
Undariopsis peterseniana Promotes Hair Growth by the Activation of Wnt/β-Catenin and ERK Pathways

Abstract
:1. Introduction
2. Results
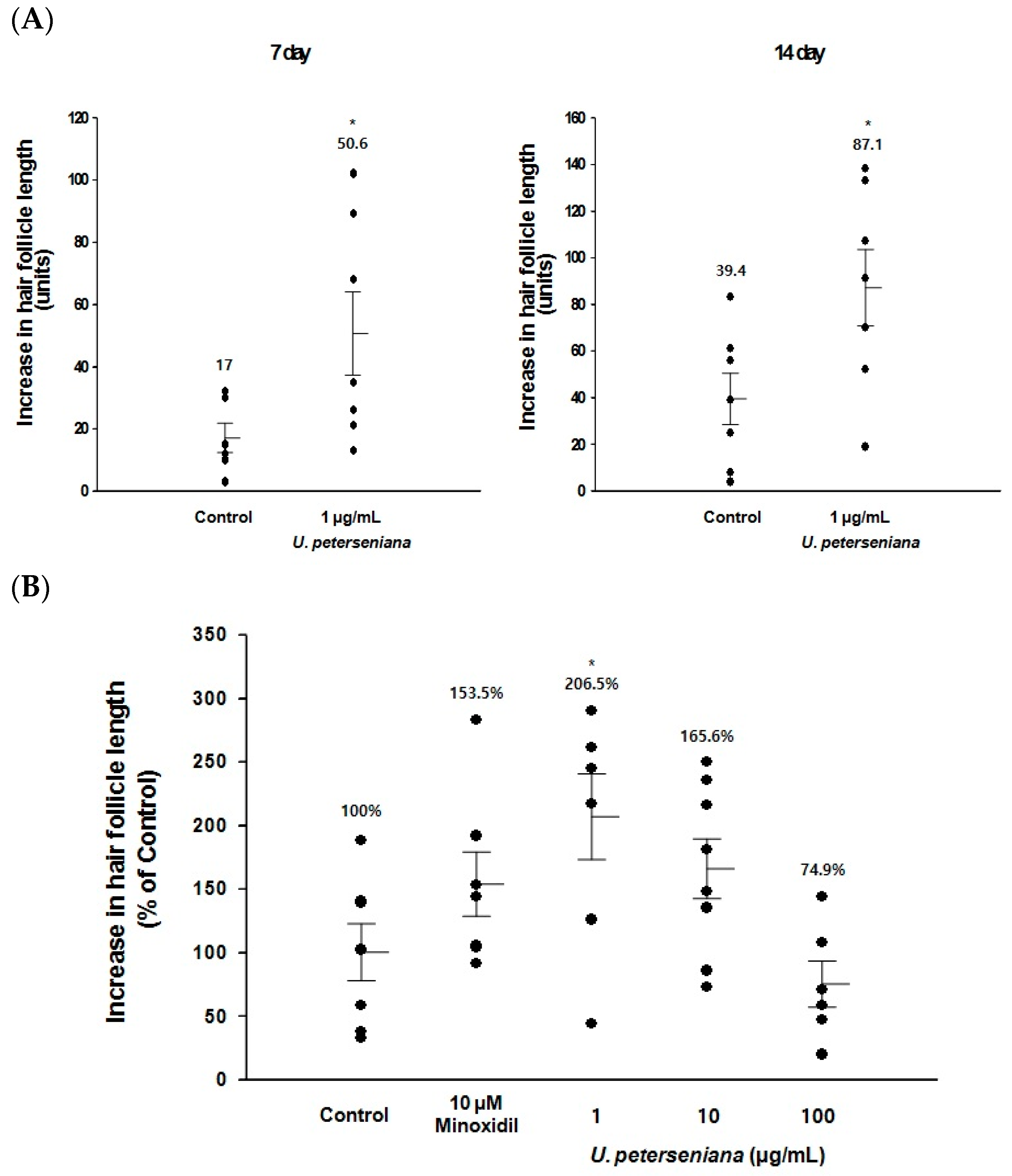
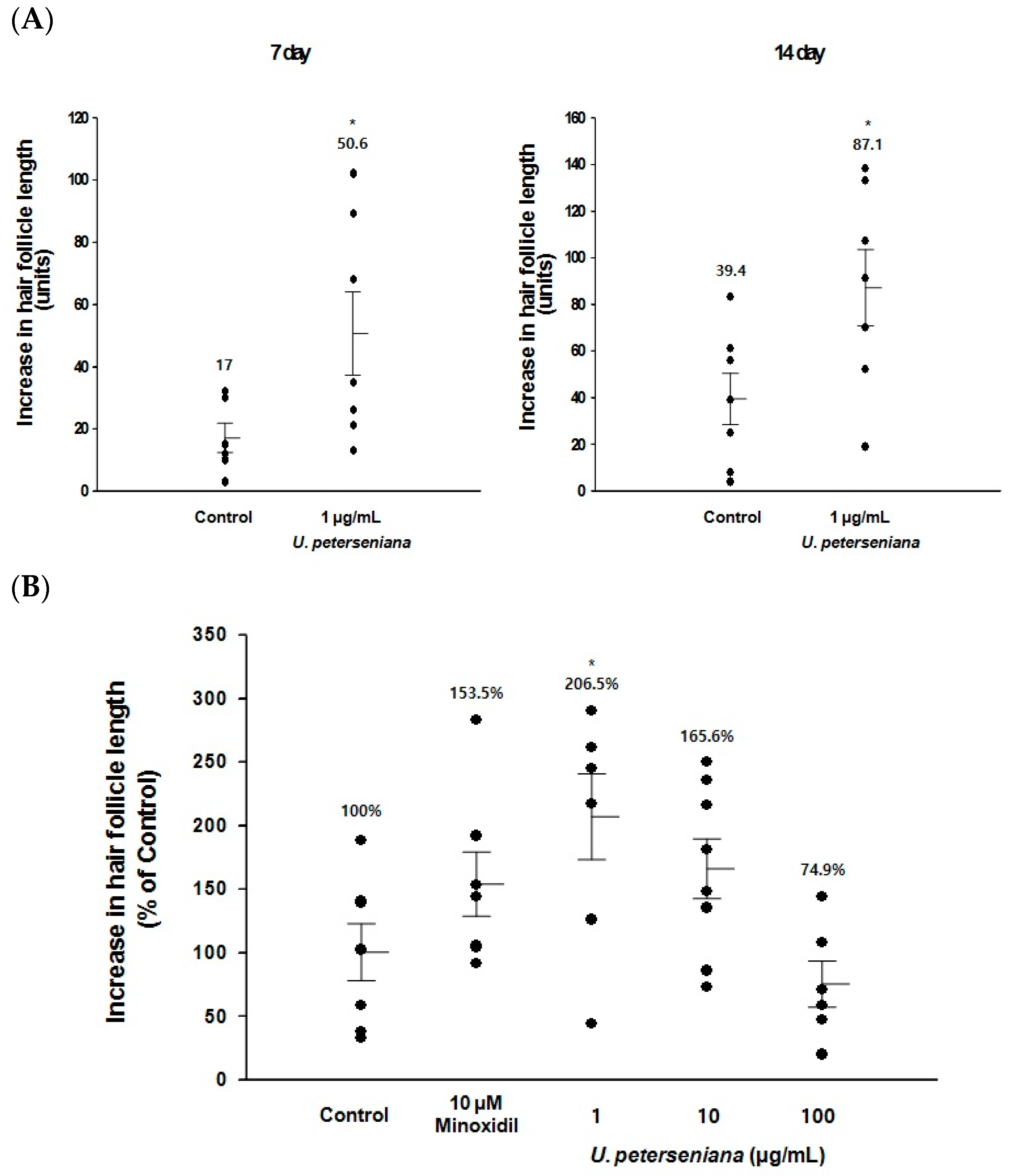
2.1. Ex Vivo Effect of U. peterseniana Extract on the Hair-Fiber Length of Rat Vibrissa Follicles
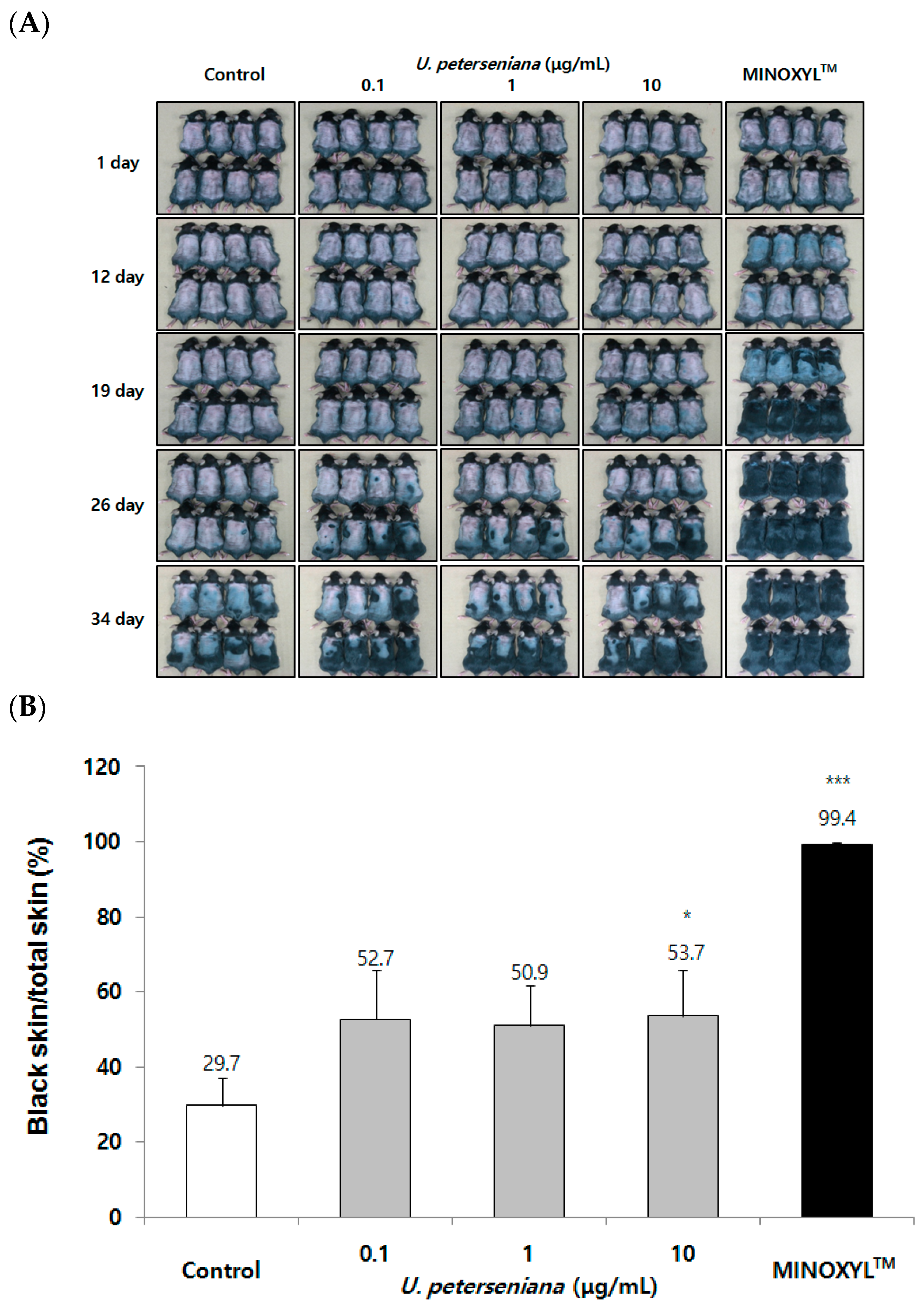
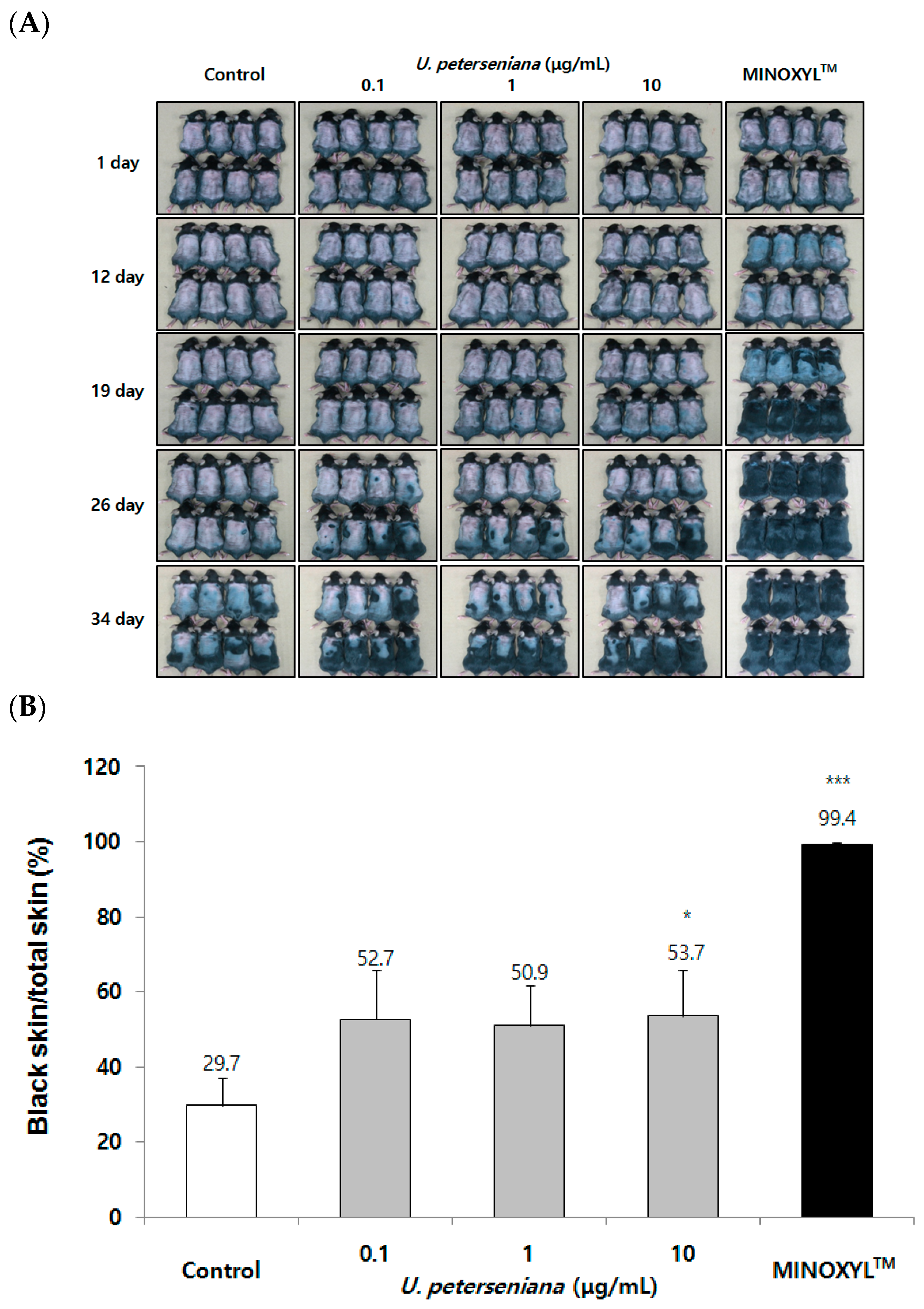
2.2. Effect of U. peterseniana Extract on the Telegen to Anagen Progression in C57BL/6 Mouse
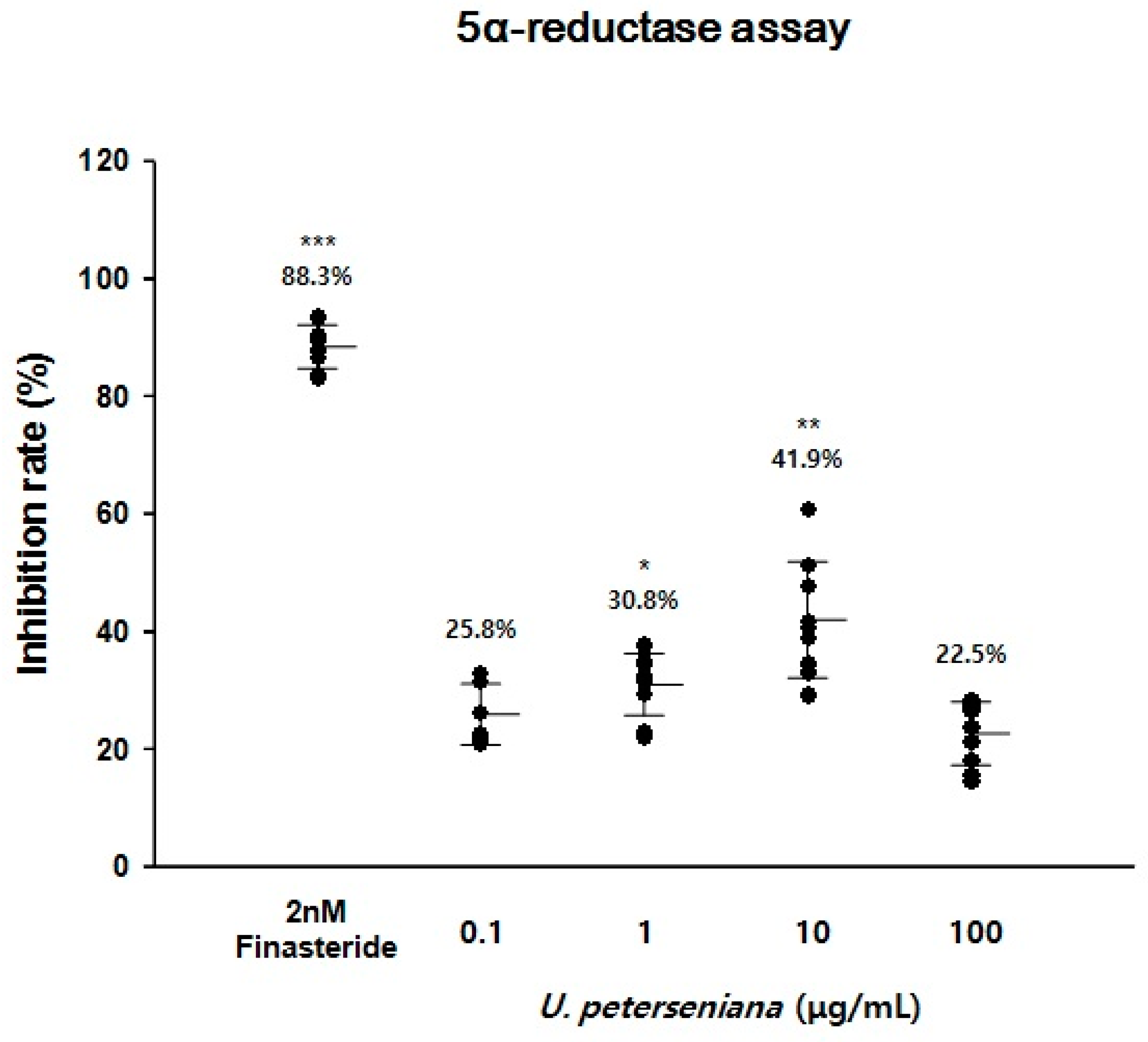
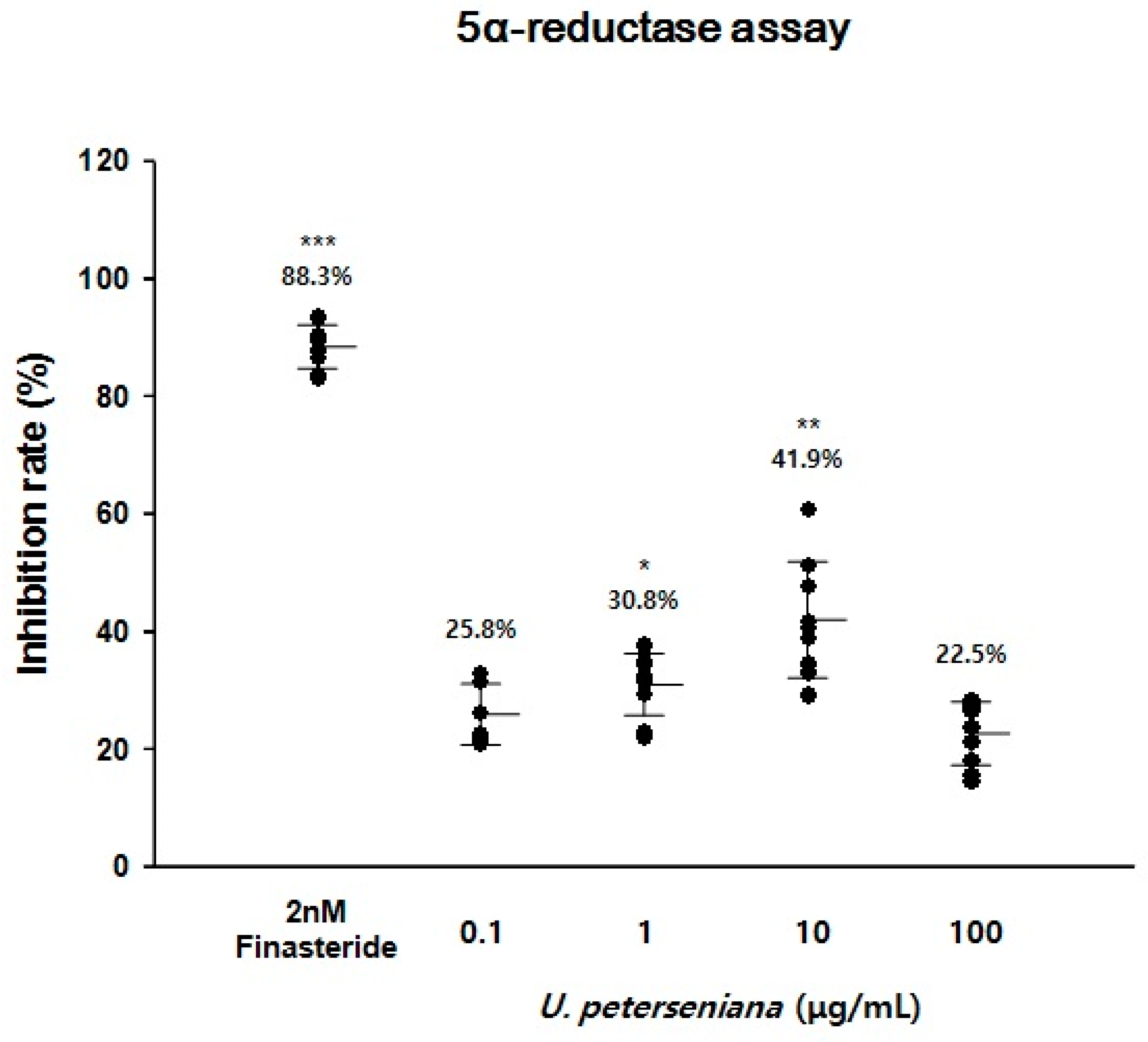
2.3. Effect of U. peterseniana Extract on the 5α-Reductase Activity
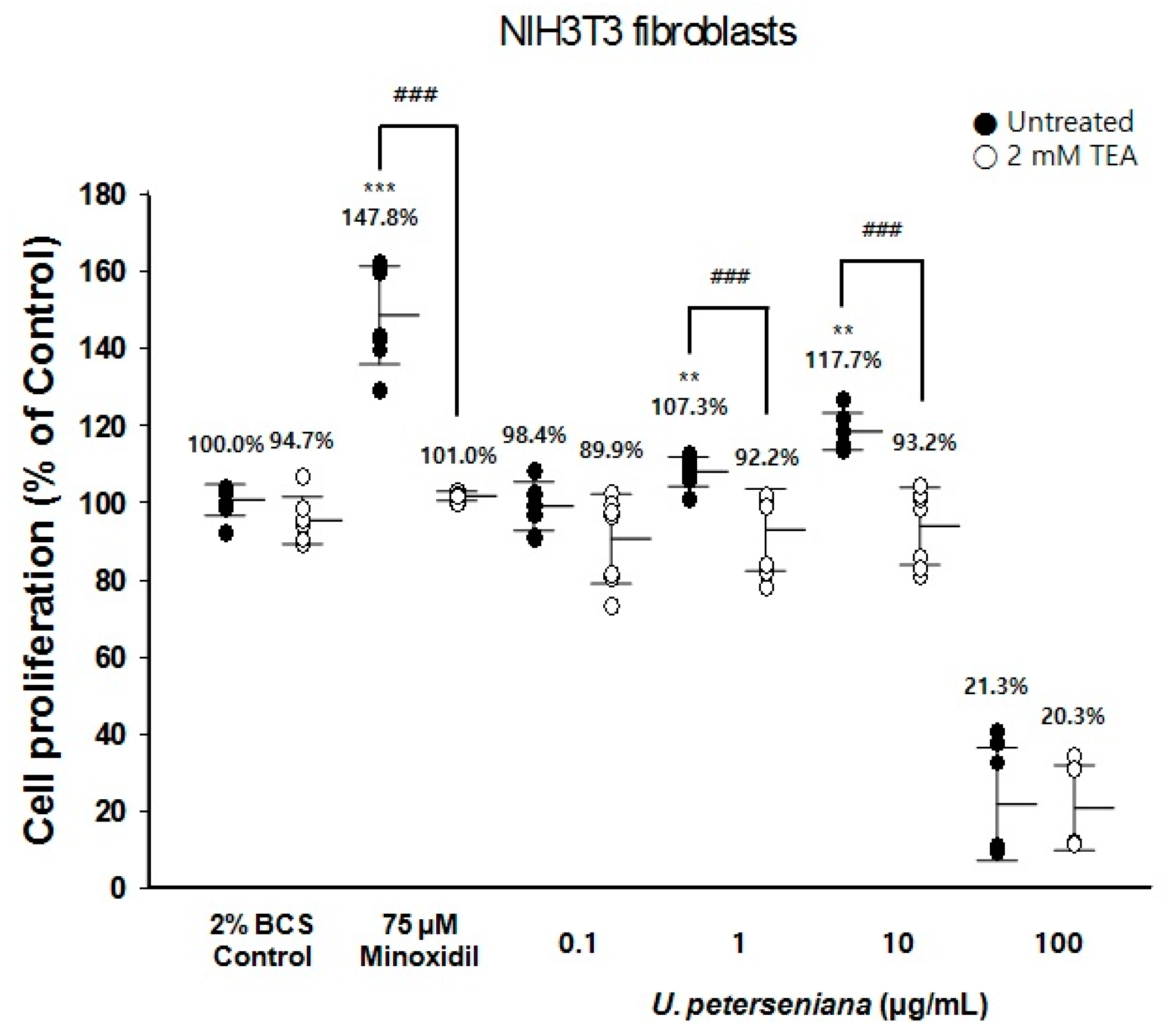
2.4. Effect of U. peterseniana Extract on the Proliferation of NIH3T3 Fibroblasts via KATP Channel Opening
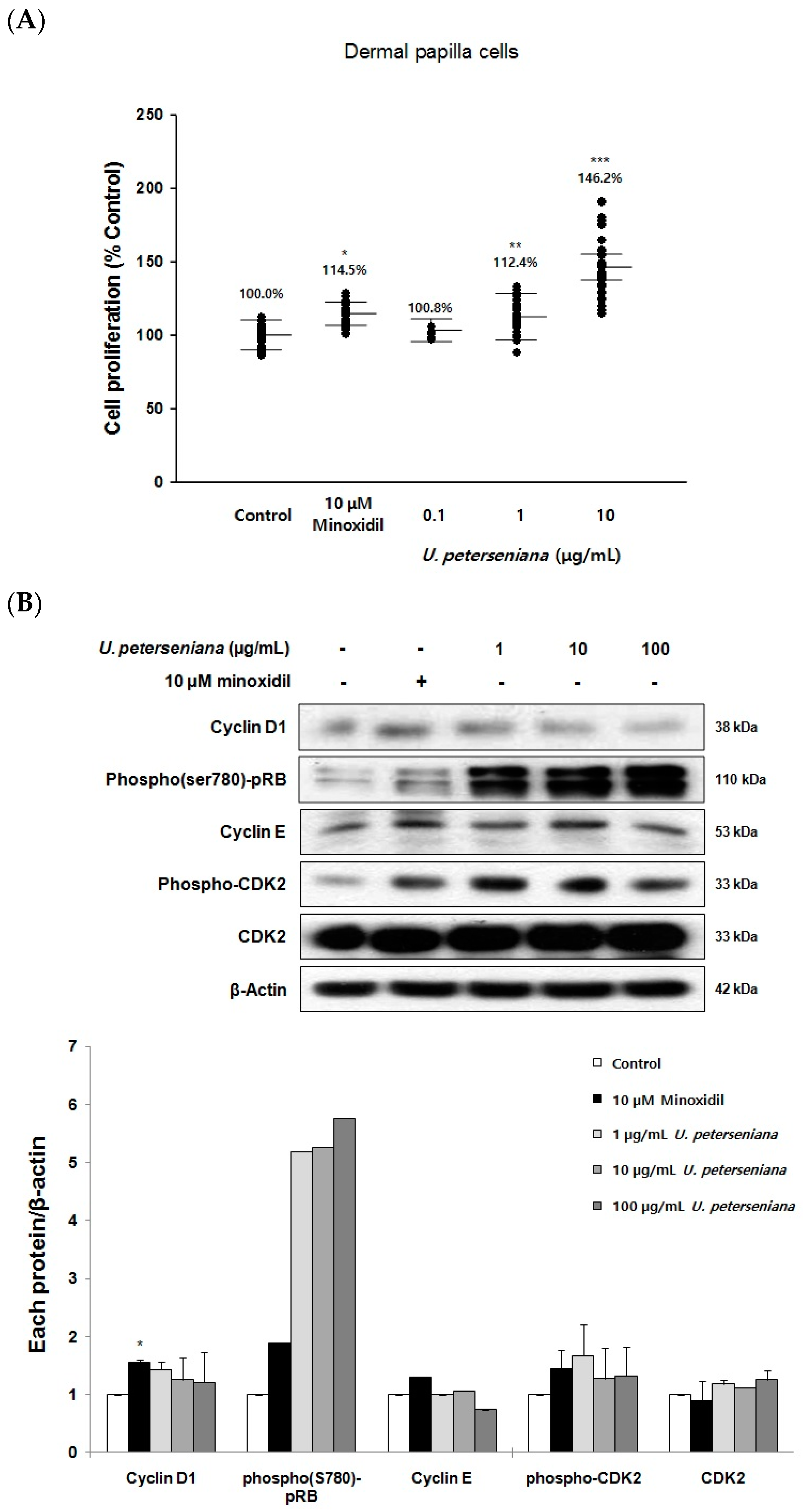
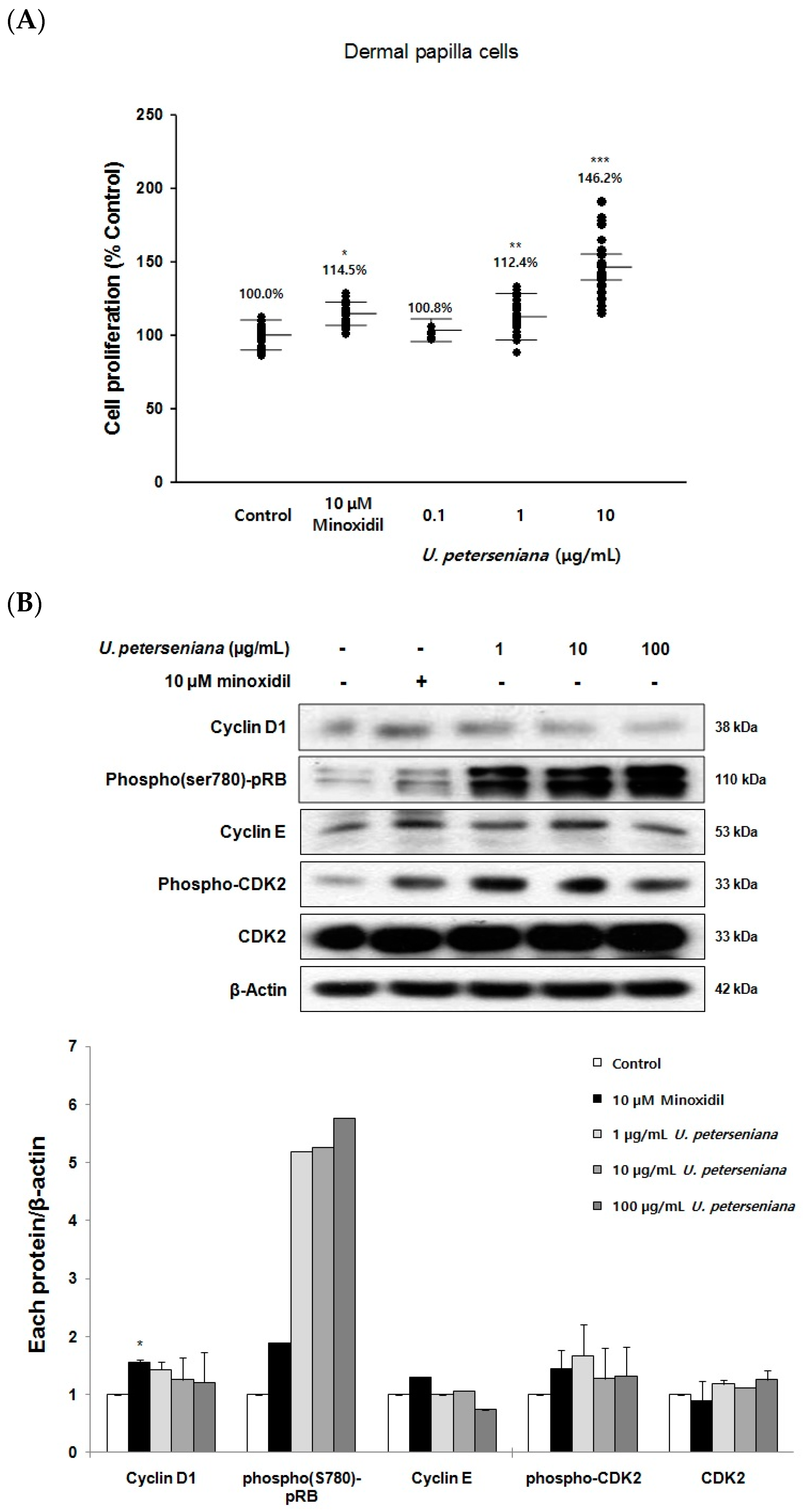
2.5. Effects of U. peterseniana Extract on the Proliferation of Dermal Papilla Cells
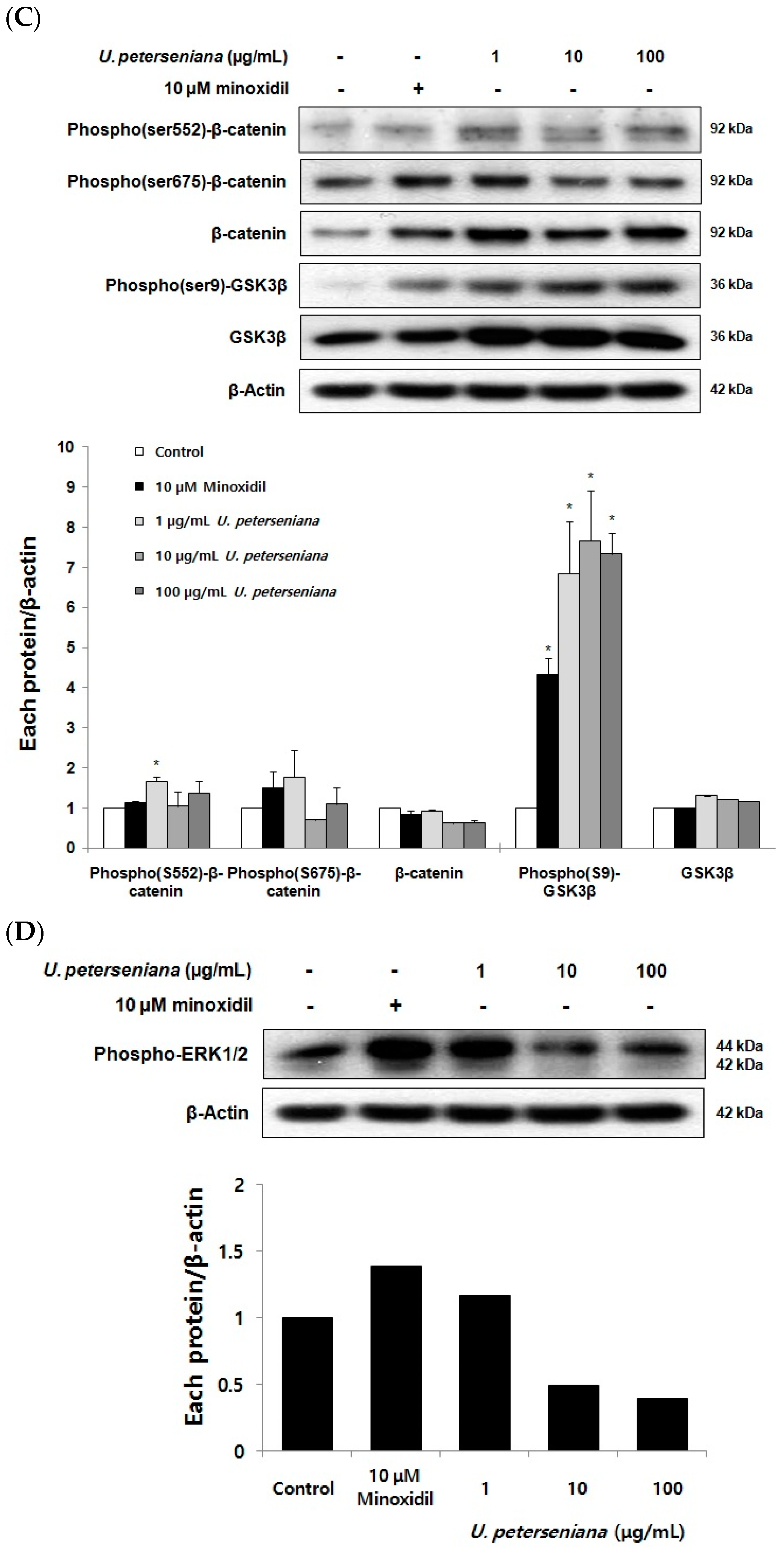
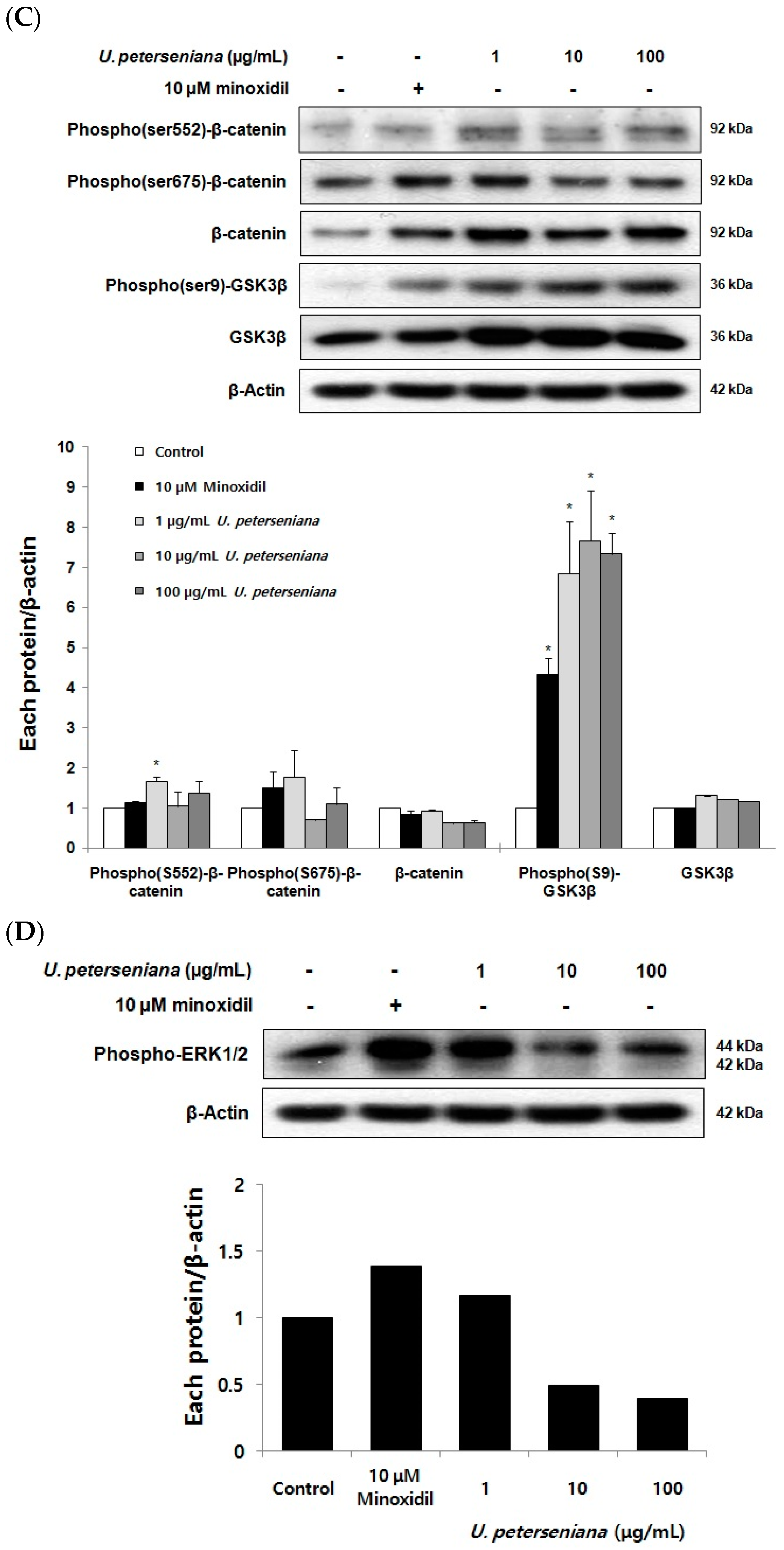
2.6. Effects of U. peterseniana Extract on the Levels of Cell Cycle Proteins, Wnt/β-Catenin Signaling, and Erk1/2 Signaling in Dermal Papilla Cells
3. Discussion
4. Materials and Methods
4.1. Reagents
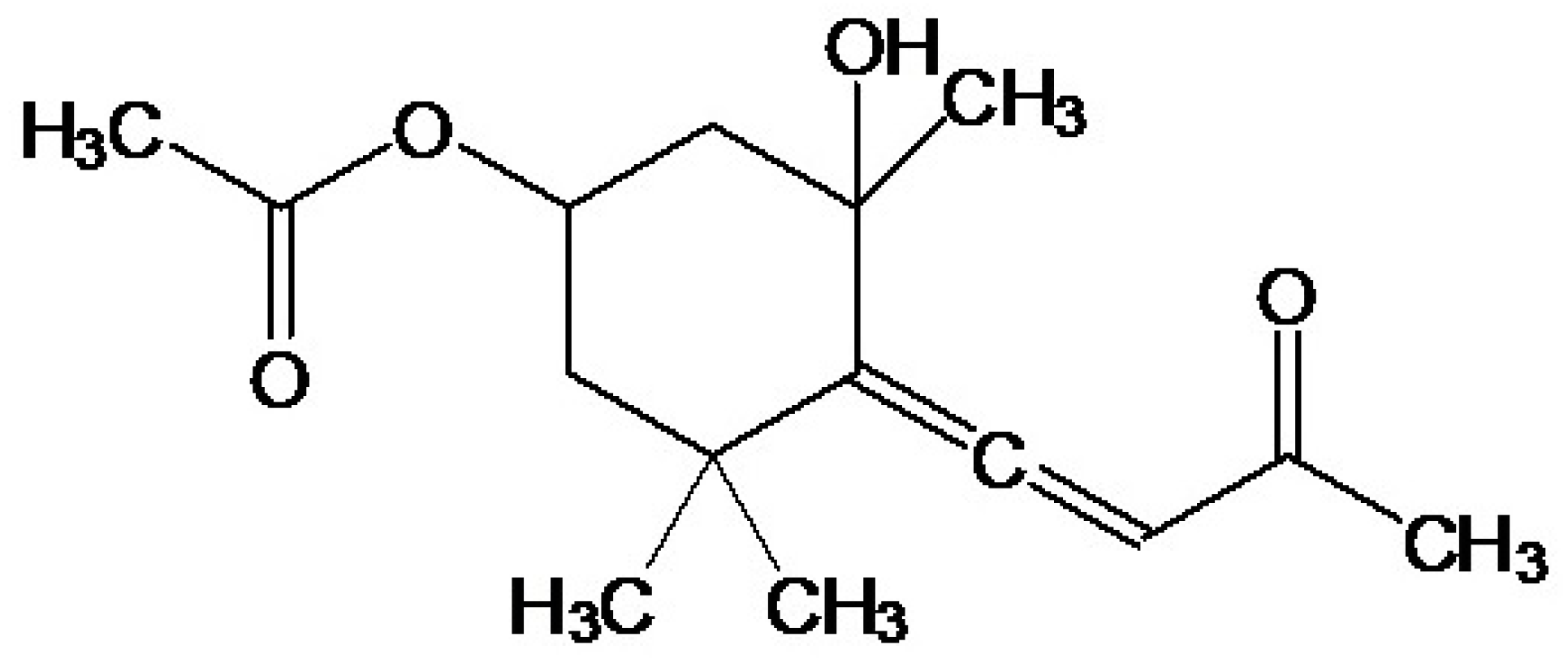
4.2. Preparation of Undariopsis peterseniana Extract
4.3. Animals
4.4. Isolation and Culture of Rat Vibrissa Follicles
4.5. Hair Growth Activity In Vivo
4.6. Assay for Prostatic 5α-Reductase Activity
