
Polycyclic Guanidine Alkaloids from Poecilosclerida Marine Sponges

Abstract
:
1. Introduction
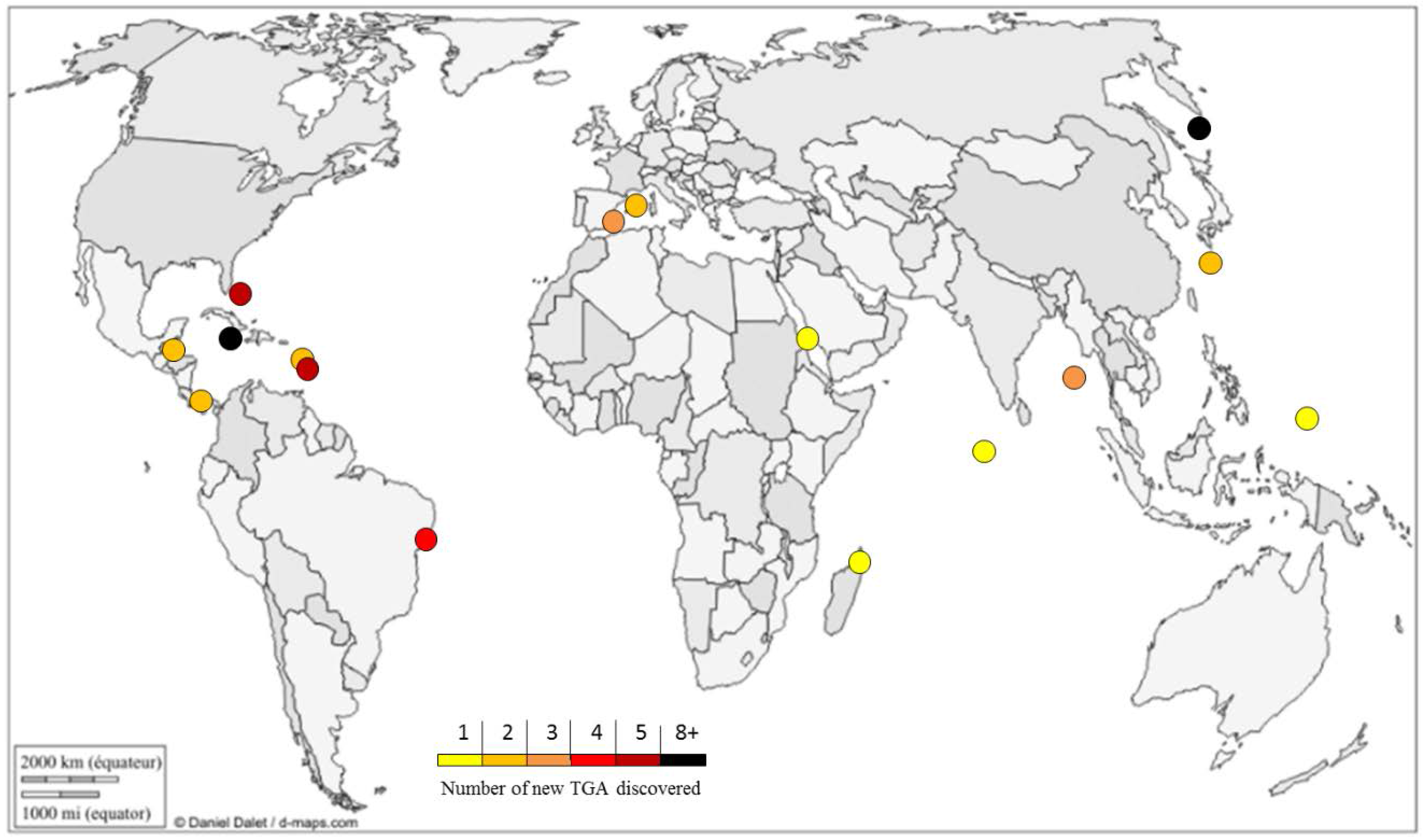
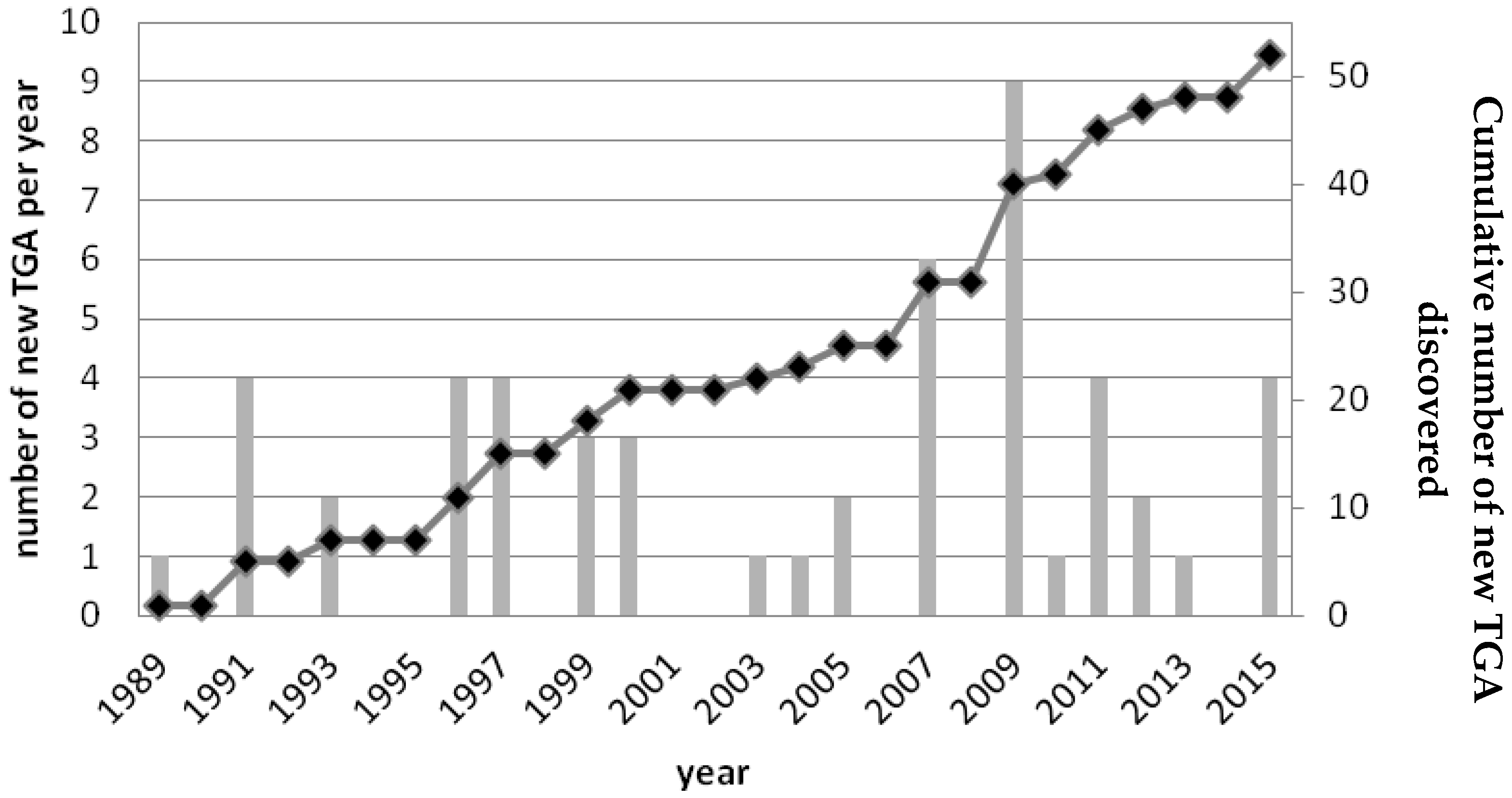
2. Structural Diversity
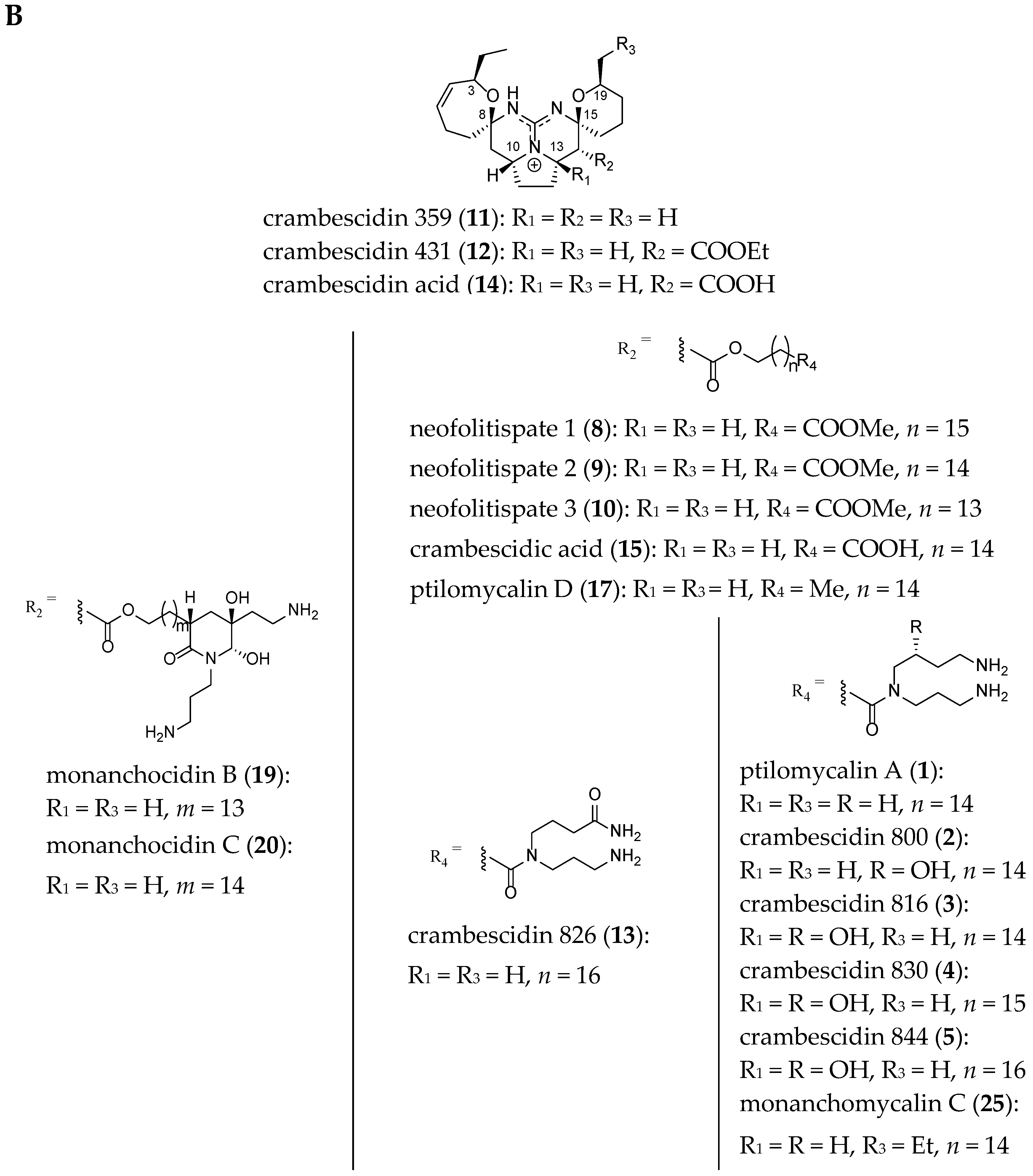
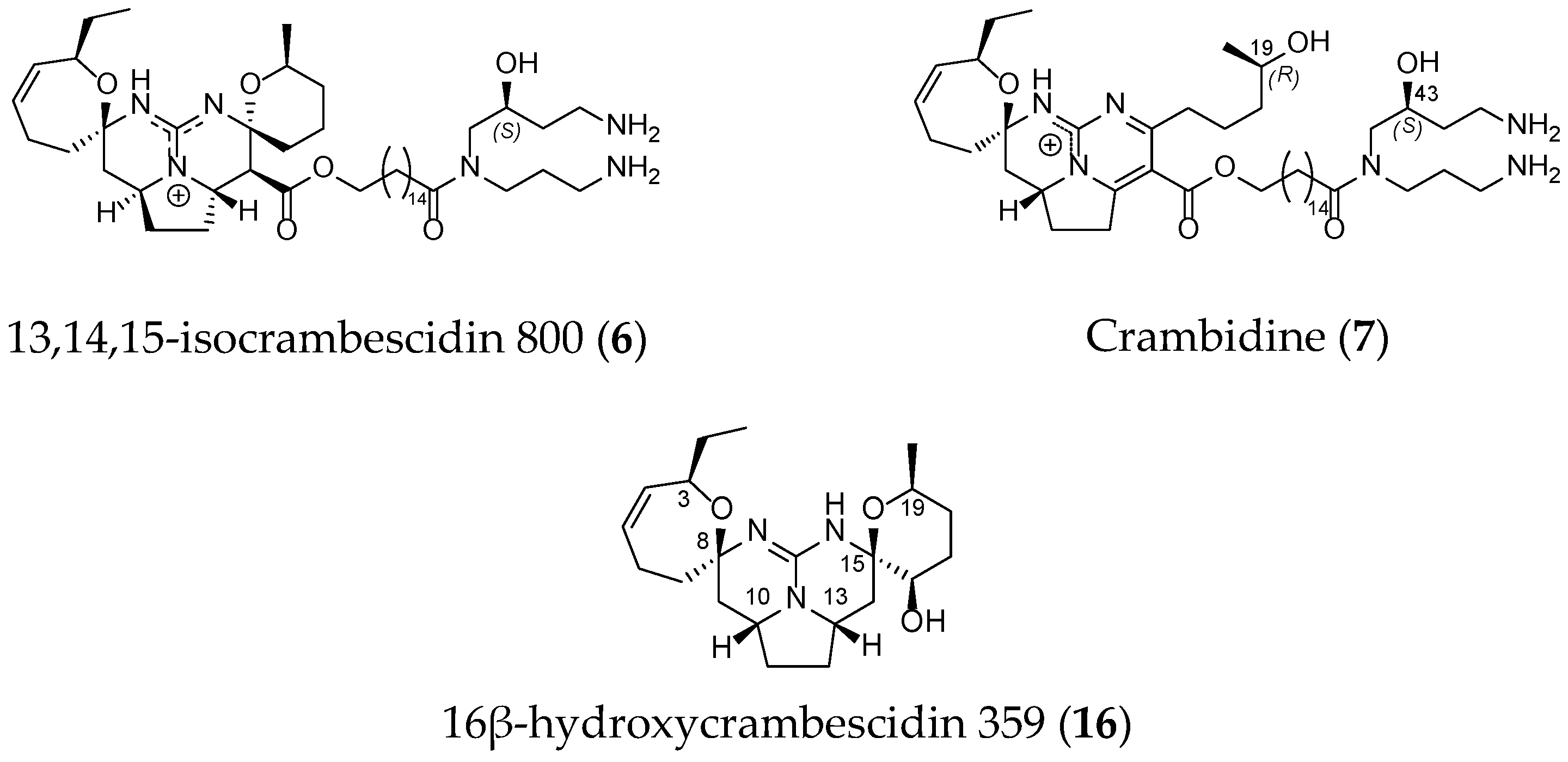
2.1. TGA Structures
2.2. TGA Classification
2.3. Analytical Tools for TGA Structural Analysis
2.3.1. NMR Spectroscopy
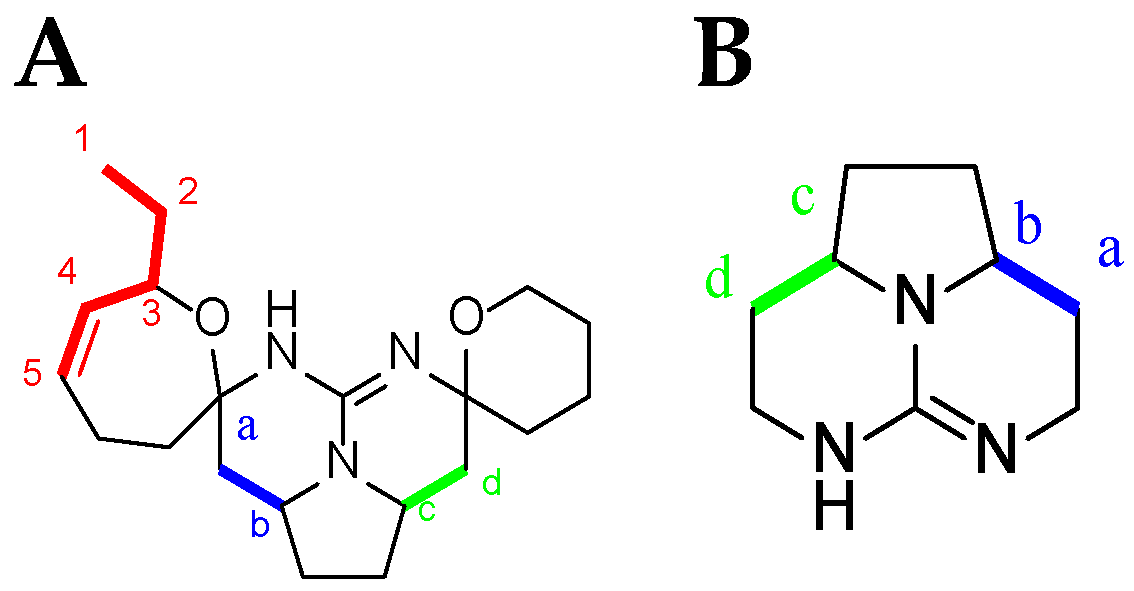
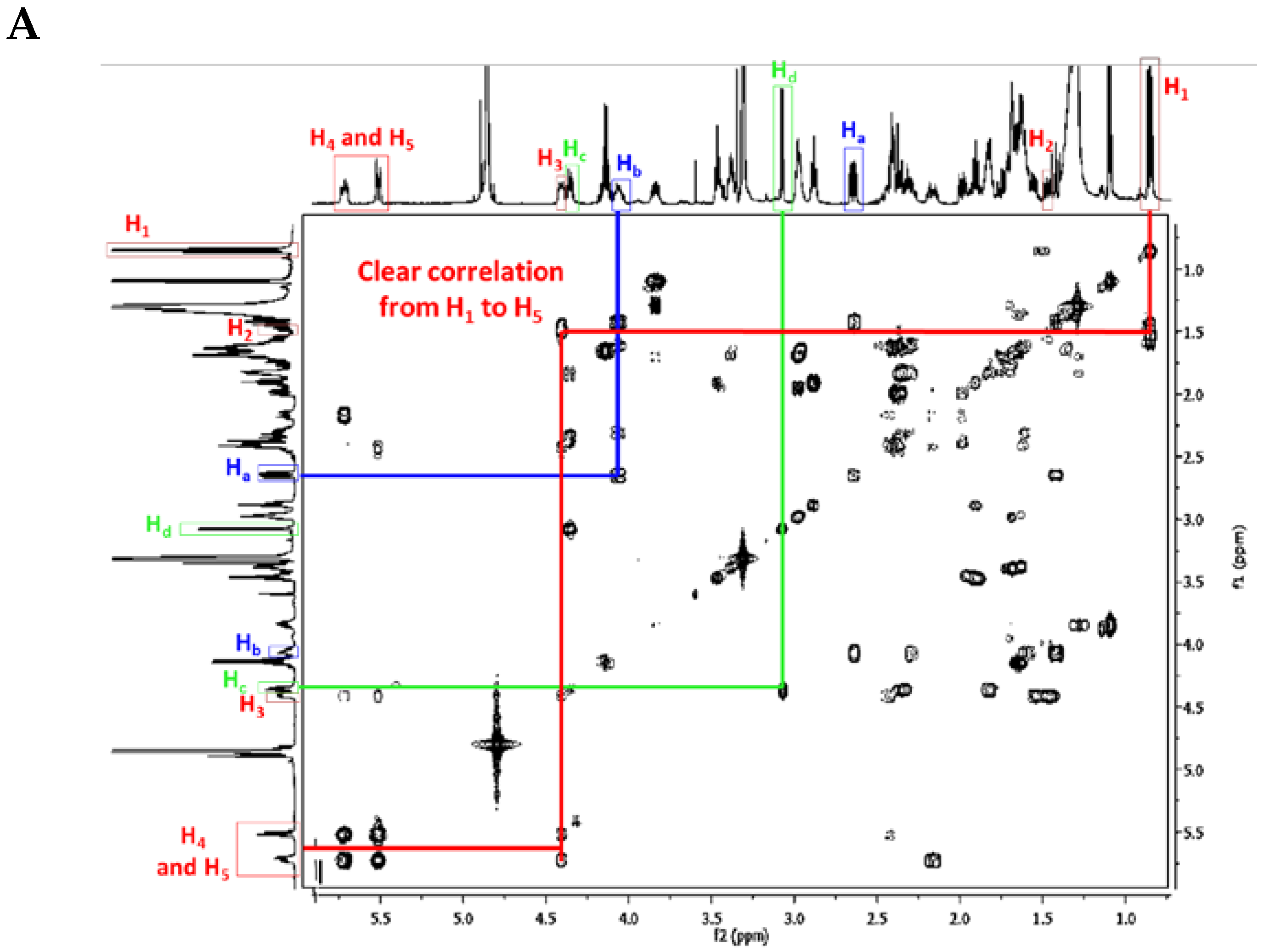
Crambescidin Case: Ptilomycalin A (1)
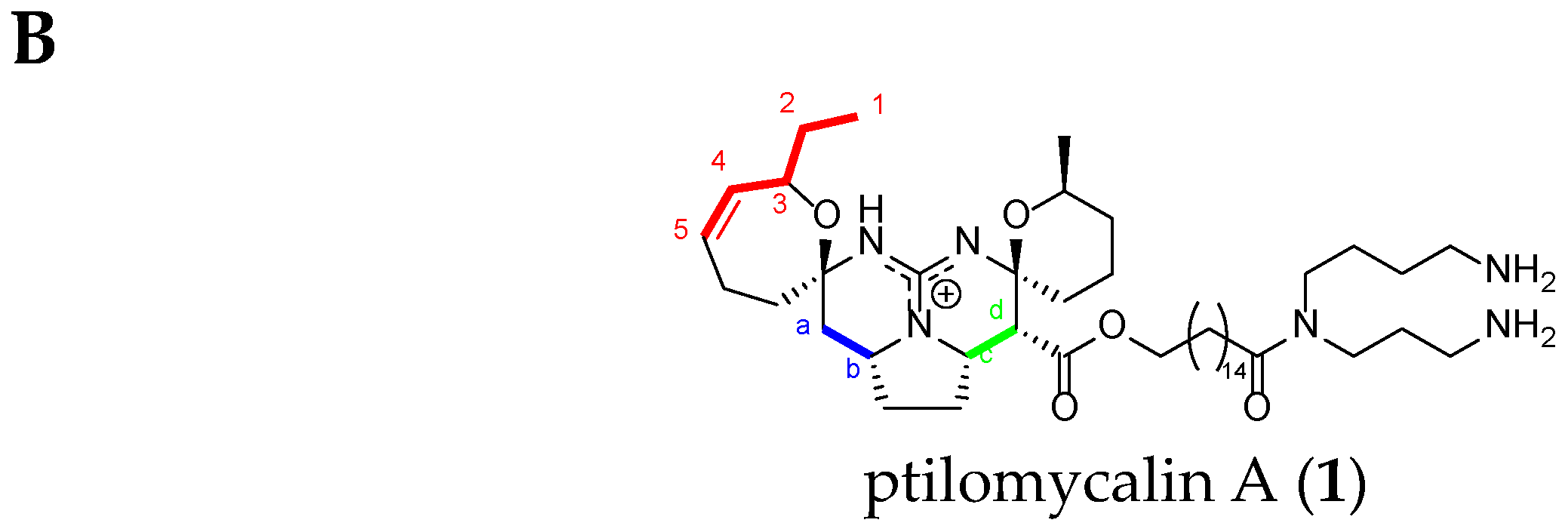
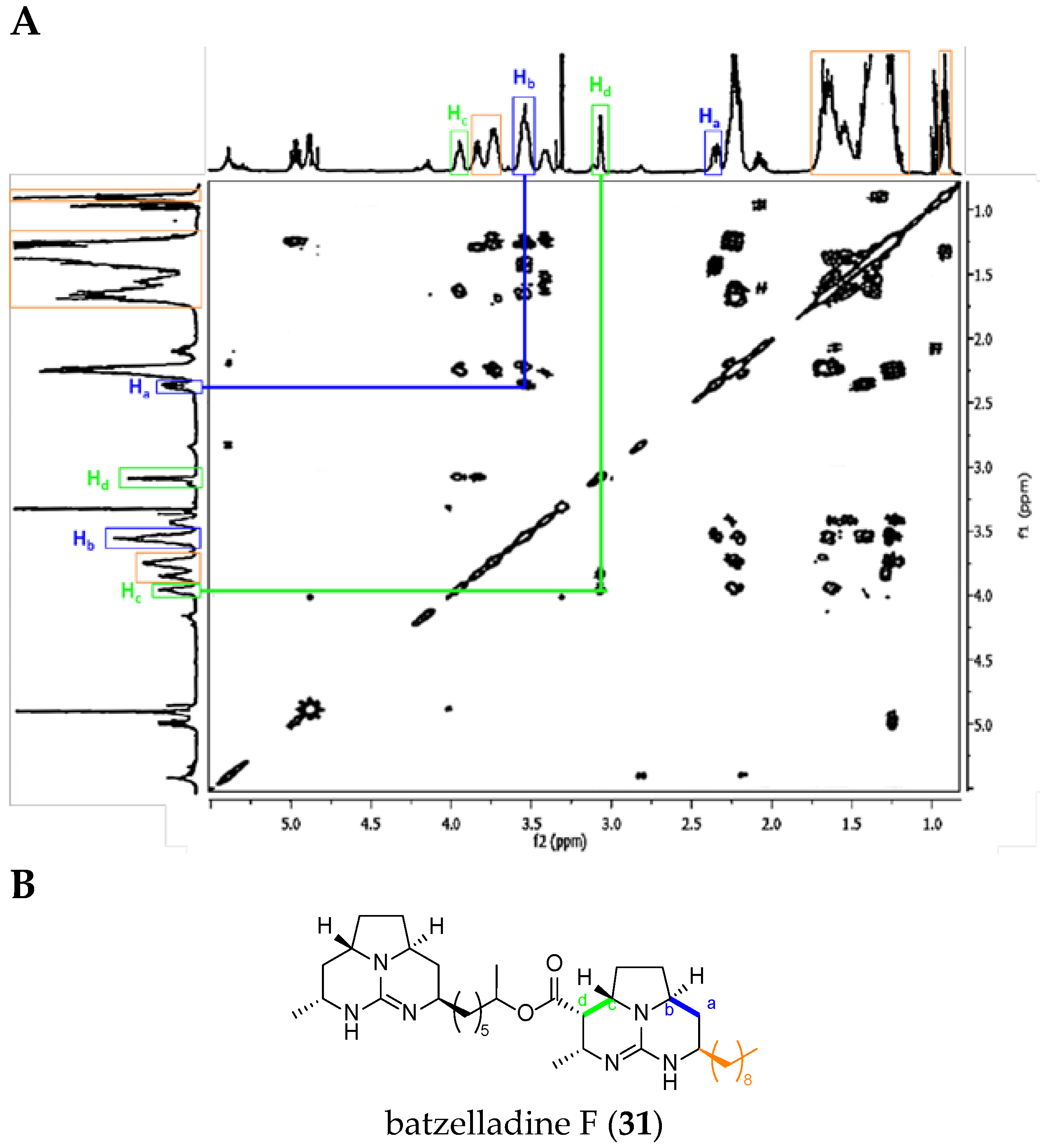
Batzelladine Case: Batzelladine F (31)
2.3.2. Mass Spectrometry
3. Biological Activities
3.1. Antiviral Activities
3.2. Antimicrobial Activities
3.3. Antitumoral Activities
4. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| A.m. | Antimicrobial activity |
| A.v. | Antiviral activity |
| A.t. | Antitumor activity |
| COSY | Correlation Spectroscopy |
| EC50 | half maximal Effective Concentration |
| GA | Guanidine Alkaloids |
| GI50 | half maximal Growth Inhibition |
| gp120 | glycoprotein 120 |
| HBV | Human hepatitis B virus |
| HIV-1 | Human Immunodeficiency Virus 1 |
| HSV-1 | Herpes simplex Virus 1 |
| IC50 | half maximal Inhibitory Concentration |
| LC50 | half maximal Lethal Concentration |
| MIC | Minimum Inhibitory Concentration |
| MS | Mass Spectrometry |
| NMR | Nuclear Magnetic Resonance |
| n.t. | Not tested |
| PBMC | Peripheral Blood Mononuclear Cells |
| SM | Secondary Metabolites |
| TC50 | half maximal Toxic Concentration |
| TGA | Triazaacenaphthylene Guanidine Alkaloids |
| TGI | Total Growth Inhibition concentration |
References
- Nijampatnam, B.; Dutta, S.; Velu, S.E. Recent advances in isolation, synthesis, and evaluation of bioactivities of bispyrroloquinone alkaloids of marine origin. Chin. J. Nat. Med. 2015, 13, 561–577. [Google Scholar] [CrossRef]
- Gul, W.; Hamann, M.T. Indole alkaloid marine natural products: An established source of cancer drug leads with considerable promise for the control of parasitic, neurological and other diseases. Life Sci. 2005, 78, 442–453. [Google Scholar] [CrossRef] [PubMed]
- Forte, B.; Malgesini, B.; Piutti, C.; Quartieri, F.; Scolaro, A.; Papeo, G. A submarine journey: The pyrrole-imidazole alkaloids. Mar. Drugs 2009, 7, 705–753. [Google Scholar] [CrossRef] [PubMed]
- Weinreb, S.M. Some recent advances in the synthesis of polycyclic imidazole-containing marine natural products. Nat. Prod. Rep. 2007, 24, 931–948. [Google Scholar] [CrossRef] [PubMed]
- Berlinck, R.G.S. Natural guanidine derivatives. Nat. Prod. Rep. 2002, 19, 617–649. [Google Scholar] [CrossRef] [PubMed]
- Berlinck, R.G.S.; Burtoloso, A.C.; Kossuga, M.H. The chemistry and biology of organic guanidine derivatives. Nat. Prod. Rep. 2008, 25, 919–954. [Google Scholar] [CrossRef] [PubMed]
- Berlinck, R.G.S.; Burtoloso, A.C.; Trindade-Silva, A.E.; Romminger, S.; Morais, R.P.; Bandeira, K.; Mizuno, C.M. The chemistry and biology of organic guanidine derivatives. Nat. Prod. Rep. 2010, 27, 1871–1907. [Google Scholar] [CrossRef] [PubMed]
- Berlinck, R.G.S.; Kossuga, M.H. Natural guanidine derivatives. Nat. Prod. Rep. 2005, 22, 516–550. [Google Scholar] [CrossRef] [PubMed]
- Berlinck, R.G.S. Natural guanidine derivatives. Nat. Prod. Rep. 1999, 16, 339–365. [Google Scholar] [CrossRef]
- Faulkner, J.D. Marine natural products. Nat. Prod. Rep. 2001, 18, 1–49. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2015, 32, 116–211. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Hu, W.-P.; Munro, M.H.G.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2007, 24, 31–86. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Hu, W.-P.; Munro, M.H.G.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2008, 25, 35–94. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Hu, W.-P.; Munro, M.H.G.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2009, 26, 170–244. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Munro, M.H.G.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2010, 27, 165–237. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Munro, M.H.G.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2011, 28, 196–268. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2012, 29, 144–222. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2013, 30, 237–323. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2014, 31, 160–258. [Google Scholar] [CrossRef] [PubMed]
