 colon adenocarcinoma cells. Br. J. Pharm. 2007, 150, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Awad, A.B.; Fink, C.S. Phytosterols as Anticancer Dietary Components: Evidence and mechanism of action. J. Nutr. 2000, 130, 2127–2130. [Google Scholar] [PubMed]
- Khalos, K.; Kangas, L.; Hiltunen, R. Ergosterol peroxide, an active compound from Inonotus radiatus. Planta Medica 1989, 55, 389–390. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.F.; Yi, Y.H.; Yao, X.S.; Xu, Q.Z.; Zhang, S.Y.; Lin, H.W. Bioactive steroids from the brown alga Sargassum carpophyllum. J. Asian Nat. Prod. Res. 2002, 4, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.; Jafri, M.; Kar, A.; Meheta, B.K. Thyroid inhibitory, antiperoxidative and hypoglycemic effects of stigmasterol isolated from Butea monosperma. Fitoterapia 2009, 80, 123–126. [Google Scholar] [CrossRef] [PubMed]
- Park, P.J.; Jung, W.K.; Nam, K.S.; Shahidi, F.; Kim, S.K. Purification and characterization of antioxidative peptides from protein hydrolysate of lecithin-free egg yolk. J. Am. Oil Chem. Soc. 2001, 78, 651–656. [Google Scholar] [CrossRef]
- WHO. Global Tuberculosis Report 2014; World Health Organisation: Geneva, Swizerland, 2014. [Google Scholar]
- Ikekawa, N.; Morisaki, N.; Tsuda, K.; Yoshida, T. Sterol compositions in some green algae and brown algae. Steroids 1968, 12, 41–48. [Google Scholar] [CrossRef]
- Francavilla, M.; Colaianna, M.; Zotti, M.; Morgese, M.; Trotta, P.; Tucci, P.; Schiavone, S.; Cuomo, V.; Trabace, L. Extraction, Characterization and in vivo neuromodulatory activity of phytosterols from microalga Dunaliella tertiolecta. Curr. Med. Chem. 2012, 19, 3058–3067. [Google Scholar] [CrossRef] [PubMed]
- Kazlowska, K.; Lin, H.T.V.; Chang, S.H.; Tasi, G.J. In vitro and in vivo anticancer effects of sterol fraction from red algae Porphyra dentata. Evid. Based Complement. Altern. Med. 2013, 2013, 10. [Google Scholar] [CrossRef] [PubMed]
- Yusnelti, Y.M.; Dharma, A.; Darwis, D.; Munaf, E. Steroids from N-Hexane fraction of the stem bark of shorea singkawang mig and anticancer activity as tested with Murin Leukemia P-388 cells. Res. J. Pharm. Biol. Chem. Sci. 2015, 6, 1315–1320. [Google Scholar]
- Mohammady, N. Different light spectral qualities influence sterol pool in Porphyridium cruentum (Rhodophyta). Am. J. Plant Physiol. 2007, 2, 115–121. [Google Scholar]
- Akihisa, T.; Hori, T.; Suzuki, H.; Sakoh, T.; Yokota, T.; Tamura, T. 24β-methyl-5α-cholest-9(11)-en-3β-ol, two 24β-alkyl-Δ5,7,9(11)-sterols and other 24β-alkylsterol from Chlorella vulgaris. Phytochemistry 1992, 31, 1769–1772. [Google Scholar] [CrossRef]
- Miller, M.B.; Haubrich, B.A.; Wang, Q.; Snell, W.J.; Nes, W.D. Evolutionarily conserved Δ25(27)-olefin ergosterol biosynthesis pathway in the alga Chlamydomonas reinhardtii. J. Lipid Res. 2012, 53, 1636–1645. [Google Scholar] [CrossRef] [PubMed]
- Ciminiello, P.; Fattorusso, E.; Magno, S.; Mangoni, A.; Pansini, M. Incisterols, a new class of highly degraded sterols from the marine sponge Dictyonella incisa. J. Am. Chem. Soc. 1990, 112, 3505–3509. [Google Scholar] [CrossRef]
- Ma, L.; Chen, H.; Dong, P.; Lu, X. Anti-inflammatory and anticancer activities of extracts and compounds from the mushroom Inonotus obliquus. Food Chem. 2013, 139, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.P.; Xie, Y.Z.; Deng, Z.; Li, X.M.; Yang, W.; Jiao, C.W.; Fang, L.; Li, S.Z.; Pan, H.H.; Yee, A.J. Ergosterol peroxide isolated from Ganoderma lucidum abolishes microRNA miR-378-mediated tumor cells on chemoresistance. PLoS ONE 2012, 7, e44579. [Google Scholar] [CrossRef] [PubMed]
