
Marine Lectins DlFBL and HddSBL Fused with Soluble Coxsackie-Adenovirus Receptor Facilitate Adenovirus Infection in Cancer Cells BUT Have Different Effects on Cell Survival

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
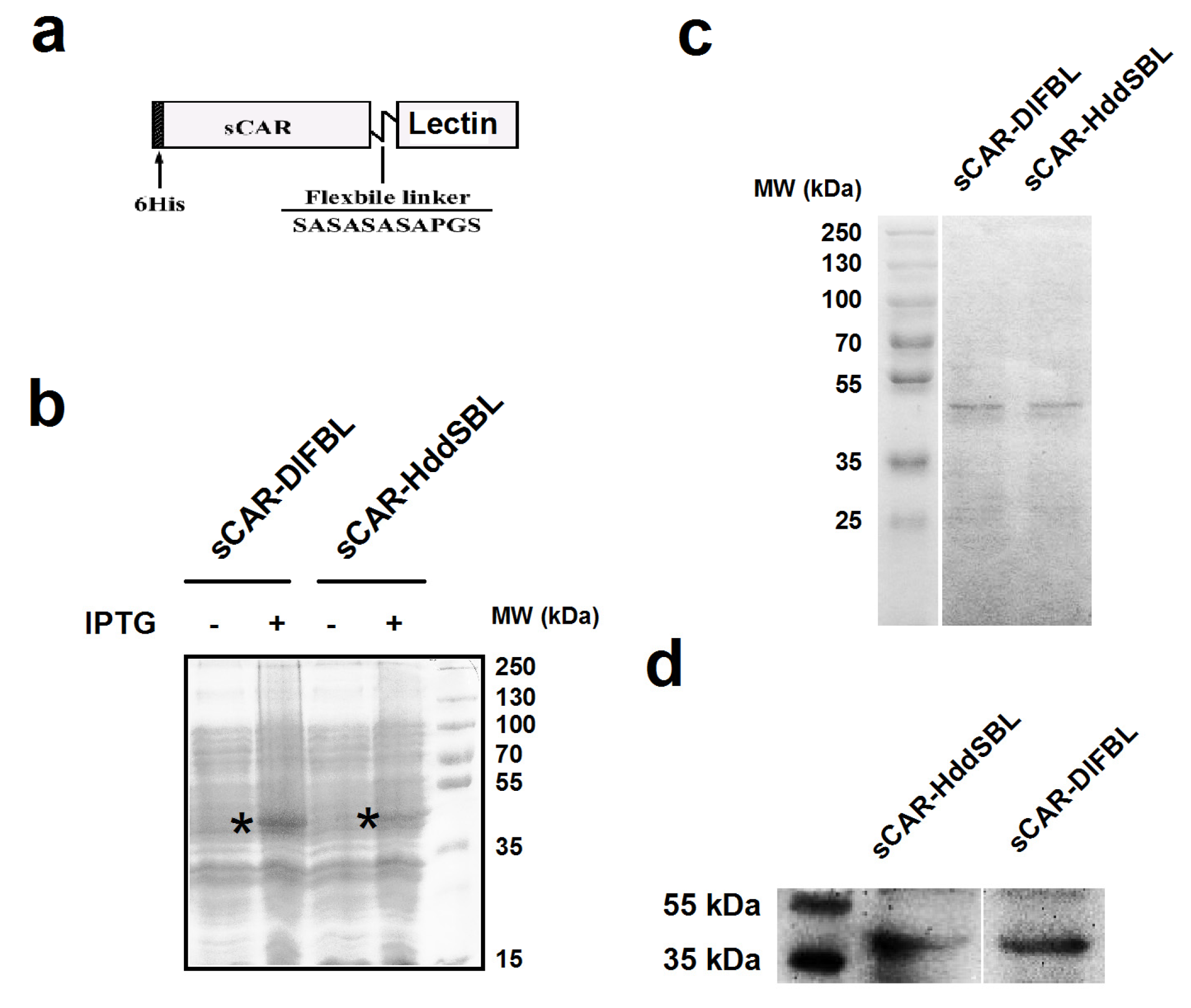
2.1. The Production of sCAR-DlFBL and sCAR-HddSBL Fusion Proteins
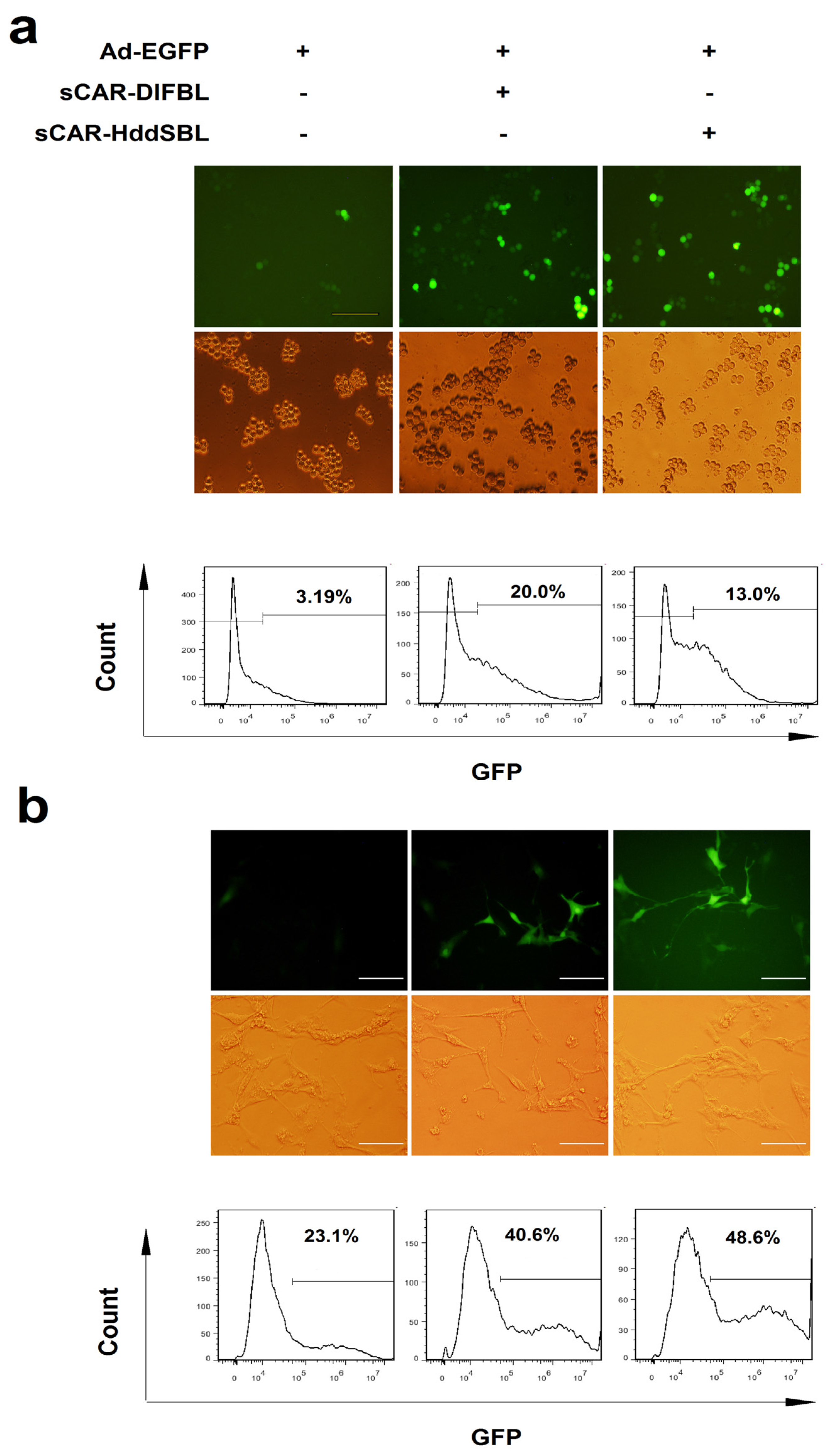
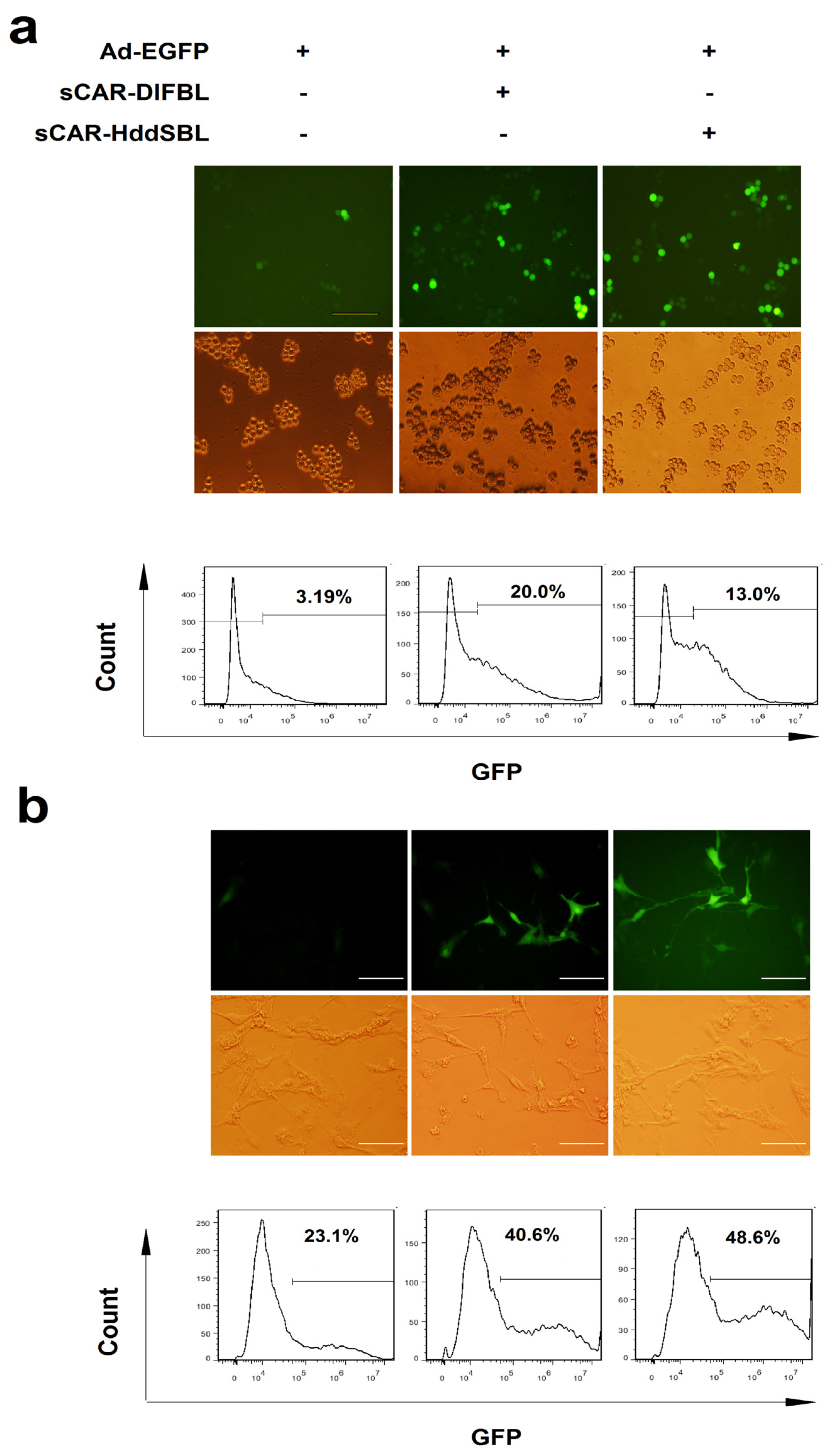
2.2. Recombinant sCAR-DlFBL and sCAR-HddSBL Proteins Facilitated Adenovirus Infection
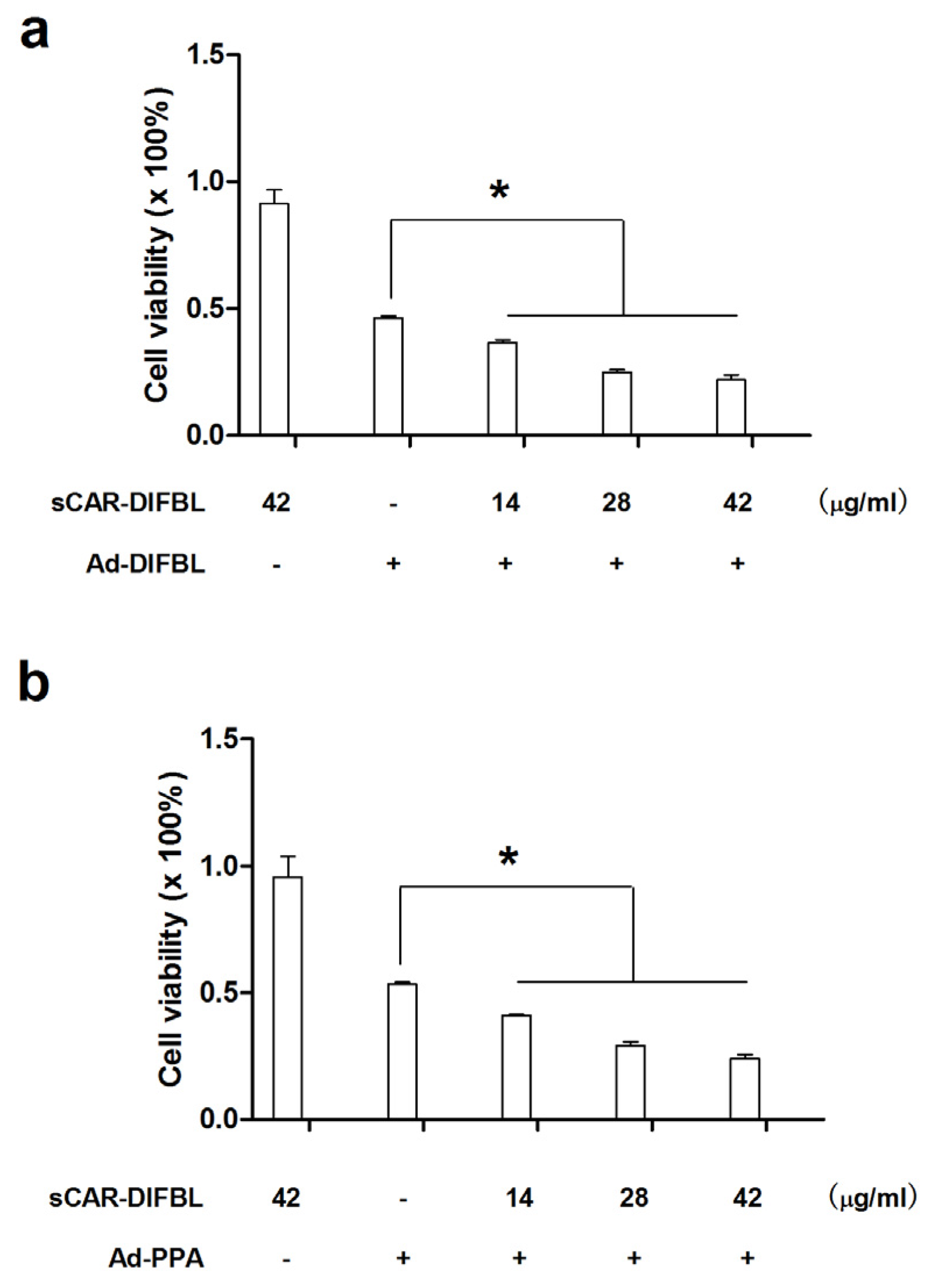
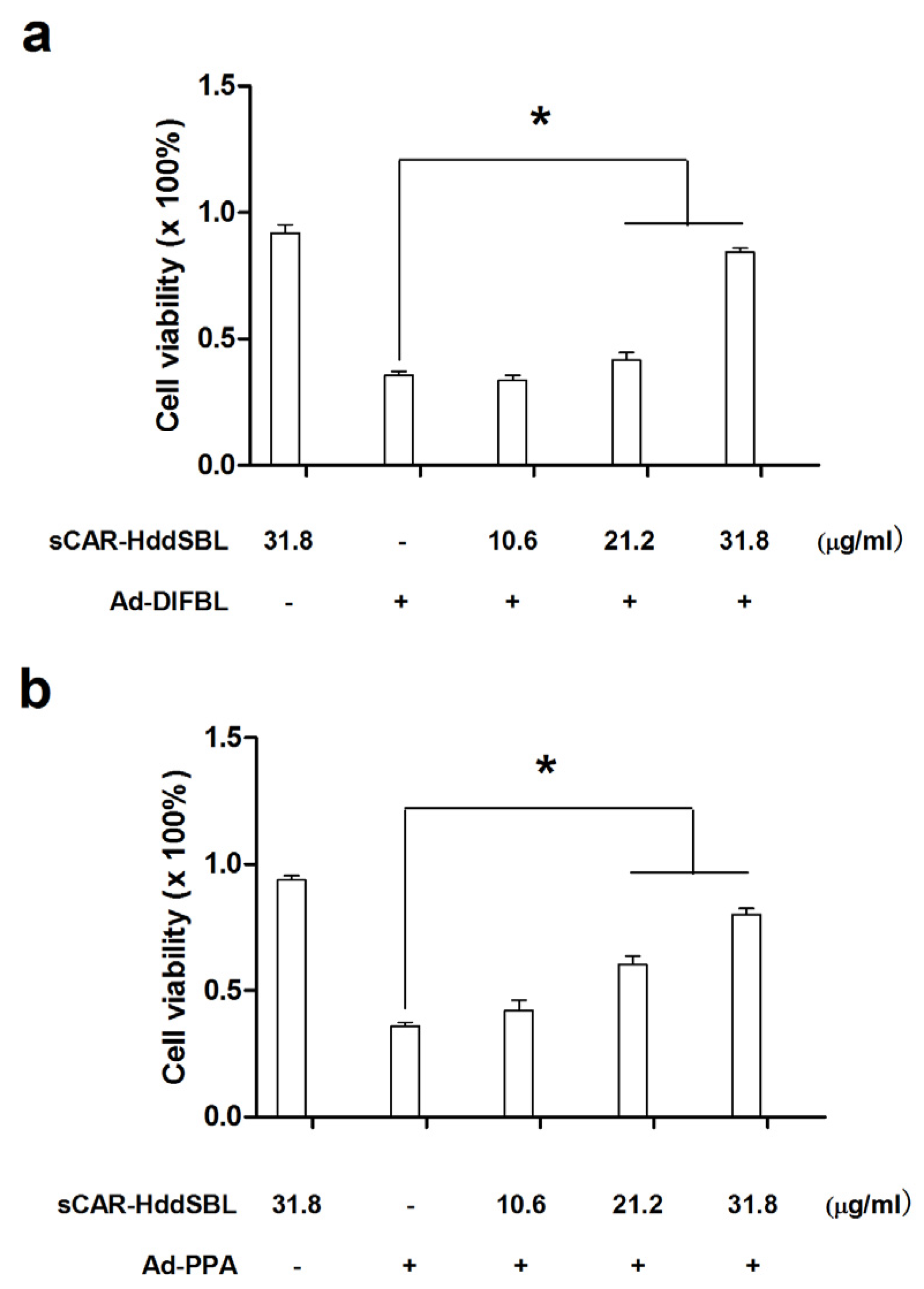
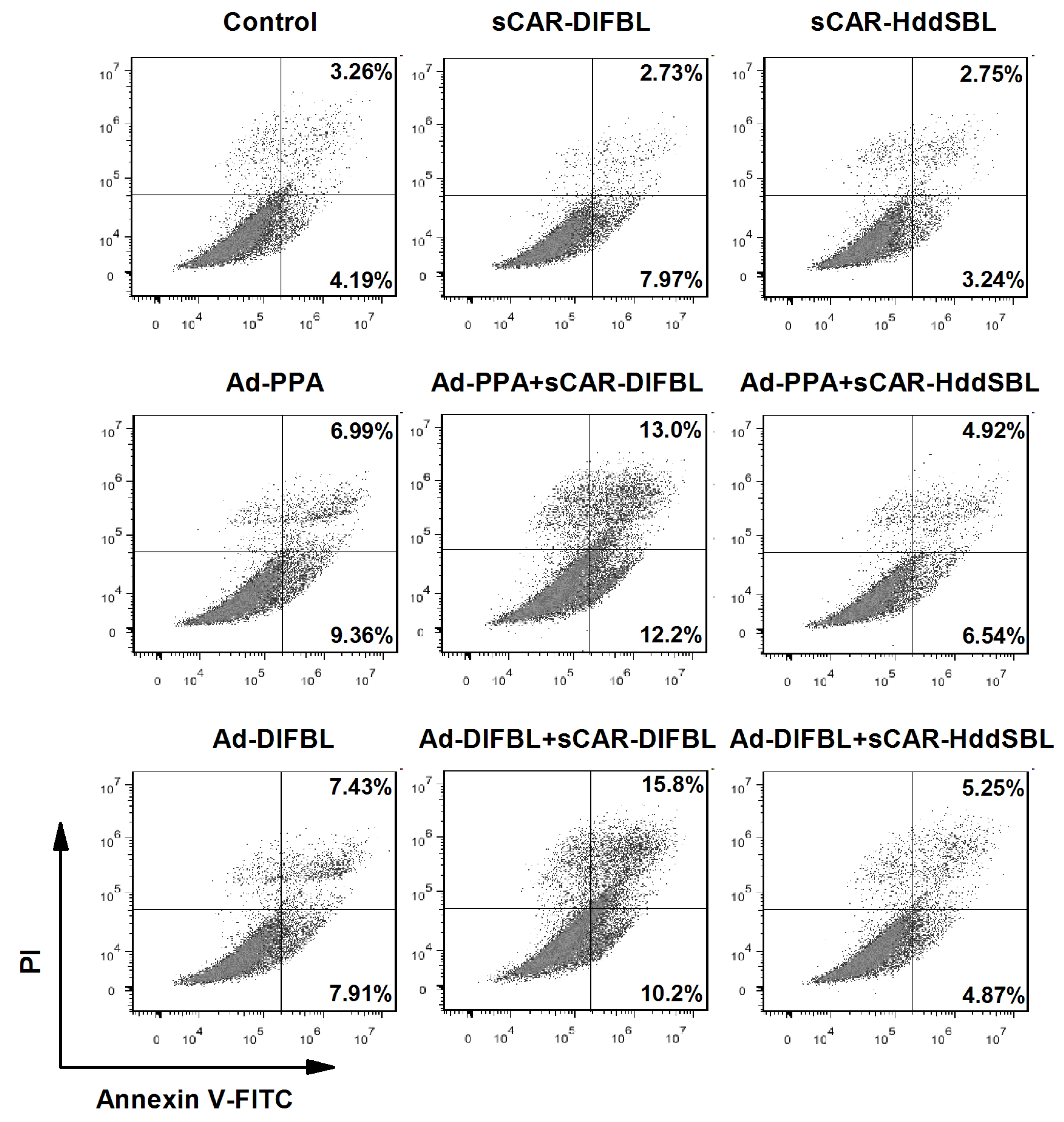
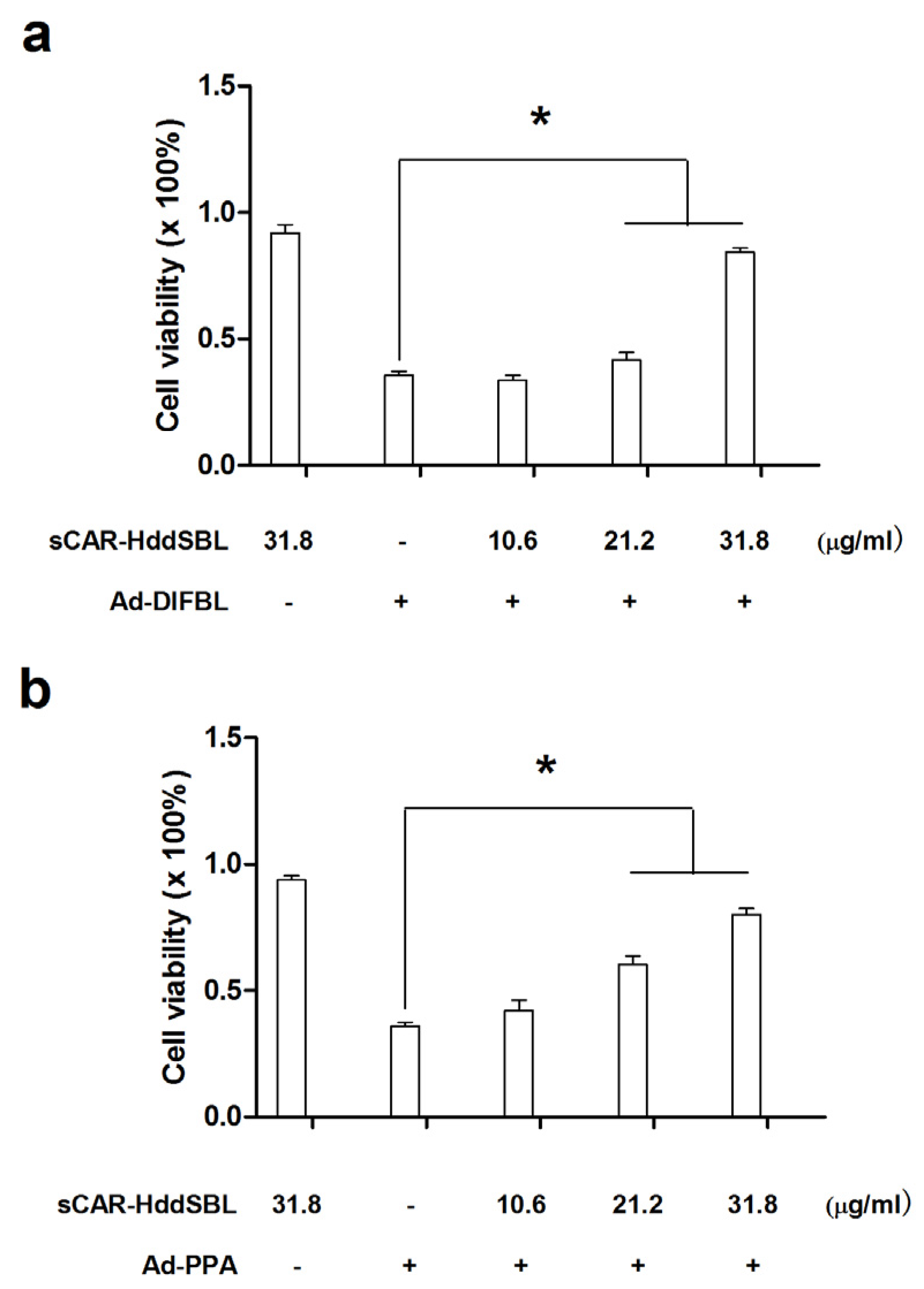
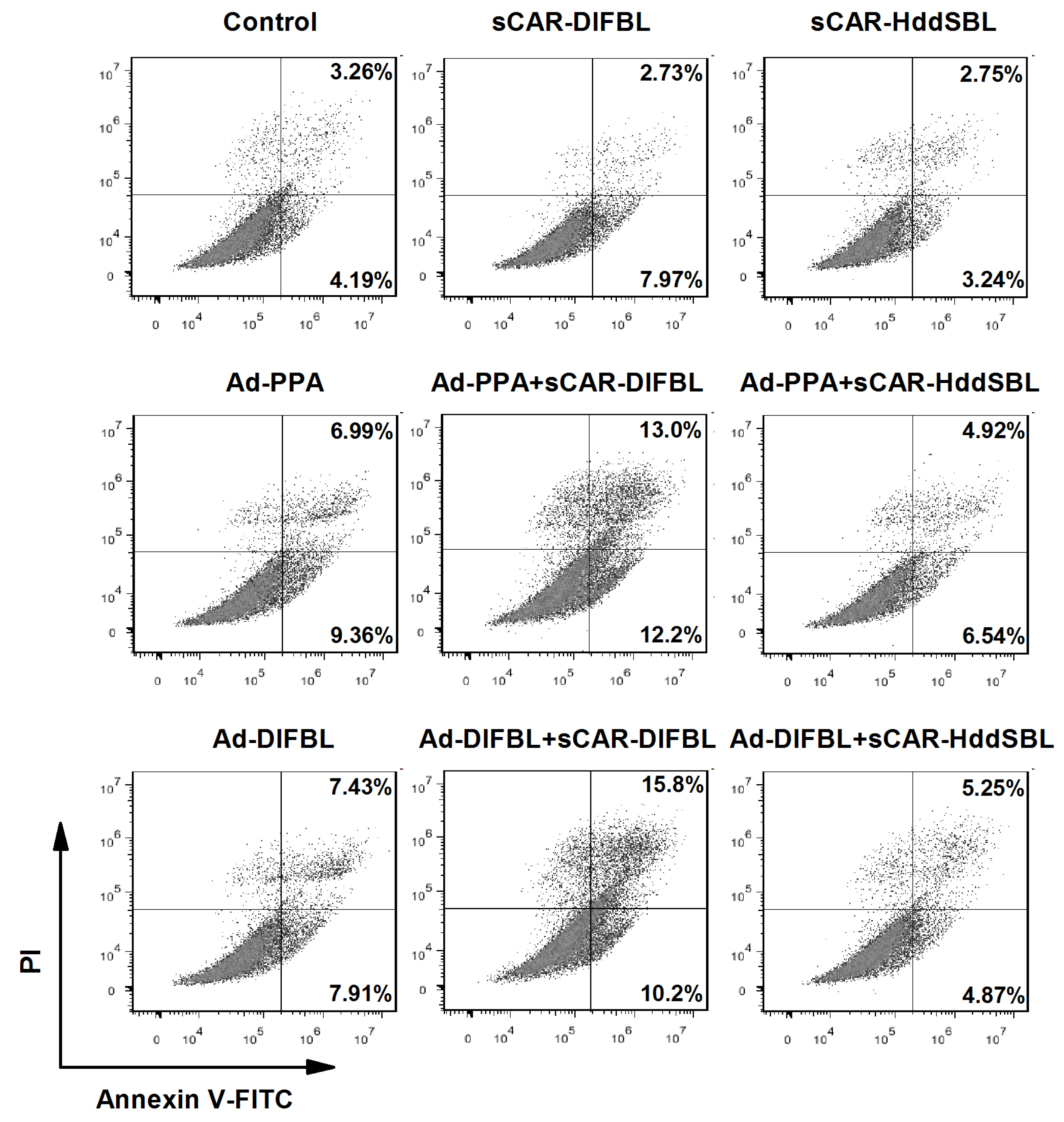
2.3. Recombinant sCAR-DlFBL and sCAR-HddSBL Had Different Effects on the Cytotoxicity of Ad-PPA and Ad-DlFBL in U87MG Cells
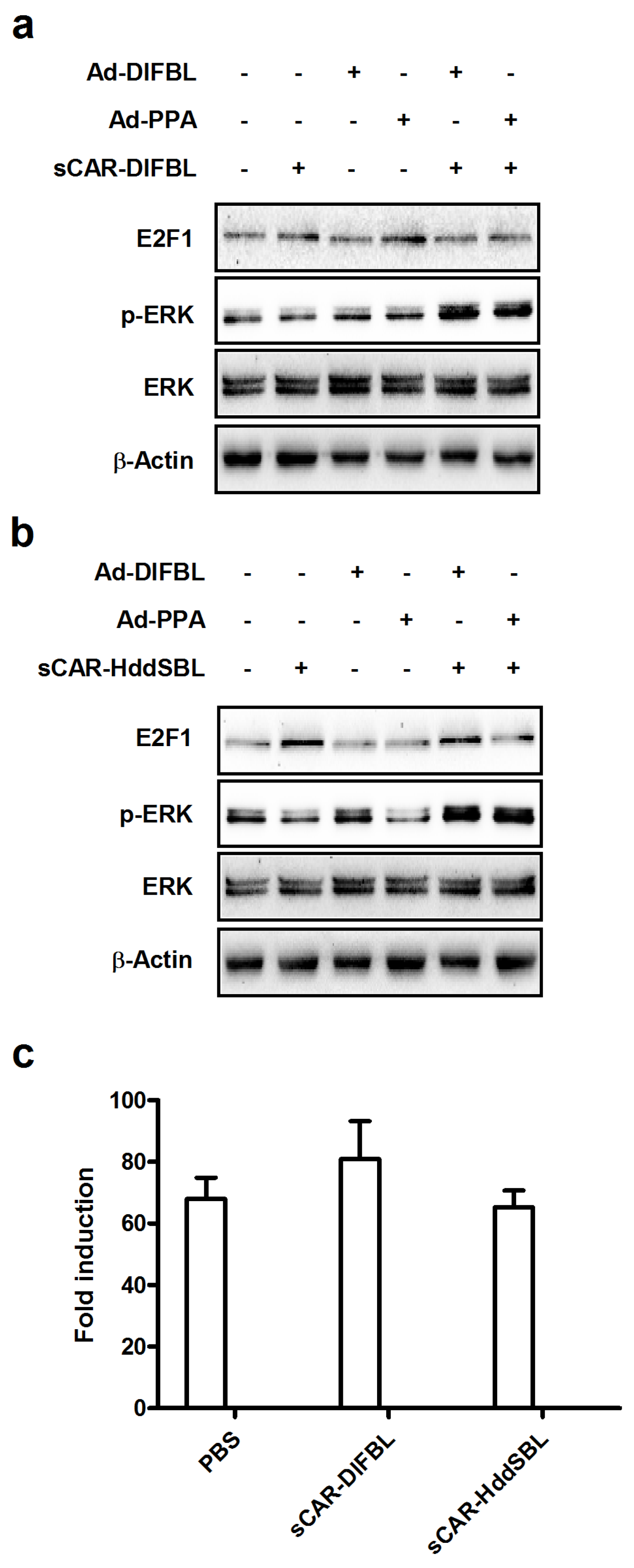
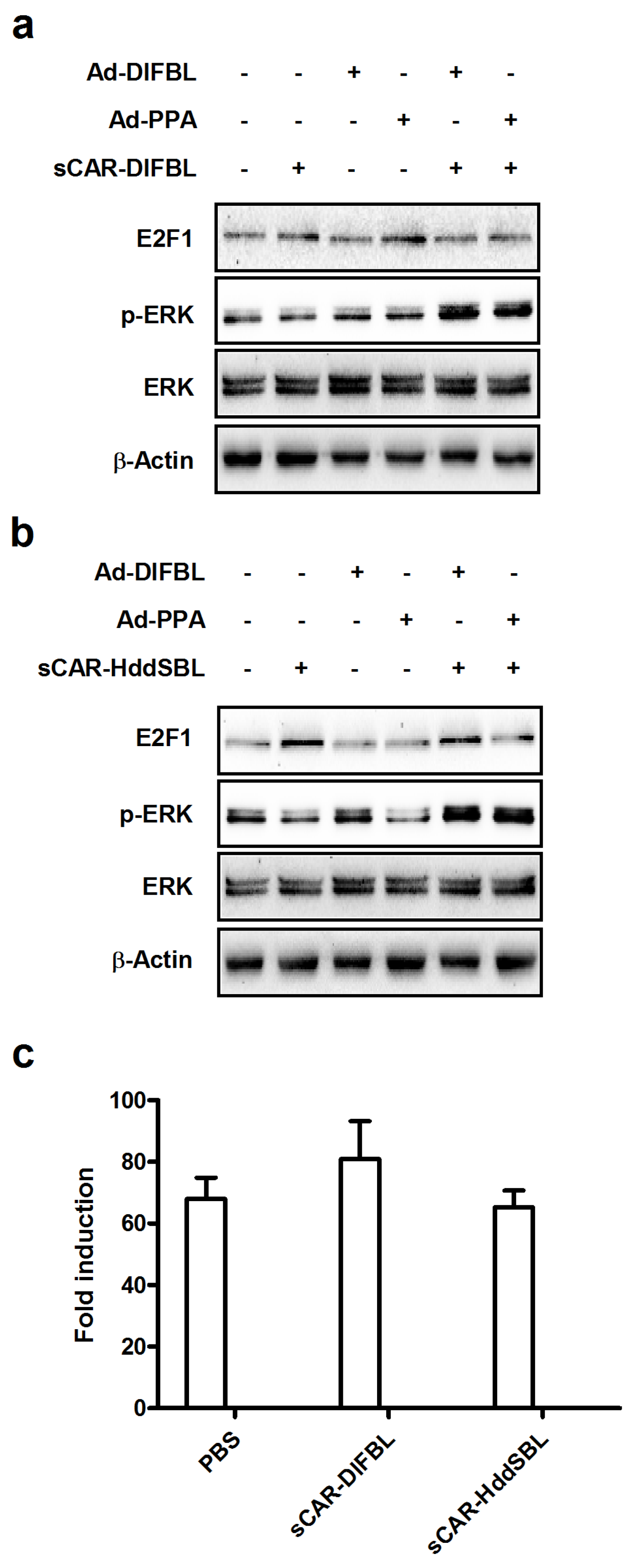
2.4. Recombinant sCAR-HddSBL Upregulated E2F1 Levels in U87MG Cells
3. Discussion
4. Materials and Methods
4.1. Production of sCAR-DlFBL and sCAR-HddSBL Fusion Proteins
4.2. Adenoviral Infection
4.3. Cytotoxicity Detection
4.4. Western Blot Analysis
4.5. Reporter Assay
4.6. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ochoa-Alvarez, J.A.; Krishnan, H.; Shen, Y.; Acharya, N.K.; Han, M.; McNulty, D.E.; Hasegawa, H.; Hyodo, T.; Senga, T.; Geng, J.G.; et al. Plant lectin can target receptors containing sialic acid, exemplified by podoplanin, to inhibit transformed cell growth and migration. PLoS ONE 2012, 7, e41845. