
The Potential Exploitation of the Mediterranean Invasive Alga Caulerpa cylindracea: Can the Invasion Be Transformed into a Gain?

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. NMR Spectoscopy
2.2. Antioxidant Activity
2.3. Antimicrobial Activity
3. Discussion
4. Materials and Methods
4.1. Study Site and Species Collection
4.2. Preparation of Lipidic Extracts from the Macroalga
4.3. NMR Spectroscopy
4.4. Antioxidant Activity
4.4.1. Oxygen Radical Absorbance Capacity Assay (ORAC)
4.4.2. Trolox Equivalent Antioxidant Capacity Assay (TEAC)
4.5. Test Microorganisms
4.6. Antimicrobial Activity
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Raniello, R.; Mollo, E.; Lorenti, M.; Gavagnin, M.; Buia, M.C. Phytotoxic activity of caulerpenyne from the Mediterranean invasive variety of Caulerpa racemosa: A potential allelochemical. Biol. Invasions 2007, 9, 361–368. [Google Scholar] [CrossRef]
- Mollo, E.; Gavagnin, M.; Carbone, M.; Castelluccio, F.; Pozone, F.; Roussis, V.; Templado, J.; Ghiselin, M.T.; Cimino, G. Factors promoting marine invasions: A chemoecological approach. Proc. Natl. Acad. Sci. USA 2008, 105, 4582–4586. [Google Scholar] [CrossRef] [PubMed]
- Terlizzi, A.; Felline, S.; Lionetto, M.G.; Caricato, R.; Perfetti, V.; Cutignano, A.; Mollo, E. Detrimental physiological effects of the invasive alga Caulerpa racemosa on the Mediterranean white seabream Diplodus sargus. Aquat. Biol. 2011, 12, 109–117. [Google Scholar] [CrossRef]
- Pusceddu, A.; Fraschetti, S.; Scopa, M.; Rizzo, L.; Danovaro, R. Meiofauna communities, nematode diversity and C degradation rates in seagrass (Posidonia oceanica L.) and unvegetated sediments invaded by the algae Caulerpa cylindracea (Sonder). Mar. Environ. Res. 2016, 119, 88–99. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, L.; Fraschetti, S.; Alifano, P.; Pizzolante, G.; Stabili, L. The alien species Caulerpa cylindracea and its associated bacteria in the Mediterranean Sea. Mar. Biol. 2016, 163, 1–12. [Google Scholar] [CrossRef]
- Rizzo, L.; Fraschetti, S.; Alifano, P.; Tredici, M.S.; Stabili, L. Association of Vibrio community with the Atlantic Mediterranean invasive alga Caulerpa cylindracea. J. Exp. Mar. Biol. Ecol. 2016, 475, 129–136. [Google Scholar] [CrossRef]
- Ciminiello, P.; Dell’Aversano, C.; Forino, M.; Tartaglione, L. Marine toxins in Italy: The more you look, the more you find. Eur. J. Org. Chem. 2014, 2014, 1357–1369. [Google Scholar] [CrossRef]
- Gorbi, S.; Giuliani, M.E.; Pittura, L.; d’Errico, G.; Terlizzi, A.; Felline, S.; Grauso, L.; Mollo, E.; Cutignano, A.; Regoli, F. Could molecular effects of Caulerpa racemosa metabolites modulate the impact on fish populations of Diplodus sargus? Mar. Environ. Res. 2014, 96, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Felline, S.; Mollo, E.; Ferramosca, A.; Zara, V.; Regoli, F.; Gorbi, S.; Terlizzi, A. Can a marine pest reduce the nutritional value of Mediterranean fish flesh? Mar. Biol. 2014, 161, 1275–1283. [Google Scholar] [CrossRef]