- Gil-Ramírez, A.; Caz, V.; Martin-Hernandez, R.; Marín, F.R.; Largo, C.; Rodríguez-Casado, A.; Tabernero, M.; Ruiz-Rodríguez, A.; Reglero, G.; Soler-Rivas, C. Modulation of cholesterol-related gene expression by ergosterol and ergosterol-enriched extracts obtained from Agaricus bisporus. Eur. J. Nutr. 2015, 1–17. [Google Scholar]
- Klosty, M.; Bergmann, W. Sterols of Algae. III. The occurrence of ergosterol in Chlorella pyranoidosa. J. Am. Chem. Soc. 1952, 74, 1601–1601. [Google Scholar] [CrossRef]
- Plaza, M.; Cifuentes, A.; Ibáñez, E. In the search of new functional food ingredients from algae. Trends Food Sci. Technol. 2008, 19, 31–39. [Google Scholar] [CrossRef]
- Kim, K.B.W.R.; Kim, M.J.; Ahn, D.H. Lipase inhibitory activity of chlorophyll a, isofucosterol and saringosterol isolated from chloroform fraction of Sargassum thunbergii. Nat. Prod. Res. 2014, 28, 1310–1312. [Google Scholar] [CrossRef] [PubMed]
- Wächter, G.A.; Franzblau, S.G.; Montenegro, G.; Hoffmann, J.J.; Maiese, W.M.; Timmermann, B.N. Inhibition of Mycobacterium tuberculosis growth by saringosterol from Lessonia nigrescens. J. Nat. Prod. 2001, 64, 1463–1464. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Liu, J.; Fu, Z.; Ye, C.; Zhang, R.; Song, Y.; Zhang, Y.; Li, H.; Ying, H.; Liu, H. 24(S)-Saringosterol from edible marine seaweed Sargassum fusiforme is a novel selective LXRβ agonist. J. Agric. Food Chem. 2014, 62, 6130–6137. [Google Scholar] [CrossRef] [PubMed]
- Awad, A.B.; Chan, K.C.; Downie, A.C.; Fink, C.S. Peanuts as a source of β-sitosterol, a sterol with anticancer properties. Nutr. Cancer 2000, 36, 238–241. [Google Scholar] [CrossRef] [PubMed]
- Byju, K.; Anuradha, V.; Vasundhara, G.; Nair, S.M.; Kumar, N.C. In vitro and in silico studies on the anticancer and apoptosis-inducing activities of the sterols identified from the soft coral, subergorgia reticulata. Pharmacogn. Mag. 2014, 10, S65–S71. [Google Scholar]
- Deepak, M.; Handa, S.S. Antiinflammatory activity and chemical composition of extracts of Verbena officinalis. Phytother. Res. 2000, 14, 463–465. [Google Scholar] [CrossRef]
- Villaseñor, I.M.; Angelada, J.; Canlas, A.P.; Echegoyen, D. Bioactivity studies on β-sitosterol and its glucoside. Phytother. Res. 2002, 16, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Jamaluddin, F.; Mohamed, S.; Lajis, M.N. Hypoglycaemic effect of Parkia speciosa seeds due to the synergistic action of β-sitosterol and stigmasterol. Food Chem. 1994, 49, 339–345. [Google Scholar] [CrossRef]
- Gordon, M.H.; Magos, P. The effect of sterols on the oxidation of edible oils. Food Chem. 1983, 10, 141–147. [Google Scholar] [CrossRef]
- Karabacak, M.; Kanbur, M.; Eraslan, G.; Soyer, S.Z. The antioxidant effect of wheat germ oil on subchronic coumaphos exposure in mice. Ecotoxicol. Environ. Saf. 2011, 74, 2119–2125. [Google Scholar] [CrossRef] [PubMed]
- Małecka, M. Antioxidant properties of the unsaponifiable matter isolated from tomato seeds, oat grains and wheat germ oil. Food Chem. 2002, 79, 327–330. [Google Scholar] [CrossRef]
- Wei, W.; Yang, Y. Preparation of two phytosterols from rape bee pollen and determination by HPLC-ELSD. Chin. J. Pharm. 2011, 6, 008. [Google Scholar]