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.P.; Yang, M.C.; Liu, H.S.; Lin, Y.S.; Lei, H.Y. Concanavalin a induces autophagy in hepatoma cells and has a therapeutic effect in a murine in situ hepatoma model. Hepatology 2007, 45, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, B.; Ghosh, K.; Yadav, N.; Kanade, S.R. A novel l-fucose-binding lectin from Fenneropenaeus indicus induced cytotoxicity in breast cancer cells. J. Biochem. 2016, 161, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Cheng, Y.; Bian, H.J.; Bao, J.K. Molecular mechanisms of Polygonatum cyrtonema lectin-induced apoptosis and autophagy in cancer cells. Autophagy 2009, 5, 253–255. [Google Scholar] [CrossRef] [PubMed]
- Terada, D.; Kawai, F.; Noguchi, H.; Unzai, S.; Hasan, I.; Fujii, Y.; Park, S.Y.; Ozeki, Y.; Tame, J.R. Crystal structure of mytilec, a galactose-binding lectin from the mussel mytilus galloprovincialis with cytotoxicity against certain cancer cell types. Sci. Rep. 2016, 6, 28344. [Google Scholar] [CrossRef] [PubMed]
- Hasan, I.; Sugawara, S.; Fujii, Y.; Koide, Y.; Terada, D.; Iimura, N.; Fujiwara, T.; Takahashi, K.G.; Kojima, N.; Rajia, S.; et al. Mytilec, a mussel r-type lectin, interacts with surface glycan gb3 on burkitt’s lymphoma cells to trigger apoptosis through multiple pathways. Mar. Drugs 2015, 13, 7377–7389. [Google Scholar] [CrossRef] [PubMed]
- Fujii, Y.; Dohmae, N.; Takio, K.; Kawsar, S.M.; Matsumoto, R.; Hasan, I.; Koide, Y.; Kanaly, R.A.; Yasumitsu, H.; Ogawa, Y.; et al. A lectin from the mussel mytilus galloprovincialis has a highly novel primary structure and induces glycan-mediated cytotoxicity of globotriaosylceramide-expressing lymphoma cells. J. Biol. Chem. 2012, 287, 44772–44783. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Li, N.; Luo, J.; Yu, M.; Huang, Y.; Wu, X.; Wu, H.; Liu, X.Y.; Li, G. Pinellia pedatisecta agglutinin interacts with the methylosome and induces cancer cell death. Oncogenesis 2012, 1, e29. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Yang, X.; Duan, X.; Cui, L.; Li, G. Exogenous expression of marine lectins dlfbl and sprbl induces cancer cell apoptosis possibly through prmt5-e2f-1 pathway. Sci. Rep. 2014, 4, 4505. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wu, L.; Duan, X.; Cui, L.; Luo, J.; Li, G. Adenovirus carrying gene encoding Haliotis discus discus sialic acid binding lectin induces cancer cell apoptosis. Mar. Drugs 2014, 12, 3994–4004. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Gao, Y.; Cui, L.; Wu, L.; Yang, X.; Chen, J. Anguilla japonica lectin 1 delivery through adenovirus vector induces apoptotic cancer cell death through interaction with prmt5. J. Gene Med. 2016, 18, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Li, X.; Wu, H.; Yang, X.; Zhang, Y.; Chen, L.; Wu, X.; Cui, L.; Wu, L.; Luo, J.; et al. Cd123 targeting oncolytic adenoviruses suppress acute myeloid leukemia cell proliferation in vitro and in vivo. Blood Cancer J. 2014, 4, e194. [Google Scholar] [CrossRef] [PubMed]
- Munkley, J.; Elliott, D.J. Hallmarks of glycosylation in cancer. Oncotarget 2016, 7, 35478–35489. [Google Scholar] [PubMed]
- Fry, S.A.; Afrough, B.; Lomax-Browne, H.J.; Timms, J.F.; Velentzis, L.S.; Leathem, A.J. Lectin microarray profiling of metastatic breast cancers. Glycobiology 2011, 21, 1060–1070. [Google Scholar] [CrossRef] [PubMed]
