
Immunomodulatory and Anti-IBDV Activities of the Polysaccharide AEX from Coccomyxa gloeobotrydiformis

Abstract
:1. Introduction
2. Results
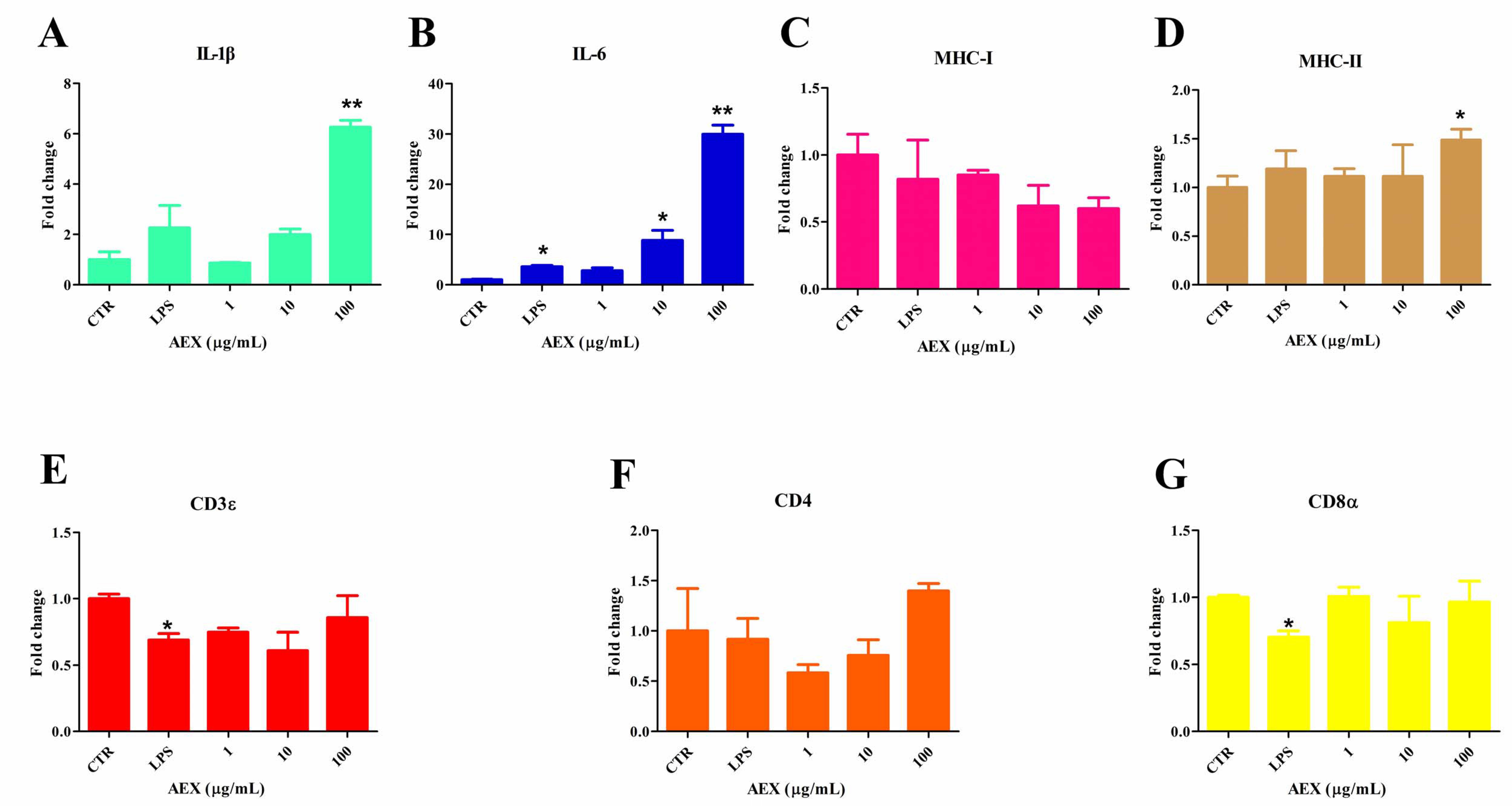
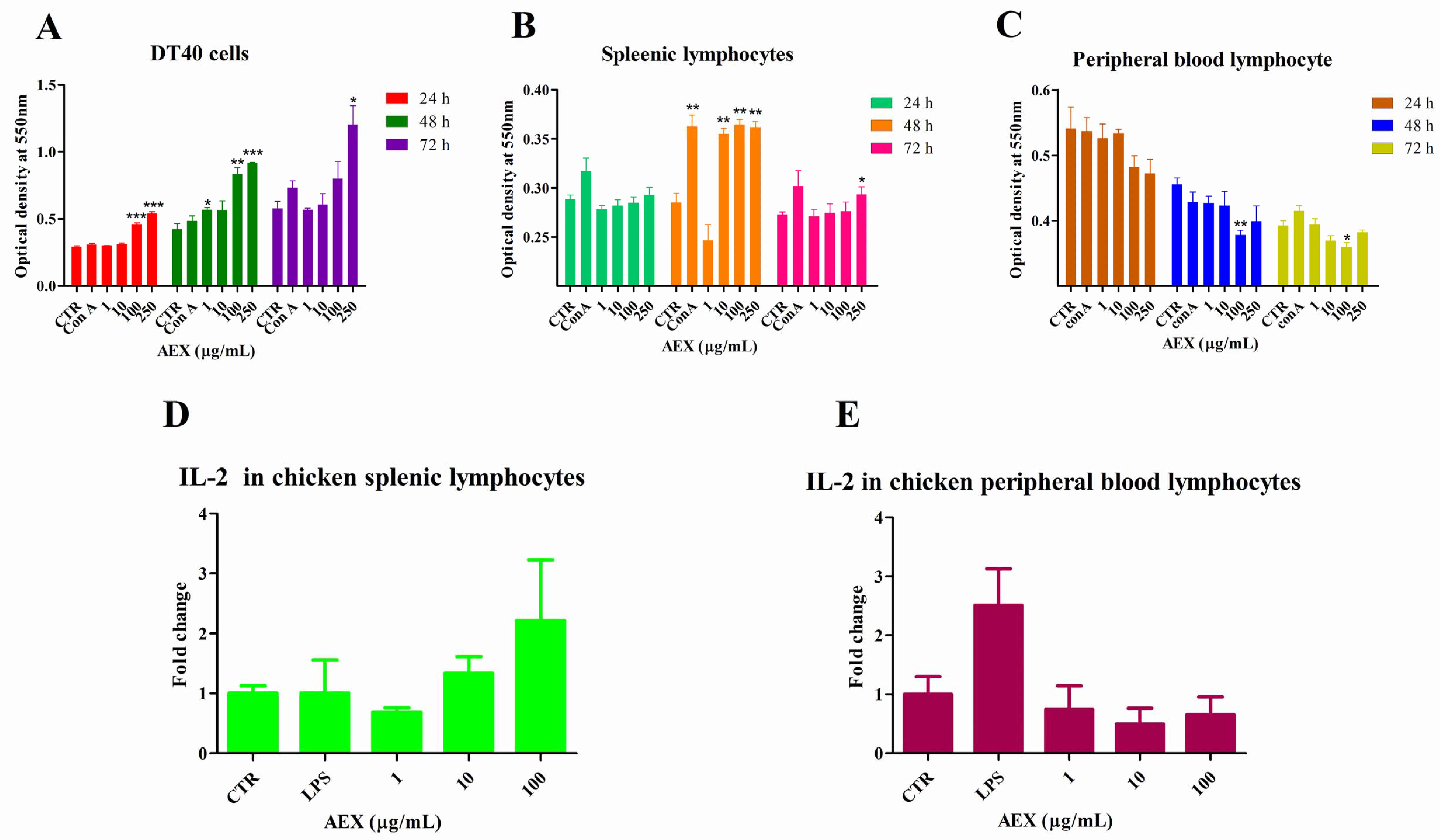
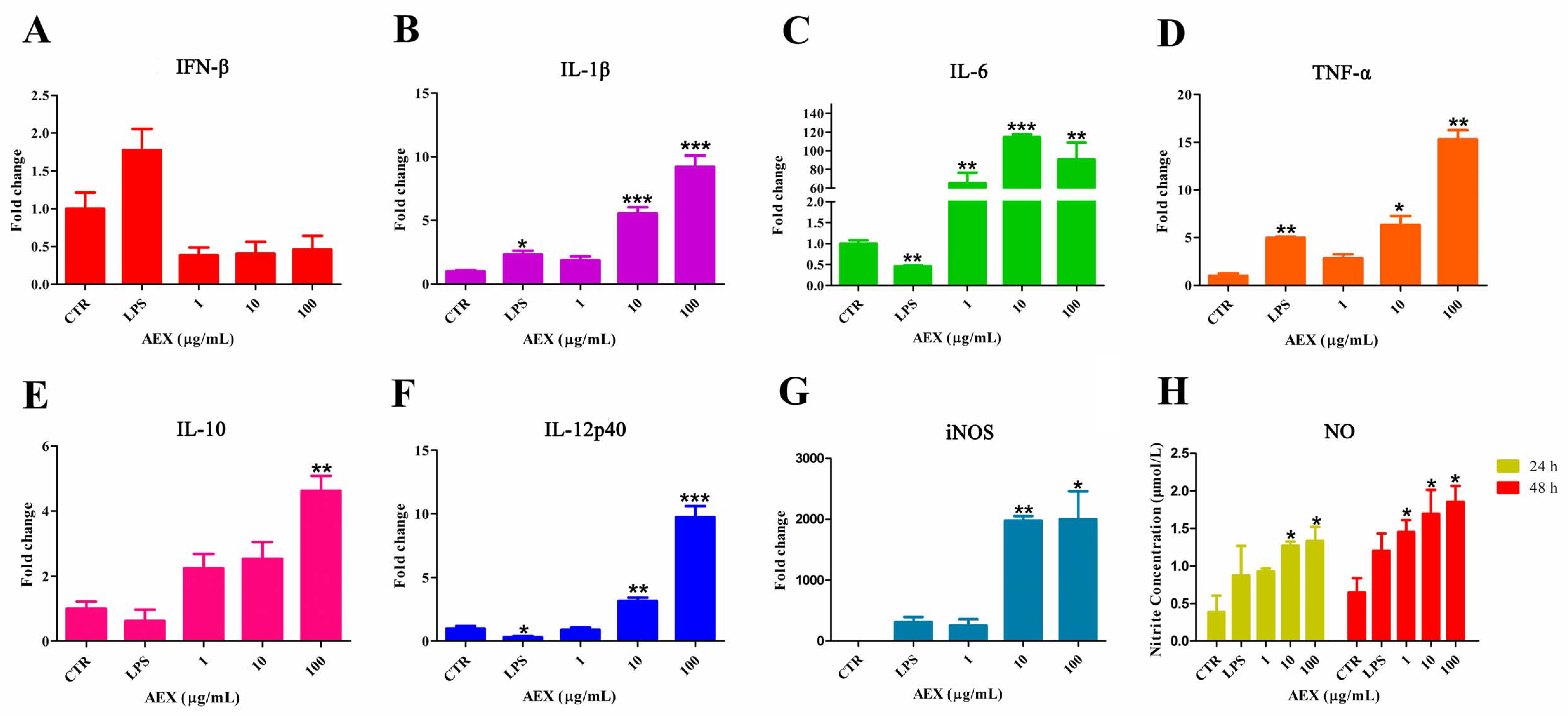
2.1. Effects of AEX on Cytokine and NO Production in Chicken PBMCs
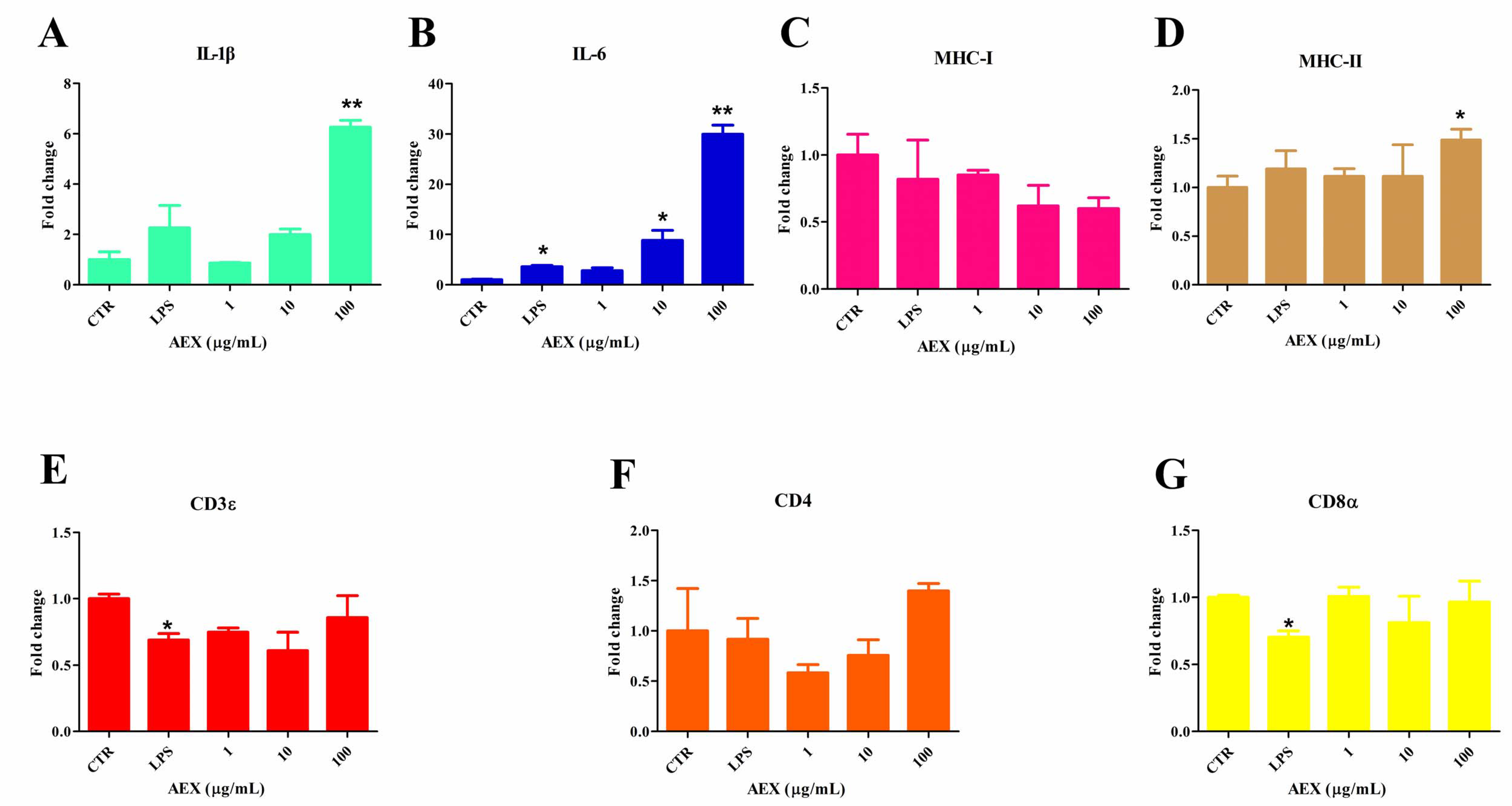
2.2. Effect of AEX on Gene Expression in Splenic Lymphocytes
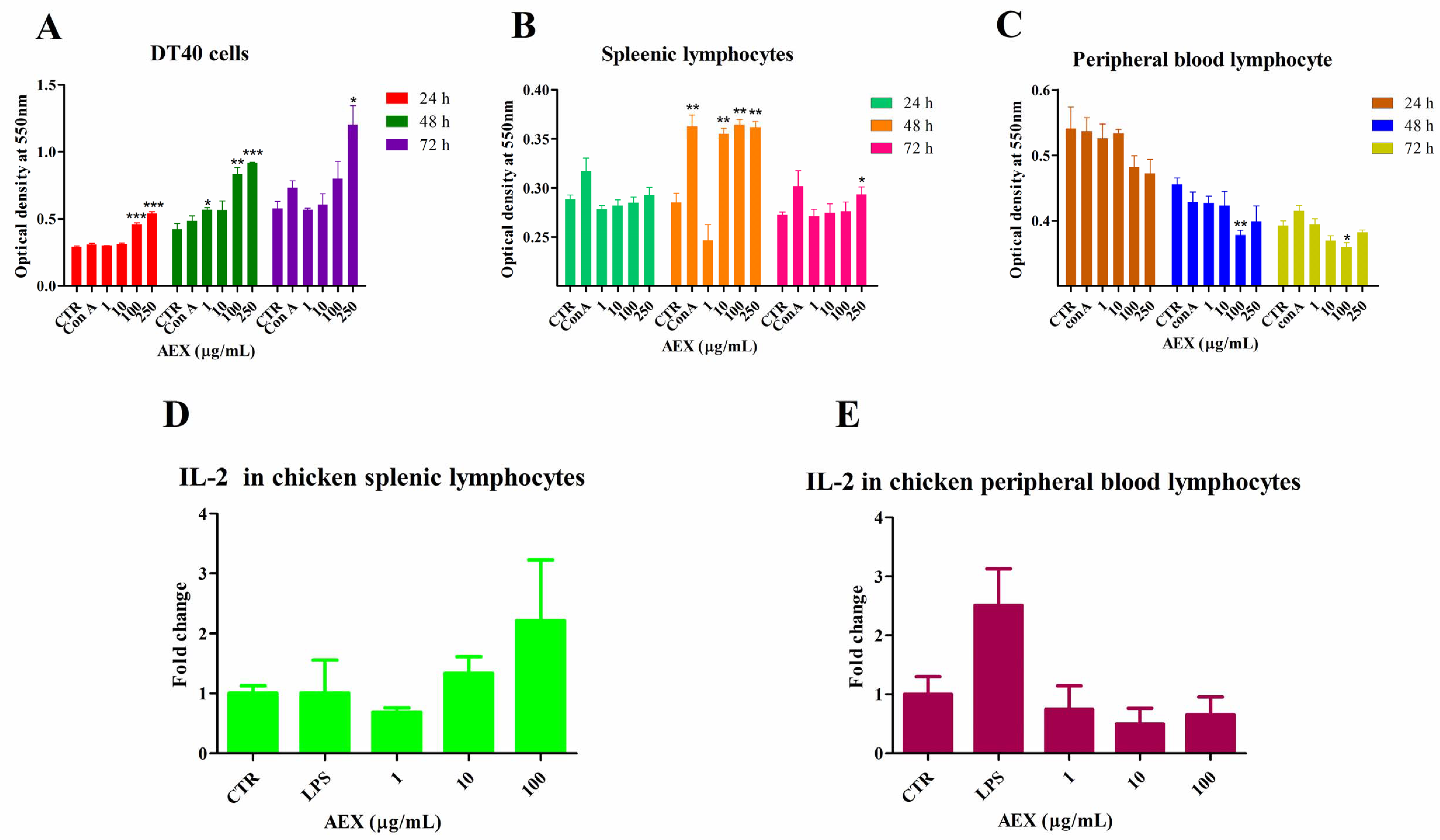
2.3. Effect of AEX on Lymphocyte Proliferation
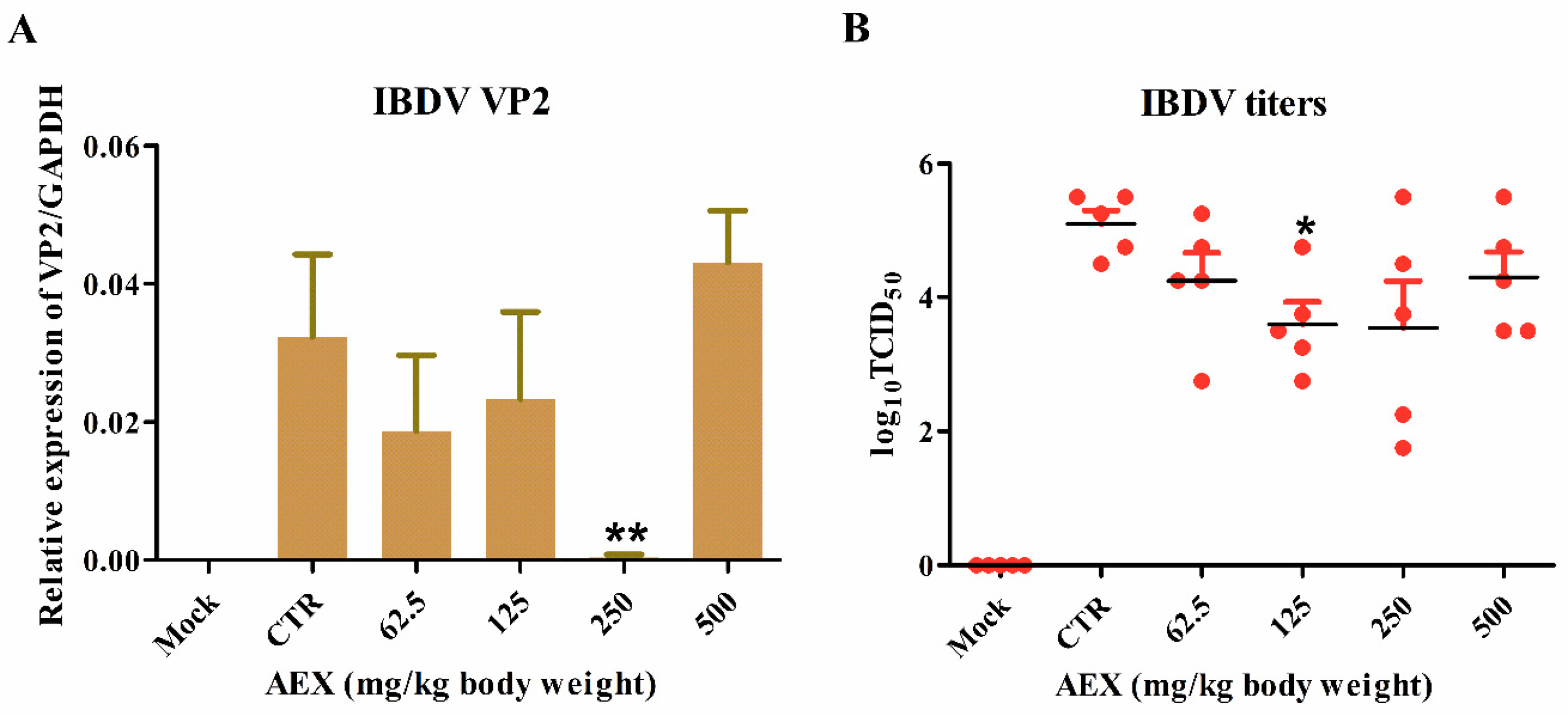
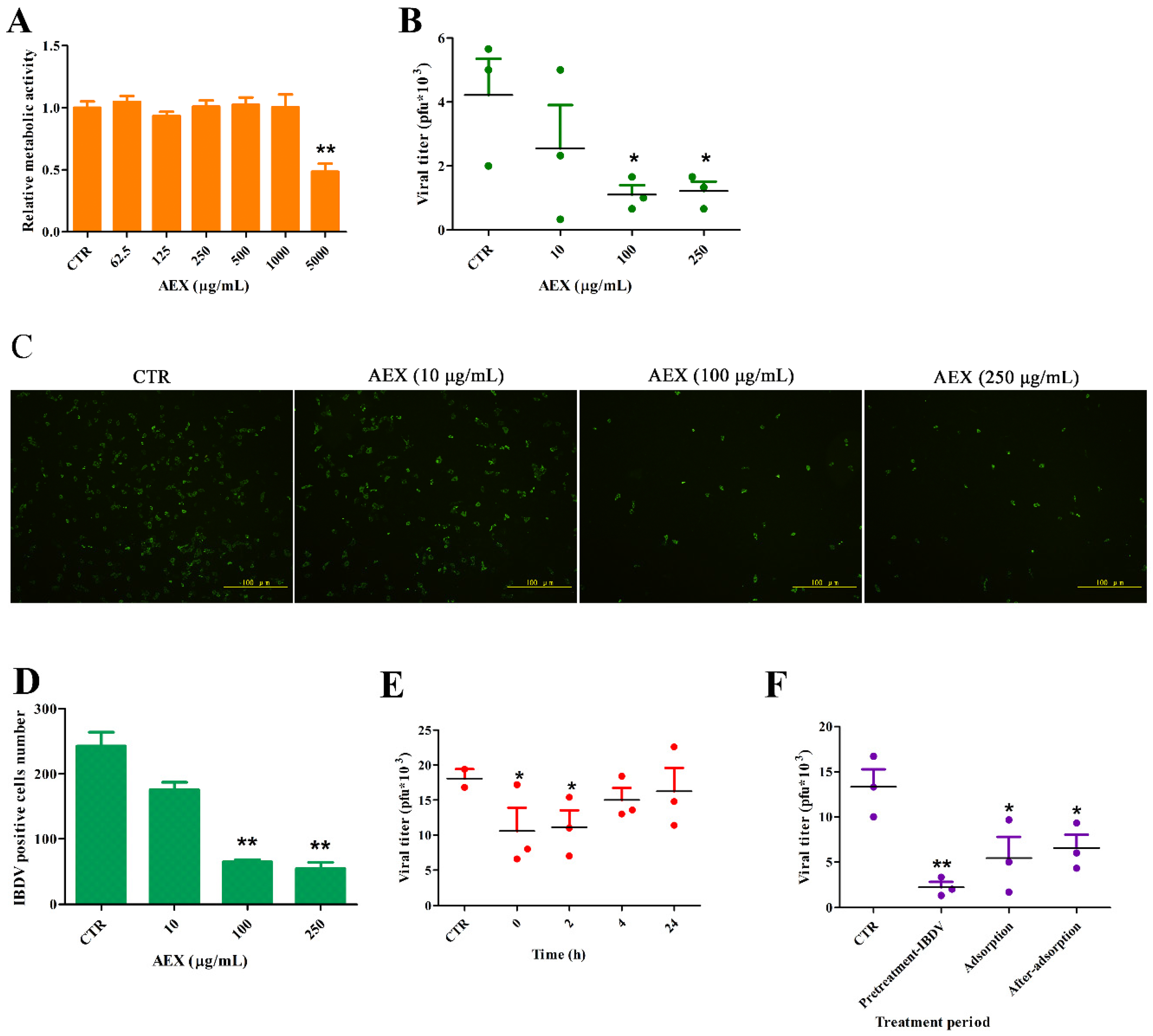
2.4. In Vitro Anti-Viral Effect of AEX on the IBDV TS Strain Virus
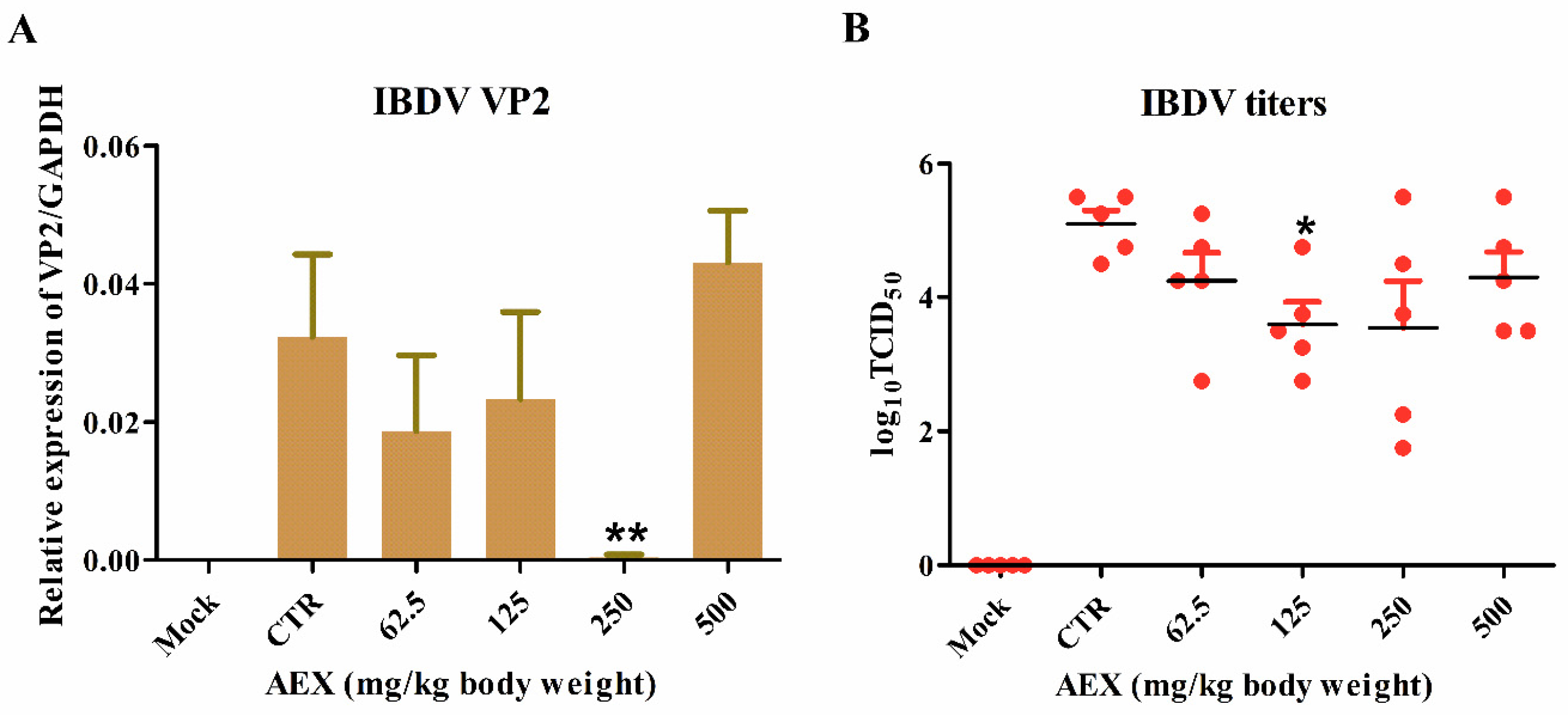
2.5. In Vivo Anti-Viral Effect of AEX on the IBDV TS Strain Virus
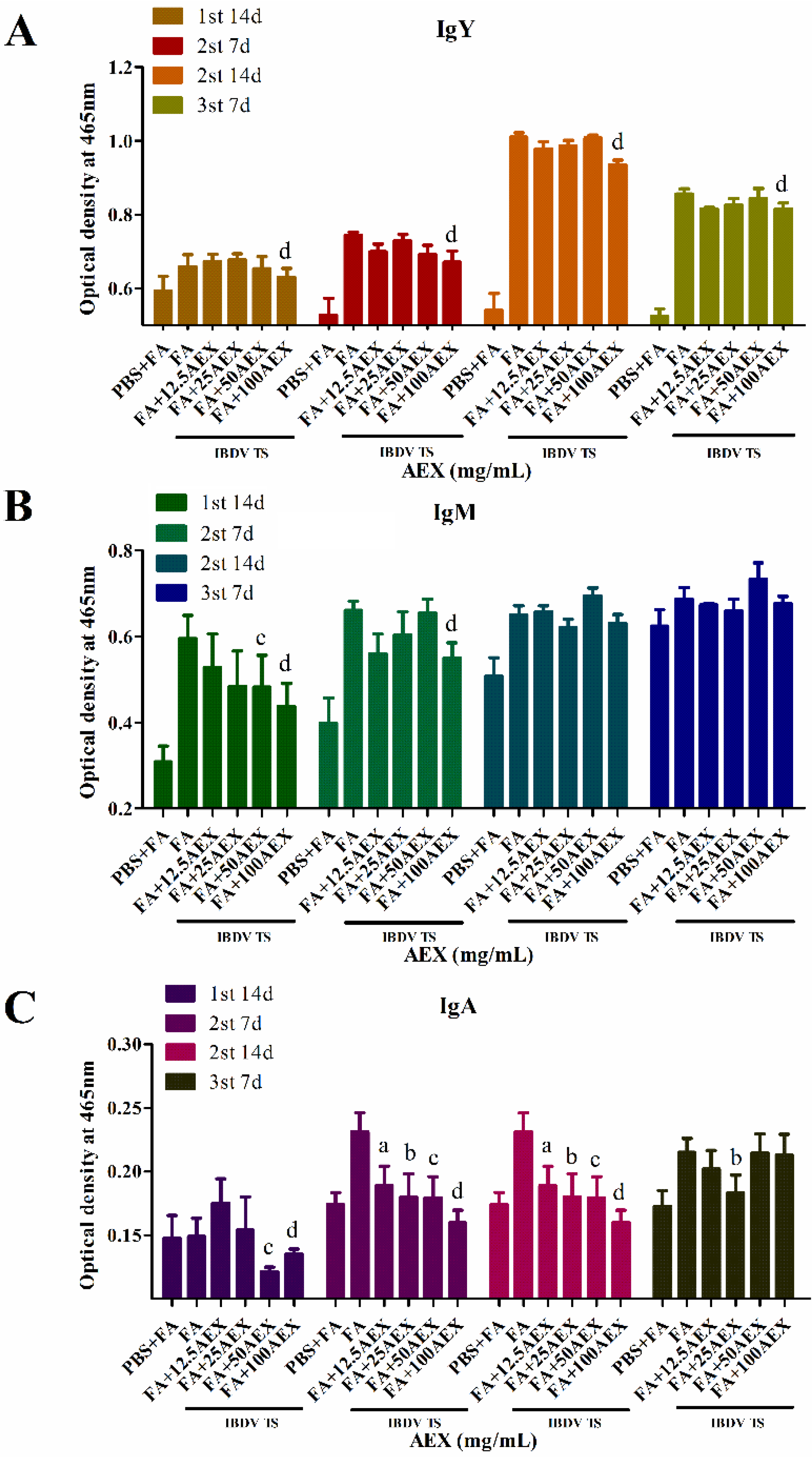
2.6. Effect of AEX on Anti-IBDV Antibodies in Immunized Chickens
3. Discussion
4. Materials and Methods
4.1. Animals, Cells, Compounds and Reagents
4.2. Cell Isolation
4.3. Lymphocyte Proliferation Assay
4.4. Nitric Oxide (NO) Production Assay
4.5. qRT-PCR Analysis
4.6. Viruses and Infection
4.7. Cytotoxicity Assay
4.8. Antiviral Assay (TCID50)
4.9. Immunization of SPF Chicken
4.10. Measurement of Specific Antibody in the Sera of Immunized Chickens
4.11. Statistics
4.12. Ethics Statements
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wang, W.; Wang, S.X.; Guan, H.S. The antiviral activities and mechanisms of marine polysaccharides: An overview. Mar. Drugs 2012, 10, 2795–2816. [Google Scholar] [CrossRef] [PubMed]
- Li, R.-J.; Qiu, S.-D.; Chen, H.-X.; Tian, H.; Wang, H.-X. The immunotherapeutic effects of astragalus polysaccharide in type 1 diabetic mice. Biol. Pharm. Bull. 2007, 30, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.-Y.; Yim, J.H.; Lee, H.K.; Pyo, S. Activation of murine peritoneal macrophages by sulfated exopolysaccharide from marine microalga Gyrodinium impudicum (strain KG03): Involvement of the NF-κB and JNK pathway. Int. Immunopharmacol. 