
Potential Antiviral Agents from Marine Fungi: An Overview

Abstract
:1. Introduction
2. Marine Fungi and Their Antiviral Activity
2.1. Enterovirus-71 (EV-71)




{kind=link}
{kind=link}
{kind=link}
| Virus | Antiviral Agent | Source | Chemical Class | Effect/Mechanism | Reference | |
|---|---|---|---|---|---|---|
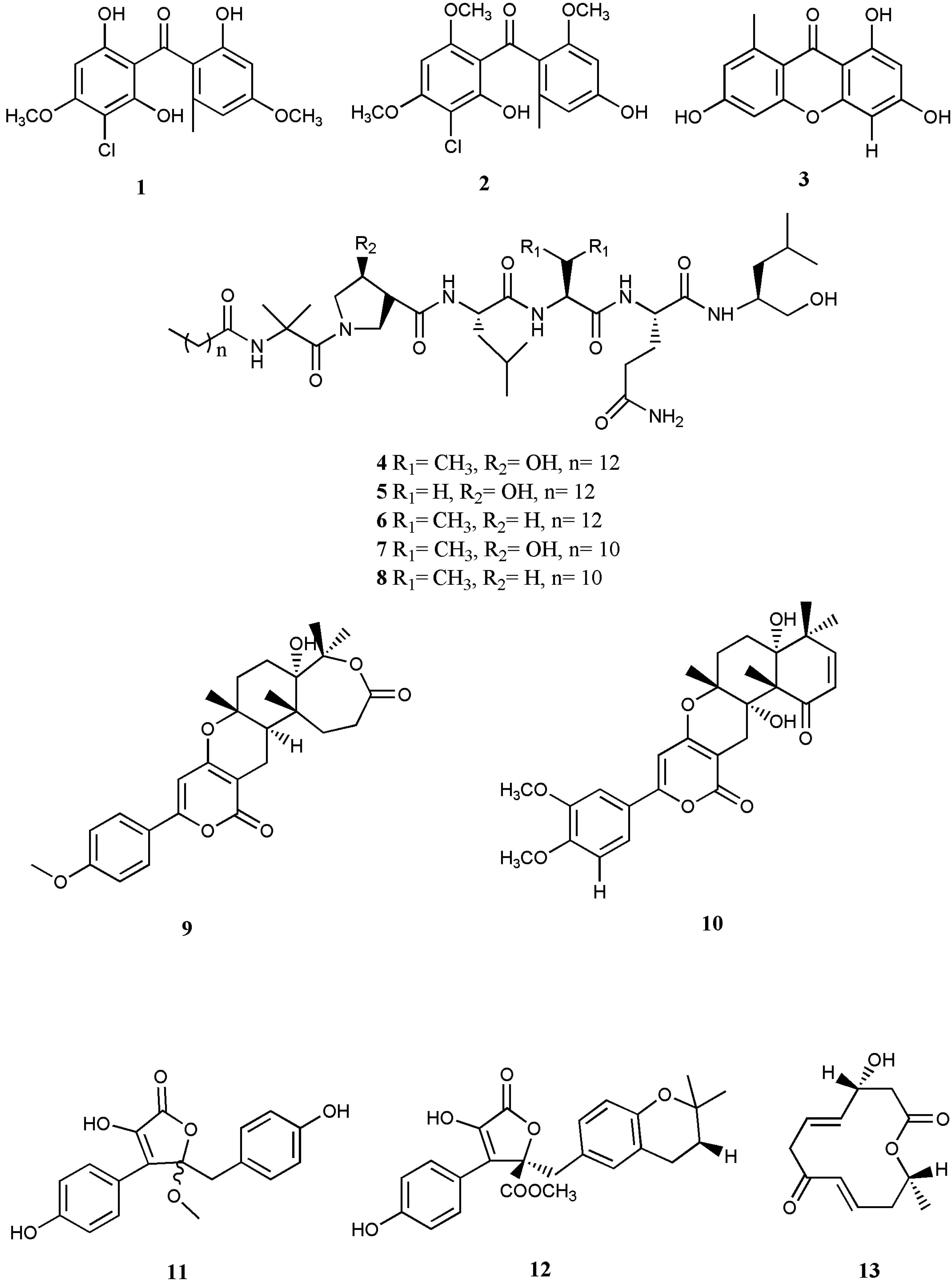
| EV71 | Stachybogrisephenone B (1) | Stachybotrys sp. | Xanthone | IC50: 30.1 µM | [34] | |
| Grisephenone A (2) | Stachybotrys sp | Xanthone | IC50: 50.0 µM | [34] | ||
| 3,6,8-Trihydroxy-1-methylxanthone (3) | Stachybotrys sp | Xanthone | IC50: 40.3 µM | [34] | ||
| HSV | Halovirs A–E (4–8) | Scytalidium sp. | Peptide | direct inactivation | [36] | |
| 11a-dehydroxyisoterreulactone A (9) | Aspergillus terreus SCSGAF0162 | Lactone | IC50: 33.38 μM | [37] | ||
| Arisugacin A (10) | Aspergillus terreus SCSGAF0162 | Lactone | IC50: 12.76 μM | [37] | ||
| Isobutyrolactone II (11) | Aspergillus terreus SCSGAF0162 | Lactone | IC50: 62.08 μM | [37] | ||
| Aspernolide A (12) | Aspergillus terreus SCSGAF0162 | Lactone | IC50: 68.16 μM | [37] | ||
| Balticolid (13) | Ascomycetous strain 222 | Macrolide | IC50: 0.45 μM | [38] | ||
| HIV | Equisetin (14) | Fusarium heterosporum | Tetramic acid | IC50: 15 µM | [39] | |
| Phomasetin (15) | Phoma sp. | Tetramic acid | IC50: 10 µM | [39] | ||
| Integric acid (16) | Xylaria sp. | Acylated eremophilane sesquiterpenoid | IC50: 10 µM | [40] | ||
| IFV | Stachyflin (17) | Stachybotrys sp. RF-7260 | Sesquiterpenoidal alkaloid | inhibition of fusion between the viral envelope and the endosome | [41,42] | |
| Oxoglyantrypine (18) | Cladosporium sp. | Indole alkaloid | IC50: 85 µM | [43] | ||
| Norquinadoline A (19) | Cladosporium sp. | Indole alkaloid | IC50: 82 µM | [43] | ||
| Deoxynortryptoquivaline (20) | Cladosporium sp. | Alkaloid | IC50: 87 µM | [43] | ||
| Deoxytryptoquivaline (21) | Cladosporium sp. | Alkaloid | IC50: 85 µM | [43] | ||
| Tryptoquivaline (22) | Cladosporium sp. | Alkaloid | IC50: 89 µM | [43] | ||
| Quinadoline B (23) | Cladosporium sp. | Alkaloid | IC50: 82 µM | [43] | ||
| Cladosin C (24) | Cladosporium sphaerospermum 2005-01-E3 | Hybrid polyketide | IC50: 276 μM | [44] | ||
| (Z)-5-(Hydroxymenthyl)-2-(6′)-methylhept-2′-en-2′-yl)-phenol (25) | A. sydowii ZSDS1-F6 | Bisabolane-type sesquiterpenoid | IC50: 57.4 µM | [45] | ||
| Diorcinol (26) | A. sydowii ZSDS1-F6 | Prenylated diphenyl ether | IC50: 66.5 µM | [45] | ||
| Cordyol C (27) | A. sydowii ZSDS1-F6 | Diphenyl ether | IC50: 78.5 µM | [45] | ||
| Rubrolide S (28) | A. terreus OUCMDZ-1925 | Rubrolide | IC50: 87.1 µM | [46] | ||
| Asperterrestide A (29) | A.terreus SCSGAF0162 | Cyclic tetrapeptide | H1N1 IC50: 15 µM; H3N2 IC50: 8.1 µM | [47] | ||
| Isoaspulvinone E (30) | A.terreus Gwq-48 | Butenolide | IC50: 101.23 µM | [48] | ||
| Aspulvinone E (31) | A.terreus Gwq-48 | Butenolide | IC50: 192.05 µM | [48] | ||
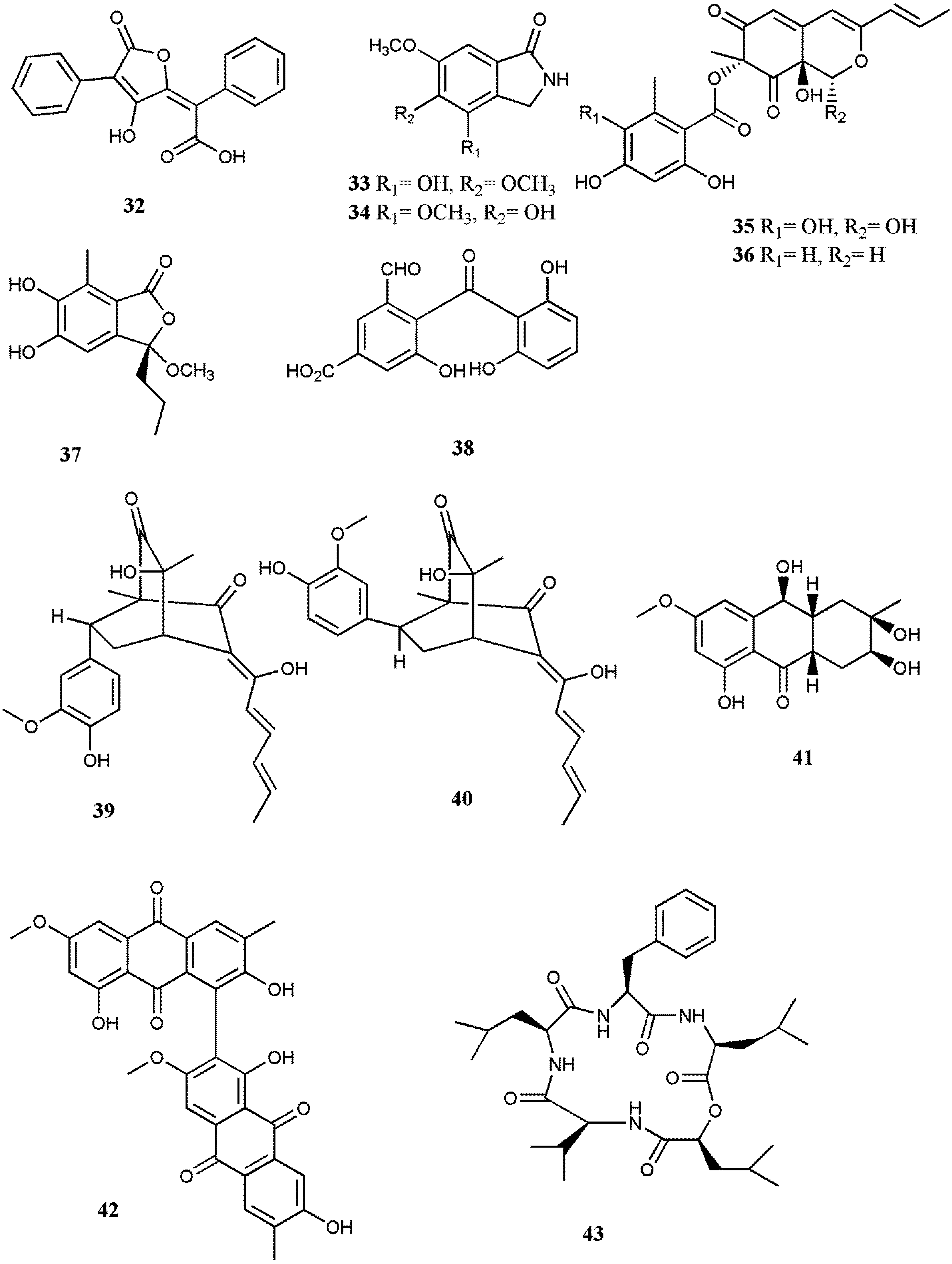