- Drake, P.M.; Schilling, B.; Niles, R.K.; Prakobphol, A.; Li, B.; Jung, K.; Cho, W.; Braten, M.; Inerowicz, H.D.; Williams, K.; et al. Lectin chromatography/mass spectrometry discovery workflow identifies putative biomarkers of aggressive breast cancers. J. Proteome Res. 2012, 11, 2508–2520. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Xie, X.; Liu, Y.; He, J.; Benitez, R.; Buckanovich, R.J.; Lubman, D.M. Identification and confirmation of differentially expressed fucosylated glycoproteins in the serum of ovarian cancer patients using a lectin array and lc-ms/ms. J. Proteome Res. 2012, 11, 4541–4552. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Simeone, D.M.; Brenner, D.E.; Anderson, M.A.; Shedden, K.A.; Ruffin, M.T.; Lubman, D.M. Pancreatic cancer serum detection using a lectin/glyco-antibody array method. J. Proteome Res. 2009, 8, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Osuga, T.; Takimoto, R.; Ono, M.; Hirakawa, M.; Yoshida, M.; Okagawa, Y.; Uemura, N.; Arihara, Y.; Sato, Y.; Tamura, F.; et al. Relationship between increased fucosylation and metastatic potential in colorectal cancer. J. Natl. Cancer Inst. 2016, 108, djw038. [Google Scholar] [CrossRef] [PubMed]
- Batabyal, S.K.; Majhi, R.; Basu, P.S. Clinical utility of the interaction between lectin and serum prostate specific antigen in prostate cancer. Neoplasma 2009, 56, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Ahn, Y.H.; Shin, P.M.; Oh, N.R.; Park, G.W.; Kim, H.; Yoo, J.S. A lectin-coupled, targeted proteomic mass spectrometry (mrm ms) platform for identification of multiple liver cancer biomarkers in human plasma. J. Proteom. 2012, 75, 5507–5515. [Google Scholar] [CrossRef] [PubMed]
