Identification and Characterization of an Isoform Antifreeze Protein from the Antarctic Marine Diatom, Chaetoceros neogracile and Suggestion of the Core Region

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Cloning and Identification of the C. neogracile Isoform AFP
2.2. Phylogenetic Analysis of Cn-isoAFP Using Multiple Alignment
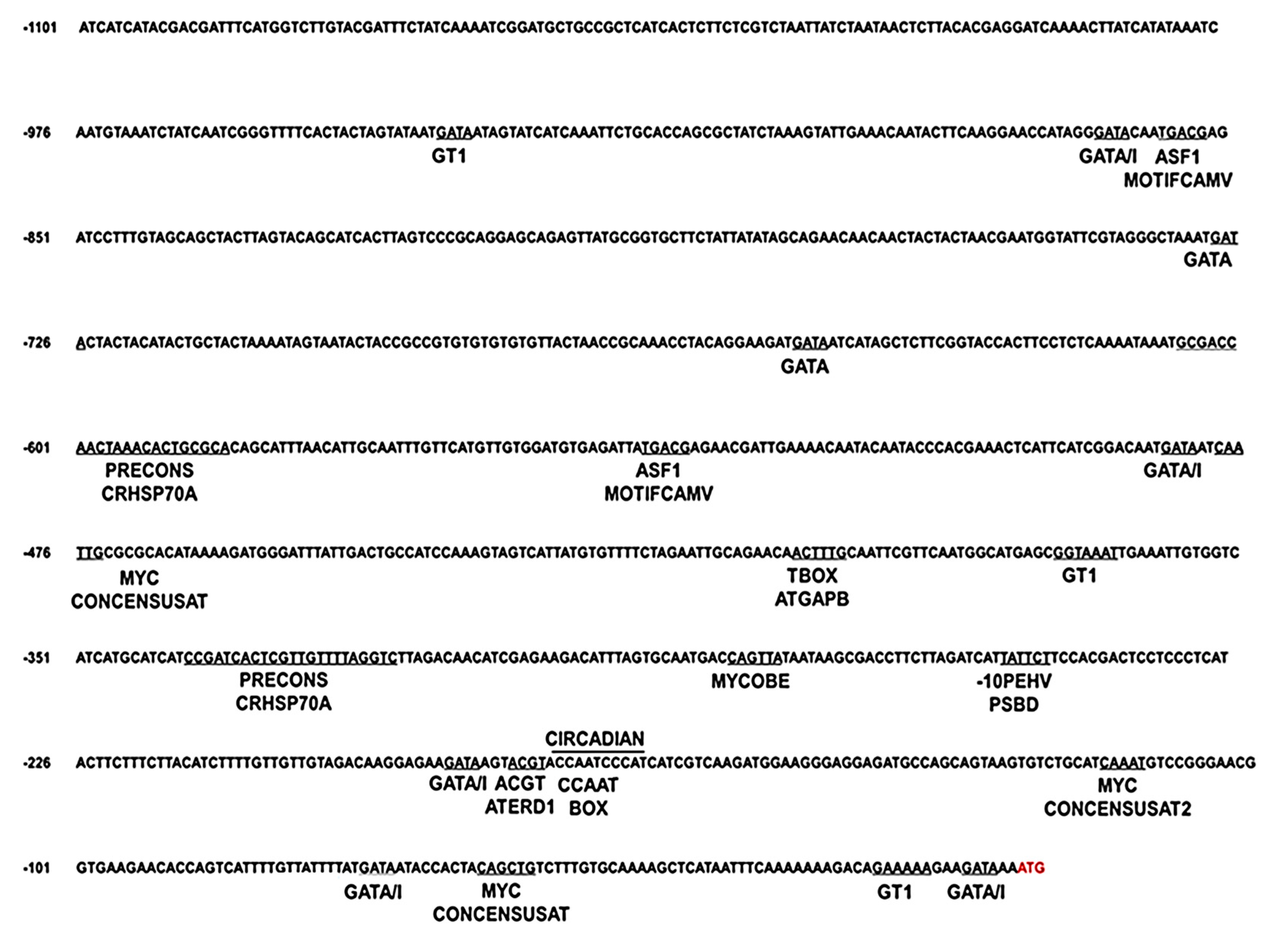
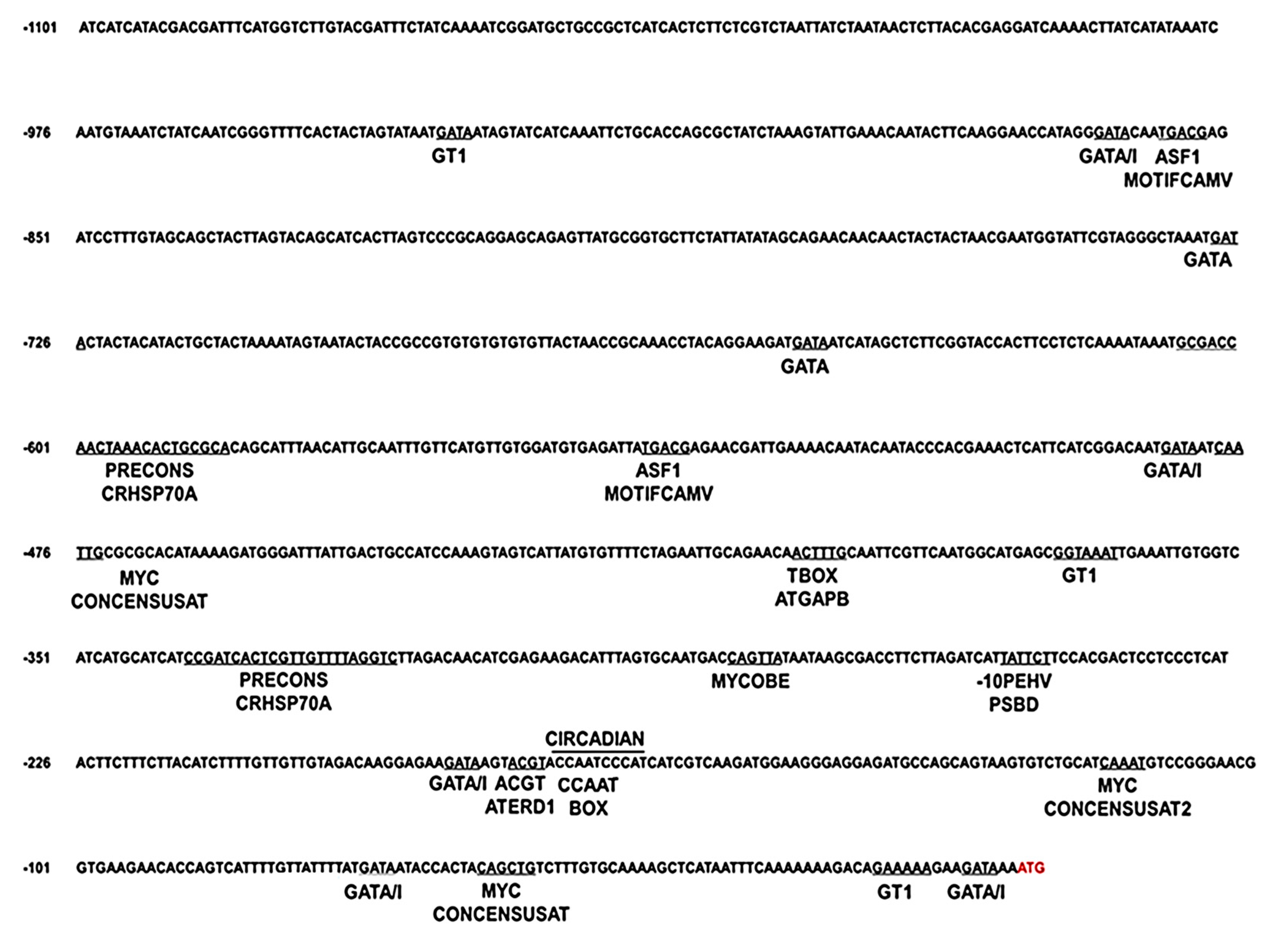
2.3. In Silico Analysis of the Cn-isoAFP Promoter
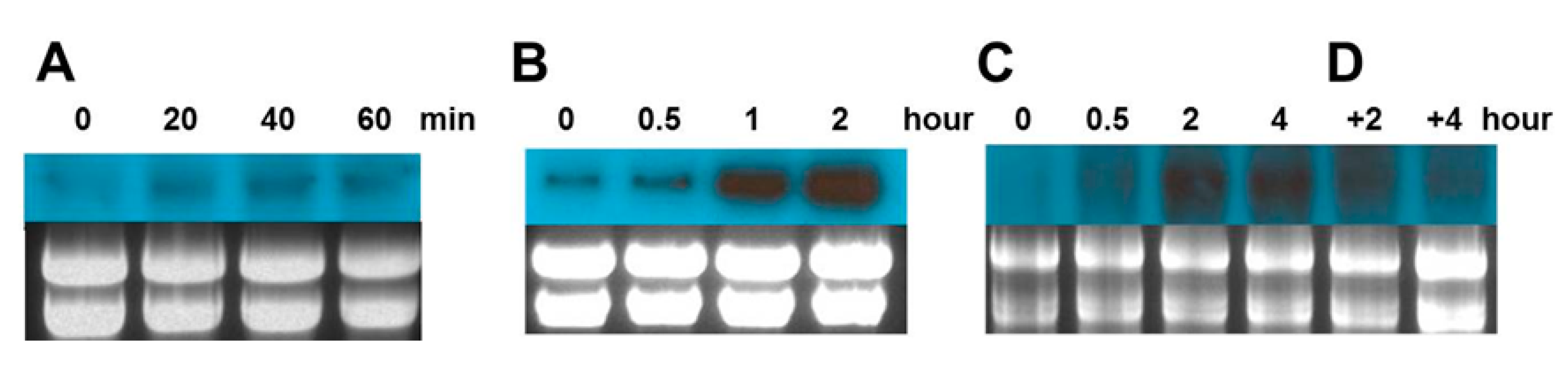
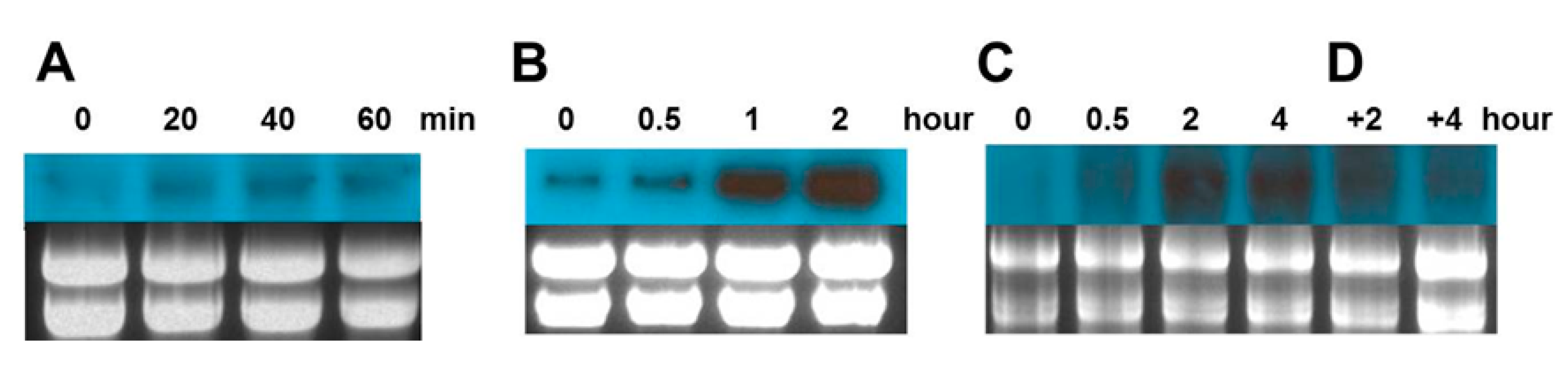
2.4. Gene Expression Analysis of Cn-isoAFP in Response to Stress Conditions
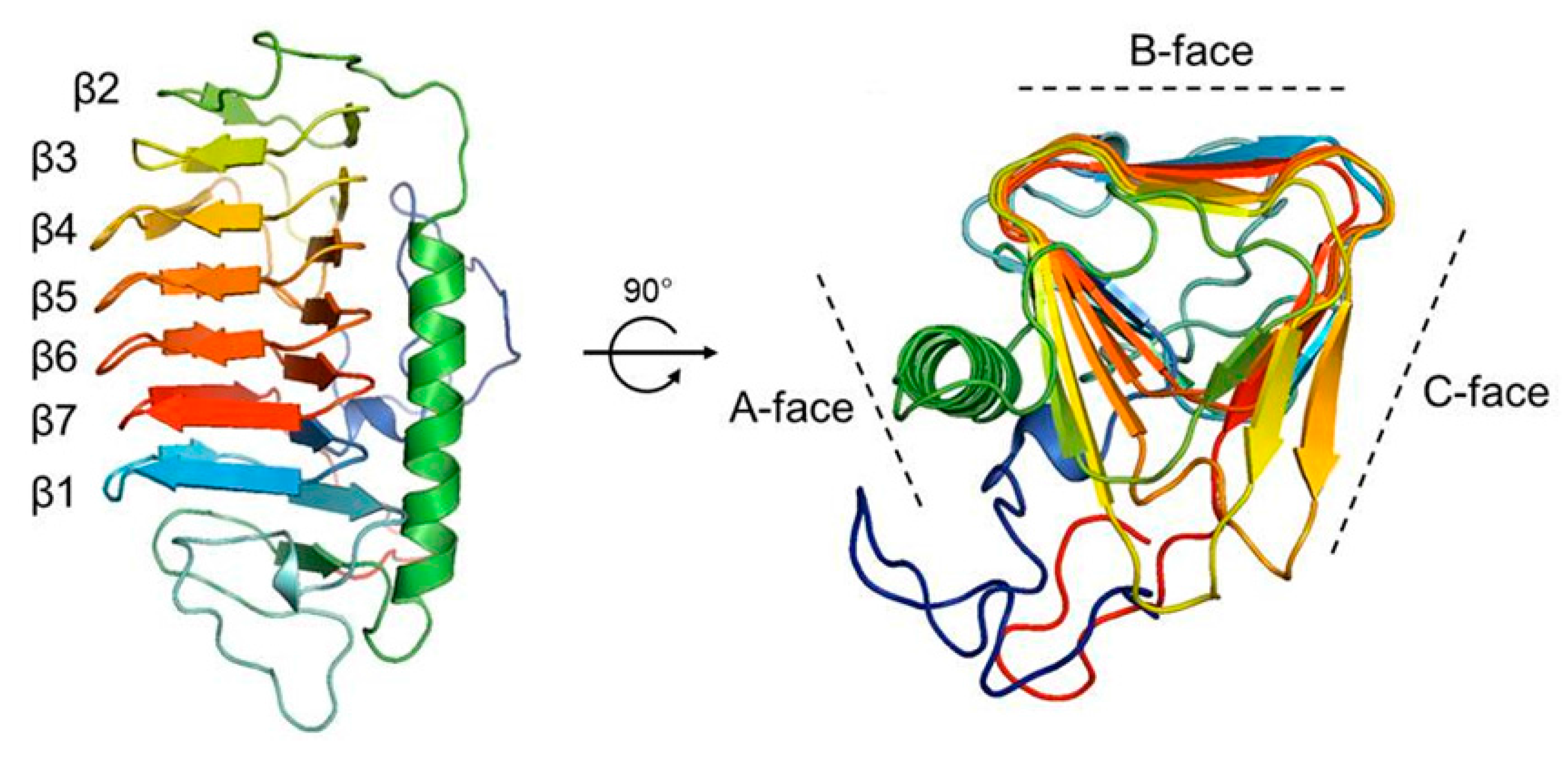
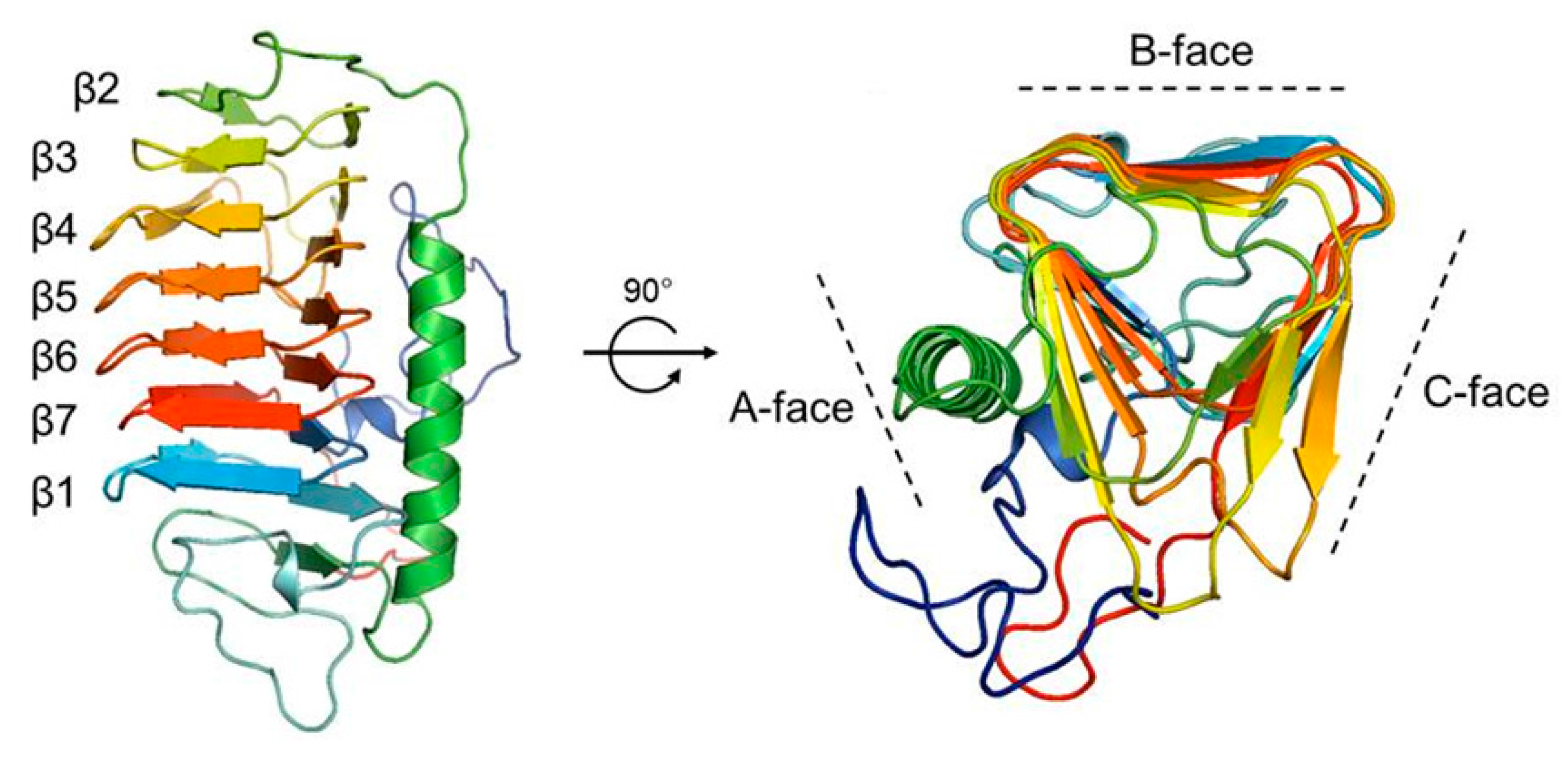
2.5. Homology Modeling of the Structure of Cn-isoAFP
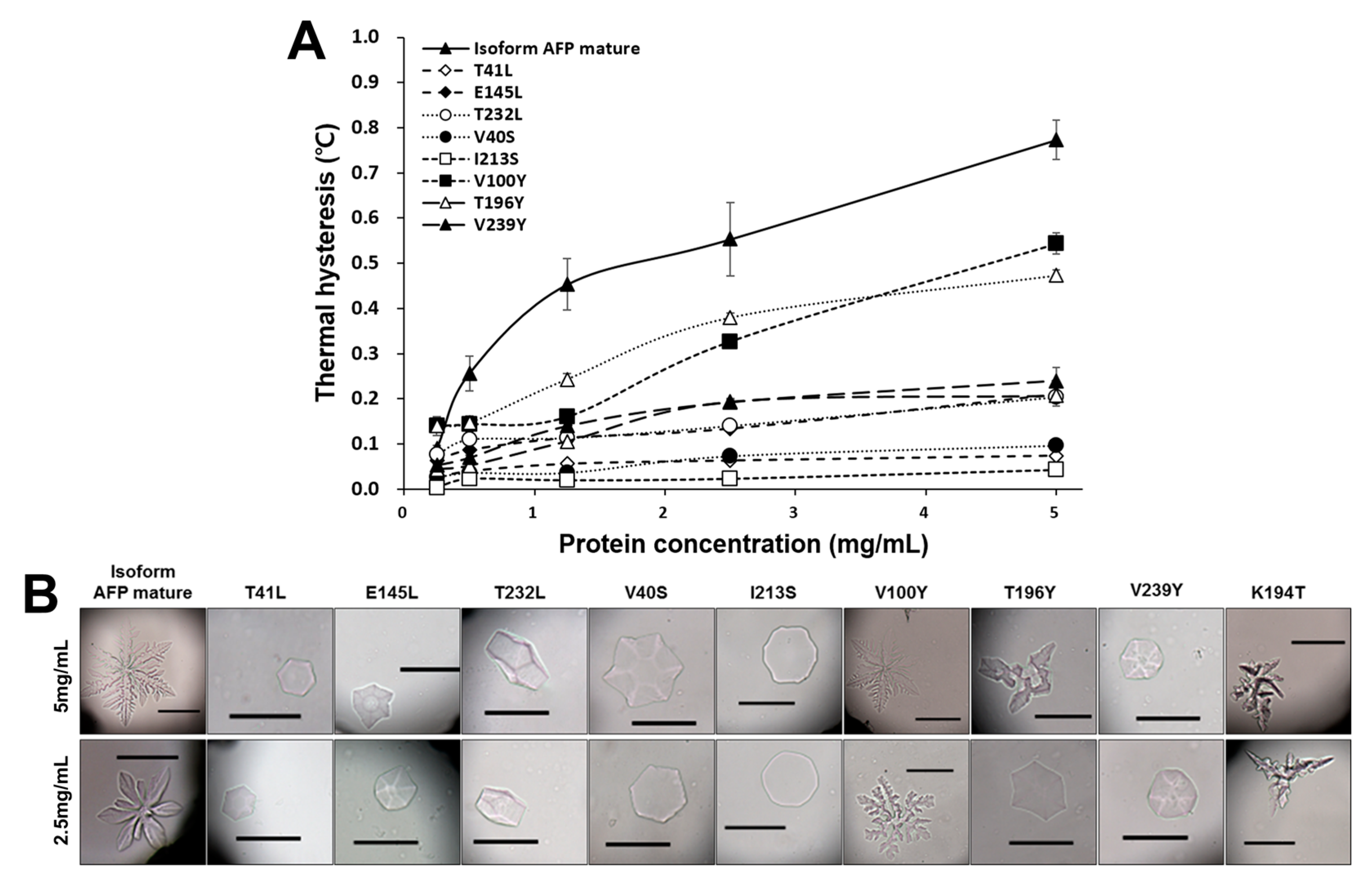
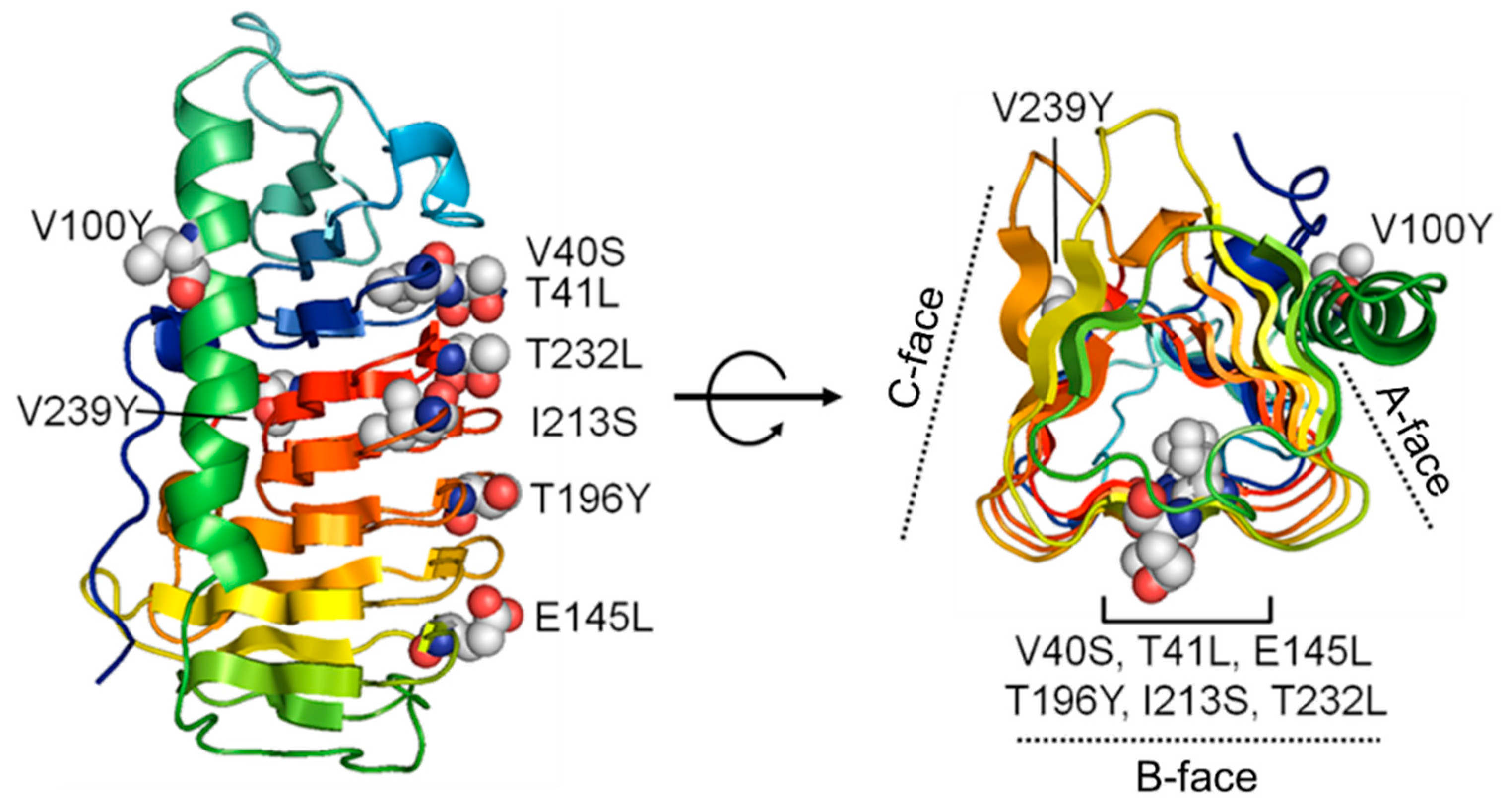
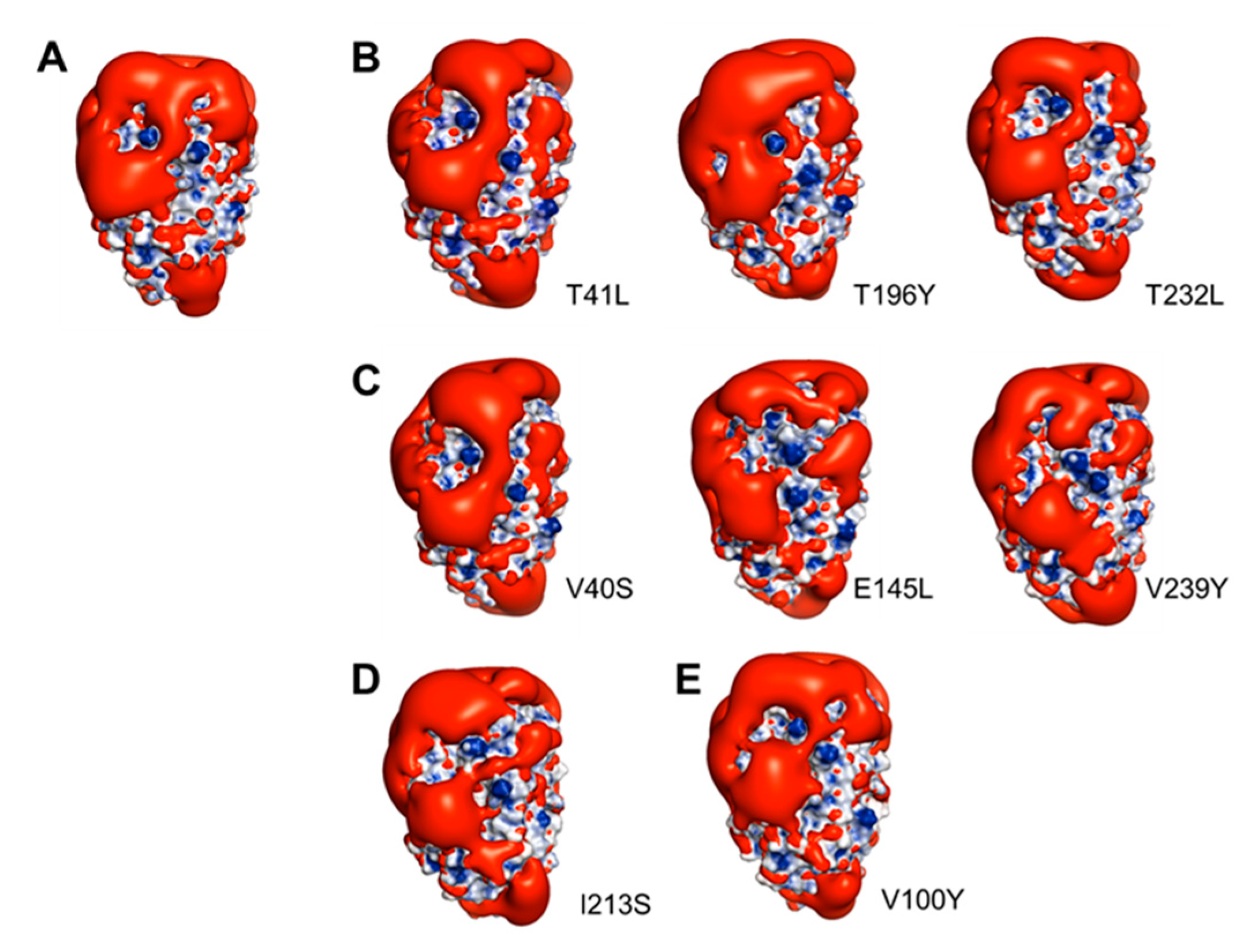
2.6. The Antifreeze Activity of Cn-isoAFP and Its Mutant Proteins
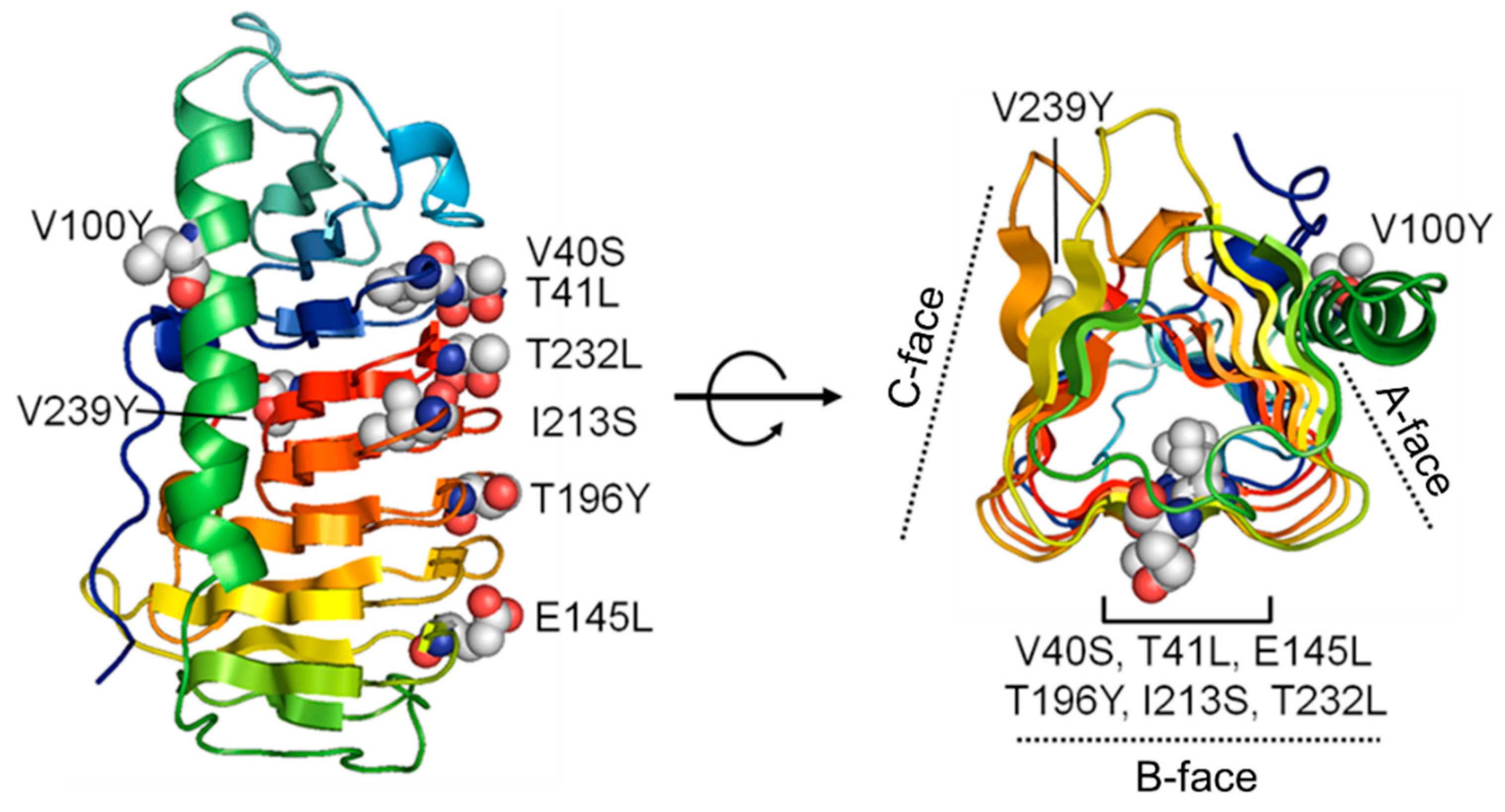
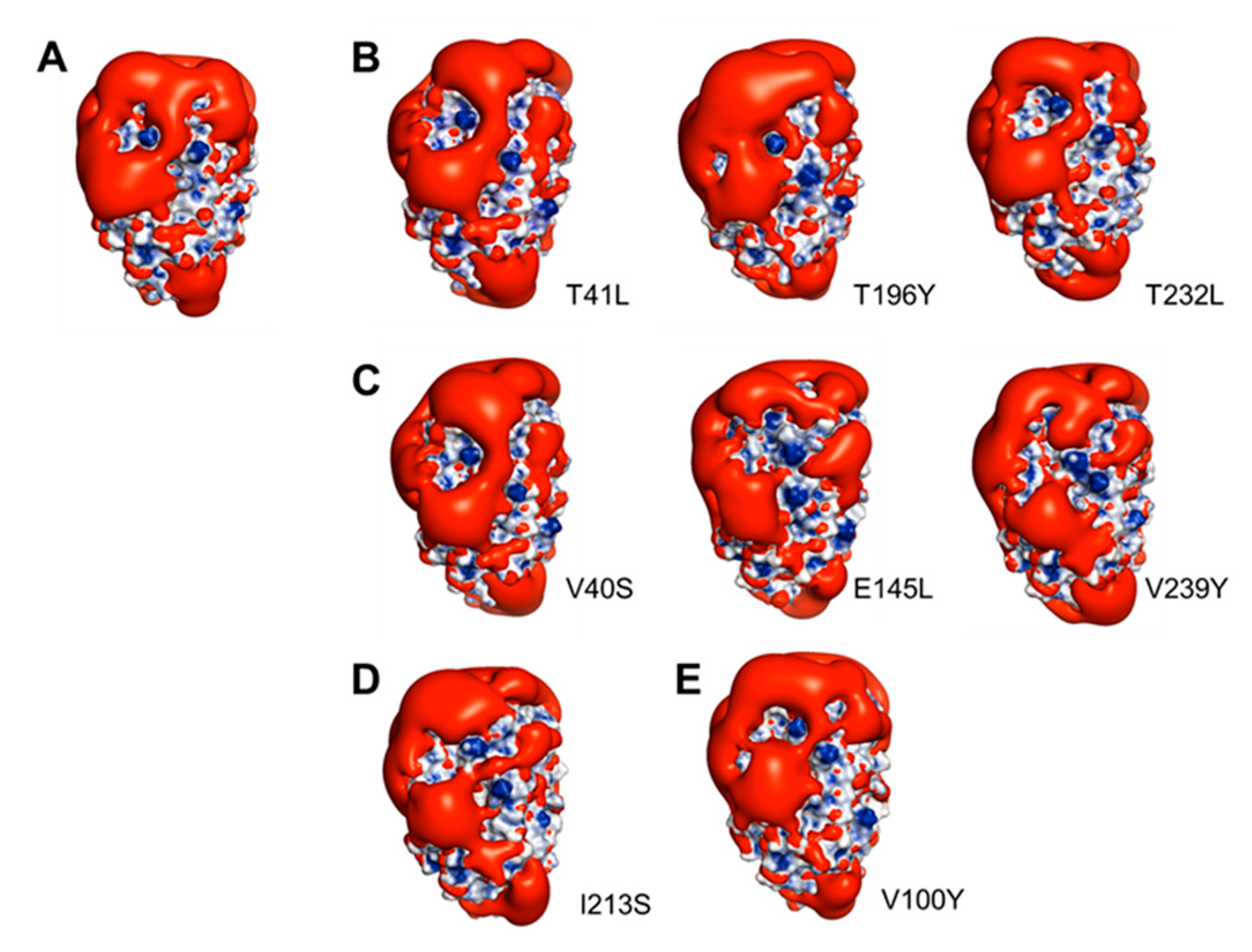
2.7. Modeling of Cn-isoAFP Mutants Generated by Site-Directed Mutagenesis
3. Discussion
4. Materials and Methods
4.1. Cell Growth and Stress Treatments
4.2. Cloning of the Cn-isoAFP Gene and Phylogenetic Analysis
4.3. Identification of the Predicted Promoter Sequence
4.4. Sourthern Blot and Northern Blot Assay
4.5. Cloning for Expression of Recombinant Proteins
4.6. Antifreeze Activity Assay
4.7. Structure Prediction of Cn-isoAFP by Homology Modeling
4.8. Site-Directed Mutagenesis of the Cn-isoAFP
4.9. Evaluation of Structures of Cn-isoAFP and Its Site-Directed Mutants
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yeh, Y.; Feeney, R.E. Antifreeze proteins: Structures and mechanisms of function. Chem. Rev. 1996, 96, 601–618. [Google Scholar] [CrossRef] [PubMed]