- Mollo, E.; Cimino, G.; Ghiselin, M.T. Alien biomolecules: A new challenge for natural product chemists. Biol. Invasions 2015, 17, 941–950. [Google Scholar] [CrossRef]
- Villarreal-Gómez, L.J.; Soria-Mercado, I.E.; Guerra-Rivas, G.; Ayala-Sánchez, N.E. Antibacterial and anticancer activity of seaweeds and bacteria associated with their surface. Rev. Biol. Mar. Oceanogr. 2010, 45, 267–275. [Google Scholar] [CrossRef]
- Pandian, P.; Selvamuthukumar, S.; Manavalan, R.; Parthasarthy, V. Screening of antibacterial and antifungal activities of red marine algae Acanthaphora spicifera (Rhodophyceae). J. Biomed. Sci. Res. 2011, 3, 444–448. [Google Scholar]
- Senthilkumar, K.; Kim, S.K. Anticancer effects of fucoidan. Adv. Food Nutr. Res. 2014, 72, 195–213. [Google Scholar] [PubMed]
- Chakraborty, K.; Joseph, D.; Joy, M.; Raola, V.K. Characterization of substituted aryl meroterpenoids from red seaweed Hypnea musciformis as potential antioxidants. Food Chem. 2016, 212, 778–788. [Google Scholar] [CrossRef] [PubMed]
- Hardouin, K.; Bedoux, G.; Burlot, A.S.; Donnay-Moreno, C.; Bergé, J.P.; Nyvall-Collén, P.; Bourgougnon, N. Enzyme-assisted extraction (EAE) for the production of antiviral and antioxidant extracts from the green seaweed Ulva armoricana (Ulvales, Ulvophyceae). Algal Res. 2016, 16, 233–239. [Google Scholar] [CrossRef]
- Jiménez-Escrig, A.; Sáchez-Muniz, F.J. Dietary fibre from edible seaweeds: Chemical structure, physicochemical properties and effects on cholesterol metabolism. Nutr. Res. 2000, 20, 585–598. [Google Scholar] [CrossRef]
- Yang, Y.J.; Nam, S.J.; Kong, G.; Kim, M.K. A case-control study on seaweed consumption and the risk of breast cancer. Br. J. Nutr. 2010, 103, 1345–1353. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, Y. Nutrition trends in Japan. Asia Pac. J. Clin. Nutr. 2001, 10, S40–S47. [Google Scholar] [CrossRef] [PubMed]
- Terrados, J.; Lopez-Jimenez, J.A. Fatty acid composition and chilling resistance in the green alga Caulerpa prolifera (Forrskal) Lamouroux (Chlorophyta, Caulerpales). Biochem. Mol. Biol. Int. 1996, 39, 863–869. [Google Scholar] [CrossRef] [PubMed]
- Blažina, M.; Iveša, L.; Najdek, M. Caulerpa racemosa: Adaptive varieties studied by fatty acid composition (Northern Adriatic Sea, Vrsar, Croatia). Eur. J. Phycol. 2009, 44, 183–189. [Google Scholar] [CrossRef]
- Kumar, M.; Gupta, V.; Kumari, P.; Reddy, C.R.K.; Jha, B. Assessment of nutrient composition and antioxidant potential of Caulerpaceae Seaweeds. J. Food Compos. Anal. 2010, 24, 270–278. [Google Scholar] [CrossRef]
- Paul, N.A.; Neveux, N.; Magnusson, M.; de Nys, R. Comparative production and nutritional value of “sea grapes”—The tropical green seaweeds Caulerpa lentillifera and C. racemosa. J. Appl. Phycol. 2014, 26, 1833. [Google Scholar] [CrossRef]
- Kumar, C.S.; Ganesan, P.; Suresh, P.V.; Bhaskar, N. Seaweeds as a Source of Nutritionally Beneficial Compounds—A Review. J. Food Sci. Technol. 2008, 45, 1–13. [Google Scholar]