| Pulvic acid (32) | A.terreus Gwq-48 | Butenolide | IC50: 94.42 µM | [48] | ||
| Emerimidine A (33) | Emericella sp. (HK-ZJ) | Isoindolone | IC50: 201.1 µM | [49] | ||
| Emerimidine B (34) | Emericella sp. (HK-ZJ) | Isoindolone | IC50: 296.62 µM | [49] | ||
| Purpurquinone B (35) | P. purpurogenum JS03-21 | Azaphilone | IC50: 61.3 μM | [50] | ||
| Purpurquinone C (36) | P. purpurogenum JS03-21 | Azaphilone | IC50: 64.0 μM | [50] | ||
| Purpuresters A (37) | P. purpurogenum JS03-21 | Benzofuran | IC50: 85.3 μM | [50] | ||
| TAN-931 (38) | P. purpurogenum JS03-21 | Nonsteroidal aromatase | IC50: 58.6 μM | [50] | ||
| Sorbicatechol A (39) | P. chrysogenum PJX-17 | Sorbicillinoids | IC50: 85 μM | [51] | ||
| Sorbicatechol B (40) | P. chrysogenum PJX-17 | Sorbicillinoids | IC50: 113 µM | [51] | ||
| PRRS | Tetrahydroaltersolanol C (41) | Alternaria sp. ZJ-2008003 | Anthraquinone derivatives | IC50: 65 µM | [52] | |
| Alterporriol Q (42) | Alternaria sp. ZJ-2008003 | Isoindolone derivatives | IC50: 39 µM | [52] | ||
| MCV | Sansalvamide A (43) | Fusarium sp. | Pentadepsipeptide | inhibition of topoisomerase | [53] | |
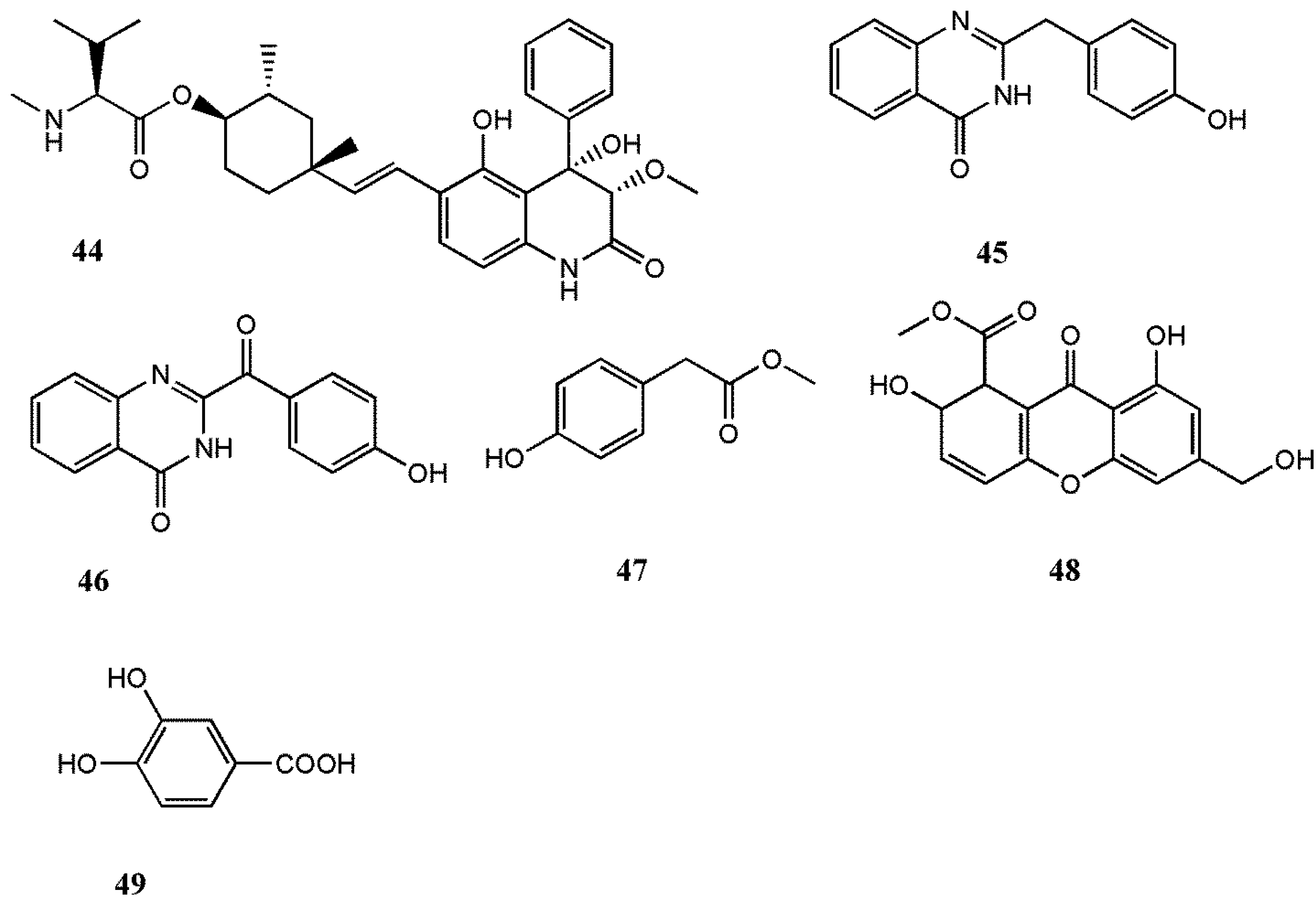
| RSV | 22- O-(N-Me-l-valyl)-21-epi-aflaquinolone B (44) | Aspergillus sp. XS-20090B15 | Prenylated dihydroquinolone derivatives | IC50: 42 nM | [54] | |
| TMV | 2-(4-hydroxybenzyl) quinazolin-4(3H)-one (45) | P. oxalicum 0312F1 | Alkaloid | EC50: 399.57 µM | [55,56] | |
| 2-(4-hydroxybenzoyl) quinazolin-4(3H)-one (46) | P. oxalicum 0312F1 | Alkaloid | EC50 not tested | [55,56] | ||
| Methyl 4-hydroxyphenylacetate (47) | P. oxalicum 0312F1 | Ester | EC50: 829.15 µM | [55,56] | ||
| AGI-B4 (48) | Neosartorya fischeri 1008F1 | Dihydroxanthenone | IC50: 260 µM | [57] | ||
| 3,4-dihydroxybenzoic acid (49) | Neosartorya fischeri 1008F1 | Polyphenol | IC50: 630 µM | [57] | ||
2.2. Herpes Simplex Viruses (HSVs)
2.3. Human Immunodeficiency Virus (HIV)
2.4. Influenza Virus
2.5. Porcine Reproductive and Respiratory Syndrome Virus (PRRSV)
2.6. Molluscum Contagiosum Virus (MCV)
2.7. Respiratory Syncytial Virus (RSV)
2.8. Tobacco Mosaic Virus (TMV)
3. Conclusions
Acknowledgments
Conflicts of Interest
References
- Vo, T.-S.; Kim, S.-K. Potential anti-HIV agents from marine resources: An overview. Mar. Drugs 2010, 8, 2871–2892. [Google Scholar] [CrossRef] [PubMed]
- Aneiros, A.; Garateix, A. Bioactive peptides from marine sources: Pharmacological properties and isolation procedures. J. Chromatogr. B 2004, 803, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Bhadury, P.; Mohammad, B.T.; Wright, P.C. The current status of natural products from marine fungi and their potential as anti-infective agents. J. Ind. Microbiol. Biotechnol. 2006, 33, 325–337. [Google Scholar] [CrossRef] [PubMed]
- Simon, C.; Daniel, R. Metagenomic analyses: Past and future trends. Appl. Environ. Microbiol. 2011, 77, 1153–1161. [Google Scholar] [CrossRef] [PubMed]