- Ewart, K.; Lin, Q.; Hew, C. Structure, function and evolution of antifreeze proteins. Cell. Mol. Life Sci. 1999, 55, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Davies, P.L.; Baardsnes, J.; Kuiper, M.J.; Walker, V.K. Structure and function of antifreeze proteins. Philos. Trans. R. Soc. Lond. B 2002, 357, 927–935. [Google Scholar] [CrossRef] [PubMed]
- Raymond, J.A.; DeVries, A.L. Adsorption inhibition as a mechanism of freezing resistance in polar fishes. Proc. Natl. Acad. Sci. USA 1977, 74, 2589–2593. [Google Scholar] [CrossRef] [PubMed]
- Kondo, H.; Hanada, Y.; Sugimoto, H.; Hoshino, T.; Garnham, C.P.; Davies, P.L.; Tsuda, S. Ice-binding site of snow mold fungus antifreeze protein deviates from structural regularity and high conservation. Proc. Natl. Acad. Sci. USA 2012, 109, 9360–9365. [Google Scholar] [CrossRef] [PubMed]
- Graether, S.P.; Kuiper, M.J.; Gagne, S.M.; Walker, V.K.; Jia, Z.; Sykes, B.D.; Davies, P.L. β-Helix structure and ice-binding properties of a hyperactive antifreeze protein from an insect. Nature 2000, 406, 325–328. [Google Scholar] [PubMed]
- Kuiper, M.J.; Davies, P.L.; Walker, V.K. A theoretical model of a plant antifreeze protein from Lolium perenne. Biophys. J. 2001, 81, 3560–3565. [Google Scholar] [CrossRef]
- Marshall, C.B.; Daley, M.E.; Graham, L.A.; Sykes, B.D.; Davies, P.L. Identification of the ice-binding face of antifreeze protein from Tenebrio molitor. FEBS Lett. 2002, 529, 261–267. [Google Scholar] [CrossRef]
- John, U.P.; Polotnianka, R.M.; Sivakumaran, K.A.; Chew, O.; Mackin, L.; Kuiper, M.J.; Talbot, J.P.; Nugent, G.D.; Mautord, J.; Schrauf, G.E. Ice recrystallization inhibition proteins (IRIPs) and freeze tolerance in the cryophilic Antarctic hair grass Deschampsia antarctica E. Desv. Plant Cell Environ. 2009, 32, 336–348. [Google Scholar] [CrossRef] [PubMed]