4.7. Cell Culture
4.8. Proliferation Assay of NIH3T3 Fibroblasts
4.9. Proliferation Assay of Dermal Papilla Cells
4.10. Western Blot Analysis
4.11. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hunt, N.; McHale, S. The psychological impact of alopecia. BMJ 2005, 331, 951–953. [Google Scholar] [CrossRef] [PubMed]
- Daniells, S.; Hardy, G. Hair loss in long-term or home parenteral nutrition: Are micronutrient deficiencies to blame? Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 690–697. [Google Scholar] [CrossRef] [PubMed]
- Cotsarelis, G.; Millar, S.E. Towards a molecular understanding of hair loss and its treatment. Trends Mol. Med. 2001, 7, 293–301. [Google Scholar] [CrossRef]
- Burton, J.L.; Marshall, A. Hypertrichosis due to minoxidil. Br. J. Dermatol. 1979, 101, 593–595. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, K.D.; Olsen, E.A.; Whiting, D.; Savin, R.; DeVillez, R.; Bergfeld, W.; Price, V.H.; Van Neste, D.; Roberts, J.L.; Hordinsky, M.; et al. Finasteride in the treatment of men with androgenetic alopecia. Finasteride male pattern hair loss study group. J. Am. Acad. Dermatol. 1998, 39, 578–589. [Google Scholar] [CrossRef]
- Rossi, A.; Anzalone, A.; Fortuna, M.C.; Caro, G.; Garelli, V.; Pranteda, G.; Carlesimo, M. Multi-therapies in androgenetic alopecia: Review and clinical experiences. Dermatol. Ther. 2016, 29, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Tu, H.Y.; Zini, A. Finasteride-induced secondary infertility associated with sperm DNA damage. Fertil. Steril. 2011, 95, 2125.e13–2125.e14. [Google Scholar] [CrossRef] [PubMed]