- Faulkner, D.J. Marine pharmacology. Antonie Leeuwenhoek 2000, 77, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Zorofchian Moghadamtousi, S.; Karimian, H.; Khanabdali, R.; Razavi, M.; Firoozinia, M.; Zandi, K.; Abdul Kadir, H. Anticancer and antitumor potential of fucoidan and fucoxanthin, two main metabolites isolated from brown algae. Sci. World J. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Faulkner, D.J. Marine natural products. Nat. Prod. Rep. 2001, 18, 1R–49R. [Google Scholar] [CrossRef]
- Fenical, W. New pharmaceuticals from marine organisms. Trends Biotechnol. 1997, 15, 339–341. [Google Scholar] [CrossRef]
- König, G.M.; Wright, A.D.; Sticher, O.; Angerhofer, C.K.; Pezzuto, J.M. Biological activities of selected marine natural products. Planta Med. 1994, 60, 532–537. [Google Scholar] [CrossRef] [PubMed]
- Bugni, T.S.; Ireland, C.M. Marine-derived fungi: A chemically and biologically diverse group of microorganisms. Nat. Prod. Rep. 2004, 21, 143–163. [Google Scholar] [CrossRef] [PubMed]
- Kornprobst, J.-M.; Ha, T.B.T. Encyclopedia of Marine Natural Products; Wiley-Blackwell: Weinheim, Germany, 2010; Volume 1. [Google Scholar]
- Hart, C. Forged in St. Anthony’s fire: Drugs for migraine. Mod. Drug Disc. 1999, 2, 20–21. [Google Scholar]
- Tresner, H.; Hayes, J.A. Sodium chloride tolerance of terrestrial fungi. Appl. Microbiol. 1971, 22, 210–213. [Google Scholar] [PubMed]
- Smith, G.W.; Ives, L.D.; Nagelkerken, I.A.; Ritchie, K.B. Caribbean sea-fan mortalities. Nature 1996, 383, 487. [Google Scholar] [CrossRef]
- Alderman, D.; Polglase, J. Are fungal diseases significant in the marine environment? In The Biology of Marine Fungi; Cambridge University Press: Cambridge, UK, 1986; p. 189. [Google Scholar]
- Porter, D. Mycoses of marine organisms: An overview of pathogenic fungi. In The Biology of Marine Fungi; Cambridge University Press: Cambridge, UK, 1986; p. 141. [Google Scholar]
- Duarte, K.; Rocha-Santos, T.A.; Freitas, A.C.; Duarte, A.C. Analytical techniques for discovery of bioactive compounds from marine fungi. Trends Analyt. Chem. 2012, 34, 97–110. [Google Scholar] [CrossRef]
- Kohlmeyer, J.; Kohlmeyer, E. Marine Mycology: The Higher Fungi; Academic Press: New York, NY, USA, 1979. [Google Scholar]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2014, 31, 160–258. [Google Scholar] [CrossRef] [PubMed]
- Gallo, M.L.; Seldes, A.M.; Cabrera, G.M. Antibiotic long-chain and α, β-unsaturated aldehydes from the culture of the marine fungus Cladosporium sp. Biochem. Syst. Ecol. 2004, 32, 545–551. [Google Scholar] [CrossRef]
- Abdel-Lateff, A.; Klemke, C.; König, G.M.; Wright, A.D. Two new xanthone derivatives from the algicolous marine fungus wardomyces anomalus. J. Nat. Prod. 2003, 66, 706–708. [Google Scholar] [CrossRef] [PubMed]
- Daferner, M.; Anke, T.; Sterner, O. Zopfiellamides A and B, antimicrobial pyrrolidinone derivatives from the marine fungus Zopfiella latipes. Tetrahedron 2002, 58, 7781–7784. [Google Scholar] [CrossRef]
