
Marine Diterpenes: Molecular Modeling of Thrombin Inhibitors with Potential Biotechnological Application as an Antithrombotic

 , ,
, ,
Abstract
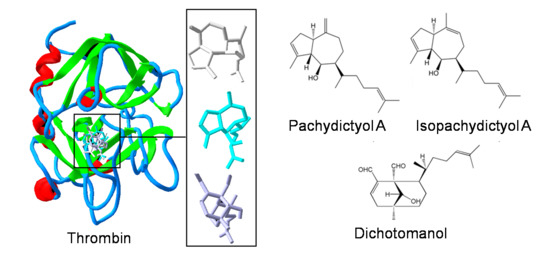
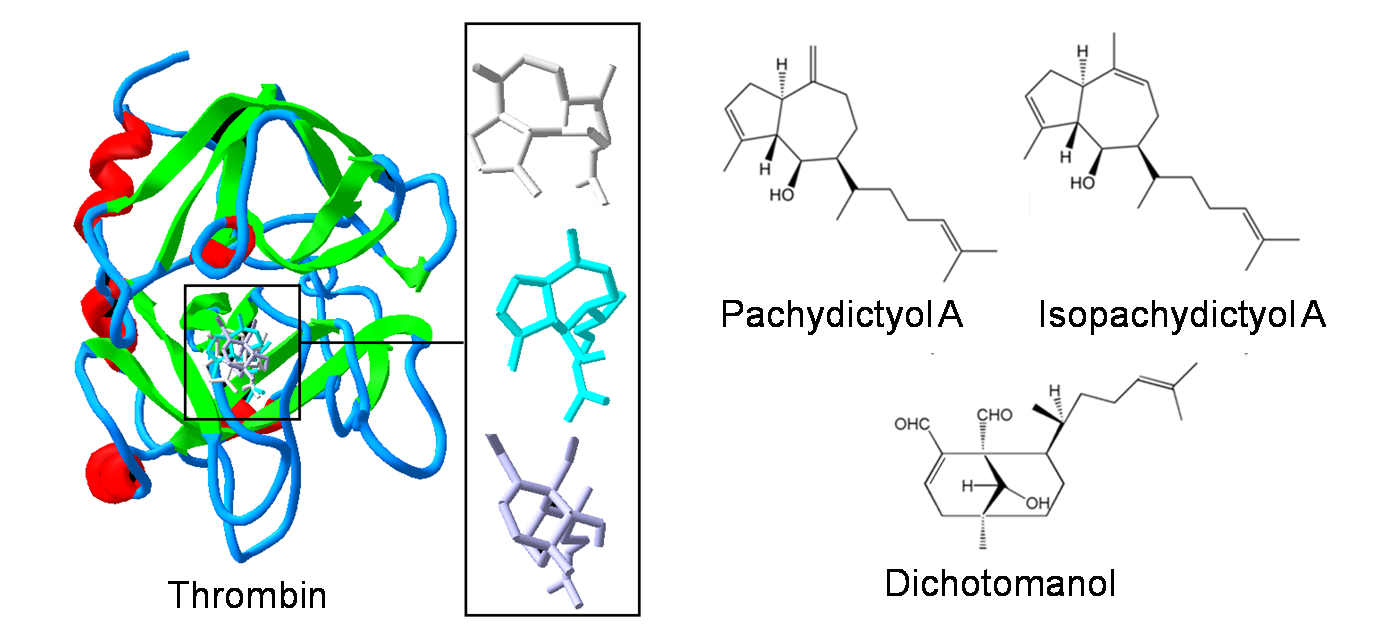
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
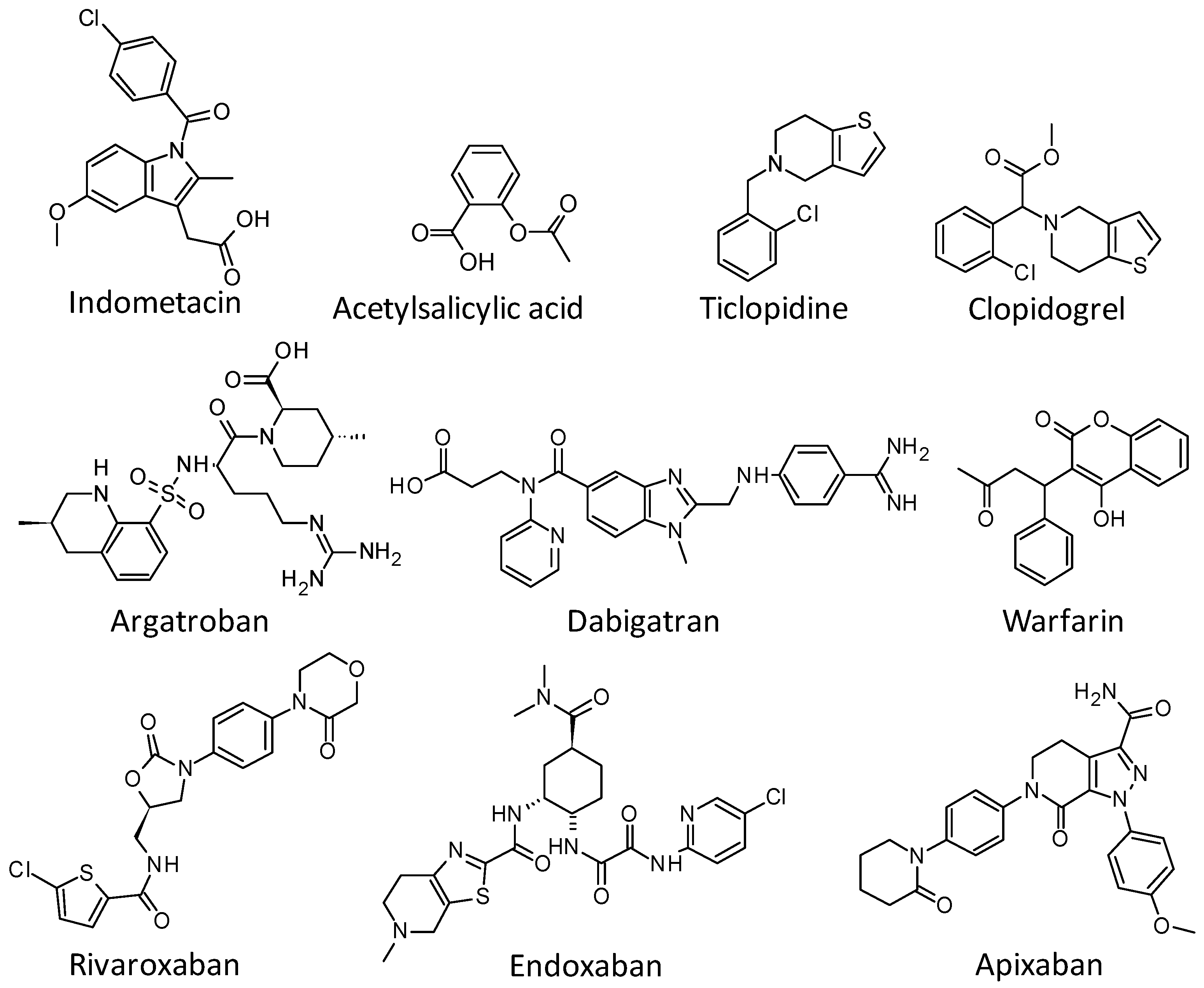
1. Introduction
2. Results and Discussion
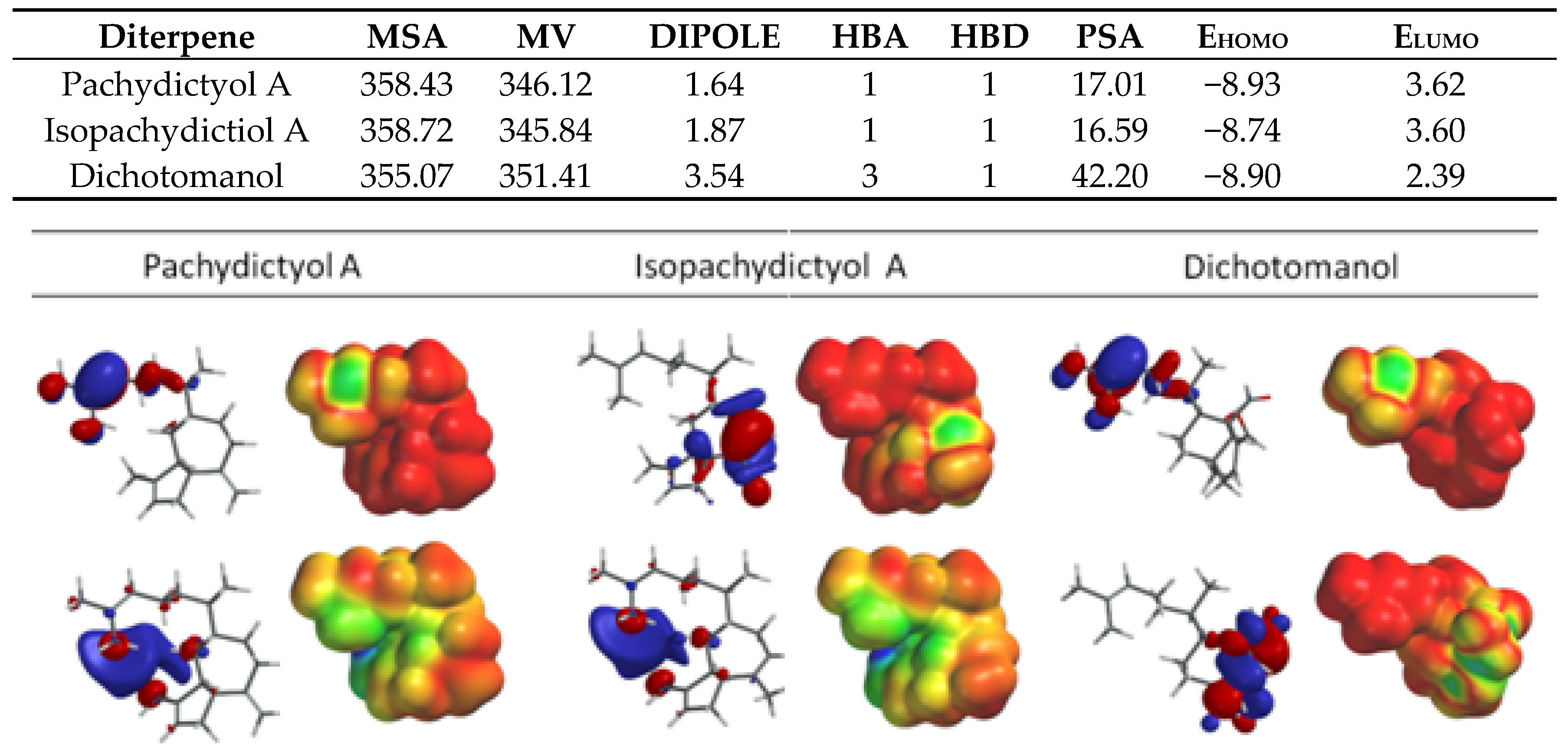
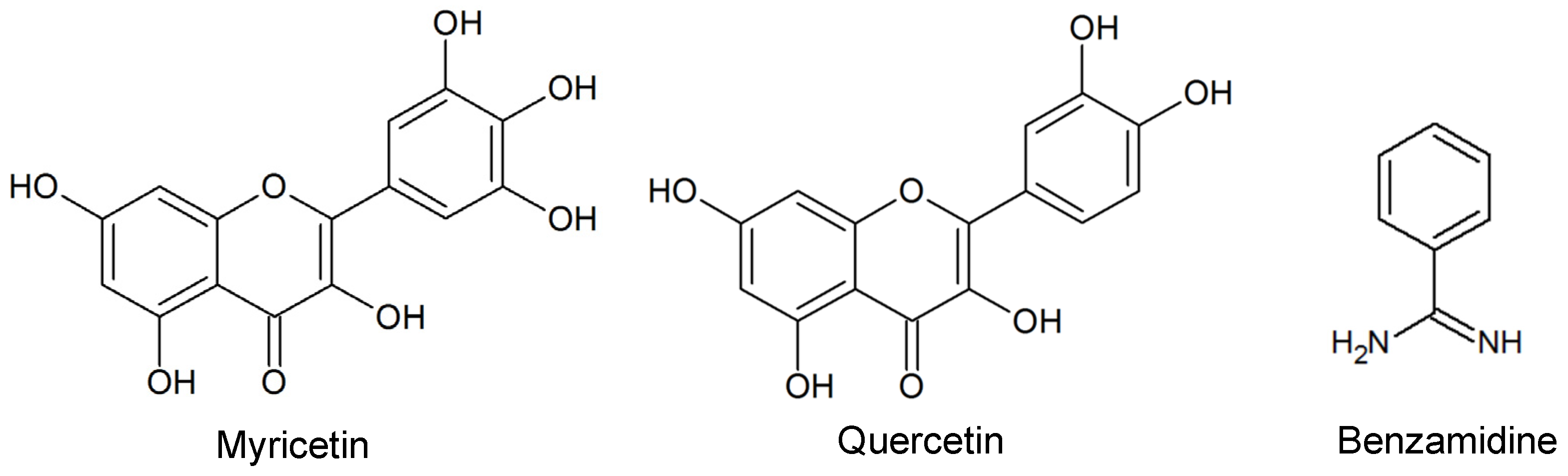
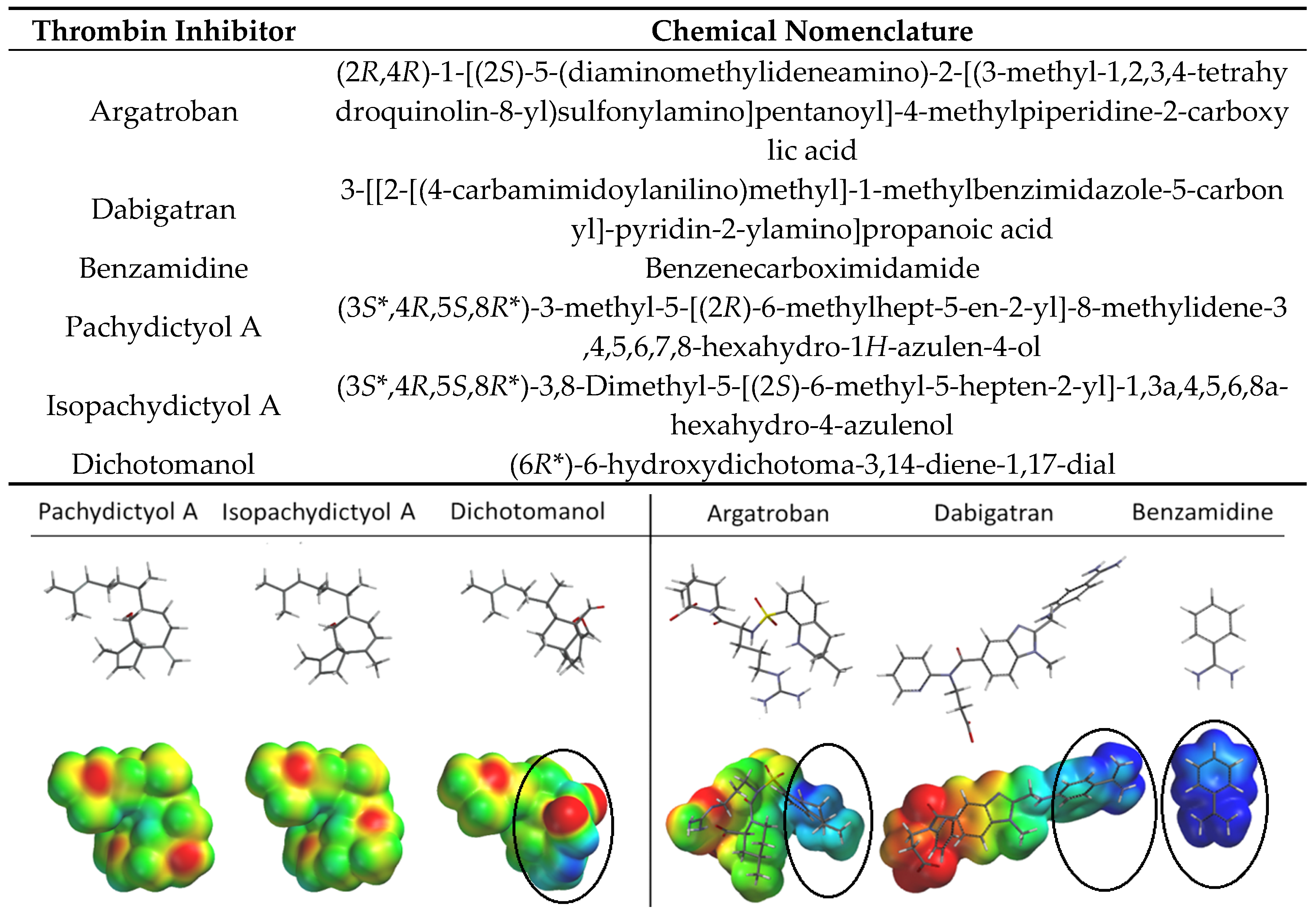
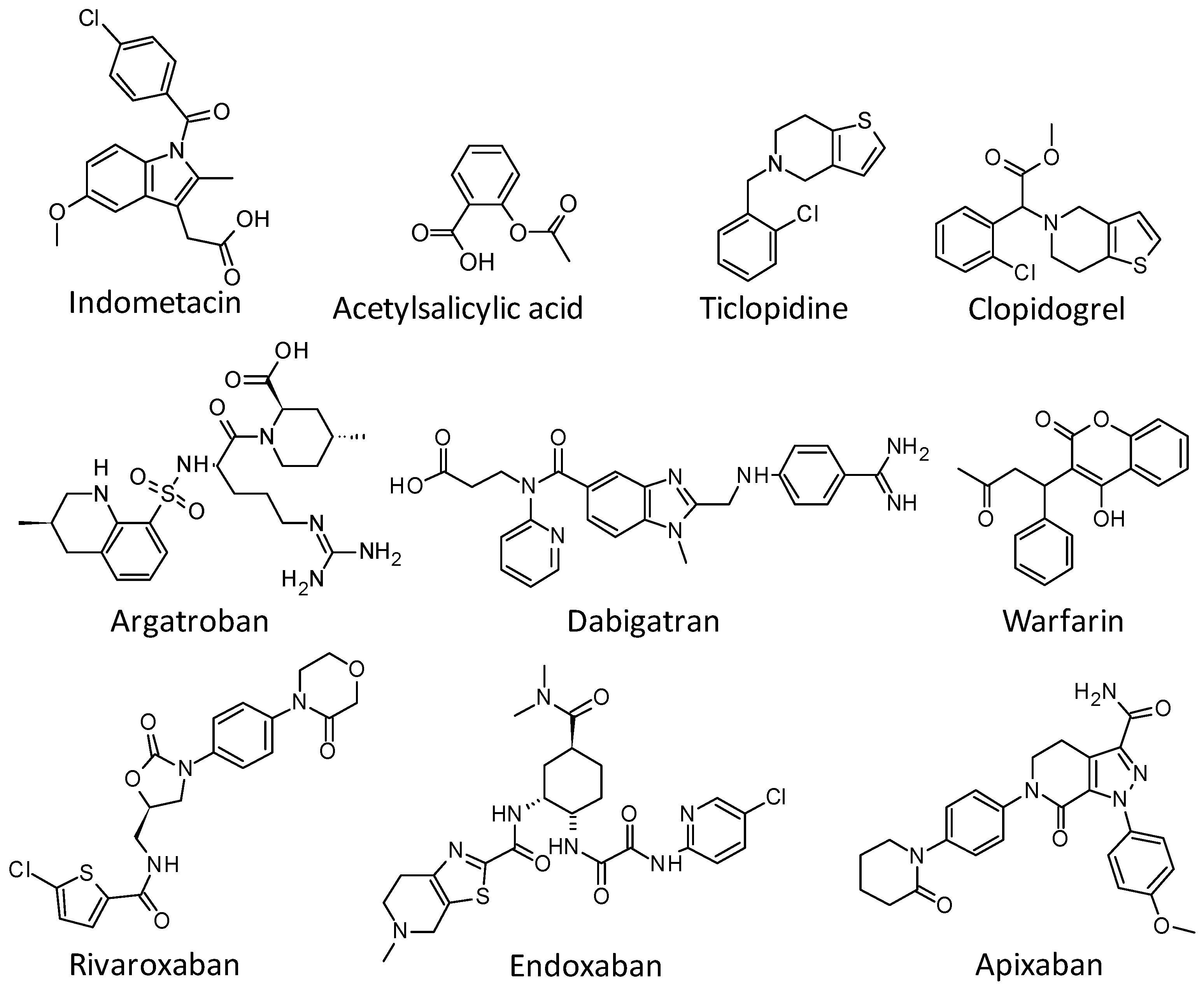
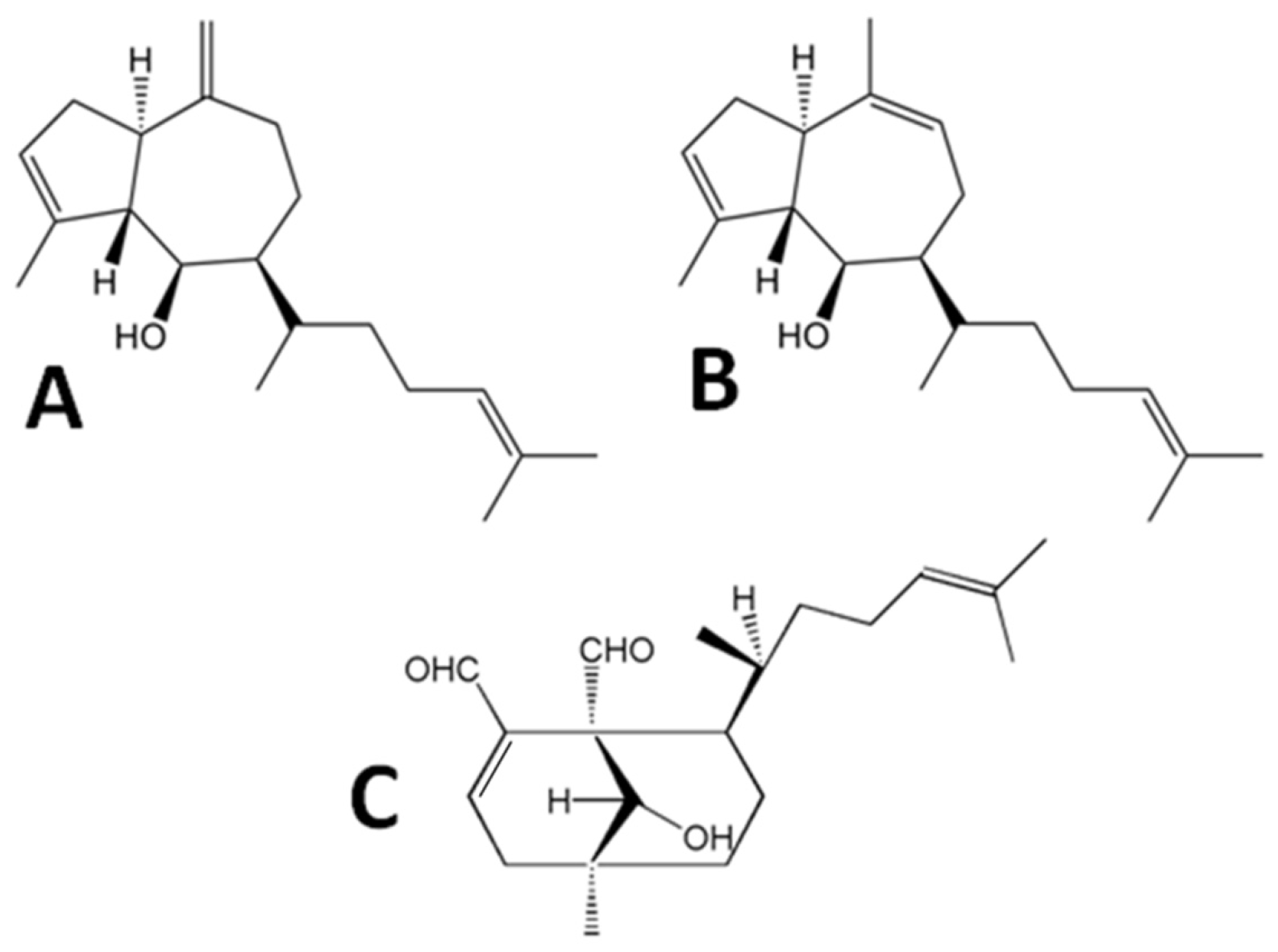
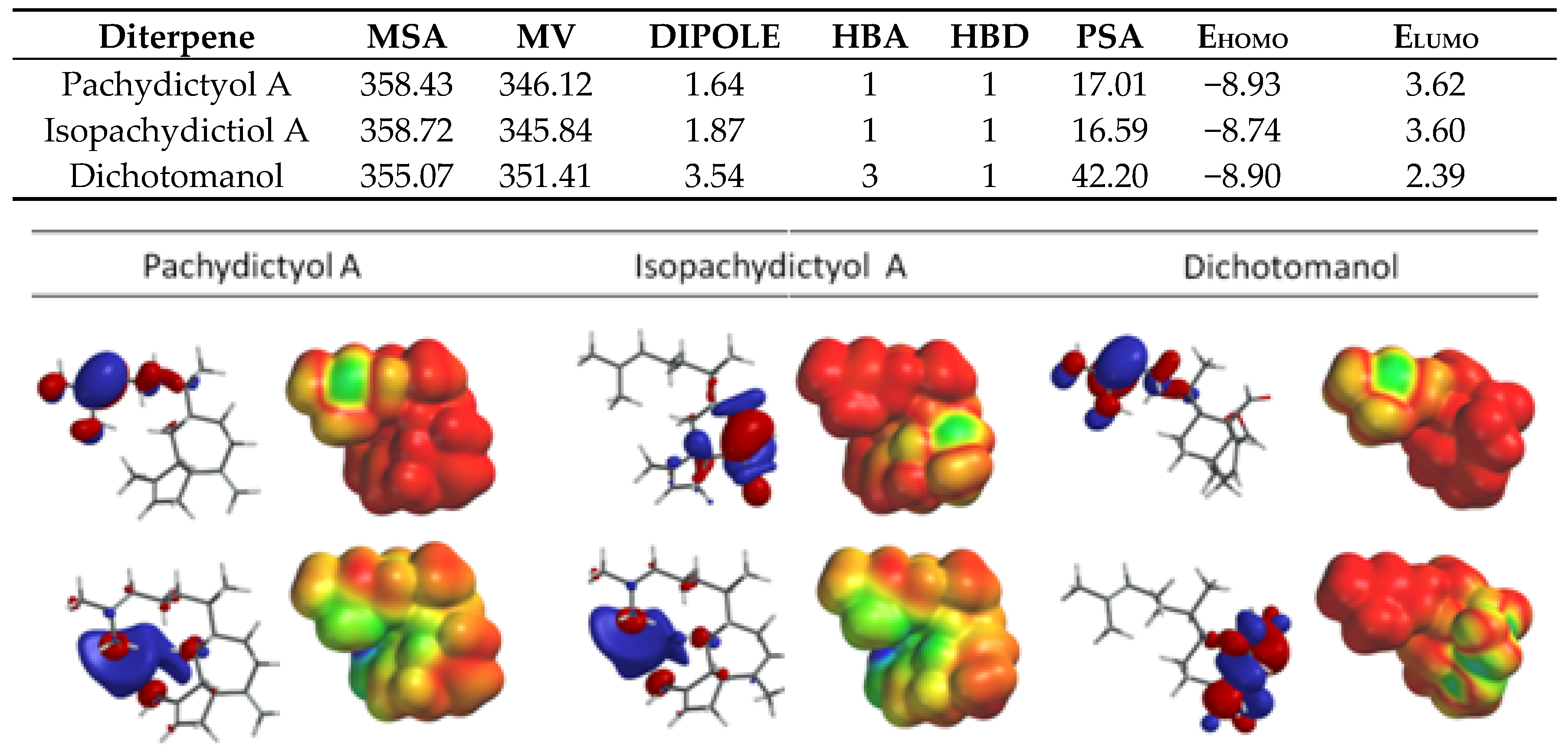
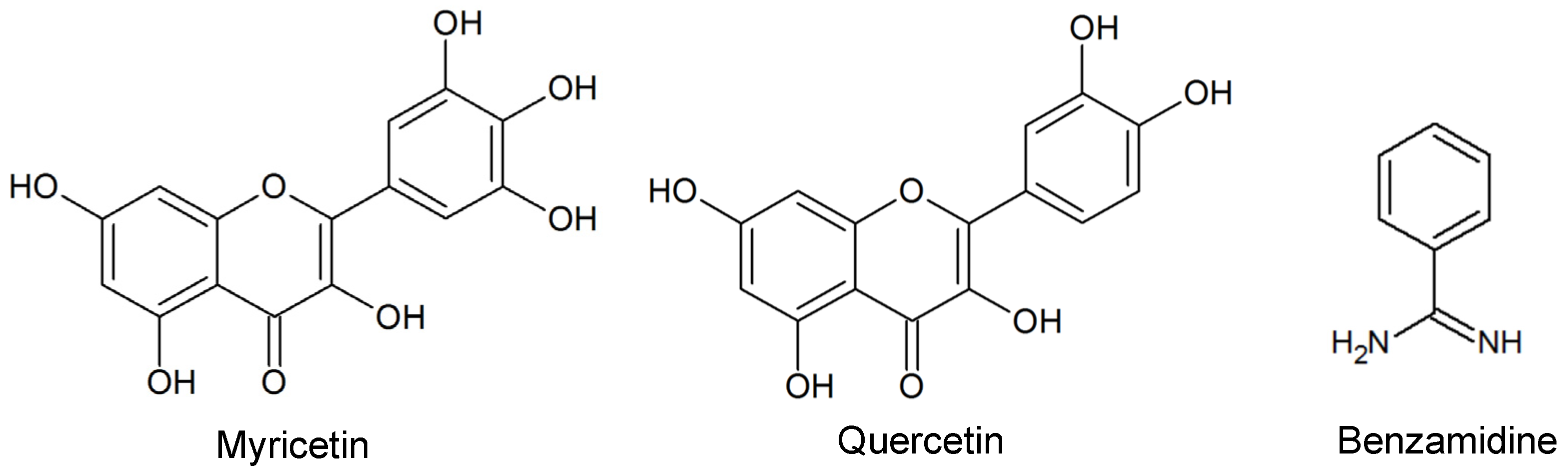
2.1. Diterpenes Structure-Activity Relationship Analysis
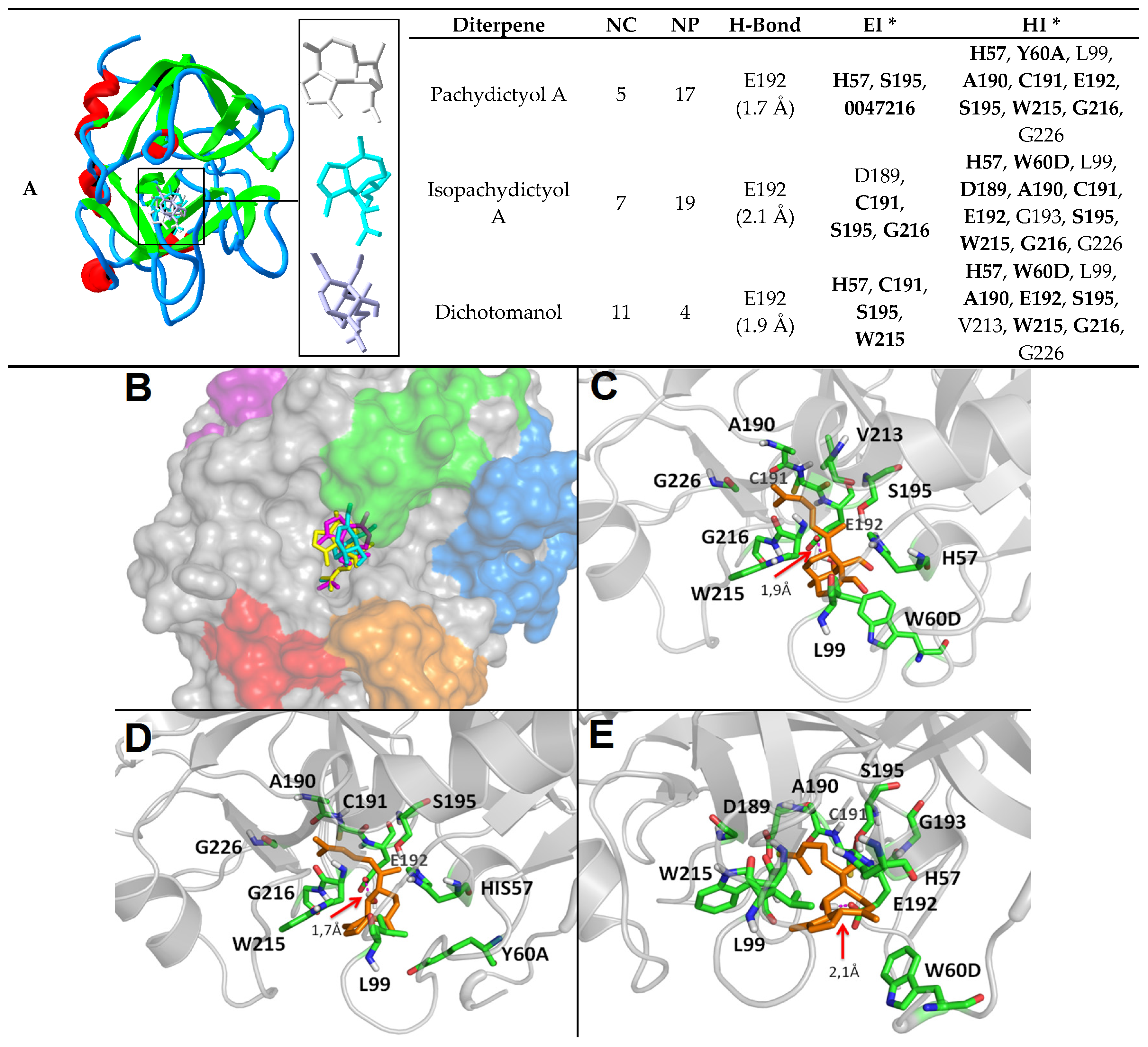
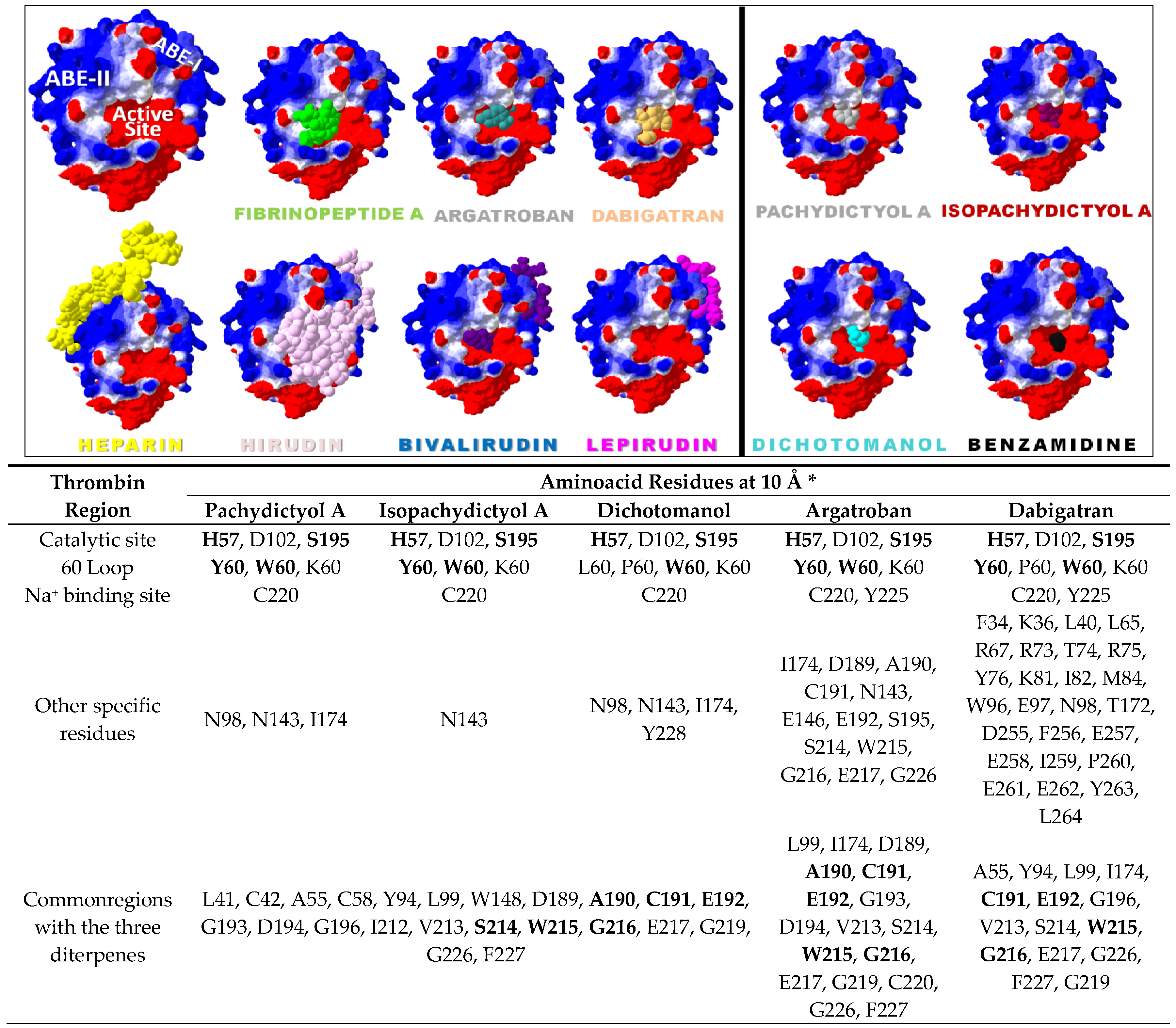
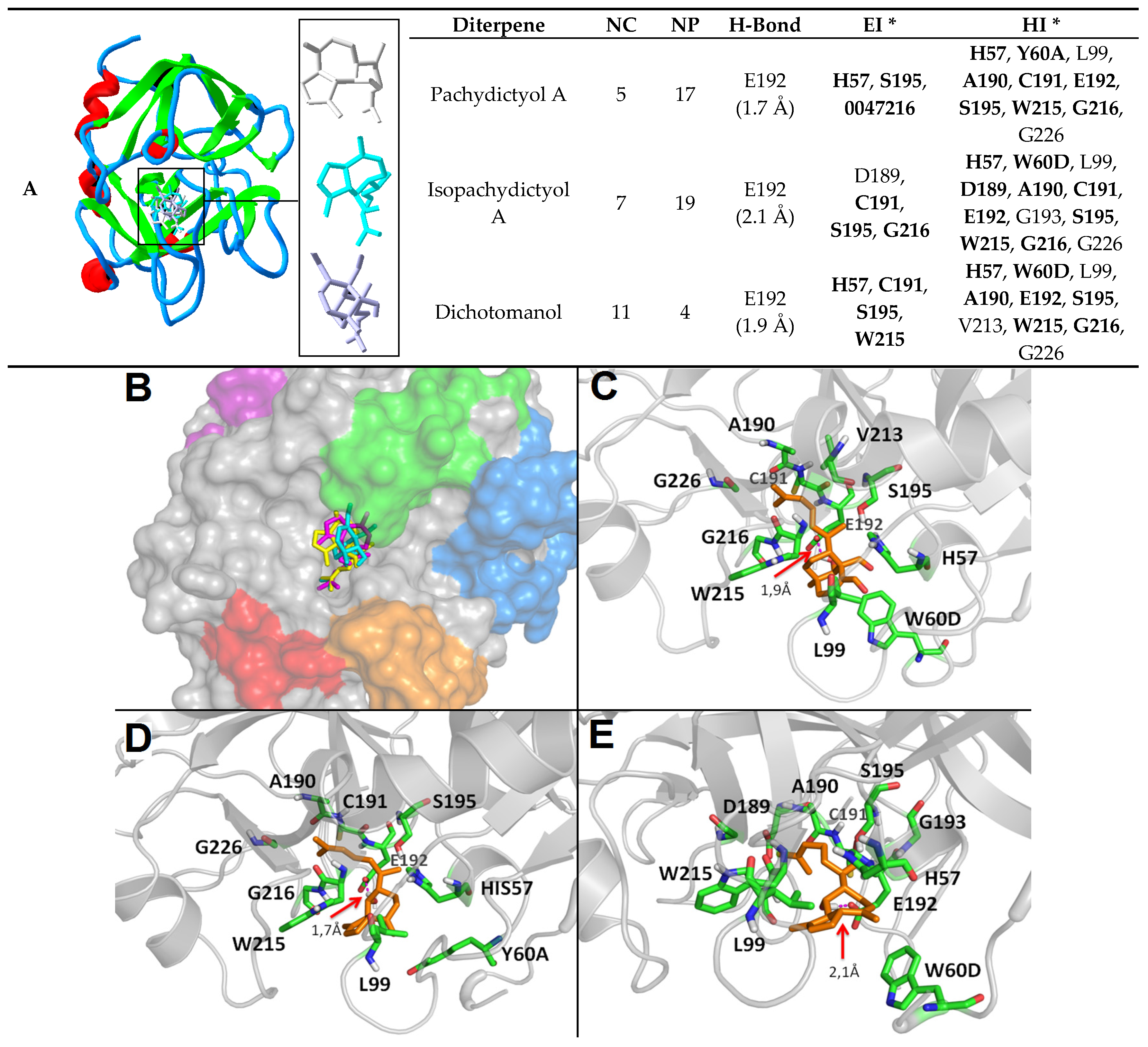
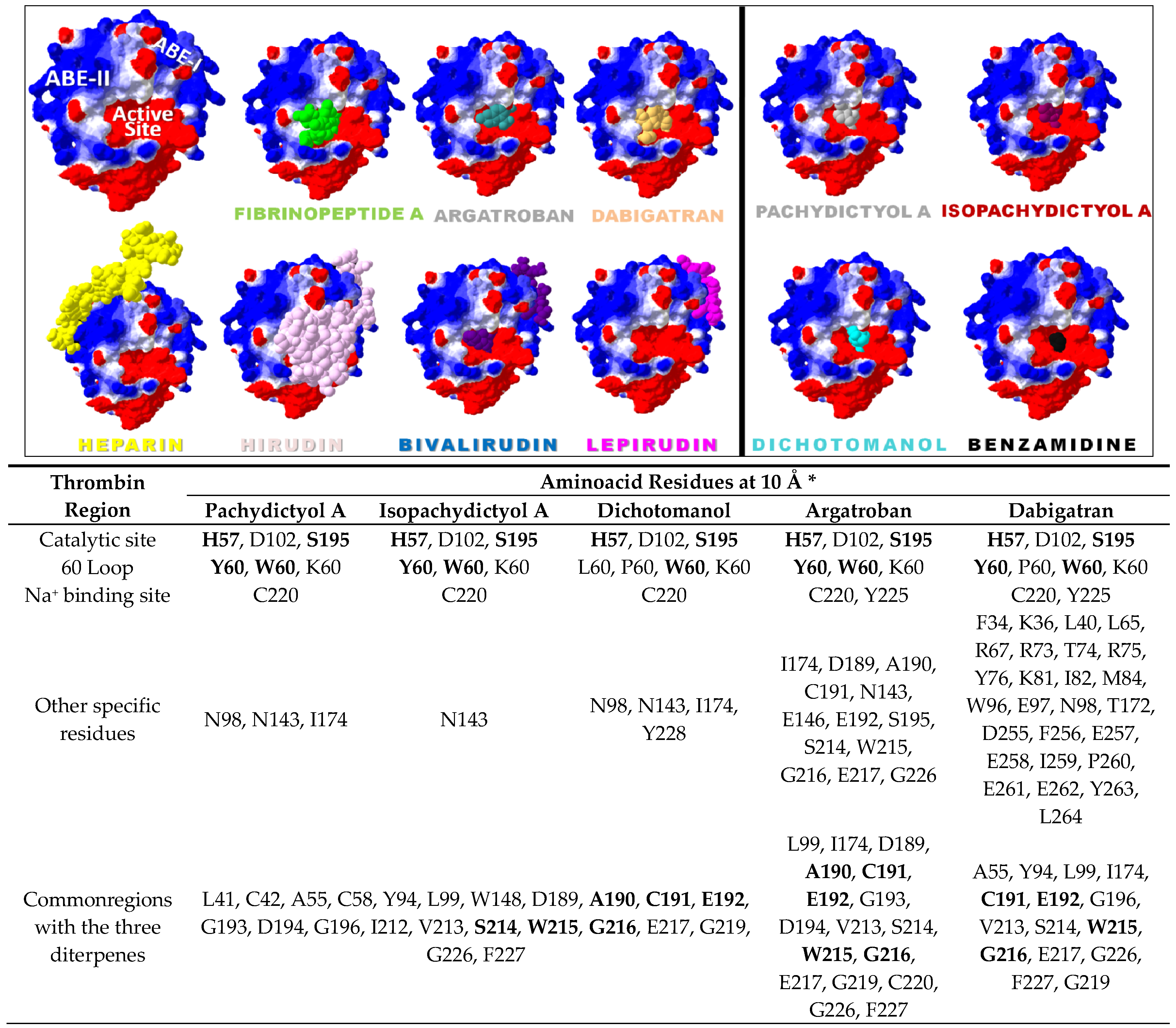
2.2. Diterpenes Molecular Docking Evaluation
3. Materials and Methods
3.1. Structure-Activity Relationship Analysis
3.2. Molecular Docking Study
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Shobharani, P.; Nanishankar, V.H.; Halami, P.M.; Sachindra, N.M. Antioxidant and anticoagulant activity of polyphenol and polysaccharides from fermented Sargassum sp. Int. J. Biol. Macromol. 2014, 65, 542–548. [Google Scholar] [CrossRef] [PubMed]
- De Andrade Moura, L.; De Almeida, A.C.M.; Domingos, T.F.S.; Ortiz-Ramirez, F.; Cavalcanti, D.N.; Teixeira, V.L.; Fuly, A.L. Antiplatelet and anticoagulant effects of diterpenes isolated from the marine alga, Dictyotamenstrualis. Mar. Drugs 2014, 12, 2471–2484. [Google Scholar] [CrossRef] [PubMed]