- Muller-Rover, S.; Handjiski, B.; van der Veen, C.; Eichmuller, S.; Foitzik, K.; McKay, I.A.; Stenn, K.S.; Paus, R. A comprehensive guide for the accurate classification of murine hair follicles in distinct hair cycle stages. J. Investig. Dermatol. 2001, 117, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.B.; Zhang, Y.V.; Tumbar, T. Gata6 promotes hair follicle progenitor cell renewal by genome maintenance during proliferation. EMBO J. 2017, 36, 61–78. [Google Scholar] [CrossRef] [PubMed]
- Greco, V.; Chen, T.; Rendl, M.; Schober, M.; Pasolli, H.A.; Stokes, N.; Dela Cruz-Racelis, J.; Fuchs, E. A two-step mechanism for stem cell activation during hair regeneration. Cell Stem Cell 2009, 4, 155–169. [Google Scholar] [CrossRef] [PubMed]
- Kwack, M.H.; Kang, B.M.; Kim, M.K.; Kim, J.C.; Sung, Y.K. Minoxidil activates beta-catenin pathway in human dermal papilla cells: A possible explanation for its anagen prolongation effect. J. Dermatol. Sci. 2011, 62, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Dastan, M.; Najafzadeh, N.; Abedelahi, A.; Sarvi, M.; Niapour, A. Human platelet lysate versus minoxidil stimulates hair growth by activating anagen promoting signaling pathways. Biomed. Pharmacother. 2016, 84, 979–986. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.I.; Kim, S.C.; Hyun, J.H.; Kang, J.H.; Park, D.B.; Lee, Y.J.; Yoo, E.S.; Kang, H.K. Promotion effect of Schisandra nigra on the growth of hair. Eur. J. Dermatol. 2009, 19, 119–125. [Google Scholar] [PubMed]