2006, 6, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Yim, J.H.; Son, E.; Pyo, S.; Lee, H.K. Novel sulfated polysaccharide derived from red-tide microalga Gyrodinium impudicum strain KG03 with immunostimulating activity in vivo. Mar. Biotechnol. 2005, 7, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yang, F.; Yang, H.; Wang, G. Water-soluble polysaccharide isolated with alkali from the stem of physalis alkekengi l.: Structural characterization and immunologic enhancement in DNA vaccine. Carbohydr. Polym. 2015, 121, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Wei, K.; Sun, Z.; Yan, Z.; Tan, Y.; Zhu, X.; Wang, X.; Sheng, P.; Zhu, R. Effects of Taishan Pinus massoniana pollen polysaccharide on immune response of rabbit haemorrhagic disease tissue inactivated vaccine and on production performance of Rex rabbits. Vaccine 2011, 29, 2530–2536. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Xue, C.; Wu, C.; Zhao, X.; Qu, T.; He, X.; Guo, Z.; Zhu, R. Immunoregulatory effects of Taishan Pinus massoniana pollen polysaccharide on chicks co-infected with avian leukosis virus and Bordetella avium early in ovo. Res. Vet. Sci. 2014, 96, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Liang, M.; Yang, P.; Guo, F.; Pan, D.; Huang, X.; Li, Y.; Wu, C.; Qu, T.; Zhu, R. Taishan Pinus massoniana pollen polysaccharides promote immune responses of recombinant Bordetella avium ompa in BALB/c mice. Int. Immunopharmacol. 2013, 17, 793–798. [Google Scholar] [CrossRef] [PubMed]
- Hidari, K.I.; Takahashi, N.; Arihara, M.; Nagaoka, M.; Morita, K.; Suzuki, T. Structure and anti-dengue virus activity of sulfated polysaccharide from a marine alga. Biochem. Biophys. Res. Commun. 