- Gupta, S.; Rajauria, G.; Abu-Ghannam, N. Study of the microbial diversity and antimicrobial properties of Irish edible brown seaweeds. Int. J. Food Sci. Technol. 2010, 45, 482–489. [Google Scholar] [CrossRef]
- Ballesteros, E.; Martin, D.; Uriz, M.J. Biological activity of extracts from some Mediterranean macrophytes. Bot. Mar. 1992, 35, 481–486. [Google Scholar] [CrossRef] [Green Version]
- Kandhasamy, M.; Arunachalam, K.D. Evaluation of in vitro antibacterial property of seaweeds of southeast coast of India. Afr. J. Biotechnol. 2008, 7, 1958–1961. [Google Scholar]
- Jebasingh, S.E.J.; Rosemary, S.; Elaiyaraja, S.; Sivaraman, K.; Lakshmikandan, M.; Murugan, A.; Raja, P. Potential antibacterial activity of selected green and red seaweeds. J. Pharmac. Biomed. Sci. 2011, 5, 1–7. [Google Scholar]
- Etcherla, M.; Narasimha Rao, G.M. In vitro study of antimicrobial activity in marine algae Caulerpa taxifolia and Caulerpa racemosa (C. Agardh). Int. J. Appl. Biol. Pharm. Technol. 2014, 5, 57–72. [Google Scholar]
- Belton, G.S.; Prud’homme van Reine, W.F.; Huisman, J.M.; Draisma, S.G.A.; Gurgel, C.F.D. Resolving phenotypic plasticity and species designation in the morphology challenging Caulerpa racemosa-peltata complex (Caulerpaceae, Chlorophyta). J. Phycol. 2014, 50, 32–54. [Google Scholar] [CrossRef] [PubMed]
- Verlaque, M.; Durand, C.; Huisman, J.M.; Boudouresque, C.F.; Le Parco, Y. On the identity and origin of the Mediterranean invasive Caulerpa racemosa (Caulerpales, Chlorophyta). Eur. J. Phycol. 2003, 38, 325–339. [Google Scholar] [CrossRef]
- Durand, C.; Manuel, M.; Boudouresque, C.F.; Meinesz, A.; Verlaque, M.; Le Parco, Y. Molecular data suggest a hybrid origin for the invasive Caulerpa racemosa (Caulerpales, Chlorophyta) in the Mediterranean Sea. J. Evol. Biol. 2002, 15, 122–133. [Google Scholar] [CrossRef]
- Sauvage, T.; Payri, C.; Draisma, S.G.A.; Prud’homme van Reine, W.F.; Verbruggen, H.; Belton, G.S.; Gabriel, D.; Sherwood, A.R.; Fredericq, S.; Gurgel, C.F.D. Molecular diversity of the Caulerpa racemosa–Caulerpa peltata complex (Caulerpaceae, Bryopsidales) in New Caledonia, with new Australasian records for C. racemosa var. cylindracea. Phycologia 2013, 52, 6–13. [Google Scholar] [CrossRef]
- Wright, J.T.; Davis, A.R. Demographic feedback between clonal growth and fragmentation in an invasive seaweed. Ecology 2006, 87, 1744–1754. [Google Scholar] [CrossRef]
- Smith, C.M.; Walters, L.J. Fragmentation as a strategy for Caulerpa species: Fates of fragments and implications for management of an invasive weed. Mar. Ecol. 1999, 20, 307–319. [Google Scholar] [CrossRef]
- Holmer, M.; Marbà, N.; Lamote, M.; Duarte, C.M. Deterioration of Sediment Quality in Seagrass Meadows (Posidonia oceanica) Invaded by Macroalgae (Caulerpa sp.). Estuaries Coasts 2009, 32, 456–466. [Google Scholar] [CrossRef]