- Berlinck, R.G.S.; Trindade-Silva, A.E.; Santos, M.F. The chemistry and biology of organic guanidine derivatives. Nat. Prod. Rep. 2012, 29, 1382–1406. [Google Scholar] [CrossRef] [PubMed]
- Snider, B.B.; Shi, Z. Biomimetic synthesis of the central tricyclic portion of ptilomycalin A. Tetrahedron Lett. 1993, 34, 2099–2102. [Google Scholar] [CrossRef]
- Rao, A.V.R.; Gurjar, M.K.; Vasudevan, J. An enantiospecific synthesis of the tricyclic guanidine segment of the anti-HIV marine alkaloid batzelladine A. J. Chem. Soc. Chem. Commun. 1995, 13, 1369–1370. [Google Scholar] [CrossRef]
- Louwrier, S.; Ostendorf, M.; Boom, A.; Hiemstra, H.; Speckamp, W.N. Studies towards the synthesis of (+)-ptilomycalin A; stereoselective N-acyliminium ion coupling reactions to enantiopure C-2 substituted lactams. Tetrahedron 1996, 52, 2603–2628. [Google Scholar] [CrossRef]
- Louwrier, S.; Ostendorf, M.; Tuynman, A.; Hiemstra, H. Studies towards the synthesis of guanidine alkaloids; synthesis of a tricyclic guanidine from succinimide. Tetrahedron Lett. 1996, 37, 905–908. [Google Scholar] [CrossRef]
- Louwrier, S.; Tuynman, A.; Hiemstra, H. Synthesis of bicyclic guanidines from pyrrolidin-2-one. Tetrahedron 1996, 52, 2629–2646. [Google Scholar] [CrossRef]
- Murphy, P.J.; Lloyd Williams, H.; Hibbs, D.E.; Hursthouse, M.B.; Malik, A.K.M. Biomimetic model studies towards ptilomycalin A. Tetrahedron 1996, 52, 8315–8332. [Google Scholar] [CrossRef]
- Snider, B.B.; Chen, J.; Patil, A.D.; Freyer, A.J. Synthesis of the tricyclic portions of batzelladines A, B and D. Revision of the stereochemistry of batzelladines A and D. Tetrahedron Lett. 1996, 37, 6977–6980. [Google Scholar] [CrossRef]
- Black, G.P.; Murphy, P.J.; Walshe, N.D.A. A short synthetic route to the tricyclic guanidinium core of the batzelladine alkaloids. Tetrahedron 1998, 54, 9481–9488. [Google Scholar] [CrossRef]
- Black, G.P.; Murphy, P.J.; Thornhill, A.J.; Walshe, N.D.A.; Zanetti, C. Synthesis of the left hand unit of batzelladine F; revision of the reported relative stereochemistry. Tetrahedron 1999, 55, 6547–6554. [Google Scholar] [CrossRef]
- Snider, B.B.; Busuyek, M.V. Revision of the stereochemistry of batzelladine F. Approaches to the tricyclic hydroxyguanidine moiety of batzelladines G, H, and I. J. Nat. Prod. 1999, 62, 1707–1711. [Google Scholar] [CrossRef] [PubMed]
- Coffey, D.S.; McDonald, A.I.; Overman, L.E.; Rabinowitz, M.H.; Renhowe, P.A. A practical entry to the crambescidin family of guanidine alkaloids. Enantioselective total syntheses of ptilomycalin A, crambescidin 657 and its methyl ester (neofolitispates 2), and crambescidin 800. J. Am. Chem. Soc. 2000, 122, 4893–4903. [Google Scholar] [CrossRef]
- Elliott, M.C.; Long, M.S. Studies towards the total synthesis of batzelladine A. Org. Biomol. Chem 2004, 2, 2003–2011. [Google Scholar] [CrossRef] [PubMed]
- Aron, Z.D.; Overman, L.E. Total synthesis and properties of the crambescidin core zwitterionic acid and crambescidin 359. J. Am. Chem. Soc. 2005, 127, 3380–3390. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Pierce, J.G. Synthesis of the 5,6-dihydroxymorpholin-3-one fragment of monanchocidin A. Org. Lett. 2015, 17, 968–971. [Google Scholar] [CrossRef] [PubMed]
- Snider, B.B.; Chen, J. Synthesis of batzelladine E and its E isomer. Tetrahedron Lett. 1998, 39, 5697–5700. [Google Scholar] [CrossRef]
- Coffey, D.S.; McDonald, A.I.; Overman, L.E.; Stappenbeck, F. Enantioselective total synthesis of 13,14,15-isocrambescidin 800. J. Am. Chem. Soc. 1999, 121, 6944–6945. [Google Scholar] [CrossRef]
- Cohen, F.; Overman, L.E.; Ly Sakata, S.K. Asymmetric total synthesis of batzelladine D. Org. Lett. 1999, 1, 2169–2172. [Google Scholar] [CrossRef] [PubMed]