- Organization, W.H. Global Atlas on Cardiovascular Disease Prevention and Control; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- Behan, M.; Storey, R. Antiplatelet therapy in cardiovascular disease. Postgrad. Med. J. 2004, 80, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Yang, E. A clinician’s perspective: Novel oral anticoagulants to reduce the risk of stroke in nonvalvular atrial fibrillation full speed ahead or proceed with caution? Vasc. Health Risk Manag. 2014, 10, 507. [Google Scholar] [CrossRef] [PubMed]
- Roderick, P.J.; Wilkes, H.C.; Meade, T.W. The gastrointestinal toxicity of aspirin: An overview of randomised controlled trials. Br. J. Clin. Pharmacol. 1993, 35, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Noble, S.; Goa, K.L. Ticlopidine: A review of its pharmacology, clinical efficacy and tolerability in the prevention of cerebral ischaemia and stroke. Drugs Aging 1996, 8, 214–232. [Google Scholar] [CrossRef] [PubMed]
- Wisler, J.W.; Becker, R.C. Antithrombotic therapy: New areas to understand efficacy and bleeding. Expert Opin. Ther. Targets 2014, 18, 1427–1434. [Google Scholar] [CrossRef] [PubMed]
- Barros Gomes Camara, R.; Silva Costa, L.; Pereira Fidelis, G.; Duarte BarretoNobre, L.T.; Dantas-Santos, N.; Lima Cordeiro, S.; Santana Santos Pereira Costa, M.; Guimaraes Alves, L.; Oliveira Rocha, H.A. Heterofucans from the Brown Seaweed Canistrocarpuscervicornis with Anticoagulant and Antioxidant Activities. Mar. Drugs 2011, 9, 124–138. [Google Scholar] [CrossRef] [PubMed]