- Garnham, C.P.; Campbell, R.L.; Walker, V.K.; Davies, P.L. Novel dimeric β-helical model of an ice nucleation protein with bridged active sites. BMC Struct. Biol. 2011, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Park, A.K.; Do, H.; Park, K.S.; Moh, S.H.; Chi, Y.M.; Kim, H.J. Structural basis for antifreeze activity of ice-binding protein from arctic yeast. J. Biol. Chem. 2012, 287, 11460–11468. [Google Scholar] [CrossRef] [PubMed]
- Janech, M.G.; Krell, A.; Mock, T.; Kang, J.S.; Raymond, J.A. Ice-binding proteins from sea ice diatoms (Bacillariophyceae) 1. J. Phycol. 2006, 42, 410–416. [Google Scholar] [CrossRef] [Green Version]
- Uhlig, C.; Kabisch, J.; Palm, G.J.; Valentin, K.; Schweder, T.; Krell, A. Heterologous expression, refolding and functional characterization of two antifreeze proteins from Fragilariopsis cylindrus (Bacillariophyceae). Cryobiology 2011, 63, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Raymond, J.A.; Janech, M.G.; Fritsen, C.H. Novel ice-binding proteins from a psychrophilic antarctic alga (Chlamydomonadaceae, Chlorophyceae) 1. J. Phycol. 2009, 45, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.; Campbell, R.L.; Gwak, Y.; Im Kim, J.; Davies, P.L.; Jin, E. New Cysteine-Rich Ice-Binding Protein Secreted from Antarctic Microalga, Chloromonas sp. PLoS ONE 2016, 11, e0154056. [Google Scholar] [CrossRef] [PubMed]
- Bayer-Giraldi, M.; Uhlig, C.; John, U.; Mock, T.; Valentin, K. Antifreeze proteins in polar sea ice diatoms: Diversity and gene expression in the genus Fragilariopsis. Environ. Microbiol. 2010, 12, 1041–1052. [Google Scholar] [CrossRef] [PubMed]
- Andorfer, C.A.; Duman, J.G. Isolation and characterization of cDNA clones encoding antifreeze proteins of the pyrochroid beetle Dendroides canadensis. J. Insect Physiol. 2000, 46, 365–372. [Google Scholar] [CrossRef]