- Franklin, A.S.; Ly, S.K.; Mackin, G.H.; Overman, L.E.; Shaka, A.J. Application of the tethered biginelli reaction for enantioselective synthesis of batzelladine alkaloids. Absolute configuration of the tricyclic guanidine portion of batzelladine B. J. Org. Chem. 1999, 64, 1512–1519. [Google Scholar] [CrossRef] [PubMed]
- Cohen, F.; Overman, L.E. Enantioselective total synthesis of batzelladine F: Structural revision and stereochemical definition. J. Am. Chem. Soc. 2001, 123, 10782–10783. [Google Scholar] [CrossRef] [PubMed]
- Ishiwata, T.; Hino, T.; Koshino, H.; Hashimoto, Y.; Nakata, T.; Nagasawa, K. Total synthesis of batzelladine D. Org. Lett. 2002, 4, 2921–2924. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, K.; Hashimoto, Y. Synthesis of marine guanidine alkaloids and their application as chemical/biological tools. Chem. Rec. 2003, 3, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Aron, Z.D.; Overman, L.E. The tethered biginelli condensation in natural product synthesis. Chem. Commun. 2004, 3, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Collins, S.K.; McDonald, A.I.; Overman, L.E.; Ho Rhee, Y. Enantioselective total synthesis of (−)-dehydrobatzelladine C. Org. Lett. 2004, 6, 1253–1255. [Google Scholar] [CrossRef] [PubMed]
- Arnold, M.A.; Duron, S.G.; Gin, D.Y. Diastereoselective [4+2] annulation of vinyl carbodiimides with N-alkyl imines. Asymmetric synthetic access to the batzelladine alkaloids. J. Am. Chem. Soc. 2005, 127, 6924–6925. [Google Scholar] [CrossRef] [PubMed]
- Overman, L.E.; Ho Rhee, Y. Total synthesis of (−)-crambidine and definition of the relative configuration of its unique tetracyclic guanidinium core. J. Am. Chem. Soc. 2005, 125, 15652–15658. [Google Scholar] [CrossRef] [PubMed]
- Shimokawa, J.; Ishiwata, T.; Shirai, K.; Koshino, H.; Tanatani, A.; Nakata, T.; Hashimoto, Y.; Nagasawa, K. Total synthesis of (+)-batzelladine A and (−)-batzelladine D, and identification of their target protein. Chemistry 2005, 11, 6878–6888. [Google Scholar] [CrossRef] [PubMed]
- Evans, P.A.; Qin, J.; Robinson, J.E.; Bazin, B. Enantioselective total synthesis of the polycyclic guanidine-containing marine alkaloid (−)-batzelladine D. Angew. Chem. 2007, 46, 7417–7419. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.G.; Murphy, P.J.; Williams, H.L.; McGown, A.T.; Smith, N.K. Synthetic studies towards ptilomycalin A: Total synthesis of crambescidin 359. Tetrahedron 2007, 63, 11771–11780. [Google Scholar] [CrossRef]
- Perl, N.R.; Ide, N.D.; Prajapati, S.; Perfect, H.; Duro, S.G.; Gin, D.Y. Annulation of thioimidates and vinyl carbodiimides to prepare 2-aminopyrimidines, competent nucleophiles for intramolecular alkyne hydroamination. Synthesis of (−)-crambidine. J. Am. Chem. Soc. 2010, 132, 1802–1803. [Google Scholar] [CrossRef] [PubMed]
- Babij, N.R.; Wolfe, J.P. Asymmetric total synthesis of (+)-merobatzelladine B. Angew. Chem. 2012, 51, 4128–4130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashman, Y.; Hirsh, S.; McConnell, O.J.; Ohtani, I.; Kusumi, T.; Kakisawa, H. Ptilomycalin A: A novel polycyclic guanidine alkaloid of marine origin. J. Am. Chem. Soc. 1989, 111, 8926–8928. [Google Scholar] [CrossRef]
- Berlinck, R.G.S.; Braekman, J.C.; Daloze, D.; Bruno, I.; Riccio, R. Polycyclic guanidine alkaloids from the marine sponge Crambe crambe and Ca++ channel blocker activity of crambescidin 816. J. Nat. Prod. 1993, 56, 1007–1015. [Google Scholar] [CrossRef] [PubMed]
- Braekman, J.C.; Daloze, D.; Tavares, R.; Hajdu, E.; Van Soest, R.W.M. Novel polycyclic guanidine alkaloids from two marine sponges of the genus Monanchora. J. Nat. Prod. 2000, 63, 193–196. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Whittaker, N.F.; Bewley, C.A. Crambescidin 826 and dehydrocrambine A: New polycyclic guanidine alkaloids from the marine sponge Monanchora sp. That inhibit HIV-1 fusion. J. Nat. Prod. 2003, 66, 1490–1494. [Google Scholar] [CrossRef] [PubMed]
- Jares-Erijman, E.A.; Sakai, R.; Rinehart, K.L. Crambescidins: New antiviral and cytotoxic compounds from the sponge Crambe crambe. J. Org. Chem. 1991, 56, 5712–5715. [Google Scholar] [CrossRef]