- Dahmer, T.; Berger, M.; Barlette, A.G.; Reck, J.; Segalin, J.; Verza, S.; Ortega, G.G.; Gnoatto, S.C.B.; Guimarães, J.A.; Verli, H.; et al. Antithrombotic Effect of ChikusetsusaponinIVa Isolated from Ilex paraguariensis (Maté). J. Med. Food 2012, 15, 1073–1080. [Google Scholar] [CrossRef] [PubMed]
- De Andrade Moura, L.; Ortiz-Ramirez, F.; Cavalcanti, D.N.; Ribeiro, S.M.; Muricy, G.; Teixeira, V.L.; Fuly, A.L. Evaluation of Marine Brown Algae and Sponges from Brazil as Anticoagulant and Antiplatelet Products. Mar. Drugs 2011, 9, 1346–1358. [Google Scholar] [CrossRef] [PubMed]
- Spronk, H.M.H.; Borissoff, J.I.; ten Cate, H. New Insights into Modulation of Thrombin Formation. Curr. Atheroscler. Rep. 2013, 15, 363. [Google Scholar] [CrossRef] [PubMed]
- Ten Cate, H.; Hemker, H.C. Thrombin Generation and Atherothrombosis: What Does the Evidence Indicate? J. Am. Heart Assoc. 2016, 5, e003553. [Google Scholar] [CrossRef] [PubMed]
- Bode, W.; Turk, D.; Karshikov, A. The refined 1.9-A X-ray crystal structure of D-Phe-Pro-Arg chloromethylketone-inhibited human alpha-thrombin: Structure analysis, overall structure, electrostatic properties, detailed active-site geometry, and structure-function relationships. Protein Sci. 1992, 1, 426–471. [Google Scholar] [CrossRef] [PubMed]
- Huntington, J.A. Molecular recognition mechanisms of thrombin. J. Thromb. Haemost. 2005, 3, 1861–1872. [Google Scholar] [CrossRef] [PubMed]