2008, 376, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Hosoya, M.; Balzarini, J.; Shigeta, S.; De Clercq, E. Differential inhibitory effects of sulfated polysaccharides and polymers on the replication of various myxoviruses and retroviruses, depending on the composition of the target amino acid sequences of the viral envelope glycoproteins. Antimicrob. Agents Chemother. 1991, 35, 2515–2520. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.B.; Hayashi, K.; Hirata, M.; Kuroda, E.; Suzuki, E.; Kubo, Y.; Hayashi, T. Antiviral sulfated polysaccharide from Navicula directa, a diatom collected from deep-sea water in Toyama Bay. Biol. Pharm. Bull. 2006, 29, 2135–2139. [Google Scholar] [CrossRef] [PubMed]
- Talarico, L.B.; Pujol, C.A.; Zibetti, R.G.; Faria, P.C.; Noseda, M.D.; Duarte, M.E.; Damonte, E.B. The antiviral activity of sulfated polysaccharides against dengue virus is dependent on virus serotype and host cell. Antivir. Res. 2005, 66, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, K.; Medeiros, V.; Queiroz, L.; Abreu, L.; Rocha, H.; Ferreira, C.; Juca, M.; Aoyama, H.; Leite, E. Inhibition of reverse transcriptase activity of HIV by polysaccharides of brown algae. Biomed. Pharmacother. 2008, 62, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.; Geng, M.; Guan, H.; Li, Z. Study on the mechanism of inhibitory action of 911 on replication of HIV-1 in vitro. Chin. J. Mar. Drugs 1999, 19, 15–18. [Google Scholar]
- Muto, S.; Niimura, K.; Oohara, M.; Oguchi, Y.; Matsunaga, K.; Hirose, K.; Kakuchi, J.; Sugita, N.; Furusho, T. Polysaccharides from Marine Algae and Antiviral Drugs Containing the Same as Active Ingredients. Eur. Patent EP295956, 21 Decmber 1988. [Google Scholar]
- Carlucci, M.; Scolaro, L.; Noseda, M.; Cerezo, A.; Damonte, E. Protective effect of a natural carrageenan on genital herpes simplex virus infection in mice. Antivir. Res. 2004, 64, 137–141. [Google Scholar] [CrossRef]
- Mundt, E.; Köllner, B.; Kretzschmar, D. VP5 of infectious bursal disease virus is not essential for viral replication in cell culture. J. Virol. 1997, 71, 5647–5651. [Google Scholar] [PubMed]