- Jares-Erijman, E.A.; Ingrum, A.L.; Carney, J.R.; Rinehart, K.L.; Sakai, R. Polycyclic guanidine-containing compounds from the mediterranean sponge Crambe crambe: The structure of 13,14,15-isocrambescidin 800 and the absolute stereochemistry of the pentacyclic guanidine moieties of the crambescidins. J. Org. Chem. 1993, 58, 4805–4808. [Google Scholar] [CrossRef]
- Venkateswarlu, Y.; Venkata Rami Reddy, M.; Ramesh, P.; Venkateswara Rao, J. Neofolitispates, pentacyclic guanidine alkaloids from the sponge Neofolitispa dianchora. Indian J. Chem. 1999, 38B, 254–256. [Google Scholar]
- Meragelman, K.M.; McKee, T.C.; McMahon, J.B. Monanchorin, a bicyclic alkaloid from the sponge Monanchora ungiculata. J. Nat. Prod. 2004, 67, 1165–1167. [Google Scholar] [CrossRef] [PubMed]
- Gallimore, W.A.; Kelly, M.; Scheuer, P.J. Alkaloids from the sponge Monanchora unguifera. J. Nat. Prod. 2005, 68, 1420–1423. [Google Scholar] [CrossRef] [PubMed]
- Bensemhoun, J.; Bombarda, I.; Aknin, M.; Vacelet, J.; Gaydou, E.M. Ptilomycalin D, a polycyclic guanidine alkaloid from the marine sponge Monanchora dianchora. J. Nat. Prod. 2007, 70, 2033–2035. [Google Scholar] [CrossRef] [PubMed]
- Guzii, A.G.; Makarieva, T.N.; Denisenko, V.A.; Dmitrenok, P.S.; Kuzmich, A.S.; Dyshlovoy, S.A.; Krasokhin, V.B.; Stonik, V.A. Monanchocidin: A new apoptosis-inducing polycyclic guanidine alkaloid from the marine sponge Monanchora pulchra. Org. Lett. 2010, 12, 4292–4295. [Google Scholar] [CrossRef] [PubMed]
- Makarieva, T.N.; Tabakmaher, K.M.; Guzii, A.G.; Denisenko, V.A.; Dmitrenok, P.S.; Shubina, L.K.; Kuzmich, A.S.; Lee, H.S.; Stonik, V.A. Monanchocidins B–E: Polycyclic guanidine alkaloids with potent antileukemic activities from the sponge Monanchora pulchra. J. Nat. Prod. 2011, 74, 1952–1958. [Google Scholar] [CrossRef] [PubMed]
- Makarieva, T.N.; Tabakmaher, K.M.; Guzii, A.G.; Denisenko, V.A.; Dmitrenok, P.S.; Kuzmich, A.S.; Lee, H.-S.; Stonik, V.A. Monanchomycalins A and B, unusual guanidine alkaloids from the sponge Monanchora pulchra. Tetrahedron Lett. 2012, 53, 4228–4231. [Google Scholar] [CrossRef]
- Tabakmakher, K.M.; Denisenko, V.A.; Guzii, A.G.; Dmitrenok, P.S.; Dyshlovoy, S.A.; Lee, H.-S.; Makarieva, T.N. Monanchomycalin C, a new pentacyclic guanidine alkaloid from the far-eastern marine sponge Monanchora pulchra. Nat. Prod. Com. 2013, 8, 1399–1402. [Google Scholar]
- Palagiano, E.; De Marino, S.; Minale, L.; Riccio, R.; Zollo, F. Ptilomycalin A, crambescidin 800 and related new highly cytotoxic guanidine alkaloids from the starfishes Fromia monilis and Celerina heffernani. Tetrahedron 1995, 51, 3675–3682. [Google Scholar] [CrossRef]
- Patil, A.D.; Kumar, N.V.; Kokke, W.C.; Bean, M.F.; Freyer, A.J.; De Brosse, C.; Mai, S.; Truneh, A.; Faulkner, D.J.; Carte, B.; et al. Novel alkaloids from the sponge Batzella sp.: Inhibitors of HIV gp120-human CD4 binding. J. Org. Chem. 1995, 60, 1182–1188. [Google Scholar] [CrossRef]
- Patil, A.D.; Freyer, A.J.; Taylor, P.B.; Carte, B.; Johnson, R.K.; Faulkner, D.J. Batzelladines F-I, novel alkaloids from the sponge Batzella sp.: Inducers of p56lck-CD4 dissociation. J. Org. Chem. 1997, 62, 1814–1819. [Google Scholar] [CrossRef]
- Laville, R.; Thomas, O.P.; Berrue, F.; Marquez, D.; Vacelet, J.; Amade, P. Bioactive guanidine alkaloids from two caribbean marine sponges. J. Nat. Prod. 2009, 72, 1589–1594. [Google Scholar] [CrossRef] [PubMed]
- Takishima, S.; Ishiyama, A.; Iwatsuki, M.; Otoguro, K.; Yamada, H.; Omura, S.; Kobayashi, H.; van Soest, R.W.M.; Matsunaga, S. Merobatzelladines A and B, anti-infective tricyclic guanidines from a marine sponge Monanchora sp. Org. Lett. 2009, 11, 2655–2658. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.F.; Harper, P.M.; Williams, D.E.; Mesquita, J.T.; Pinto, E.G.; da Costa-Silva, T.A.; Hajdu, E.; Ferreira, A.G.; Santos, R.A.; Murphy, P.J.; et al. Anti-parasitic guanidine and pyrimidine alkaloids from the marine sponge Monanchora arbuscula. J. Nat. Prod. 2015, 78, 1101–1112. [Google Scholar] [CrossRef] [PubMed]
- Kornprobst, J.-M. Encyclopedia of Marine Natural Products, 2nd ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2014; Volume 2, pp. 869–877. [Google Scholar]
- Daniel Dalet. Planisphère centré Europe Afrique. Available online: http://d-maps.com/carte.php ?num_car=126803&lang=fr (access on 5 November 2015).