- Koh, C.Y.; Kumar, S.; Kazimirova, M.; Nuttall, P.A.; Radhakrishnan, U.P.; Kim, S.; Jagadeeswaran, P.; Imamura, T.; Mizuguchi, J.; Iwanaga, S.; et al. Crystal structure of thrombin in complex with s-variegin: Insights of a novel mechanism of inhibition and design of tunable thrombin inhibitors. PLoS ONE 2011, 6, e26367. [Google Scholar] [CrossRef] [PubMed]
- Huntington, J.A. How Na+ activates thrombin—A review of the functional and structural data. Biol. Chem. 2008, 389, 1025–1035. [Google Scholar] [CrossRef] [PubMed]
- Tsiang, M.; Jain, A.K.; Dunn, K.E.; Rojas, M.E.; Leung, L.L.K.; Gibbs, C.S. Functional Mapping of the Surface Residues of Human Thrombin. J. Biol. Chem. 1995, 270, 16854–16863. [Google Scholar] [CrossRef] [PubMed]
- Chahal, G.; Thorpe, M.; Hellman, L. The Importance of Exosite Interactions for Substrate Cleavage by Human Thrombin. PLoS ONE 2015, 10, e0129511. [Google Scholar] [CrossRef] [PubMed]
- Fitton, J.; Stringer, D.; Karpiniec, S. Therapies from Fucoidan: An Update. Mar. Drugs 2015, 13, 5920–5946. [Google Scholar] [CrossRef] [PubMed]
- Krichen, F.; Ghlissi, Z.; Amor, I.B.; Sayari, N.; Kallel, R.; Gargouri, J.; Sahnoun, Z.; Boudawara, T.; Bougatef, A. In vitro and in vivo anti-coagulant activity and toxicological studies of marine sulfated glycosaminoglycans. Exp. Toxicol. Pathol. 2017, 69, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Yeh, R.W.; Jang, I.-K. Argatroban: Update. Am. Heart J. 2006, 151, 1131–1138. [Google Scholar] [CrossRef] [PubMed]