- Lauersen, K.J.; Brown, A.; Middleton, A.; Davies, P.L.; Walker, V.K. Expression and characterization of an antifreeze protein from the perennial rye grass, Lolium perenne. Cryobiology 2011, 62, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Raymond, J.A.; Wilson, P.; DeVries, A.L. Inhibition of growth of nonbasal planes in ice by fish antifreezes. Proc. Natl. Acad. Sci. USA 1989, 86, 881–885. [Google Scholar] [CrossRef] [PubMed]
- DeVries, A.L. Antifreeze glycopeptides and peptides: Interactions with ice and water. Method Enzymol. 1986, 127, 293–303. [Google Scholar]
- Knight, C.; Cheng, C.; DeVries, A. Adsorption of alpha-helical antifreeze peptides on specific ice crystal surface planes. Biophys. J. 1991, 59, 409–418. [Google Scholar] [CrossRef]
- Dalal, P.; Sönnichsen, F.D. Source of the ice-binding specificity of antifreeze protein type I. J. Chem. Inf. Comp. Sci. 2000, 40, 1276–1284. [Google Scholar] [CrossRef]
- Garnham, C.P.; Campbell, R.L.; Davies, P.L. Anchored clathrate waters bind antifreeze proteins to ice. Proc. Natl. Acad. Sci. USA 2011, 108, 7363–7367. [Google Scholar] [CrossRef] [PubMed]
- Gwak, Y.; Park, J.-I.; Kim, M.; Kim, H.S.; Kwon, M.J.; Oh, S.J.; Kim, Y.-P.; Jin, E. Creating anti-icing surfaces via the direct immobilization of antifreeze proteins on aluminum. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Gwak, I.G.; Sic Jung, W.; Kim, H.J.; Kang, S.-H.; Jin, E. Antifreeze protein in Antarctic marine diatom, Chaetoceros neogracile. Mar. Biotechnol. 2010, 12, 630–639. [Google Scholar] [CrossRef] [PubMed]
- Gwak, Y.; Jung, W.; Lee, Y.; Kim, J.S.; Kim, C.G.; Ju, J.-H.; Song, C.; Hyun, J.-K.; Jin, E. An intracellular antifreeze protein from an Antarctic microalga that responds to various environmental stresses. FASEB J. 2014, 28, 4924–4935. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, H.; Engelbrecht, J.; Brunak, S.; von Heijne, G. Identification of prokaryotic and eukaryotic signal peptides and prediction of their cleavage sites. Protein Eng. 1997, 10, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.