- Hua, H.-M.; Peng, J.; Dunbar, D.C.; Schinazi, R.F.; de Castro Andrews, A.G.; Cuevas, C.; Garcia-Fernandez, L.F.; Kelly, M.; Hamann, M.T. Batzelladine alkaloids from the caribbean sponge Monanchora unguifera and the significant activities against HIV-1 and AIDS opportunistic infectious pathogens. Tetrahedron 2007, 63, 11179–11188. [Google Scholar] [CrossRef]
- Suna, H.; Aoki, S.; Setiawan, A.; Kobayashi, M. Crambescidin 800, a pentacyclic guanidine alkaloid, protects a mouse hippocampal cell line against glutamate-induced oxidative stress. J. Nat. Med. 2007, 61, 288–295. [Google Scholar] [CrossRef]
- Rubiolo, J.A.; Lopez-Alonso, H.; Roel, M.; Vieytes, M.R.; Thomas, O.; Ternon, E.; Vega, F.V.; Botana, L.M. Mechanism of cytotoxic action of crambescidin-816 on human liver-derived tumour cells. Br. J. Pharmacol. 2014, 171, 1655–1667. [Google Scholar] [CrossRef] [PubMed]
- Bewley, C.A.; Ray, S.; Cohen, F.; Collins, S.K.; Overman, L.E. Inhibition of HIV-1 envelope-mediated fusion by synthetic batzelladine analogues. J. Nat. Prod. 2004, 67, 1319–1324. [Google Scholar] [CrossRef] [PubMed]
- Overman, L.E.; Wolfe, J.P. Synthesis of polycyclic guanidines by cyclocondensation reactions of N-amidinyliminium ions. J. Org. Chem. 2001, 66, 3167–3175. [Google Scholar] [CrossRef] [PubMed]
- Rubiolo, J.A.; Ternon, E.; Lopez-Alonso, H.; Thomas, O.P.; Vega, F.V.; Vieytes, M.R.; Botana, L.M. Crambescidin-816 acts as a fungicidal with more potency than crambescidin-800 and -830, inducing cell cycle arrest, increased cell size and apoptosis in Saccharomyces cerevisiae. Mar. Drugs 2013, 11, 4419–4434. [Google Scholar] [CrossRef] [PubMed]
- Croue, J.; West, N.J.; Escande, M.L.; Intertaglia, L.; Lebaron, P.; Suzuki, M.T. A single betaproteobacterium dominates the microbial community of the crambescidine-containing sponge Crambe crambe. Sci. Rep. 2013, 3, 2583–2590. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | Species | Sampling Site | Discovery Year | Guanidine Moiety | Biological Activity | Synthesis Described | Ref. |
|---|---|---|---|---|---|---|---|
| ptilomycalin A (1) | Hemimycale sp. Ptilocaulis spiculifer | Red sea | 1989 | 1 | Av, Am, At | Yes | [42,51,73] |
| crambescidin 800 (2) | Crambe crambe | Palma de Mallorca, Mediterranean sea | 1991 | 1 | Av, Am, At | Yes | [54,55,56,73,74] |
| crambescidin 816 (3) | Crambe crambe | Palma de Mallorca, Mediterranean sea | 1991 | 1 | Av, At, Ca2+ antagonist | No | [52,55,56,75] |
| crambescidin 830 (4) | Crambe crambe | Palma de Mallorca, Mediterranean sea | 1991 | 1 | n.t. | No | [55] |
| crambescidin 844 (5) | Crambe crambe | Palma de Mallorca, Mediterranean sea | 1991 | 1 | Av | No | [55,56] |
| 13,14,15-isocrambescidine 800 (6) | Crambe crambe | Banyuls, Mediterranean sea | 1993 | 1 | Not active | Yes | [36,42,56] |
| crambidine (7) | Crambe crambe | Banyuls, Mediterranean sea | 1993 | 1 | n.t. | Yes | [45,49,52] |
| neofolitispate 1 (8) | Neofolitispa dianchora | Andaman Islands, Indian Ocean | 1999 | 1 | Av | No | [57] |
| neofolitispate 2 (9) | Neofolitispa dianchora | Andaman Islands, Indian Ocean | 1999 | 1 | Av | Yes | [57] |
| neofolitispate 3 (10) | Neofolitispa dianchora | Andaman Islands, Indian Ocean | 1999 | 1 | Av | No | [57] |
| crambescidin 359 (11) | Monanchora ungiculata | Belize, North Atlantic Ocean | 2000 | 1 | n.t. | Yes | [33,41,48,53] |
| crambescidin 431 (12) | Monanchora ungiculata | Belize, North Atlantic Ocean | 2000 | 1 | n.t. | No | [53] |
| crambescidin 826 (13) | Monanchora sp. | Palau, Pacific Ocean | 2003 | 1 | Av | No | [54] |
| crambescidin acid (14) | Monanchora ungiculata | Maldive Islands, Indian Ocean | 2004 | 1 | At | No | [58] |
| crambescidic acid (15) | Monanchora unguifera Monanchora dianchora | Panama, Caribbean side, Atlantic Ocean | 2005 | 1 | n.t. | No | [59] |
| 16β-hydroxycrambescidin 359 (16) | Monanchora unguifera | Jamaica, North Atlantic Ocean | 2007 | 1 | Am | No | [73] |
| ptilomycalin D (17) | Monanchora dianchora | Madagascar, Indian Ocean | 2007 | 1 | n.t. | No | [60] |
| monanchocidin A (18) | Monanchora pulchra | Urup Island, North Pacific Ocean | 2010 | 1 | At | No | [61] |
| monanchocidin B (19) | Monanchora pulchra | Urup Island, North Pacific Ocean | 2011 | 1 | At | No | [62] |
| monanchocidin C (20) | Monanchora pulchra | Urup Island, North Pacific Ocean | 2011 | 1 | At | No | [62] |
| monanchocidin D (21) | Monanchora pulchra | Urup Island, North Pacific Ocean | 2011 | 1 | At | No | [62] |
| monanchocidin E (22) | Monanchora pulchra | Urup Island, North Pacific Ocean | 2011 | 1 | At | No | [62] |
| monanchomycalin A (23) | Monanchora pulchra | Urup Island, North Pacific Ocean | 2012 | 1 | At | No | [63] |
| monanchomycalin B (24) | Monanchora pulchra | Urup Island, North Pacific Ocean | 2012 | 1 | n.t. | No | [63] |
| monanchomycalin C (25) | Monanchora pulchra | Urup Island, North Pacific Ocean | 2013 | 1 | n.t. | No | [64] |
| Metabolites | Species | Sampling Site | Discovery Year | Guanidine Moiety | Biological Activities | Synthesis Described | Ref. |
|---|---|---|---|---|---|---|---|
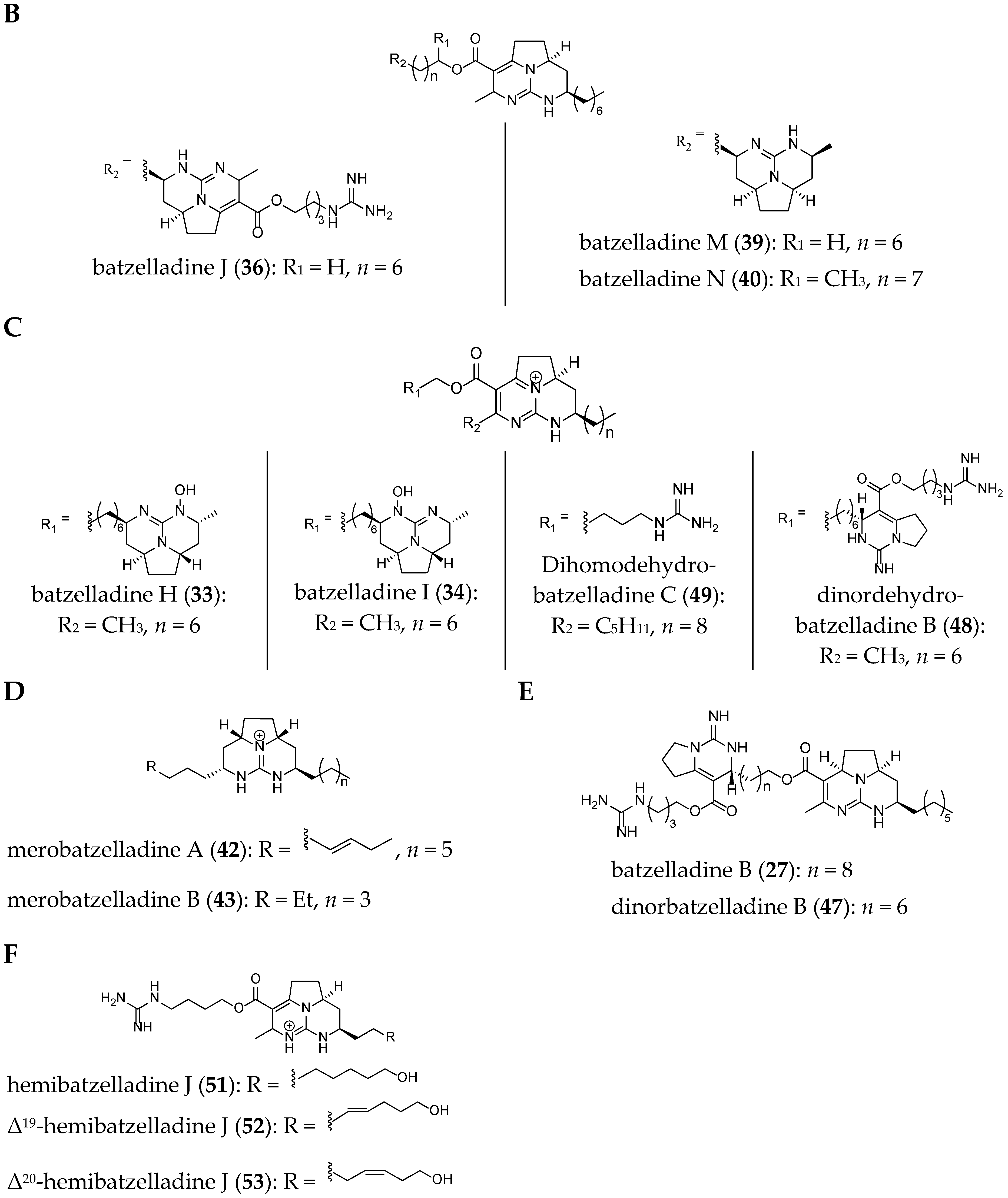
| batzelladine A (26) | Batzella sp. | Bahamas, North Atlantic Ocean | 1996 | 3 | Am, Av | Yes | [32,46,66] |
| batzelladine B (27) | Batzella sp. | Bahamas, North Atlantic Ocean | 1996 | 3 | Av | No | [66] |
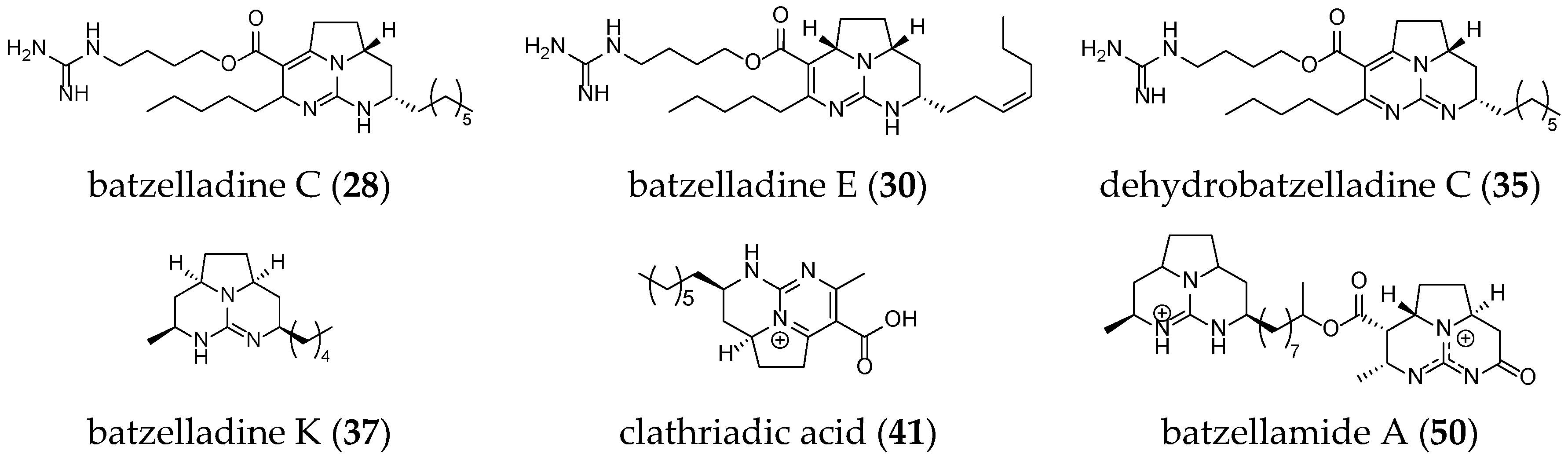
| batzelladine C (28) | Batzella sp. | Bahamas, North Atlantic Ocean | 1996 | 2 | Av, Am, At | No | [66,73] |
| batzelladine D (29) | Batzella sp. | Bahamas, North Atlantic Ocean | 1996 | 2 | Av, Am | Yes | [37,40,46,47,66] |
| batzelladine E (30) | Batzella sp. | Bahamas, North Atlantic Ocean | 1996 | 2 | n.t. | Yes | [35,66] |
| batzelladine F (31) | Batzella sp. | Jamaica, North Atlantic Ocean | 1997 | 2 | Am | Yes | [30,39,42,67] |
| batzelladine G (32) | Batzella sp. | Jamaica, North Atlantic Ocean | 1997 | 2 | Av | No | [67] |
| batzelladine H (33) | Batzella sp. | Jamaica, North Atlantic Ocean | 1997 | 2 | Av | No | [67] |
| batzelladine I (34) | Batzella sp. | Jamaica, North Atlantic Ocean | 1997 | 2 | Av | No | [67] |
| dehydrobatzelladine C (35) | Monanchora arbuscula | Belize, North Atlantic Ocean | 2000 | 2 | Av, Am, At | Yes | [43,53,73] |
| batzelladine J (36) | Monanchora unguifera | Panama, North Atlantic Ocean | 2005 | 3 | n.t. | No | [59] |