- Warkentin, T.E.; Greinacher, A.; Koster, A. Bivalirudin. Thromb. Haemost. 2008, 99, 830–839. [Google Scholar] [CrossRef] [PubMed]
- Coppens, M.; Eikelboom, J.W.; Gustafsson, D.; Weitz, J.I.; Hirsh, J. Translational success stories: Development of direct thrombin inhibitors. Circ. Res. 2012, 111, 920–929. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, D.; Bylund, R.; Antonsson, T.; Nilsson, I.; Nyström, J.-E.; Eriksson, U.; Bredberg, U.; Teger-Nilsson, A.-C. Case history: A new oral anticoagulant: the 50-year challenge. Nat. Rev. Drug Discov. 2004, 3, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Behnes, M.; Fastner, C.; Ansari, U.; Akin, I. New Oral Anticoagulants in Coronary Artery Disease. Cardiovasc. Haematol. Disord. Drug Targets 2015, 15, 101–105. [Google Scholar] [CrossRef]
- Becattini, C.; Lignani, A.; Agnelli, G. New Oral Anticoagulants for Venous Thromboembolism: Focus on Factor Xa and Thrombin Inhibitors. Curr. Drug Discov. Technol. 2012, 9, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Mozzicafreddo, M.; Cuccioloni, M.; Eleuteri, A.M.; Fioretti, E.; Angeletti, M. Flavonoids inhibit the amidolytic activity of human thrombin. Biochimie 2006, 88, 1297–1306. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Gao, Y.-H.; Cao, J.; Zhang, H.-X.; Fan, L.; Hu, G.-L.; Hu, Y.-X.; Li, X.-L.; Zou, X.; Li, J.-H. High prevalence of aspirin resistance in elderly patients with cardiovascular disease and metabolic syndrome. J. Geriatr. Cardiol. 2016, 13, 531–536. [Google Scholar] [PubMed]