-C.; Lee, T.-Y.; Huang, H.-D.; Huang, H.-Y.; Pan, R.-L. PlantPAN: Plant promoter analysis navigator, for identifying combinatorial cis-regulatory elements with distance constraint in plant gene groups. BMC Genom. 2008, 9, 1. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.-H.; Henderson, D.A.; Zhu, J.-K. The Arabidopsis cold-responsive transcriptome and its regulation by ICE1. Plant Cell 2005, 17, 3155–3175. [Google Scholar] [CrossRef] [PubMed]
- Teakle, G.R.; Manfield, I.W.; Graham, J.F.; Gilmartin, P.M. Arabidopsis thaliana GATA factors: Organisation, expression and DNA-binding characteristics. Plant Mol. Biol. 2002, 50, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Donald, R.; Cashmore, A.R. Mutation of either G box or I box sequences profoundly affects expression from the Arabidopsis rbcS-1A promoter. EMBO J. 1990, 9, 1717–1726. [Google Scholar] [PubMed]
- Thum, K.E.; Kim, M.; Morishige, D.T.; Eibl, C.; Koop, H.-U.; Mullet, J.E. Analysis of barley chloroplast psbD light-responsive promoter elements in transplastomic tobacco. Plant Mol. Biol. 2001, 47, 353–366. [Google Scholar] [CrossRef] [PubMed]
- Terzaghi, W.B.; Cashmore, A.R. Light-regulated transcription. Annu. Rev. Plant Biol. 1995, 46, 445–474. [Google Scholar] [CrossRef]
- Chan, C.-S.; Guo, L.; Shih, M.-C. Promoter analysis of the nuclear gene encoding the chloroplast glyceraldehyde-3-phosphate dehydrogenase B subunit of Arabidopsis thaliana. Plant Mol. Biol. 2001, 46, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Simpson, S.D.; Nakashima, K.; Narusaka, Y.