- Lasher, H.; Shane, S. Infectious bursal disease. Worlds Poult. Sci. J. 1994, 50, 133–166. [Google Scholar] [CrossRef]
- Abdu, P.; Abdullahi, S.; Adesiyun, A.; Ezeokoli, C. Infectious bursal disease. Worlds Poult. Sci. J. 1986, 42, 219–231. [Google Scholar] [CrossRef]
- Berg, T.P.V.D. Acute infectious bursal disease in poultry: A review. Avian Pathol. 2000, 29, 175–194. [Google Scholar] [CrossRef] [PubMed]
- Käufer, I.; Weiss, E. Significance of bursa of fabricius as target organ in infectious bursal disease of chickens. Infect. Immun. 1980, 27, 364–367. [Google Scholar] [PubMed]
- Kibenge, F.; Dhillon, A.; Russell, R. Biochemistry and immunology of infectious bursal disease virus. J. Gen. Virol. 1988, 69, 1757–1775. [Google Scholar] [CrossRef] [PubMed]
- Reisigl, H. Bodenalgen-studien II. Plant Syst. Evol. 1969, 116, 492–506. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. Coccomyxa gloeobotrydiformis Reisigl. Available online: http://www.algaebase.org/search/species/detail/?species_id=H72dc4e4819414c40 (accessed on 1 January 2017).
- Sun, L.; Jin, Y.; Dong, L.; Sumi, R.; Jahan, R.; Li, Z. The neuroprotective effects of Coccomyxa gloeobotrydiformis on the ischemic stroke in a rat model. Int. J. Biol. Sci. 2013, 9, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Jin, Y.; Dong, L.; Sui, H.J.; Sumi, R.; Jahan, R.; Hu, D.; Li, Z. Coccomyxa gloeobotrydiformis improves learning and memory in intrinsic aging rats. Int. J. Biol. Sci. 2015, 11, 825–832. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.M.; Jin, Y.; Liu, Y.L. Inhibitory effect of Cocoomyxa gloeobotrydifomis on benign prostate hyperplasia in aged rats and its action mechanism. Zhonghua Nan Ke Xue 2013, 19, 506–510. [Google Scholar] [PubMed]
- Liu, D.; Sui, H.; Zhang, L.; Jin, Y.; Pharmacology, D.O.; University, L.M. Protective mechanism of Cocoomyxa gloeobotrydifomis in cerebral ischemia-reperfusion injury model rats. Pharmacol. Clin. Chin. Mater. Med. 2014, 6, 036. [Google Scholar]