- Gautschi, J.T.; Amagata, T.; Amagata, A.; Valeriote, F.A.; Mooberry, S.L.; Crews, P. Expanding the strategies in natural product studies of marine-derived fungi: A chemical investigation of Penicillium obtained from deep water sediment. J. Nat. Prod. 2004, 67, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Tziveleka, L.-A.; Vagias, C.; Roussis, V. Natural products with anti-HIV activity from marine organisms. Curr. Top. Med. Chem. 2003, 3, 1512–1535. [Google Scholar] [CrossRef] [PubMed]
- Lou, Z.; Sun, Y.; Rao, Z. Current progress in antiviral strategies. Trends Pharmacol. Sci. 2014, 35, 86–102. [Google Scholar] [CrossRef] [PubMed]
- Tantillo, C.; Ding, J.; Jacobo-Molina, A.; Nanni, R.G.; Boyer, P.L.; Hughes, S.H.; Pauwels, R.; Andries, K.; Janssen, P.A.; Arnold, E. Locations of anti-aids drug binding sites and resistance mutations in the three-dimensional structure of HIV-1 reverse transcriptase: Implications for mechanisms of drug inhibition and resistance. J. Mol. Biol. 1994, 243, 369–387. [Google Scholar] [CrossRef] [PubMed]
- Morfin, F.; Thouvenot, D. Herpes simplex virus resistance to antiviral drugs. J. Clin. Virol. 2003, 26, 29–37. [Google Scholar] [CrossRef]
- Gilbert, C.; Boivin, G. Human cytomegalovirus resistance to antiviral drugs. Antimicrob. Agents Chemother. 2005, 49, 873–883. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the 30 years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef] [PubMed]
- Taishi, T.; Takechi, S.; Mori, S. First total synthesis of (±)-stachyflin. Tetrahedron Lett. 1998, 39, 4347–4350. [Google Scholar] [CrossRef]
- McMinn, P.; Stratov, I.; Nagarajan, L.; Davis, S. Neurological manifestations of Enterovirus 71 infection in children during an outbreak of hand, foot, and mouth disease in Western Australia. Clin. Infect. Dis. 2001, 32, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.-Y.; Chang, L.-Y.; Hsia, S.-H.; Huang, Y.-C.; Chiu, C.-H.; Hsueh, C.; Shih, S.-R.; Liu, C.-C.; Wu, M.-H. The 1998 Enterovirus 71 outbreak in Taiwan: Pathogenesis and management. Clin. Infect. Dis. 2002, 34, S52–S57. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.X.; Ng, M.M.-L.; Chu, J.J. Developments towards antiviral therapies against Enterovirus 71. Drug. Discov. Today 2010, 15, 1041–1051. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Lin, X.; Lu, X.; Wan, J.; Zhou, X.; Liao, S.; Tu, Z.; Xu, S.; Liu, Y. Sesquiterpenoids and xanthones derivatives produced by sponge-derived fungus Stachybotry sp. HH1 ZSDS1F1-2. J. Antibiot. 2014, 68, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Rabenau, H.F.; Richter, M.; Doerr, H.W. Hand, foot and mouth disease: Seroprevalence of Coxsackie A16 and Enterovirus 71 in Germany. Med. Microbiol. Immun. 2010, 199, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Rowley, D.C.; Kelly, S.; Kauffman, C.A.; Jensen, P.R.; Fenical, W. Halovirs A–E, new antiviral agents from a marine-derived fungus of the genus Scytalidium. Bioorgan. Med. Chem. 2003, 11, 4263–4274. [Google Scholar] [CrossRef]