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Two different novel cis-acting elements of erd1, a clpA homologous Arabidopsis gene function in induction by dehydration stress and dark-induced senescence. Plant J. 2003, 33, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Urao, T.; Yamaguchi-Shinozaki, K.; Urao, S.; Shinozaki, K. An Arabidopsis myb homolog is induced by dehydration stress and its gene product binds to the conserved MYB recognition sequence. Plant Cell 1993, 5, 1529–1539. [Google Scholar] [CrossRef] [PubMed]
- von Gromoff, E.D.; Schroda, M.; Oster, U.; Beck, C.F. Identification of a plastid response element that acts as an enhancer within the Chlamydomonas HSP70A promoter. Nucleic Acids Res. 2006, 34, 4767–4779. [Google Scholar] [CrossRef] [PubMed]
- Piechulla, B.; Merforth, N.; Rudolph, B. Identification of tomato Lhc promoter regions necessary for circadian expression. Plant Mol. Biol. 1998, 38, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Lovell, S.C.; Davis, I.W.; Arendall, W.B.; de Bakker, P.I.; Word, J.M.; Prisant, M.G.; Richardson, J.S.; Richardson, D.C. Structure validation by Cα geometry: ϕ, ψ and Cβ deviation. Proteins Struct. Funct. Bioinform. 2003, 50, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Baker, N.A.; Sept, D.; Joseph, S.; Holst, M.J.; McCammon, J.A. Electrostatics of nanosystems: Application to microtubules and the ribosome. Proc. Natl. Acad. Sci. USA 2001, 98, 10037–10041. [Google Scholar] [CrossRef] [PubMed]
- Scott, G.K.; Hew, C.L.; Davies, P.L. Antifreeze protein genes are tandemly linked and clustered in the genome of the winter flounder. Proc. Natl. Acad. Sci. USA 1985, 82, 2613–2617. [Google Scholar] [CrossRef] [PubMed]
- Gourlie, B.; Lin, Y.; Price, J.; DeVries, A.; Powers, D.; Huang, R. Winter flounder antifreeze proteins: A multigene family. J. Biol. Chem. 1984, 259, 14960–14965. [Google Scholar] [PubMed]