- Cebrian, E.; Linares, C.; Marchal, C.; Garrabou, J. Exploring the effects of invasive algae on the persistence of gorgonian populations. Biol. Invasions 2012, 14, 2647–2656. [Google Scholar] [CrossRef]
- Box, A.; Deudero, S.; Sureda, A.; Blanco, A.; Alòs, J.; Terrados, J.; Grau, A.M.; Riera, F. Diet and physiological responses of Spondyliosoma cantharus (Linnaeus, 1758) to the Caulerpa racemosa var. cylindracea invasion. J. Exp. Mar. Biol. Ecol. 2009, 380, 11–19. [Google Scholar] [CrossRef]
- Gupta, M.P.; Gómez, N.E.; Santana, A.I.; Solis, P.N.; Palacios, G. Antimicrobial activity of various algae of the Panamanian Atlantic coast. Rev. Med. Panama 1991, 16, 64–68. [Google Scholar] [PubMed]
- Dawczynski, C. Amino acids, fatty acids, and dietary fibre in edible seaweed products. Food Chem. 2007, 103, 891–899. [Google Scholar] [CrossRef]
- Stabili, L.; Acquaviva, M.I.; Biandolino, F.; Cavallo, R.A.; De Pascali, S.A.; Fanizzi, F.P.; Narracci, M.; Cecere, E.; Petrocelli, A. Biotechnological potential of the seaweed Cladophora rupestris (Chlorophyta, Cladophorales) lipidic extract. New Biotechnol. 2014, 31, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Luengo, J.M.; Garcia, B.; Sandoval, A.; Naharro, G.; Olivera, E.R. Bioplastics from microorganisms. Curr. Opin. Microbiol. 2003, 6, 251–260. [Google Scholar] [CrossRef]
- Pachekoski, W.M.; Marcondes Agnelli, J.A.; Belem, L.P. Thermal, mechanical and morphological properties of poly (hydrohybutyrate) and polypropylene blends after processing. Mater. Res. 2009, 12, 159–164. [Google Scholar] [CrossRef]
- Noreen, A.; Zia, K.M.; Zuber, M.; Ali, M.; Mujahid, M. A critical review of algal biomass: A versatile platform of bio-based polyesters from renewable resources. Int. J. Biol. Macromol. 2016, 86, 937–949. [Google Scholar] [CrossRef] [PubMed]
- De Pascali, S.A.; Del Coco, L.; Felline, S.; Mollo, E.; Terlizzi, A.; Fanizzi, F.P. 1H NMR spectroscopy and MVA analysis of Diplodus sargus eating the exotic pest Caulerpa cylindracea. Mar. Drugs 2015, 13, 3550–3566. [Google Scholar] [CrossRef] [PubMed]
- Schmid, M.; Guihéneuf, F.; Stengel, D. Fatty acid contents and profiles of 16 macroalgae collected from the Irish Coast at two seasons. J. Appl. Phycol. 2014, 26, 451–463. [Google Scholar] [CrossRef]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Stabili, L.; Acquaviva, M.I.; Biandolino, F.; Cavallo, R.A.; De Pascali, S.A.; Fanizzi, F.P.; Narracci, M.; Petrocelli, A.; Cecere, E. The lipidic extract of the seaweed Gracilariopsis longissima (Rhodophyta, Gracilariales): A potential resource for biotechnological purposes? New Biotechnol. 2012, 29, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Pridgeon, J.W.; Klesius, P.H. Major bacterial diseases in aquaculture and their vaccine development. Anim. Sci. Rev. 2012, 7, 1–16. [Google Scholar] [CrossRef]