- Ouji, Y.; Yoshikawa, M.; Moriya, K.; Nishiofuku, M.; Matsuda, R.; Ishizaka, S. Wnt-10b, uniquely among Wnts, promotes epithelial differentiation and shaft growth. Biochem. Biophys. Res. Commun. 2008, 367, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Huelsken, J.; Vogel, R.; Erdmann, B.; Cotsarelis, G.; Birchmeier, W. β-catenin controls hair follicle morphogenesis and stem cell differentiation in the skin. Cell 2001, 105, 533–545. [Google Scholar] [CrossRef]
- Molavi, O.; Narimani, F.; Asiaee, F.; Sharifi, S.; Tarhriz, V.; Shayanfar, A.; Hejazi, M.; Lai, R. Silibinin sensitizes chemo-resistant breast cancer cells to chemotherapy. Pharm. Biol. 2017, 55, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Gunda, V.; Sarosiek, K.A.; Brauner, E.; Kim, Y.S.; Amin, S.; Zhou, Z.; Letai, A.; Parangi, S. Inhibition of MAPKinase pathway sensitizes thyroid cancer cells to ABT-737 induced apoptosis. Cancer Lett. 2017, 395, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Man, X.Y.; Li, C.M.; Chen, J.Q.; Zhou, J.; Cai, S.Q.; Lu, Z.F.; Zheng, M. VEGF induces proliferation of human hair follicle dermal papilla cells through VEGFR-2-mediated activation of ERK. Exp. Cell Res. 2012, 318, 1633–1640. [Google Scholar] [CrossRef] [PubMed]