- Norton, P.; Comunale, M.A.; Herrera, H.; Wang, M.; Houser, J.; Wimmerova, M.; Romano, P.R.; Mehta, A. Development and application of a novel recombinant aleuria aurantia lectin with enhanced core fucose binding for identification of glycoprotein biomarkers of hepatocellular carcinoma. Proteomics 2016, 16, 3126–3136. [Google Scholar] [CrossRef] [PubMed]
- Munkley, J.; Mills, I.G.; Elliott, D.J. The role of glycans in the development and progression of prostate cancer. Nat. Rev. Urol. 2016, 13, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Sawanobori, A.; Moriwaki, K.; Takamatsu, S.; Kamada, Y.; Miyoshi, E. A glycoproteomic approach to identify novel glycomarkers for cancer stem cells. Proteomics 2016, 16, 3073–3080. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Huang, D.; Chen, K.Y.; Cui, M.; Wang, W.; Huang, X.; Awadellah, A.; Li, Q.; Friedman, A.; Xin, W.W.; et al. Fucosylation deficiency in mice leads to colitis and adenocarcinoma. Gastroenterology 2017, 152, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.P.; Xu, X.Y.; Fang, M.; Wang, H.; You, Q.; Yi, C.H.; Ji, J.; Gu, X.; Zhou, P.T.; Cheng, C.; et al. Decreased core-fucosylation contributes to malignancy in gastric cancer. PLoS ONE 2014, 9, e94536. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Woods, E.C.; Vukojicic, P.; Bertozzi, C.R. Precision glycocalyx editing as a strategy for cancer immunotherapy. Proc. Natl. Acad. Sci. USA 2016, 113, 10304–10309. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Dong, G.; Wang, S.; Zhu, S.; Shen, Y.; Li, G. Pinellia pedatisecta agglutinin-based lectin blot analysis distinguishes between glycosylation patterns in various cancer cell lines. Oncol. Lett. 2014, 8, 837–840. [Google Scholar] [PubMed]