- Sudheesh, P.S.; Al-Ghabshi, A.; Al-Mazrooei, N.; Al-Habsi, S. Comparative pathogenomics of bacteria causing infectious diseases in fish. Int. J. Evol. Biol. 2012, 2012, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Labreuche, Y.; Soudant, P.; Gonçalves, M.; Lambert, C.; Nicolas, J.L. Effects of extracellular products from the pathogenic Vibrio aestuarianus strain 01/32 on lethality and cellular immune responses of the oyster Crassostrea gigas. Dev. Comp. Immunol. 2006, 30, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Kushmaro, A.; Banin, E.; Loya, Y.; Stackebrandt, E.; Rosenberg, E. Vibrio shiloi sp. nov., the causative agent of bleaching of the coral Oculina patagonica. Int. J. Syst. Evol. Microbiol. 2001, 51, 1383–1388. [Google Scholar] [CrossRef] [PubMed]
- Strom, M.S.; Paranjpye, R.N. Epidemiology and pathogenesis of Vibrio vulnificus. Microbes Infect. 2000, 2, 177–188. [Google Scholar] [CrossRef]
- Vatsos, I.N.; Rebours, C. Seaweed extracts as antimicrobial agents in aquaculture. J. Appl. Phycol. 2015, 27, 2017–2035. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekaran, M.; Venkatesalu, V.; Raj, G.A. Antibacterial activity of selected marine macro algae against vancomycin resistant Enterococcus faecalis. J. Coast. Life Med. 2014, 2, 940–946. [Google Scholar]
- Salem, W.M.; Galal, H.; Nasr El-deen, F. Screening for antibacterial activities in some marine algae from the red sea (Hurghada, Egypt). Afr. J. Microbiol. Res. 2011, 5, 2160–2167. [Google Scholar] [CrossRef]
- Kindleysides, S.; Quek, S.Y.; Miller, M.R. Inhibition of fish oil oxidation and the radical scavenging activity of New Zealand sea weed extracts. Food Chem. 2012, 133, 1624–1631. [Google Scholar] [CrossRef]
- Frankel, E.N.; Meyer, A.S. The problems of using one-dimensional methods to evaluate multifunctional food and biological antioxidants. J. Sci. Food Agric. 2000, 80, 1925–1941. [Google Scholar] [CrossRef]
- Prior, R.L.; Cao, G. In vivo total antioxidant capacity: Comparison of different analytical methods. Free Radic. Biol. Med. 1999, 27, 1173–1181. [Google Scholar] [CrossRef]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef] [PubMed]
- Lakmal, H.C.; Samarakoon, K.W.; Lee, W.; Lee, J.H.; Abeytunga, D.T.U.; Lee, H.S.; Jeon, Y.J. Anticancer and antioxidant effects of selected Sri Lankan marine algae. J. Natl. Sci. Found. Sri Lanka 2014, 42, 315–323. [Google Scholar] [CrossRef]
- Zulueta, A.; Esteve, M.A.; Frígola, A. ORAC and TEAC assay comparison to measure the antioxidant capacity of food product. Food Chem. 2009, 114, 310–316. [Google Scholar] [CrossRef]
- Ou, B.; Huang, D.; Hampsch-Woodill, M.; Flanagan, J.A.; Deemer, E.K. Analysis of antioxidant activities of common vegetables employing oxygen radical absorbance capacity (ORAC) and ferric reducing antioxidant power (FRAP) assays: A comparative study. J. Agric. Food Chem. 2002, 50, 3122–3128. [Google Scholar] [CrossRef] [PubMed]
- Endo, Y.; Usuki, R.; Kaneda, T. Antioxidant effects of chlorophyll and pheophytin on the autooxidation of oils in the dark. II. The mechanism of antioxidative action of chlorophyll. J. Am. Oil Chem. Soc. 1985, 62, 1387–1390. [Google Scholar] [CrossRef]