- Hwang, K.A.; Hwang, Y.L.; Lee, M.H.; Kim, N.R.; Roh, S.S.; Lee, Y.; Kim, C.D.; Lee, J.H.; Choi, K.C. Adenosine stimulates growth of dermal papilla and lengthens the anagen phase by increasing the cysteine level via fibroblast growth factors 2 and 7 in an organ culture of mouse vibrissae hair follicles. Int. J. Mol. Med. 2012, 29, 195–201. [Google Scholar] [PubMed]
- Hwang, E.K.; Gong, Y.G.; Park, C.S. Cultivation of a brown alga, Undariopsis peterseniana (Kjellman) Miyabe and Okamura, as a warm-temperature species by artificial seed production in Korea. J. Appl. Phycol. 2011, 23, 449–455. [Google Scholar] [CrossRef]
- Hwang, E.K.; Gong, Y.G.; Park, C.S. Cultivation of a hybrid of free-living gametophytes between Undariopsis peterseniana and Undaria pinnatifida: Morphological aspects and cultivation period. J. Appl. Phycol. 2012, 24, 401–408. [Google Scholar] [CrossRef]
- Cho, M.; Yoon, S.J.; Kim, Y.-B. The nutritional composition and antioxidant activity from Undariopsis peterseniana. Ocean Polar Res. 2013, 35, 273–280. [Google Scholar] [CrossRef]
- Philpott, M.P.; Kealey, T. Cyclical changes in rat vibrissa follicles maintained in vitro. J. Investig. Dermatol. 2000, 115, 1152–1155. [Google Scholar] [CrossRef] [PubMed]
- Ohnemus, U.; Uenalan, M.; Conrad, F.; Handjiski, B.; Mecklenburg, L.; Nakamura, M.; Inzunza, J.; Gustafsson, J.A.; Paus, R. Hair cycle control by estrogens: Catagen induction via estrogen receptor (ER)-α is checked by ERβ signaling. Endocrinology 2005, 146, 1214–1225. [Google Scholar] [CrossRef] [PubMed]