- Komatsu, T.; Kido, N.; Sugiyama, T.; Yokochi, T. Antiviral activity of acidic polysaccharides from Coccomyxa gloeobotrydiformi, a green alga, against an in vitro human influenza a virus infection. Immunopharmacol. Immunotoxicol. 2013, 35, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Patil, U.S.; Jaydeokar, A.V.; Bandawane, D.D. Immunomodulators: A pharmacological review. Int. J. Pharm. Pharm. Sci. 2012, 4, 30–36. [Google Scholar]
- Jiang, M.H.; Zhu, L.; Jiang, J.G. Immunoregulatory actions of polysaccharides from Chinese herbal medicine. Expert Opin. Ther. Targets 2010, 14, 1367–1402. [Google Scholar] [CrossRef] [PubMed]
- Duerksen-Hughes, P.J. Both tumor necrosis factor and nitric oxide participate in lysis of simian virus 40-transformed cells by activated macrophages. J. Immunol. 1992, 149, 2114–2122. [Google Scholar] [PubMed]
- Farias-Eisner, R.; Chaudhuri, G. Nitric oxide is an important mediator for tumoricidal activity in vivo. Proc. Natl. Acad. Sci. USA 1994, 91, 9407–9411. [Google Scholar] [CrossRef] [PubMed]
- Lorsbach, R.B.; Murphy, W.J.; Lowenstein, C.J.; Snyder, S.H.; Russell, S.W. Expression of the nitric oxide synthase gene in mouse macrophages activated for tumor cell killing. Molecular basis for the synergy between interferon-gamma and lipopolysaccharide. J. Biol. Chem. 1993, 268, 1908–1913. [Google Scholar] [PubMed]
- Baigent, S.J.; Zhang, G.; Fray, M.D.; Flick-Smith, H.; Goodbourn, S.; McCauley, J.W. Inhibition of beta interferon transcription by noncytopathogenic bovine viral diarrhea virus is through an interferon regulatory factor 3-dependent mechanism. J. Virol. 2002, 76, 8979–8988. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.Y.; Gao, J.S.; Yuan, F.; Zhang, W.X.; Shao, Y.J.; Sakurai, F.; Li, Z. Effects of Sophy β-glucan on growth performance, carcass traits, meat composition, and immunological responses of Peking ducks. Poult. Sci. 2011, 90, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Hoffmannfezer, G.; Rodt, H.; Götze, D.; Thierfelder, S. Anatomical distribution of T and B lymphocytes identified by immunohistochemistry in the chicken spleen. Int. Arch. Allergy Appl. Immunol. 1977, 55, 86–95. [Google Scholar] [CrossRef]
- Fair, J.M.; Taylor-McCabe, K.J.; Shou, Y.; Marrone, B.L. Immunophenotyping of chicken peripheral blood lymphocyte subpopulations: Individual variability and repeatability. Vet. Immunol. Immunopathol. 2008, 125, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.L.; Miao, D.Y.; Gong, Y.M.; Zhang, P.J. Effect of three kinds of plant amylose on the immunity with infectious bursal disease live vccine in chickens. Hua Bei Nong Xue Bao 2006, 21, 106–109. [Google Scholar]
- Kuang, L.; Zhan-Qiang, S.U.; Lei, C.H.; Long, L.I.; Shi-Wei, L.I.; Jian, L.I.; Ning-Ning, S.I.; Zhang, Y.H. Study on the immune-promoting effects of Astragalus polysaccharide and epimedium polysaccharide on living vaccine against IBD in chicks. J. Tradit. Chin. Vet. Med. 2013, 2, 009. [Google Scholar]
- Gai-Ru, L.U.; Wei, S.P.; Yao-Di, N.I.; Liu, R.X. Study on active components in Echinacea-Astragalus on relieving immune suppression of IBD chickens. Dongwu Yixue Jinzhan 2010, 5, 002. [Google Scholar]
- Cobb, B.A.; Wang, Q.; Tzianabos, A.O.; Kasper, D.L. Polysaccharide processing and presentation by the mhcii pathway. Cell 2004, 117, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Bohn, J.A.; Bemiller, J.N. (1→3)-β-d-glucans as biological response modifiers: A review of structure-functional activity relationships. Carbohydr. Polym. 1995, 28, 3–14. [Google Scholar] [CrossRef]
- Hashimoto, T.; Ohno, N.; Adachi, Y.; Yadomae, T. Enhanced production of inducible nitric oxide synthase by beta-glucans in mice. FEMS Immunol. Med. Microbiol. 1997, 19, 131–135. [Google Scholar] [CrossRef]