- Gutierrez-Rosales, F.; Garrido-Fernandez, J.; Gallardo-Guerrero, L.; Gandul-Rojas, B.; Mınguez-Mosquera, M.I. Action of chlorophylls on the stability of the virgin olive oil. J. Am. Oil Chem. Soc. 1992, 69, 866–871. [Google Scholar] [CrossRef]
- Le Tutour, B.; Brunel, C.; Quemeneur, F. Synergistic effect of chlorophyll a on the oxidation properties of vitamin E. New J. Chem. 1996, 20, 707–721. [Google Scholar]
- Le Tutour, B.; Benslimane, F.; Gouleau, M.P.; Gouygou, J.P.; Saadan, B.; Quemeneur, F. Antioxidant and pro-oxidant activities of the brown algae, Laminaria digitata, Himanthalia elongata, Fucus vesiculosus, Fucus serratus and Ascophyllum nodosum. J. Appl. Phycol. 1998, 19, 121–129. [Google Scholar] [CrossRef]
- Pires-Cavalcante, K.M.D.S.; de Alencar, D.B.; de Sousa, M.B.; Sampaio, A.H.; Saker-Sampaio, S. Seasonal changes of a-tocopherol in green marine algae (Caulerpa genus). J. Food Sci. 2011, 76, C775–C781. [Google Scholar] [CrossRef] [PubMed]
- Ornano, L.; Donno, Y.; Sanna, C.; Ballero, M.; Serafini, M.; Bianco, A. Phytochemical study of Caulerpa racemosa (Forsk.) j. Agarth, an invading alga in the habitat of La Maddalena Archipelago. Nat. Prod. Res. 2014, 28, 1795–1799. [Google Scholar] [CrossRef] [PubMed]
- Matanjun, P.; Mohamed, S.; Mustapha, N.M.; Muhammad, K.; Ming, C.H. Antioxidant activities and phenolics content of eight species of seaweeds from north Borneo. J. Appl. Phycol. 2008, 20, 367–373. [Google Scholar] [CrossRef] [Green Version]
- Vinayak, R.C.; Sudha, S.A.; Chatterji, A. Bio-screening of a few green seaweeds from India for their cytotoxic and antioxidant potential. J. Sci. Food Agric. 2011, 91, 2471–2476. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, B.; Zhang, Q.; Qu, Y.; Xu, H.; Li, G. Preparation and antioxidant property of extract and semipurified fractions of Caulerpa racemosa. J. Appl. Phycol. 2012, 24, 1527–1536. [Google Scholar] [CrossRef]
- Anderson, L.W. California’s reaction to Caulerpa taxifolia: A model for invasive species rapid response. Biol. Invasions 2005, 7, 1003–1016. [Google Scholar] [CrossRef]
- Piazzi, L.; Ceccherelli, G.; Cinelli, F. Threat to macroalgal diversity: Effects of the introduced green alga Caulerpa racemosa in the Mediterranean. Mar. Ecol. Prog. Ser. 2001, 210, 161–165. [Google Scholar] [CrossRef]
- Ruitton, S.; Verlaque, M.; Boudouresque, C.F. Seasonal changes of the introduced Caulerpa racemosa var. cylindracea (Caulerpales, Chlorophyta) at the northwest limit of its Mediterranean distribution. Aquat. Bot. 2005, 82, 55–70. [Google Scholar] [CrossRef]
- Klein, J.; Verlaque, M. The Caulerpa racemosa invasion: A critical review. Mar. Pollut. Bull. 2008, 56, 205–225. [Google Scholar] [CrossRef] [PubMed]
- Guillén, J.; Jiménez, S.; Martinez, J.; Triviño, A.; Múgica, Y.; Argilés, J.; Bueno, M. Expansion of the invasive algae Caulerpa racemosa var. cylindracea (Sonder) Verlaque, Huisman & Boudouresque, 2003 on the region of Valencia seabed. Thalassas 2010, 26, 135–149. [Google Scholar]