- Nong, X.-H.; Wang, Y.-F.; Zhang, X.-Y.; Zhou, M.-P.; Xu, X.-Y.; Qi, S.-H. Territrem and butyrolactone derivatives from a marine-derived fungus Aspergillus terreus. Mar. Drugs 2014, 12, 6113–6124. [Google Scholar] [CrossRef] [PubMed]
- Shushni, M.A.; Singh, R.; Mentel, R.; Lindequist, U. Balticolid: A new 12-membered macrolide with antiviral activity from an Ascomycetous fungus of marine origin. Mar. Drugs 2011, 9, 844–851. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.B.; Zink, D.; Polishook, J.; Valentino, D.; Shafiee, A.; Silverman, K.; Felock, P.; Teran, A.; Vilella, D.; Hazuda, D.J. Structure and absolute stereochemistry of HIV-1 integrase inhibitor integric acid. A novel eremophilane sesquiterpenoid produced by a Xylaria sp. Tetrahedron Lett. 1999, 40, 8775–8779. [Google Scholar] [CrossRef]
- Rowley, D.C.; Kelly, S.; Jensen, P.; Fenical, W. Synthesis and structure–activity relationships of the halovirs, antiviral natural products from a marine-derived fungus. Bioorgan. Med. Chem. 2004, 12, 4929–4936. [Google Scholar] [CrossRef] [PubMed]
- Minagawa, K.; Kouzuki, S.; Yoshimoto, J.; Kawamura, Y.; Tani, H.; Iwata, T.; Terui, Y.; Nakai, H.; Yagi, S.; Hattori, N. Stachyflin and acetylstachyflin, novel anti-influenza a virus substances, produced by Stachybotrys sp. RF-7260. I. Isolation, structure elucidation and biological activities. J. Antibiot. 2002, 55, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Yagi, S.; Ono, J.; Yoshimoto, J.; Sugita, K.-I.; Hattori, N.; Fujioka, T.; Fujiwara, T.; Sugimoto, H.; Hirano, K.; Hashimoto, N. Development of anti-influenza virus drugs I: Improvement of oral absorption and in vivo anti-influenza activity of stachyflin and its derivatives. Pharmaceut. Res. 1999, 16, 1041–1046. [Google Scholar] [CrossRef]
- Peng, J.; Lin, T.; Wang, W.; Xin, Z.; Zhu, T.; Gu, Q.; Li, D. Antiviral alkaloids produced by the mangrove-derived fungus Cladosporium sp. PJX-41. J. Nat. Prod. 2013, 76, 1133–1140. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Sun, X.; Yu, G.; Wang, W.; Zhu, T.; Gu, Q.; Li, D. Cladosins A–E, hybrid polyketides from a deep-sea-derived fungus, Cladosporium sphaerospermum. J. Nat. Prod. 2014, 77, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-F.; Lin, X.-P.; Qin, C.; Liao, S.-R.; Wan, J.-T.; Zhang, T.-Y.; Liu, J.; Fredimoses, M.; Chen, H.; Yang, B. Antimicrobial and antiviral sesquiterpenoids from sponge-associated fungus, Aspergillus sydowii ZSDS1-F6. J. Antibiot. 2014, 67, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Chen, Z.; Liu, P.; Wang, Y.; Xin, Z.; Zhu, W. New rubrolides from the marine-derived fungus Aspergillus terreus OUCMDZ-1925. J. Antibiot. 2014, 67, 315–318. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Bao, J.; Zhang, X.-Y.; Tu, Z.-C.; Shi, Y.-M.; Qi, S.-H. Asperterrestide A, a cytotoxic cyclic tetrapeptide from the marine-derived fungus Aspergillus terreus SCSGAF0162. J. Nat. Prod. 2013, 76, 1182–1186. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Guo, W.; Wang, Q.; Zhang, L.; Zhu, M.; Zhu, T.; Gu, Q.; Wang, W.; Li, D. Aspulvinones from a mangrove rhizosphere soil-derived fungus Aspergillus terreus GWQ-48 with anti-influenza a viral (H1N1) activity. Bioorg. Med. Chem. Lett. 2013, 23, 1776–1778. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Sun, S.; Zhu, T.; Lin, Z.; Gu, J.; Li, D.; Gu, Q. Antiviral isoindolone derivatives from an endophytic fungus Emericella sp. Associated with Aegiceras corniculatum. Phytochemistry 2011, 72, 1436–1442. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, Y.; Wang, W.; Fu, P.; Liu, P.; Zhu, W. Anti-influenza virus polyketides from the acid-tolerant fungus Penicillium purpurogenum JS03-21. J. Nat. Prod. 2011, 74, 2014–2018. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Zhang, X.; Du, L.; Wang, W.; Zhu, T.; Gu, Q.; Li, D. Sorbicatechols A and B, antiviral sorbicillinoids from the marine-derived fungus Penicillium chrysogenum PJX-17. J. Nat. Prod. 2014, 77, 424–428. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.-J.; Shao, C.-L.; Guo, Z.-Y.; Chen, J.-F.; Deng, D.-S.; Yang, K.-L.; Chen, Y.-Y.; Fu, X.-M.; She, Z.-G.; Lin, Y.-C. Bioactive hydroanthraquinones and anthraquinone dimers from a soft coral-derived Alternaria sp. Fungus. J. Nat. Prod. 2012, 75, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.; Rowley, D.; Rhodes, D.; Gertsch, J.; Fenical, W.; Bushman, F. Mechanism of inhibition of a poxvirus topoisomerase by the marine natural product sansalvamide A. Mol. Pharmacol. 1999, 55, 1049–1053. [Google Scholar] [PubMed]
- Prieto, C.; Castro, J.M. Porcine reproductive and respiratory syndrome virus infection in the boar: A review. Theriogenology 2005, 63, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Li, W.; Wang, J. A novel and other bioactive secondary metabolites from a marine fungus Penicillium oxalicum 0312f1. Nat. Prod. Res. 2013, 27, 2286–2291. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Li, W.; Ouyang, M.A.; Wu, Z.; Lin, Q.; Xie, L. Identification of two marine fungi and evaluation of their antivirus and antitumor activities. Acta Microbiol. Sinic. 2009, 49, 1240–1246. [Google Scholar]
- Tan, Q.-W.; Ouyang, M.-A.; Shen, S.; Li, W. Bioactive metabolites from a marine-derived strain of the fungus Neosartorya fischeri. Nat. Prod. Res. 2012, 26, 1402–1407. [Google Scholar] [CrossRef] [PubMed]
- Whitley, R.J.; Roizman, B. Herpes simplex virus infections. Lancet 2001, 357, 1513–1518. [Google Scholar] [CrossRef]
- Taylor, R.; Manandhar, N.; Hudson, J.; Towers, G. Antiviral activities of nepalese medicinal plants. J. Ethnopharmacol. 1996, 52, 157–163. [Google Scholar] [CrossRef]
- Armstrong, G.L.; Conn, L.A.; Pinner, R.W. Trends in infectious disease mortality in the United States during the 20th century. J. Am. Med. Assoc. 1999, 281, 61–66. [Google Scholar] [CrossRef]
- Kanters, S.; Mills, E.; Thorlund, K.; Bucher, H.; Ioannidis, J. Antiretroviral therapy for initial human immunodeficiency virus/AIDS treatment: Critical appraisal of the evidence from over 100 randomized trials and 400 systematic reviews and meta-analyses. Clin. Microbiol. Infect. 2014, 20, 114–122. [Google Scholar] [CrossRef] [PubMed]