- Scott, G.K.; Hayes, P.H.; Fletcher, G.L.; Davies, P.L. Wolffish antifreeze protein genes are primarily organized as tandem repeats that each contain two genes in inverted orientation. Mol. Cell. Biol. 1988, 8, 3670–3675. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.S.; Hon, W.-C.; Bubanko, S.; Xue, Y.; Seetharaman, J.; Hew, C.L.; Sicheri, F. Identification of the ice-binding surface on a type III antifreeze protein with a “flatness function” algorithm. Biophys. J. 1998, 74, 2142–2151. [Google Scholar] [CrossRef]
- Graham, L.A.; Lougheed, S.C.; Ewart, K.V.; Davies, P.L. Lateral transfer of a lectin-like antifreeze protein gene in fishes. PLoS ONE 2008, 3, e2616. [Google Scholar] [CrossRef] [PubMed]
- Xiao, N.; Suzuki, K.; Nishimiya, Y.; Kondo, H.; Miura, A.; Tsuda, S.; Hoshino, T. Comparison of functional properties of two fungal antifreeze proteins from Antarctomyces psychrotrophicus and Typhula ishikariensis. FEBS J. 2010, 277, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Garnham, C.P.; Gilbert, J.A.; Hartman, C.P.; Campbell, R.L.; Laybourn-Parry, J.; Davies, P.L. A Ca2+-dependent bacterial antifreeze protein domain has a novel β-helical ice-binding fold. Biochem. J. 2008, 411, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.; Tyshenko, M.G.; Doucet, D.; Walker, V.K. Characterization of antifreeze protein gene expression in summer spruce budworm larvae. Insect Biochem. Mol. 2006, 36, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Dehury, B.; Patra, M.C.; Maharana, J.; Sahu, J.; Sen, P.; Modi, M.K.; Choudhury, M.D.; Barooah, M. Structure-based computational study of two disease resistance gene homologues (Hm1 and Hm2) in maize (Zea mays L.) with implications in plant-pathogen interactions. PLoS ONE 2014, 9, e97852. [Google Scholar] [CrossRef] [PubMed]