| batzelladine K (37) | Monanchora unguifera | Jamaica, North Atlantic Ocean | 2007 | 1 | n.t. | No | [73] |
| batzelladine L (38) | Monanchora unguifera | Jamaica, North Atlantic Ocean | 2007 | 2 | Av, Am, At | No | [73] |
| batzelladine M (39) | Monanchora unguifera | Jamaica, North Atlantic Ocean | 2007 | 2 | Av, Am, At | No | [73] |
| batzelladine N (40) | Monanchora unguifera | Jamaica, North Atlantic Ocean | 2007 | 2 | Av, At | No | [73] |
| clathriadic acid (41) | Clathria calla | Martinique, North Atlantic Ocean | 2009 | 1 | Am | No | [68] |
| merobatzelladine A (42) | Monanchora sp. | Amami-Oshima Island, North Pacific Ocean | 2009 | 1 | Am | No | [50,69] |
| merobatzelladine B (43) | Monanchora sp. | Amami-Oshima Island, North Pacific Ocean | 2009 | 1 | Am | Yes | [69] |
| norbatzelladine A (44) | Monanchora arbuscula | Guadeloupe Island, North Atlantic Ocean | 2009 | 3 | Am, At | No | [68] |
| norbatzelladine L (45) | Clathria calla | Martinique, North Atlantic Ocean | 2009 | 2 | Am, At | No | [68] |
| dinorbatzelladine A (46) | Monanchora arbuscula | Guadeloupe island, North Atlantic Ocean | 2009 | 3 | Am, At | No | [68] |
| dinorbatzelladine B (47) | Monanchora arbuscula | Guadeloupe island, North Atlantic Ocean | 2009 | 3 | n.t. | No | [68] |
| dinordehydrobatzelladine B (48) | Monanchora arbuscula | Guadeloupe island, North Atlantic Ocean | 2009 | 3 | Am, At | No | [68] |
| dihomodehydrobatzelladine C (49) | Monanchora arbuscula | Guadeloupe island, North Atlantic Ocean | 2009 | 2 | Am, At | No | [68] |
| batzellamide A (50) | Monanchora arbuscula | Rio de Janeiro state, South Atlantic Ocean | 2015 | 2 | n.t. | No | [70] |
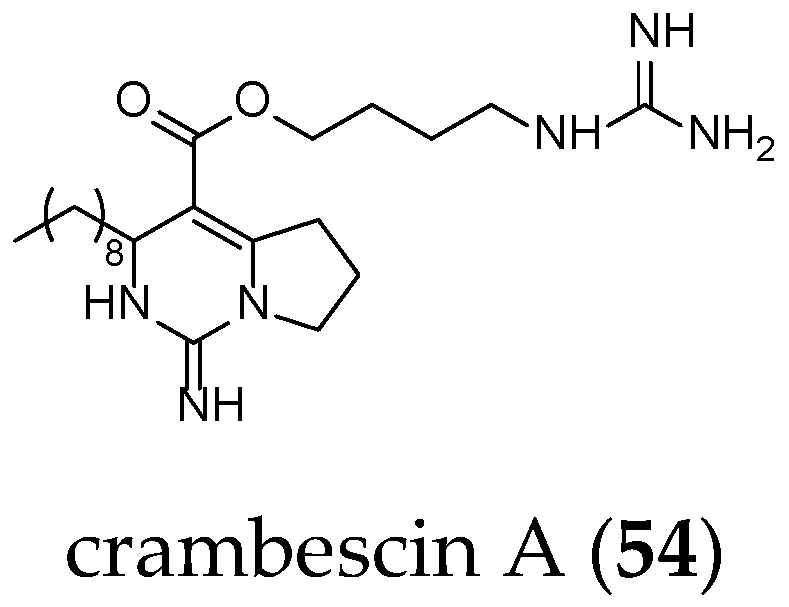
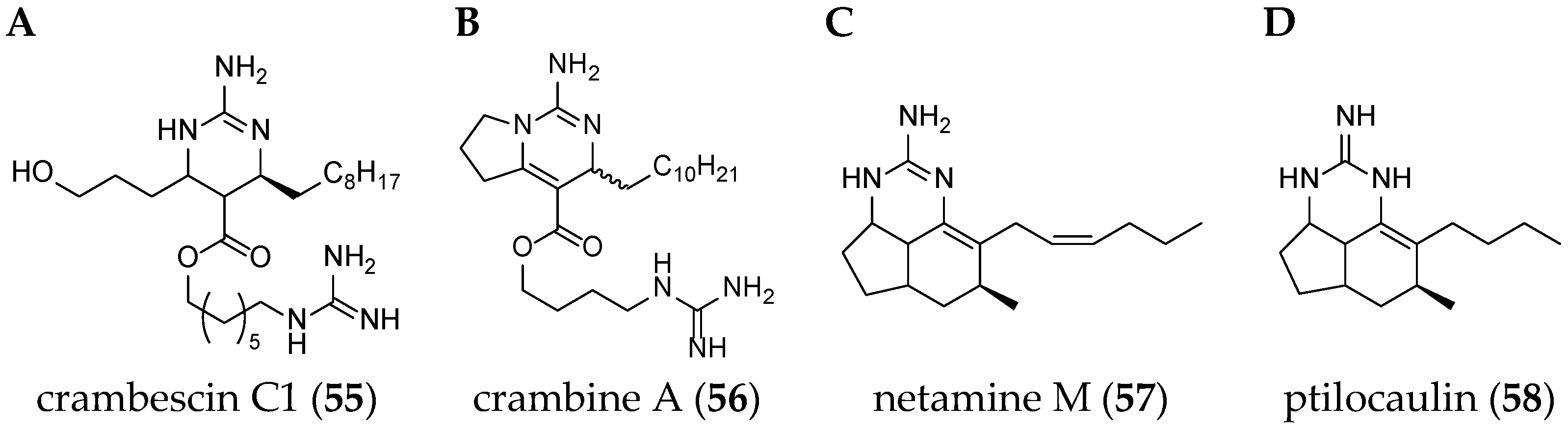
| hemibatzelladine J (51) | Monanchora arbuscula | Rio de Janeiro state, South Atlantic Ocean | 2015 | 2 | n.t. | No | [70] |
| Δ19-hemibatzelladine J (52) | Monanchora arbuscula | Rio de Janeiro state, South Atlantic Ocean | 2015 | 2 | n.t. | No | [70] |
| Δ200-hemibatzelladine J (53) | Monanchora arbuscula | Rio de Janeiro state, South Atlantic Ocean | 2015 | 2 | n.t. | No | [70] |
| No | Class 1 | Class 2 | Class 3 | Class 4 | Ref. |
|---|---|---|---|---|---|
| 1 |  |  | +1 or more guanidine moiety |  | [70] |
| Sponges * | Batzella Crambe Pseudaxinella (Ptilocaulis) | Clathria Crambe Hemimycale Monanchora Neofolitispa Ptilocaulis | Batzella Monanchora | Acanthella Arenochalina Batzella Biemna Clathria Monanchora Ptilocaulis | - |
| 2 | | |  n = 1 or 3 | | - |
| Sponges * | Batzella Crambe Pseudaxinella (Ptilocaulis) | Batzella Clathria Monanchora | Crambe Hemimycale Monanchora Neofolitispa Ptilocaulis | Acanthella Arenochalina Batzella Biemna Clathria Monanchora Ptilocaulis | - |
| Atom Number | Crambescidin-Like GA Signals | Batzelladine-Like GA Signals |
|---|---|---|
| Ha * | dd from 2.8 to 2.5 ppm | m from 2.8 to 2.5 ppm |
| Hb * | m from 4.6 to 3.9 ppm | m from 4.6 to 3.9 ppm |
| Hc * | dt toward 4.3 ppm | m toward 4.3 ppm |
| Hd * | d from 3.5 to 2.9 ppm | dd from 3.5 to 2.9 ppm |
| H4 * et H5 * double bond | 2 m toward 5.5 ppm | No signal |
| Metabolites | m/z ([M + H]+ Unless Specified) and Δppm Found | Ref. |
|---|---|---|
| ptilomycalin A (1) | 977.7915 (1.0 mmu) for the bis(trifluoroacetyl) derivative | [51] |
| crambescidin 800 (2) | 801.6205 (1.3 mmu) | [55] |
| crambescidin 816 (3) | 817.6151 (1.6 mmu) | [55] |
| crambescidin 830 (4) | 831.6300 (2.3 mmu) | [55] |
| crambescidin 844 (5) | 845.6471 (0.9 mmu) | [55] |
| 13, 14, 15 -isocrambescidine 800 (6) | 927.6521 (1.3 mmu) for the acetylated compound | [52] |
| crambidine (7) | 967.6415 (6.8 mmu) for the acetylated compound | [52] |
| neofolitispate 1 (8) | 686 (no HRMS data) | [57] |
| neofolitispate 2 (9) | 672 (no HRMS data) | [57] |
| neofolitispate 3 (10) | 658 (no HRMS data) | [57] |
| crambescidin 359 (11) | 359.2567 (0.6 mmu) | [53] |
| crambescidin 431 (12) | 431.2780 (0.4 mmu) | [53] |
| crambescidin 826 (13) | 827.6389 (1.5 mmu) | [54] |
| crambescidin acid (14) | 404.2541 (2.2 mmu) | [58] |
| crambescidic acid (15) | 658.4781 (1.4 mmu) | [59] |
| 16β-hydroxycrambescidin 359 (16) | 376.2617 (1.7 mmu) | [73] |
| ptilomycalin D (17) | 627.4994 * | [60] |
| monanchocidin (A) (18) | 859.6267 (3.0 mmu) | [61] |
| monanchocidin B (19) | 831.5978 (3.4 mmu) | [62] |
| monanchocidin C (20) | 845.6150 (4.0 mmu) | [62] |
| monanchocidin D (21) | 831.5920 (3.4 mmu) | [62] |
| monanchocidin E (22) | 845.6120 (1.0 mmu) | [62] |
| monanchomycalin A (23) | 813.6574 (0.2 mmu) | [63] |
| monanchomycalin B (24) | 785.6259 (0.4 mmu) | [63] |
| monanchomycalin C (25) | 813.6578 (0.3 mmu) and [M + 2H]2+ 407.3336 (0.7 mmu) | [64] |
| batzelladine A (26) | 768.5839 (2.4 mmu) | [66] |
| batzelladine B (27) | 738.5356 (3.8 mmu) | [66] |
| batzelladine C (28) | 489.3903 (1.4 mmu) | [66] |
| batzelladine D (29) | 463.3740 ** | [66] |
| batzelladine E (30) | 487.3728 (3.2 mmu) | [66] |
| batzelladine F (31) | 624.5096 (0.6 mmu) | [67] |
| batzelladine G (32) | 668,5353 (1.3 mmu) | [67] |
| batzelladine H (33) | 609.4488 (0.4 mmu) | [67] |
| batzelladine I (34) | [67] | |
| dehydrobatzelladine C (35) | 487.3711 (4.9 mmu) | [53] |
| batzelladine J (36) | 750.5361 (3.3 mmu) | [59] |
| batzelladine K (37) | 250.2322 (3.9 mmu) | [73] |
| batzelladine L (38) | 653.5458 (2.4 mmu) and [M + 2H]2+ 327.2798 (1.8 mmu) | [73] |
| batzelladine M (39) | [M + 2H]2+ 298.2399 (1.0 mmu) | [73] |
| batzelladine N (40) | [M + 2H]2+ 312.2546 (1.0 mmu) | [73] |
| clathriadic acid (41) | 318.2173 (1.0 mmu) | [68] |
| merobatzelladine A (42) | 360.3444 (6.6 mmu) | [69] |
| merobatzelladine B (43) | 306.2909 (7.0 mmu) | [69] |
| norbatzelladine A (44) | 754.5705 (0.7 mmu) | [68] |
| norbatzelladine L (45) | 639.5327 (0.3 mmu) | [68] |
| dinorbatzelladine A (46) | 740.5547 (0.9 mmu) | [68] |
| dinorbatzelladine B (47) | 710.5074 (1.2 mmu) | [68] |
| dinordehydrobatzelladine B (48) | 708.4919 (1.0 mmu) | [68] |
| dihomodehydrobatzelladine C (49) | 515.4064 (1.3 mmu) | [68] |
| batzellamide A (50) | 541.3878 (1.2 mmu) | [70] |
| hemibatzelladine J (51) | 449.3238 (0.2 mmu) | [70] |
| Δ19-hemibatzelladine J (52) | 447.3092 (0.8 mmu) | [70] |
| Δ200-hemibatzelladine J (53) | [70] |
| m/z Fragment | m/z Fragment Loss | Fragment | Ref. |