- Chen, J.; Seviour, R. Medicinal importance of fungal beta-(1→3), (1→6)-glucans. Mycol. Res. 2007, 111, 635–652. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Chen, X.; Liu, X.; Zhang, F.; Hu, L.; Yue, Y.; Li, K.; Li, P. Characterization and comparison of the structural features, immune-modulatory and anti-avian influenza virus activities conferred by three algal sulfated polysaccharides. Mar. Drugs 2016, 14, 4. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Withers, D.R.; Young, J.R.; Davison, T.F. Infectious bursal disease virus-induced immunosuppression in the chick is associated with the presence of undifferentiated follicles in the recovering bursa. Viral Immunol. 2005, 18, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, M.; Han, H.; Yuan, J.; Li, Z. Comparison of the expression of cytokine genes in the bursal tissues of the chickens following challenge with infectious bursal disease viruses of varying virulence. Virol. J. 2010, 7, 364. [Google Scholar] [CrossRef] [PubMed]
- Shao, Q.; Xu, W.; Yan, L.; Liu, J.; Rui, L.; Xiao, X.; Yu, X.; Lu, Y.; Li, Z. Function of duck RIG-I in induction of antiviral response against IBDV and avian influenza virus on chicken cells. Virus Res. 2014, 191, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar]
- Hida, S.; Miura, N.N.; Adachi, Y.; Ohno, N. Effect of candida albicans cell wall glucan as adjuvant for induction of autoimmune arthritis in mice. J. Autoimmun. 2005, 25, 93–101. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Direction | Sequence | Accession NO. in GenBank |
|---|---|---|---|
| GAPDH a | Forward | TGCCATCACAGCCACACAGAAG | AF047874.1 |
| Reverse | ACTTTCCCCACAGCCTTAGCAG | ||
| IFN-β | Forward | ACAACTTCCTACAGCACAACAACTA | X92479.1 |
| Reverse | GCCTGGAGGCGGACATG | ||
| IL-1β | Forward | TTGCTGGTTTCCATCTCGTATGTA | NM_204524 |
| Reverse | CCCAGAGCGGCTATTCCA | ||
| IL-2 b | Forward | TTCTGGGACCACTGTATGCTCTT | AF000631.1 |
| Reverse | TACCGACAAAGTGAGAATCAATCAG | ||