- Pietrzyk, A.J.; Bujacz, A.; Łochynska, M.; Jaskolski, M.; Bujacz, G. Crystal structure of Bombyx mori lipoprotein 6: Comparative structural analysis of the 30-kDa lipoprotein family. PLoS ONE 2014, 9, e108761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burdisso, P.; Milia, F.; Schapire, A.L.; Bologna, N.G.; Palatnik, J.F.; Rasia, R.M. Structural determinants of Arabidopsis thaliana hyponastic leaves 1 function in vivo. PLoS ONE 2014, 9, e113243. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Mamiatis, T.; Fritsch, E.F.; Sambrook, J.; Engel, J. Molecular Cloning—A Laboratory Manual; Cold Spring Harbor Laboratory: New York, NY, USA, 1985. [Google Scholar]
- Jones, D.T. Protein secondary structure prediction based on position-specific scoring matrices. J. Mol. Biol. 1999, 292, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Kelley, L.A.; Sternberg, M.J. Protein structure prediction on the Web: A case study using the Phyre server. Nat. Protoc. 2009, 4, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Šali, A.; Potterton, L.; Yuan, F.; van Vlijmen, H.; Karplus, M. Evaluation of comparative protein modeling by MODELLER. Proteins Struct. Funct. Bioinform. 1995, 23, 318–326. [Google Scholar] [CrossRef] [PubMed]

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.; Gwak, Y.; Jung, W.; Jin, E. Identification and Characterization of an Isoform Antifreeze Protein from the Antarctic Marine Diatom, Chaetoceros neogracile and Suggestion of the Core Region. Mar. Drugs 2017, 15, 318. https://doi.org/10.3390/md15100318
Kim M, Gwak Y, Jung W, Jin E. Identification and Characterization of an Isoform Antifreeze Protein from the Antarctic Marine Diatom, Chaetoceros neogracile and Suggestion of the Core Region. Marine Drugs. 2017; 15(10):318. https://doi.org/10.3390/md15100318
Chicago/Turabian StyleKim, Minjae, Yunho Gwak, Woongsic Jung, and EonSeon Jin. 2017. "Identification and Characterization of an Isoform Antifreeze Protein from the Antarctic Marine Diatom, Chaetoceros neogracile and Suggestion of the Core Region" Marine Drugs 15, no. 10: 318. https://doi.org/10.3390/md15100318