- Giner, J.L.; Gunasekera, S.P.; Pomponi, S.A. Sterols of the marine sponge Petrosia weinbergi: Implications for the absolute configurations of the antiviral orthoesterols and weinbersterols. Steroids 1999, 64, 820–824. [Google Scholar] [CrossRef]
- Demonty, I.; Haddeman, E.; van der Put, N.; Duchateau, G.; Steenbergen, H.; Diks, R.; Trautwein, E. Spreads fortified with a brassicasterol-rich phytosterol mixture from rapeseed oil lower serum total and LDL-cholesterol concentrations in mildly hypercholesterolemic subjects. J. Fed. Am. Soc. Exp. Biol. 2007, 21, A1089. [Google Scholar]
- Napolitano, G.E.; Ackman, R.G.; Silva-Serra, M.A. Incorporation of dietary sterols by the sea scallop Placopecten magellanicus (Gmelin) fed on microalgae. Mar. Biol. 1993, 117, 647–654. [Google Scholar] [CrossRef]
- Chu, F.L.E.; Lund, E.D.; Littreal, P.R.; Ruck, K.E.; Harvey, E.; Le Coz, J.; Marty, Y.; Moal, J.; Soudant, P. Sterol production and phytosterol bioconversion in two species of heterotrophic protists, Oxyrrhis marina and Gyrodinium dominans. Mar. Biol. 2008, 156, 155–169. [Google Scholar] [CrossRef]
- Ho, D.S.S. Recovery of Phytonutrients from Oils. EP Patent 1689353 B1, 9 January 2005. [Google Scholar]
- Smith, F.E. Separation of Tocopherols and Sterols from Deodorizer Sludge and the Like. U.S. Patent 3335154 A, 8 August 1967. [Google Scholar]
- Fizet, C. Process for Tocopherols and Sterols from Natural Sources. U.S. Patent 5487817 A, 30 January 1996. [Google Scholar]
- Huibers, D.T.A.; Robbins, A.M.; Sullivan, D.H. Method for Separating Sterols from Tall Oil. U.S. Patent 6107456, 22 August 2000. [Google Scholar]
- Mouahid, A.; Crampon, C.; Toudji, S.A.; Badens, E. Supercritical CO2 extraction of neutral lipids from microalgae: Experiments and modelling. J. Supercrit. Fluids 2013, 77, 7–16. [Google Scholar]
- Snyder, J.M.; King, J.W.; Taylor, S.L.; Neese, A.L. Concentration of phytosterols for analysis by supercritical fluid extraction. J. Am. Oil Chem. Soc. 1999, 76, 717–721. [Google Scholar] [CrossRef]
- Dunford, N.T.; King, J.W. Phytosterol Enrichment of Rice Bran Oil by a Supercritical Carbon Dioxide Fractionation Technique. J. Food Sci. 2000, 65, 1395–1399. [Google Scholar] [CrossRef]
- Nyam, K.L.; Tan, C.P.; Lai, O.M.; Long, K.; Che, M.Y.B. Optimization of supercritical fluid extraction of phytosterol from roselle seeds with a central composite design model. Food Bioprod. Process. 2010, 88, 239–246. [Google Scholar] [CrossRef]
- Sajfrtová, M.; Lickova, I.; Wimmerova, M.; Sovova, H.; Wimmer, Z. β-Sitosterol: Supercritical carbon dioxide extraction from sea buckthorn (Hippophae rhamnoides L.) seeds. Int. J. Mol. Sci. 2010, 11, 1842–1850. [Google Scholar] [CrossRef] [PubMed]
- Herrero, M.; Cifuentes, A.; Ibañez, E. Sub- and supercritical fluid extraction of functional ingredients from different natural sources: Plants, food-by-products, algae and microalgae: A review. Food Chem. 2006, 98, 136–148. [Google Scholar] [CrossRef]
- Capuzzo, A.; Maffei, M.; Occhipinti, A. Supercritical fluid extraction of plant flavors and fragrances. Molecules 2013, 18, 7194–7238. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Weller, C.L. Recent advances in extraction of nutraceuticals from plants. Trends Food Sci. Technol. 2006, 17, 300–312. [Google Scholar] [CrossRef]