- Sawaya, M.E.; Price, V.H. Different levels of 5α-reductase type I and II, aromatase, and androgen receptor in hair follicles of women and men with androgenetic alopecia. J. Investig. Dermatol. 1997, 109, 296–300. [Google Scholar] [CrossRef] [PubMed]
- Sanders, D.A.; Fiddes, I.; Thompson, D.M.; Philpott, M.P.; Westgate, G.E.; Kealey, T. In the absence of streptomycin, minoxidil potentiates the mitogenic effects of fetal calf serum, insulin-like growth factor 1, and platelet-derived growth factor on NIH 3T3 fibroblasts in a K+ channel-dependent fashion. J. Investig. Dermatol. 1996, 107, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, S.R.; Mallinger, A.; Workman, P.; Clarke, P.A. Inhibitors of cyclin-dependent kinases as cancer therapeutics. Pharmacol. Ther. 2017, 173, 83–105. [Google Scholar] [CrossRef] [PubMed]
- Leirós, G.J.; Ceruti, J.M.; Castellanos, M.L.; Kusinsky, A.G.; Balañá, M.E. Androgens modify Wnt agonists/antagonists expression balance in dermal papilla cells preventing hair follicle stem cell differentiation in androgenetic alopecia. Mol. Cell. Endocrinol. 2017, 439, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Whelchel, A.; Evans, J.; Posada, J. Inhibition of ERK activation attenuates endothelin-stimulated airway smooth muscle cell proliferation. Am. J. Respir. Cell Mol. Biol. 1997, 16, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Dallob, A.L.; Sadick, N.S.; Unger, W.; Lipert, S.; Geissler, L.A.; Gregoire, S.L.; Nguyen, H.H.; Moore, E.C.; Tanaka, W.K. The effect of finasteride, a 5 α-reductase inhibitor, on scalp skin testosterone and dihydrotestosterone concentrations in patients with male pattern baldness. J. Clin. Endocrinol. Metab. 