| IL-4 b | Forward | AATGACATCCAGGGAGAGGTTTC | AJ621249.1 |
| Reverse | AGGCTTTGCATAAGAGCTCAGTTT | ||
| IL-6 a | Forward | GACGAGGAGAAATGCCTGACG | AJ309540.1 |
| Reverse | CCGAGTCTGGGATGACCACTTC | ||
| IL-10 b | Forward | GCTGAGGGTGAAGTTTGAGGAA | AF000631.1 |
| Reverse | GAAGCGCAGCATCTCTGACA | ||
| IL-12p40 b | Forward | CGAAGTGAAGGAGTTCCCAGAT | AY262752.1 |
| Reverse | GACCGTATCATTTGCCCATTG | ||
| TNF-α | Forward | GGAATGAACCCTCCGCAGTA | AY765397.1 |
| Reverse | CACTACGGGTTGCTGCACAT | ||
| iNOS | Forward | GCCCCTCCAGCTGATCAGA | D85422.1 |
| Reverse | AGGCCTGTGAGAGTGTGCAA | ||
| CD3ε | Forward | TGCCAAAGTGTGTGCAAACTG | NM_206904.1 |
| Reverse | AGTACCCCCAAGGTGATGAGAA | ||
| CD4 | Forward | GCTGTGTGTTTGCGGTCATC | Y12012.1 |
| Reverse | CCTTTCCTGCAATCCCAATC | ||
| CD8α | Forward | CGACAATGGTGTCTCCTGGAT | NM_205235.1 |
| Reverse | GGGAAAGTGGTCCGGGATAA | ||
| MHC-I | Forward | TGGTTGGTGTTGGATTCATCAT | KF294514.1 |
| Reverse | GCTGGATCCACCTTCCTTGTC | ||
| MHC-II | Forward | GTGGGCTCAGTTCGGTTTTC | DQ207939.1 |
| Reverse | AATTCGGGCAGCCTCCATA | ||
| TS vp2 b | Forward | ACCGGCACCGACAACCTTA | AF076230.1 |
| Reverse | CCCTGCCTGACCACCACTT |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Q.; Shao, Q.; Xu, W.; Rui, L.; Sumi, R.; Eguchi, F.; Li, Z. Immunomodulatory and Anti-IBDV Activities of the Polysaccharide AEX from Coccomyxa gloeobotrydiformis. Mar. Drugs 2017, 15, 36. https://doi.org/10.3390/md15020036
Guo Q, Shao Q, Xu W, Rui L, Sumi R, Eguchi F, Li Z. Immunomodulatory and Anti-IBDV Activities of the Polysaccharide AEX from Coccomyxa gloeobotrydiformis. Marine Drugs. 2017; 15(2):36. https://doi.org/10.3390/md15020036
Chicago/Turabian StyleGuo, Qiang, Qiang Shao, Wenping Xu, Lei Rui, Ryo Sumi, Fumio Eguchi, and Zandong Li. 2017. "Immunomodulatory and Anti-IBDV Activities of the Polysaccharide AEX from Coccomyxa gloeobotrydiformis" Marine Drugs 15, no. 2: 36. https://doi.org/10.3390/md15020036