- Chen, J.; Wang, M.Z. Potent and Orally Bioavailable Antiplatelet Agent, PLD-301, with the Potential of Overcoming Clopidogrel Resistance. Lett. Drug Des. Discov. 2016, 13, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Cabral, K.; Ansell, J. The role of factor Xa inhibitors in venous thromboembolism treatment. Vasc. Health Risk Manag. 2015, 11, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.H.; Singh, K.P. Inhibition of Factor Xa: A potential target for the development of new anticoagulants. Am. J. Cardiovasc. Drugs Drugs Devices Interv. 2005, 5, 279–290. [Google Scholar] [CrossRef]
- Yorkgitis, B.K. Non-vitamin K antagonist oral anticoagulant reversal: Hope is on the horizon. Am. J. Surg. 2016, 212, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Desai, N.R.; Krumme, A.A.; Schneeweiss, S.; Shrank, W.H.; Brill, G.; Pezalla, E.J.; Spettell, C.M.; Brennan, T.A.; Matlin, O.S.; Avorn, J.; et al. Patterns of Initiation of Oral Anticoagulants in Patients with Atrial Fibrillation—Quality and Cost Implications. Am. J. Med. 2014, 127, 1075–1082. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Huntington, J.A. Crystal structures of protease nexin-1 in complex with heparin and thrombin suggest a 2-step recognition mechanism. Blood 2012, 120, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Daniel, K.G.; Chen, M.S.; Kuhn, D.J.; Landis-Piwowar, K.R.; Dou, Q.P. Dietary flavonoids as proteasome inhibitors and apoptosis inducers in human leukemia cells. Biochem. Pharmacol. 2005, 69, 1421–1432. [Google Scholar] [CrossRef] [PubMed]