1994, 79, 703–706. [Google Scholar] [PubMed]
- Lee, J.-H.; Ko, J.-Y.; Kim, E.-A.; Hwang, E.-K.; Park, C.S.; Lee, J.-S.; Kim, C.-Y.; Lee, H.-S.; Kang, H.-K.; Cha, S.-H. Identification and large isolation of an anti-inflammatory compound from an edible brown seaweed, Undariopsis peterseniana, and evaluation on its anti-inflammatory effect in in vitro and in vivo zebrafish. J. Appl. Phycol. 2016, 1–10. [Google Scholar] [CrossRef]
- Kang, J.I.; Yoo, E.S.; Hyun, J.W.; Koh, Y.S.; Lee, N.H.; Ko, M.H.; Ko, C.S.; Kang, H.K. Promotion effect of apo-9′-fucoxanthinone from sargassum muticum on hair growth via the activation of Wnt/β-catenin and VEGF-R2. Biol. Pharm. Bull. 2016, 39, 1273–1283. [Google Scholar] [CrossRef] [PubMed]
- Shorter, K.; Farjo, N.P.; Picksley, S.M.; Randall, V.A. Human hair follicles contain two forms of ATP-sensitive potassium channels, only one of which is sensitive to minoxidil. FASEB J. 2008, 22, 1725–1736. [Google Scholar] [CrossRef] [PubMed]
- Buhl, A.E.; Waldon, D.J.; Kawabe, T.T.; Holland, J.M. Minoxidil stimulates mouse vibrissae follicles in organ culture. J. Investig. Dermatol. 1989, 92, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Elliott, K.; Stephenson, T.J.; Messenger, A.G. Differences in hair follicle dermal papilla volume are due to extracellular matrix volume and cell number: Implications for the control of hair follicle size and androgen responses. J. Investig. Dermatol. 