- El-Sadr, W.M.; Holmes, C.B.; Mugyenyi, P.; Thirumurthy, H.; Ellerbrock, T.; Ferris, R.; Sanne, I.; Asiimwe, A.; Hirnschall, G.; Nkambule, R.N. Scale-up of HIV treatment through pepfar: A historic public health achievement. J. Acquir. Immune Defic. Syndr. 2012, 60, S96–S104. [Google Scholar] [CrossRef] [PubMed]
- Holmes, C.B.; Blandford, J.M.; Sangrujee, N.; Stewart, S.R.; DuBois, A.; Smith, T.R.; Martin, J.C.; Gavaghan, A.; Ryan, C.A.; Goosby, E.P. Pepfar’s past and future efforts to cut costs, improve efficiency, and increase the impact of global HIV programs. Health Affairs 2012, 31, 1553–1560. [Google Scholar] [CrossRef] [PubMed]
- Hazuda, D.; Blau, C.U.; Felock, P.; Hastings, J.; Pramanik, B.; Wolfe, A.; Bushman, F.; Farnet, C.; Goetz, M.; Williams, M. Isolation and characterization of novel human immunodeficiency virus integrase inhibitors from fungal metabolites. Antivir. Chem. Chemother. 1999, 10, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.-N.; Lu, H.-Z.; Cao, B.; Du, B.; Shang, H.; Gan, J.-H.; Lu, S.-H.; Yang, Y.-D.; Fang, Q.; Shen, Y.-Z. Clinical findings in 111 cases of influenza a (H7N9) virus infection. N. Engl. J. Med. 2013, 368, 2277–2285. [Google Scholar] [CrossRef] [PubMed]
- Pica, N.; Palese, P. Toward a universal influenza virus vaccine: Prospects and challenges. Annu. Rev. Med. 2013, 64, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Scalera, N.M.; Mossad, S.B. The first pandemic of the 21st century: A review of the 2009 pandemic variant influenza a (H1N1) virus. Postgrad. Med. 2009, 121, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Tellier, R. Review of aerosol transmission of influenza a virus. Emerg. Infect. Dis. 2006, 12, 1657–1662. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Yoo, D. Cysteine residues of the porcine reproductive and respiratory syndrome virus small envelope protein are non-essential for virus infectivity. J. Gen. Virol. 2005, 86, 3091–3096. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, C.; Bøtner, A.; Takai, H.; Nielsen, J.P.; Jorsal, S. Experimental airborne transmission of PRRS virus. Vet. Microbiol. 2004, 99, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Anstey, A.V.; Bugert, J.J. Molluscum contagiosum virus infection. Lancet Infect. Dis. 2013, 13, 877–888. [Google Scholar] [CrossRef]
- Heiferman, M.J.; Salabat, M.R.; Ujiki, M.B.; Strouch, M.J.; Cheon, E.C.; Silverman, R.B.; Bentrem, D.J. Sansalvamide induces pancreatic cancer growth arrest through changes in the cell cycle. Anticancer Res. 2010, 30, 73–78. [Google Scholar] [PubMed]
- Thompson, C.H.; de Zwart-Steffe, R.T.; Donovan, B. Clinical and molecular aspects of Molluscum contagiosum infection in HIV-1 positive patients. Int. J. STD AIDS 1992, 3, 101–106. [Google Scholar] [PubMed]