- Li, G.C.; Li, N.; Zhang, Y.H.; Li, X.; Wang, Y.G.; Liu, X.Y.; Qian, W.B.; Liu, X.C. Mannose-exposing myeloid leukemia cells detected by the scar-ppa fusion protein. Int. J. Hematol. 2009, 89, 611–617. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Yang, X.; Wu, L.; Yu, M.; Li, X.; Li, N.; Wang, S.; Li, G. Pinellia pedatisecta agglutinin targets drug resistant k562/adr leukemia cells through binding with sarcolemmal membrane associated protein and enhancing macrophage phagocytosis. PLoS ONE 2013, 8, e74363. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Y. Targeting gene-virotherapy of cancer and its prosperity. Cell Res. 2006, 16, 879–886. [Google Scholar] [CrossRef] [PubMed]
- Curiel, D.T. The development of conditionally replicative adenoviruses for cancer therapy. Clin. Cancer Res. 2000, 6, 3395–3399. [Google Scholar] [PubMed]
- Hawkins, L.K.; Lemoine, N.R.; Kirn, D. Oncolytic biotherapy: A novel therapeutic plafform. Lancet Oncol. 2002, 3, 17–26. [Google Scholar] [CrossRef]
- Kirn, D.; Martuza, R.L.; Zwiebel, J. Replication-selective virotherapy for cancer: Biological principles, risk management and future directions. Nat. Med. 2001, 7, 781–787. [Google Scholar] [CrossRef] [PubMed]
- Bewley, M.C.; Springer, K.; Zhang, Y.B.; Freimuth, P.; Flanagan, J.M. Structural analysis of the mechanism of adenovirus binding to its human cellular receptor, car. Science 1999, 286, 1579–1583. [Google Scholar] [CrossRef] [PubMed]