- Belda, M.; Sanchez, D.; Bover, E.; Prieto, B.; Padrón, C.; Cejalvo, D.; Lloris, J.M. Extraction of polyphenols in Himanthaliaelongata and determination by high performance liquid chromatography with diode array detector prior to its potential use against oxidative stress. J. Chromatogr. B 2016, 1033–1034, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Ma, H.; Yang, N.; Tang, Y.; Guo, J.; Tao, W.; Duan, J. A series of natural flavonoids as thrombin inhibitors: Structure-activity relationships. Thromb. Res. 2010, 126, e365–e378. [Google Scholar] [CrossRef] [PubMed]
- Senthilvel, P.; Lavanya, P.; Kumar, K.M.; Swetha, R.; Anitha, P.; Bag, S.; Sarveswari, S.; Vijayakumar, V.; Ramaiah, S.; Anbarasu, A. Flavonoid from Carica papaya inhibits NS2B-NS3 protease and prevents Dengue 2 viral assembly. Bioinformation 2013, 9, 889–895. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Voet, A.; Zhang, K.Y.J. Fragment Based Drug Design: From Experimental to Computational Approaches. Curr. Med. Chem. 2012, 19, 5128–5147. [Google Scholar] [CrossRef] [PubMed]
- Baum, B.; Muley, L.; Heine, A.; Smolinski, M.; Hangauer, D.; Klebe, G. Think Twice: Understanding the High Potency of Bis(phenyl)methane Inhibitors of Thrombin. J. Mol. Biol. 2009, 391, 552–564. [Google Scholar] [CrossRef] [PubMed]
- David, W.B.; Paul, H. Crystallographic analysis at 3.0-A resolution of the binding to human thrombin of four active site-directed inhibitors. J. Biol. Chem. 1991, 266, 20085–20093. [Google Scholar]