- Falsey, A.R.; Walsh, E.E. Respiratory syncytial virus infection in adults. Clin. Microbiol. Rev. 2000, 13, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Feldman, S.A.; Hendry, R.M.; Beeler, J.A. Identification of a linear heparin binding domain for human respiratory syncytial virus attachment glycoprotein G. J. Virol. 1999, 73, 6610–6617. [Google Scholar] [PubMed]
- Bos, L. Crop losses caused by viruses. Crop Prot. 1982, 1, 263–282. [Google Scholar] [CrossRef]
- Klug, A. The tobacco mosaic virus particle: Structure and assembly. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 1999, 354, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Creager, A.N.; Scholthof, K.-B.G.; Citovsky, V.; Scholthof, H.B. Tobacco mosaic virus: Pioneering research for a century. Plant Cell Online 1999, 11, 301–308. [Google Scholar] [CrossRef]
- Ritzenthaler, C. Resistance to plant viruses: Old issue, news answers? Curr. Opin. Biotechnol. 2005, 16, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.-H.; Chen, J.; Di, Y.-T.; Fang, X.; Dong, J.-H.; Sang, P.; Wang, Y.-H.; He, H.-P.; Zhang, Z.-K.; Hao, X.-J. Anti-tobacco mosaic virus (TMV) quassinoids from Brucea javanica (l.) Merr. J. Agric. Food. Chem. 2010, 58, 1572–1577. [Google Scholar] [CrossRef] [PubMed]
- Megens, S.; Laethem, K.V. Antiretroviral therapy and drug resistance in human immunodeficiency virus type 2 infection. Expert. Rev. Anti. Infect. Ther. 2013, 11, 1159–1178. [Google Scholar] [CrossRef] [PubMed]
- Menéndez-Arias, L.; Álvarez, M. Antiretroviral therapy and drug resistance in human immunodeficiency virus type 2 infection. Antiviral Res. 2014, 102, 70–86. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, T.E.; Pikis, A.; Naeger, L.K.; Harrington, P.R. Resistance of human cytomegalovirus to ganciclovir/valganciclovir: A comprehensive review of putative resistance pathways. Antiviral Res. 2014, 101, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Wang, Y.; Liu, P.; Fu, P.; Zhu, T.; Wang, W.; Zhu, W. Indole-diterpenoids with anti-H1N1 activity from the aciduric fungus Penicillium camemberti OUCMDZ-1492. J. Nat. Prod. 2013, 76, 1328–1336. [Google Scholar] [CrossRef] [PubMed]
- Gong, K.-K.; Tang, X.-L.; Zhang, G.; Cheng, C.-L.; Zhang, X.-W.; Li, P.-L.; Li, G.-Q. Polyhydroxylated steroids from the south China sea soft coral Sarcophyton sp. And their cytotoxic and antiviral activities. Mar. Drugs 2013, 11, 4788–4798. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moghadamtousi, S.Z.; Nikzad, S.; Kadir, H.A.; Abubakar, S.; Zandi, K. Potential Antiviral Agents from Marine Fungi: An Overview. Mar. Drugs 2015, 13, 4520-4538. https://doi.org/10.3390/md13074520
Moghadamtousi SZ, Nikzad S, Kadir HA, Abubakar S, Zandi K. Potential Antiviral Agents from Marine Fungi: An Overview. Marine Drugs. 2015; 13(7):4520-4538. https://doi.org/10.3390/md13074520
Chicago/Turabian StyleMoghadamtousi, Soheil Zorofchian, Sonia Nikzad, Habsah Abdul Kadir, Sazaly Abubakar, and Keivan Zandi. 2015. "Potential Antiviral Agents from Marine Fungi: An Overview" Marine Drugs 13, no. 7: 4520-4538. https://doi.org/10.3390/md13074520