- Bergelson, J.M.; Cunningham, J.A.; Droguett, G.; Kurt-Jones, E.A.; Krithivas, A.; Hong, J.S.; Horwitz, M.S.; Crowell, R.L.; Finberg, R.W. Isolation of a common receptor for coxsackie b viruses and adenoviruses 2 and 5. Science 1997, 275, 1320–1323. [Google Scholar] [CrossRef] [PubMed]
- Qian, W.; Liu, J.; Tong, Y.; Yan, S.; Yang, C.; Yang, M.; Liu, X. Enhanced antitumor activity by a selective conditionally replicating adenovirus combining with mda-7/interleukin-24 for b-lymphoblastic leukemia via induction of apoptosis. Leukemia 2008, 22, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Wu, H.; Cui, L.; Gao, Y.; Chen, L.; Li, X.; Liang, T.; Yang, X.; Cheng, J.; Luo, J. Cd47-retargeted oncolytic adenovirus armed with melanoma differentiation-associated gene-7/interleukin-24 suppresses in vivo leukemia cell growth. Oncotarget 2015, 6, 43496–43507. [Google Scholar] [PubMed]
- La Thangue, N.B. The yin and yang of e2f-1: Balancing life and death. Nat. Cell Biol. 2003, 5, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Gala, S.; Marreiros, A.; Stewart, G.J.; Williamson, P. Overexpression of e2f-1 leads to cytokine-independent proliferation and survival in the hematopoietic cell line baf-b03. Blood 2001, 97, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Levine, A.J. P53 and e2f-1 cooperate to mediate apoptosis. Proc. Natl. Acad. Sci. USA 1994, 91, 3602–3606. [Google Scholar] [CrossRef] [PubMed]
- Irwin, M.; Marin, M.C.; Phillips, A.C.; Seelan, R.S.; Smith, D.I.; Liu, W.; Flores, E.R.; Tsai, K.Y.; Jacks, T.; Vousden, K.H.; et al. Role for the p53 homologue p73 in e2f-1-induced apoptosis. Nature 2000, 407, 645–648. [Google Scholar] [PubMed]