- Hauel, N.H.; Nar, H.; Priepke, H.; Ries, U.; Stassen, J.-M.; Wienen, W. Structure-based design of novel potent nonpeptide thrombin inhibitors. J. Med. Chem. 2002, 45, 1757–1766. [Google Scholar] [CrossRef] [PubMed]
- Diderot, N.T.; Silvere, N.; Yasin, A.; Zareen, S.; Fabien, Z.; Etienne, T.; Choudhary, M.I. Atta-Ur-Rahman Prolylendopeptidase and thrombin inhibitory diterpenoids from the bark of Xylopiaaethiopica. Biosci. Biotechnol. Biochem. 2005, 69, 1763–1766. [Google Scholar] [CrossRef] [PubMed]

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, R.C.C.; Lourenço, A.L.; Terra, L.; Abreu, P.A.; Laneuville Teixeira, V.; Castro, H.C. Marine Diterpenes: Molecular Modeling of Thrombin Inhibitors with Potential Biotechnological Application as an Antithrombotic. Mar. Drugs 2017, 15, 79. https://doi.org/10.3390/md15030079
Pereira RCC, Lourenço AL, Terra L, Abreu PA, Laneuville Teixeira V, Castro HC. Marine Diterpenes: Molecular Modeling of Thrombin Inhibitors with Potential Biotechnological Application as an Antithrombotic. Marine Drugs. 2017; 15(3):79. https://doi.org/10.3390/md15030079
Chicago/Turabian StylePereira, Rebeca Cristina Costa, André Luiz Lourenço, Luciana Terra, Paula Alvarez Abreu, Valéria Laneuville Teixeira, and Helena Carla Castro. 2017. "Marine Diterpenes: Molecular Modeling of Thrombin Inhibitors with Potential Biotechnological Application as an Antithrombotic" Marine Drugs 15, no. 3: 79. https://doi.org/10.3390/md15030079