- Uddin, M.S.; Sarker, M.Z.I.; Ferdosh, S.; Akanda, M.J.H.; Easmin, M.S.; Bt Shamsudin, S.H.; Yunus, K.B. Phytosterols and their extraction from various plant matrices using supercritical carbon dioxide: A review. J. Sci. Food Agric. 2015, 95, 1385–1394. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Liu, T.; Chen, X.; Chen, L.; Zhang, W.; Wang, J.; Gao, L.; Chen, Y.; Peng, X. Subcritical co-solvents extraction of lipid from wet microalgae pastes of Nannochloropsis sp. Eur. J. Lipid Sci. Technol. 2012, 114, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Dejoye, C.; Vian, M.A.; Lumia, G.; Bouscarle, C.; Charton, F.; Chemat, F. Combined extraction processes of lipid from Chlorella vulgaris microalgae: Microwave prior to supercritical carbon dioxide extraction. Int. J. Mol. Sci. 2011, 12, 9332–9341. [Google Scholar] [CrossRef] [PubMed]
- Ktari, L.; Blond, A.; Guyot, M. 16β-Hydroxy-5α-cholestane-3,6-dione, a novel cytotoxic oxysterol from the red alga Jania rubens. Bioorganic Med. Chem. Lett. 2000, 10, 2563–2565. [Google Scholar] [CrossRef]
- Sheu, J.H.; Wang, G.H.; Sung, P.J.; Chiu, Y.H.; Duh, C.Y. Cytotoxic sterols from the formosan brown alga Turbinaria ornata. Planta Medica 1997, 63, 571–572. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.S.; Engel, S.; Smith, B.A.; Fairchild, C.R.; Aalbersberg, W.; Hay, M.E.; Kubanek, J. Structure and biological evaluation of novel cytotoxic sterol glycosides from the marine red alga Peyssonnelia sp. Bioorganic Med. Chem. 2010, 18, 8264–8269. [Google Scholar] [CrossRef] [PubMed]
- Nes, W.D. Biosynthesis of cholesterol and other sterols. Chem. Rev. 2011, 111, 6423–6451. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Jiang, Y.; Liang, Y.; Tian, X.; Peng, C.; Ma, K.Y.; Liu, J.; Huang, Y.; Chen, Z.Y. DPA n-3, DPA n-6 and DHA improve lipoprotein profiles and aortic function in hamsters fed a high cholesterol diet. Atherosclerosis 2012, 221, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Pistocchi, R.; Guerrini, F.; Pezzolesi, L.; Riccardi, M.; Vanucci, S.; Ciminiello, P.; Dell’Aversano, C.; Forino, M.; Fattorusso, E.; Tartaglione, L. Toxin levels and profiles in microalgae from the North-Western Adriatic Sea—15 years of studies on cultured species. Mar. Drugs 2012, 10, 140–162. [Google Scholar] [CrossRef] [PubMed]
- Dorantes-Aranda, J.J.; Waite, T.D.; Godrant, A.; Rose, A.L.; Tovar, C.D.; Woods, G.M.; Hallegraeff, G.M. Novel application of a fish gill cell line assay to assess ichthyotoxicity of harmful marine microalgae. Harmful Algae 2011, 10, 366–373. [Google Scholar] [CrossRef]
- Alemany, L.; Barbera, R.; Alegria, A.; Laparra, J.M. Plant sterols from foods in inflammation and risk of cardiovascular disease: A real threat? Food Chem. Toxicol. 2014, 69, 140–149. [Google Scholar] [CrossRef] [PubMed]
- González-Larena, M.; Garcia-Llatas, G.; Clemente, G.; Barbera, R.; Lagarda, M.J. Plant sterol oxides in functional beverages: Influence of matrix and storage. Food Chem. 2015, 173, 881–889. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, X.; Su, P.; Zhang, W. Advances in Microalgae-Derived Phytosterols for Functional Food and Pharmaceutical Applications. Mar. Drugs 2015, 13, 4231-4254. https://doi.org/10.3390/md13074231
Luo X, Su P, Zhang W. Advances in Microalgae-Derived Phytosterols for Functional Food and Pharmaceutical Applications. Marine Drugs. 2015; 13(7):4231-4254. https://doi.org/10.3390/md13074231
Chicago/Turabian StyleLuo, Xuan, Peng Su, and Wei Zhang. 2015. "Advances in Microalgae-Derived Phytosterols for Functional Food and Pharmaceutical Applications" Marine Drugs 13, no. 7: 4231-4254. https://doi.org/10.3390/md13074231