|---|---|---|---|
| 358 (or 359) | Crambescidin core | [53,54,55,59] | |
| 322 or 336 or 350 | Batzelladine core + n = 6, 7 or 8 carbon side chain | [53,66,67,68,73] | |
| 114 | 113 |  | [53,59,66] |
| 101 |  | [61,62] | |
| Intense 18 | 17 | Carboxylic acid | - |
| Intense 48 | 47 |
| EC50 (µM Unless Specified) | HIV-1 | HSV-1 | HBV | Ref. | ||
|---|---|---|---|---|---|---|
| Human PBMC | Envelope-Mediated Fusion | gp120 Binding to CD4 | ||||
| ptilomycalin A (1) | 0.011 | n.t. | n.t. | 0.25 * | n.t. | [51] |
| crambescidin 800 (2) | 0.04 | 1–3 | n.t. | 1.25 µg/well a | n.t. | [54,55,73] |
| crambescidin 816 (3) | n.t. | n.t. | n.t. | 1.25 µg/well a | n.t. | [55] |
| crambescidin 844 (5) | n.t. | n.t. | n.t. | 1.25 µg/well a | n.t. | [55] |
| 13,14,15-isocrambescidin 800 (6) | n.t. | n.t. | n.t. | NA | n.t. | [56] |
| neofolitispate 1 (8) | n.t. | n.t. | n.t. | n.t. | 7.4 ** | [57] |
| neofolitispate 2 (9) | n.t. | n.t. | n.t. | n.t. | ||
| neofolitispate 3 (10) | n.t. | n.t. | n.t. | n.t. | ||
| crambescidin 826 (13) | n.t. | 1–3 | n.t. | n.t. | n.t. | [54] |
| batzelladine A (26) | n.t. | n.t. | 29 | n.t. | n.t. | [66] |
| batzelladine B (27) | n.t. | n.t. | 31 | n.t. | n.t. | [66] |
| batzelladine C (28) | 7.7 | n.t. | n.t. | n.t. | n.t. | [73] |
| batzelladine D (29) | n.t. | n.t. | 72 | n.t. | n.t. | [66] |
| dehydrobatzelladine C (35) | 5.5 | n.t. | n.t. | n.t. | n.t. | [73] |
| batzelladine L (38) | 1.6 | n.t. | n.t. | n.t. | n.t. | [73] |
| batzelladine M (39) | 7.7 | n.t. | n.t. | n.t. | n.t. | [73] |
| batzelladine N (40) | 2.4 | n.t. | n.t. | n.t. | n.t. | [73] |
| IC50 (Values are Expressed in µg/mL Unless Specified) | Bacteria | Yeast | Fungi | Parasites | Ref. | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S. aureus | MRSA | P. aeruginosa | M. tuberculosis | M. intracellulare | V. anguillarum | C. albicans | C. neoformans | A. fumigatus (AC) | P. falciparum | L. infatum | L. donovani | T. cruzi | T. brucei brucei | ||||
| D6 Clone | W2 Clone | FcB1 | |||||||||||||||
| ptilomycalin A (1) | 0.25 | 0.30 | 1.0 | >128 | 10 | n.t. | 0.15 | 0.10 | 1.25 | 0.12 | 0.11 | 0.08 * | n.t. | 5.9 | n.t. | n.t. | [68,73] |
| crambescidin 800 (2) | 0.20 | 0.35 | 0.95 | 46.5 | 15 | n.t. | 0.15 | 0.10 | 1.25 | 0.11 | 0.13 | n.t. | n.t. | 6.80 | n.t. | n.t. | [73] |
| 16β-hydroxycrambescidin 800 (16) | NA | NA | NA | >128 | NA | n.t. | NA | NA | NA | 3.8 | NA | NA | n.t. | n.t. | n.t. | n.t. | [73] |
| batzelladine A (26) | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | 0.2* | n.t. | n.t. | n.t. | n.t. | [68] |
| batzelladine C (28) | 0.20 | 0.30 | 10 | 34.7 | 0.9 | n.t. | 0.90 | 0.40 | 5.0 | 0.09 | 0.11 | n.t. | n.t. | 5.5 | n.t. | n.t. | [73] |
| batzelladine D (29) | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | 0.9 * | n.t. | 29 * | n.t. | [70] |
| batzelladine F (31) | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | 2.5 * | n.t. | 3.1 * | n.t. | [70] |
| dehydrobatzelladine C (35) | 0.40 | 0.70 | NA. | 37.7 | 1.0 | n.t. | 1.0 | 0.6 | 20 | 0.073 | 0.13 | n.t. | n.t. | 5.70 | n.t. | n.t. | [73] |
| batzelladine L (38) | 0.35 | 0.40 | 3.50 | 1.68 | 0.25 | n.t. | 0.40 | 0.55 | 2.5 | 0.073 | 0.10 | 0.2 * | 1.3 * | 1.90 | 1.3 * | n.t. | [68,73] |
| batzelladine M (39) | 3.0 | 5.0 | NA | 28.5 | 3.50 | n.t. | 6.0 | 8.0 | NA | 0.21 | 0.27 | n.t. | n.t. | 8.50 | n.t. | n.t. | [73] |
| batzelladine N (40) | n.t. | n.t. | n.t. | 3.18 | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | [73] |
| clathriadic acid (41) | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | 1.4 * | n.t. | n.t. | n.t. | n.t. | [68] |
| merobatzelladine A (42) | n.t. | n.t. | n.t. | n.t. | n.t. | a | n.t. | n.t. | n.t. | 0.48 | n.t. | n.t. | n.t. | 0.24 | [69] | ||
| merobatzelladine B (43) | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | 0.97 | n.t. | n.t. | n.t. | 0.24 | [69] | |||
| norbatzelladine A (44) | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | 0.2 * | n.t. | n.t. | n.t. | n.t. | [68] |
| norbatzelladine L (45) | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | 0.3 * | 1.3 * | n.t. | 4.4 * | n.t. | [68,70] |
| dinorbatzelladine A (46) | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | 1.7 * | n.t. | n.t. | n.t. | n.t. | [68] |
| dinordehydrobatzelladine B (48) | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | 0.6 * | n.t. | n.t. | n.t. | n.t. | [68] |
| dihomodehydrobatzelladine C (49) | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | 2.3 * | n.t. | n.t. | n.t. | n.t. | [68] |
| Prostate | Ovary | Breast | Melanoma | Lung | Leukemia | Pancreas | Colon | Cervix | Ref. | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DU-145 | IGROV | SK-BR3 | MDA-MB-231 | SK-MEL-28 | NSCL A549 | L-562 | HL-60 | THP-1 | PANCL | HT29 | HCT-16 | LOVO | LOVO-DOX | HeLa | |||
| ptilomycalin A (1) | GI50 | 0.05 | 0.04 | 0.07 | n.t. | 0.03 | 0.08 | 0.04 | n.t. | n.t. | 0.04 | 0.03 | n.t. | 0.05 | 0.05 | 0.04 | [73] |
| TGI | 1.22 | 1.74 | 0.54 | n.t. | 0.11 | 1.23 | 1.33 | n.t. | n.t. | 0.99 | 0.19 | n.t. | 2.14 | 2.04 | 0.22 | ||
| LC50 | 5.21 | n.t. | n.t. | n.t. | 0.98 | 9.79 | 9.72 | n.t. | n.t. | 0.98 | 5.37 | n.t. | 9.79 | 8.48 | 3.21 | ||
| crambescidin 800 (2) | GI50 | 0.19 | 0.05 | 0.16 | n.t. | 0.04 | 0.11 | 0.02 | n.t. | n.t. | 0.04 | 0.04 | n.t. | 0.08 | 0.08 | 0.05 | [73] |
| TGI | 1.38 | 2.50 | 0.56 | n.t. | 0.11 | 1.36 | 0.06 | n.t. | n.t. | 1.53 | 0.23 | n.t. | 2.29 | 2.02 | 0.21 | ||
| LC50 | 7.01 | n.t. | n.t. | n.t. | 1.70 | 9.68 | 6.73 | n.t. | n.t. | 8.66 | 5.75 | n.t. | 8.97 | 8.50 | 1.58 | ||
| crambescidin 816 (3) | GI50 | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | IC50 0.24 | n.t. | n.t. | n.t. | [52] |
| TGI | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | |||
| LC50 | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | |||
| monanchocidin A (18) | GI50 | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | IC50 4.4 * | n.t. | n.t. | n.t. | n.t. | n.t. | IC50 10.1 * | [61] |
| TGI | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | ||||
| LC50 | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | ||||
| monanchocidin B (19) | GI50 | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | IC50 0.17 * | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | [62] |
| TGI | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | |||