- Stevens, C.; Smith, L.; La Thangue, N.B. Chk2 activates e2f-1 in response to DNA damage. Nat. Cell Biol. 2003, 5, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.C.; Zheng, S.; Munro, S.; Liu, G.; Carr, S.M.; Moehlenbrink, J.; Lu, Y.C.; Stimson, L.; Khan, O.; Konietzny, R.; et al. Arginine methylation controls growth regulation by e2f-1. EMBO J. 2012, 31, 1785–1797. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Moehlenbrink, J.; Lu, Y.C.; Zalmas, L.P.; Sagum, C.A.; Carr, S.; McGouran, J.F.; Alexander, L.; Fedorov, O.; Munro, S.; et al. Arginine methylation-dependent reader-writer interplay governs growth control by e2f-1. Mol. Cell 2013, 52, 37–51. [Google Scholar] [CrossRef] [PubMed]



© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, B.; Mei, S.; Cui, L.; Zhao, Z.; Chen, J.; Wu, T.; Li, G. Marine Lectins DlFBL and HddSBL Fused with Soluble Coxsackie-Adenovirus Receptor Facilitate Adenovirus Infection in Cancer Cells BUT Have Different Effects on Cell Survival. Mar. Drugs 2017, 15, 73. https://doi.org/10.3390/md15030073
Wu B, Mei S, Cui L, Zhao Z, Chen J, Wu T, Li G. Marine Lectins DlFBL and HddSBL Fused with Soluble Coxsackie-Adenovirus Receptor Facilitate Adenovirus Infection in Cancer Cells BUT Have Different Effects on Cell Survival. Marine Drugs. 2017; 15(3):73. https://doi.org/10.3390/md15030073
Chicago/Turabian StyleWu, Bingbing, Shengsheng Mei, Lianzhen Cui, Zhenzhen Zhao, Jianhong Chen, Tao Wu, and Gongchu Li. 2017. "Marine Lectins DlFBL and HddSBL Fused with Soluble Coxsackie-Adenovirus Receptor Facilitate Adenovirus Infection in Cancer Cells BUT Have Different Effects on Cell Survival" Marine Drugs 15, no. 3: 73. https://doi.org/10.3390/md15030073