- Verlaque, M.; Boudouresque, C.F.; Meinesz, A.; Gravez, V. The Caulerpa racemosa complex (Caulerpales, Ulvophyceae) in the Mediterranean Sea. Bot. Mar. 2000, 43, 49–68. [Google Scholar] [CrossRef]
- Kientz, B.; Thabard, M.; Cragg, S.M.; Pope, J.; Hellio, C. A new method for removing microflora from macroalgal surfaces: An important step for natural product discovery. Bot. Mar. 2011, 54, 457–469. [Google Scholar] [CrossRef]
- Standal, I.B.; Axelson, A.E.; Aursand, M. Differentiation of fish oils according to species by 13C-NMR regiospecific analyses of triacyglycerols. J. Am. Oil Chem. Soc. 2009, 86, 401–407. [Google Scholar] [CrossRef]
- Del Coco, L.; Papadia, P.; De Pascali, S.A.; Bressani, G.; Storelli, C.; Zonno, V.; Fanizzi, F.P. Comparison among different gilthead sea bream (Sparus aurata) farming systems: Activity of intestinal and hepatic enzymes and 13C-NMR analysis of lipids. Nutrients 2009, 1, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Kamal, C.; Sethuraman, M.G. Caulerpin—A bis-Indole Alkaloid as a Green Inhibitor for the Corrosion of Mild Steel in 1 M HCl Solution from the Marine Alga Caulerpa racemosa. Ind. Eng. Chem. Res. 2012, 51, 10399–10407. [Google Scholar] [CrossRef]
- Dávalos, A.; Gómez-Cordovés, C.; Bartolomé, B. Extending applicability of the oxygen radical absorbance capacity (ORAC-fluorescein) assay. J. Agric. Food Chem. 2004, 52, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improvement ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Cavallo, R.A.; Stabili, L. Presence of vibrios in seawater and Mytilus galloprovincialis (Lam.) from the Mar Piccolo of Taranto (Ionian Sea). Water Res. 2002, 36, 3719–3726. [Google Scholar] [CrossRef]
- Stabili, L.; Acquaviva, M.I.; Cavallo, R.A. Mytilus galloprovincialis filter feeding on the bacterial community in a Mediterranean coastal area (Northern Ionian Sea, Italy). Water Res. 2005, 39, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Turcck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [PubMed]
- Cavallo, R.; Acquaviva, M.; Stabili, L.; Cecere, E.; Petrocelli, A.; Narracci, M. Antibacterial activity of marine macroalgae against fish pathogenic Vibrio species. Open Life Sci. 2013, 8, 646–653. [Google Scholar] [CrossRef]
- Ely, R.; Supriya, T.; Naik, C.G. Antimicrobial activity of marine organisms collected off the coast of South East India. J. Exp. Biol. Ecol. 2004, 309, 121–127. [Google Scholar] [CrossRef]
- Kim, I.H.; Lee, D.G.; Lee, S.H.; Ha, J.M.; Ha, B.J.; Kim, S.K.; Lee, J.H. Antibacterial activity of Ulva lactuca against methicillin resistant Staphylococcus aureus (MRSA). Biotechnol. Bioprocess Eng. 2007, 112, 579–582. [Google Scholar] [CrossRef]
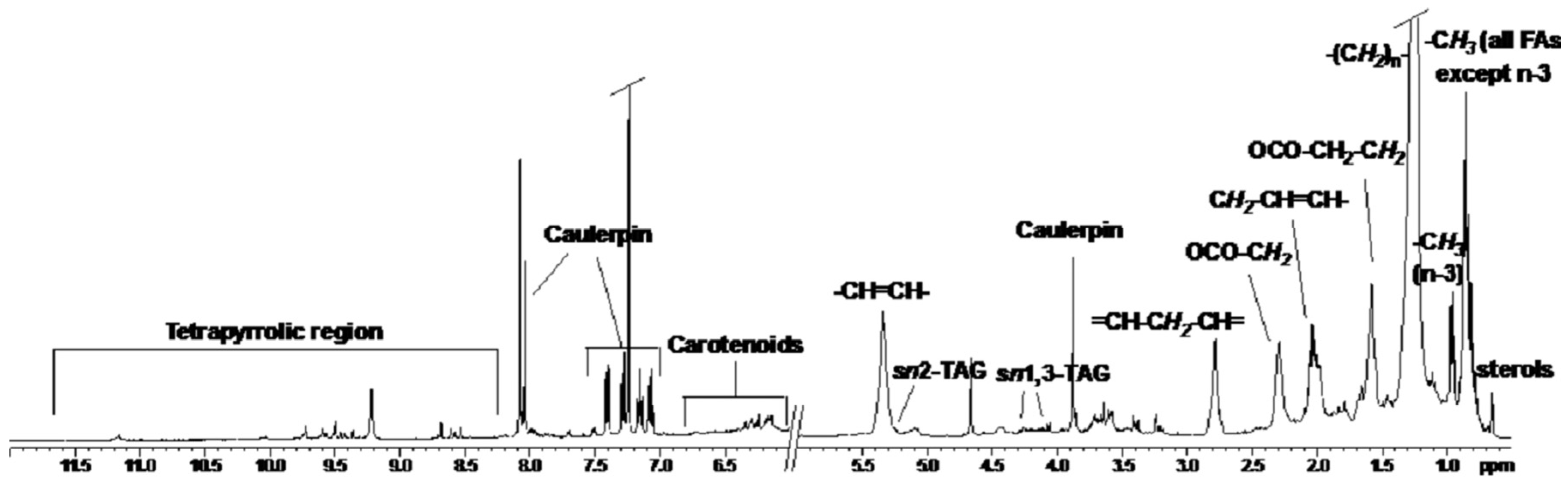

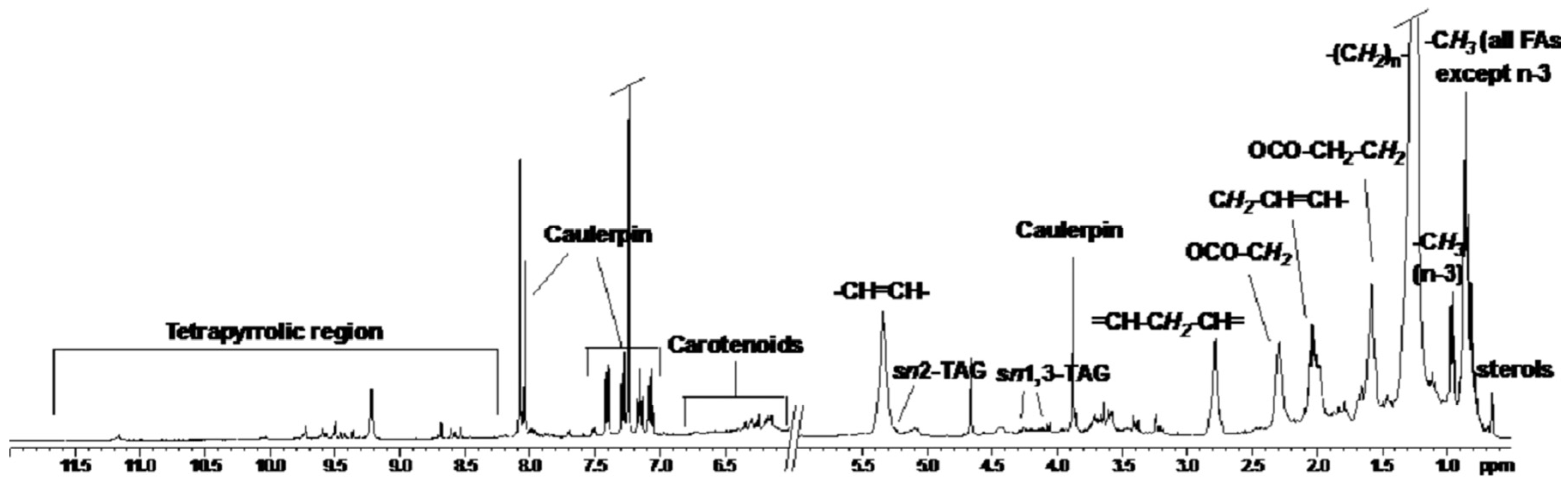{kind=link}