| LC50 | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | |||
| monanchocidin C (20) | GI50 | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | IC50 0.09 * | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | [62] |
| TGI | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | |||
| LC50 | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | |||
| monanchocidin D (21) | GI50 | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | IC50 0.69 * | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | [62] |
| TGI | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | |||
| LC50 | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | |||
| monanchocidin E (22) | GI50 | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | IC50 0.55 * | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | [62] |
| TGI | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | |||
| LC50 | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | |||
| monanchomycalin A (23) | GI50 | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | IC50 0.10 * | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | [63] |
| TGI | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | |||
| LC50 | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | |||
| monanchomycalin B (24) | GI50 | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | IC50 0.11 * | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | [63] |
| TGI | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | |||
| LC50 | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | n.t. | |||
| batzelladine C (28) | GI50 | 0.68 | 0.81 | 0.66 | n.t. | 1.45 | 1.40 | 0.62 | n.t. | n.t. | 0.55 | 0.65 | n.t. | 2.06 | 2.25 | 0.70 | [73] |
| TGI | 2.27 | 3.43 | 2.89 | n.t. | 3.67 | 3.42 | 3.01 | n.t. | n.t. | 2.03 | 2.16 | n.t. | 4.37 | 4.72 | 2.22 | ||
| LC50 | 0.69 | n.t. | n.t. | n.t. | 9.24 | 8.31 | n.t. | n.t. | n.t. | 7.14 | 6.70 | n.t. | 9.29 | 0.99 | 6.50 | ||
| dehydrobatzelladine C (35) | GI50 | 0.46 | 0.73 | 0.23 | n.t. | 0.89 | 1.19 | 0.48 | n.t. | n.t. | 0.43 | 0.48 | n.t. | 1.60 | 2.07 | 0.48 | [73] |
| TGI | 1.91 | 4.17 | 1.14 | n.t. | 3.48 | 3.24 | 2.42 | n.t. | n.t. | 1.83 | 1.77 | n.t. | 3.68 | 4.48 | 1.52 | ||
| LC50 | 7.15 | n.t. | n.t. | n.t. | n.t. | 8.81 | n.t. | n.t. | n.t. | 8.66 | 7.50 | n.t. | 8.47 | 9.69 | 5.45 | ||
| batzelladine L (38) | GI50 | 0.44 | 0.52 | 0.23 | n.t. | 0.88 | 1.30 | n.t. | n.t. | n.t. | 0.34 | 4.96 | n.t. | 1.09 | n.t. | 0.38 | [73] |
| TGI | 1.39 | 1.74 | 0.56 | n.t. | 2.18 | 9.99 | n.t. | n.t. | n.t. | 1.33 | n.t. | n.t. | 2.41 | n.t. | 1.16 | ||
| LC50 | 3.78 | 5.01 | 2.10 | n.t. | 4.95 | n.t. | n.t. | n.t. | n.t. | 4.38 | n.t. | n.t. | 5.36 | n.t. | 3.58 | ||
| batzelladine M (39) | GI50 | 1.77 | 2.28 | 1.12 | n.t. | 1.18 | 3.80 | n.t. | n.t. | n.t. | 1.22 | 3.56 | n.t. | 1.99 | n.t. | 1.64 | [73] |
| TGI | 3.44 | 5.08 | 2.51 | n.t. | 4.66 | n.t. | 0.00 | n.t. | n.t. | 3.58 | n.t. | n.t. | 3.56 | n.t. | 3.05 | ||
| LC50 | 6.66 | n.t. | 5.59 | n.t. | n.t. | n.t. | 0.00 | n.t. | n.t. | n.t. | n.t. | n.t. | 6.37 | n.t. | 5.68 | ||
| batzelladine N (40) | GI50 | 1.39 | 1.78 | 1.12 | n.t. | 1.47 | 1.94 | 0.66 | n.t. | n.t. | 1.37 | 1.31 | n.t. | 1.96 | 4.42 | 0.59 | [73] |
| TGI | 3.12 | 4.97 | 3.84 | n.t. | 3.41 | 4.29 | 3.27 | n.t. | n.t. | 3.50 | 3.11 | n.t. | 4.16 | n.t. | 1.80 | ||
| LC50 | 7.04 | n.t. | n.t. | n.t. | 7.97 | 9.47 | n.t. | n.t. | n.t. | 8.97 | 7.35 | n.t. | 8.85 | n.t. | 5.13 | ||
| clathriadic acid (41) | GI50 | n.t. | n.t. | n.t. | 13.5 | n.t. | >30 | n.t. | n.t. | n.t. | n.t. | >30 | n.t. | n.t. | n.t. | n.t. | [68] |
| TGI | n.t. | n.t. | n.t. | >30 | n.t. | >30 | n.t. | n.t. | n.t. | n.t. | >30 | n.t. | n.t. | n.t. | n.t. | ||
| LC50 | n.t. | n.t. | n.t. | >30 | n.t. | >30 | n.t. | n.t. | n.t. | n.t. | >30 | n.t. | n.t. | n.t. | n.t. | ||
| norbatzelladine A (44) | GI50 | n.t. | n.t. | n.t. | 3.8 | n.t. | 2.1 | n.t. | n.t. | n.t. | n.t. | 1.6 | n.t. | TC50 4.7 µM | n.t. | n.t. | [68] |
| TGI | n.t. | n.t. | n.t. | 6.4 | n.t. | 4.6 | n.t. | n.t. | n.t. | n.t. | 3.2 | n.t. | n.t. | n.t. | |||
| LC50 | n.t. | n.t. | n.t. | 11.4 | n.t. | 8.6 | n.t. | n.t. | n.t. | n.t. | 5.7 | n.t. | n.t. | n.t. | |||
| norbatzelladine L (45) | GI50 | n.t. | n.t. | n.t. | 0.7 | n.t. | 1.1 | n.t. | n.t. | n.t. | n.t. | 1.9 | n.t. | n.t. | n.t. | n.t. | [68] |
| TGI | n.t. | n.t. | n.t. | 1.9 | n.t. | 2.1 | n.t. | n.t. | n.t. | n.t. | 4.2 | n.t. | n.t. | n.t. | n.t. | ||
| LC50 | n.t. | n.t. | n.t. | 4.8 | n.t. | 4.2 | n.t. | n.t. | n.t. | n.t. | 7.6 | n.t. | n.t. | n.t. | n.t. | ||
| dinorbatzelladine A (46) | GI50 | n.t. | n.t. | n.t. | 3.0 | n.t. | 1.9 | n.t. | n.t. | n.t. | n.t. | 1.9 | n.t. | n.t. | n.t. | n.t. | [68] |
| TGI | n.t. | n.t. | n.t. | 3.8 | n.t. | 4.2 | n.t. | n.t. | n.t. | n.t. | 4.2 | n.t. | n.t. | n.t. | n.t. | ||
| LC50 | n.t. | n.t. | n.t. | 5.4 | n.t. | 7.6 | n.t. | n.t. | n.t. | n.t. | 7.6 | n.t. | n.t. | n.t. | n.t. | ||
| dinordehydro-batzelladine B (48) | GI50 | n.t. | n.t. | n.t. | n.t. | n.t. | 7.9 | n.t. | n.t. | n.t. | n.t. | 6.2 | n.t. | n.t. | n.t. | n.t. | [68] |
| TGI | n.t. | n.t. | n.t. | n.t. | n.t. | >14 | n.t. | n.t. | n.t. | n.t. | > 14 | n.t. | n.t. | n.t. | n.t. | ||
| LC50 | n.t. | n.t. | n.t. | n.t. | n.t. | >14 | n.t. | n.t. | n.t. | n.t. | > 14 | n.t. | n.t. | n.t. | n.t. | ||
| dihomodehydro-batzelladine C (49) | GI50 | n.t. | n.t. | n.t. | 6.1 | n.t. | >30 | n.t. | n.t. | n.t. | n.t. | >30 | n.t. | n.t. | n.t. | n.t. | [68] |
| TGI | n.t. | n.t. | n.t. | 9.8 | n.t. | >30 | n.t. | n.t. | n.t. | n.t. | >30 | n.t. | n.t. | n.t. | n.t. | ||
| LC50 | n.t. | n.t. | n.t. | 15.6 | n.t. | >30 | n.t. | n.t. | n.t. | n.t. | >30 | n.t. | n.t. | n.t. | n.t. | ||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sfecci, E.; Lacour, T.; Amade, P.; Mehiri, M. Polycyclic Guanidine Alkaloids from Poecilosclerida Marine Sponges. Mar. Drugs 2016, 14, 77. https://doi.org/10.3390/md14040077
Sfecci E, Lacour T, Amade P, Mehiri M. Polycyclic Guanidine Alkaloids from Poecilosclerida Marine Sponges. Marine Drugs. 2016; 14(4):77. https://doi.org/10.3390/md14040077
Chicago/Turabian StyleSfecci, Estelle, Thierry Lacour, Philippe Amade, and Mohamed Mehiri. 2016. "Polycyclic Guanidine Alkaloids from Poecilosclerida Marine Sponges" Marine Drugs 14, no. 4: 77. https://doi.org/10.3390/md14040077