| ORAC value | 552.14 ± 69.13 μmol Trolox Equivalent/g extract |
| TEAC value | 70.03 ± 2.67 μmol Trolox Equivalent/g extract |
| Microbial Strain | Diameter of Growth Inhibition (cm) | ||||||
|---|---|---|---|---|---|---|---|
| 10 μL | 20 μL | 30 μL | 40 μL | 60 μL | 80 μL | 100 μL | |
| Candida albicans | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Candida glabrata | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Enterococcus sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Escherichia coli | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Staphilococcus sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Streptococcus sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Vibrio aestuarianus | 0.7 | 0.7 | 0.7 | 0.7 | 0.7 | 0.8 | 0.85 |
| Vibrio campbelli | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Vibrio carchariae | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Vibrio diazotrophicus | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Vibrio fischeri | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 | 0.9 |
| Vibrio fluvialis | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Vibrio furnissi | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Vibrio harveyi | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Vibrio inusitatus | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 | 0.9 | 0.9 |
| Vibrio litoralis | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 | 0.9 |
| Vibrio mediterranei | 0 | 0 | 0.7 | 0.7 | 0.7 | 0.8 | 0.8 |
| Vibrio natriegens | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Vibrio ordalii | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Vibrio salmonicida | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Vibrio splendidus II | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Vibrio vulnificus | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 | 0.8 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stabili, L.; Fraschetti, S.; Acquaviva, M.I.; Cavallo, R.A.; De Pascali, S.A.; Fanizzi, F.P.; Gerardi, C.; Narracci, M.; Rizzo, L. The Potential Exploitation of the Mediterranean Invasive Alga Caulerpa cylindracea: Can the Invasion Be Transformed into a Gain? Mar. Drugs 2016, 14, 210. https://doi.org/10.3390/md14110210
Stabili L, Fraschetti S, Acquaviva MI, Cavallo RA, De Pascali SA, Fanizzi FP, Gerardi C, Narracci M, Rizzo L. The Potential Exploitation of the Mediterranean Invasive Alga Caulerpa cylindracea: Can the Invasion Be Transformed into a Gain? Marine Drugs. 2016; 14(11):210. https://doi.org/10.3390/md14110210
Chicago/Turabian StyleStabili, Loredana, Simonetta Fraschetti, Maria Immacolata Acquaviva, Rosa Anna Cavallo, Sandra Angelica De Pascali, Francesco Paolo Fanizzi, Carmela Gerardi, Marcella Narracci, and Lucia Rizzo. 2016. "The Potential Exploitation of the Mediterranean Invasive Alga Caulerpa cylindracea: Can the Invasion Be Transformed into a Gain?" Marine Drugs 14, no. 11: 210. https://doi.org/10.3390/md14110210