1999, 113, 873–877. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.I.; Kim, E.J.; Kim, M.K.; Jeon, Y.J.; Kang, S.M.; Koh, Y.S.; Yoo, E.S.; Kang, H.K. The promoting effect of ishige sinicola on hair growth. Mar. Drugs 2013, 11, 1783–1799. [Google Scholar] [CrossRef] [PubMed]
- Nunez, M.; Medina, V.; Cricco, G.; Croci, M.; Cocca, C.; Rivera, E.; Bergoc, R.; Martin, G. Glibenclamide inhibits cell growth by inducing G0/G1 arrest in the human breast cancer cell line MDA-MB-231. BMC Pharmacol. Toxicol. 2013, 14, 6. [Google Scholar] [CrossRef] [PubMed]
- Lien, W.H.; Polak, L.; Lin, M.; Lay, K.; Zheng, D.; Fuchs, E. In vivo transcriptional governance of hair follicle stem cells by canonical Wnt regulators. Nat. Cell Biol. 2014, 16, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Polakis, P. Wnt signaling in cancer. Cold Spring Harb. Perspect. Biol. 2012, 4, a008052. [Google Scholar] [CrossRef] [PubMed]
- Hedgepeth, C.M.; Conrad, L.J.; Zhang, J.; Huang, H.C.; Lee, V.M.; Klein, P.S. Activation of the Wnt signaling pathway: A molecular mechanism for lithium action. Dev. Biol. 1997, 185, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Lin, M.; Bu, Y.; Ling, H.; He, Y.; Huang, C.; Shen, Y.; Song, B.; Cao, D. p53-inducible long non-coding RNA PICART1 mediates cancer cell proliferation and migration. Int. J. Oncol. 2017, 50, 1671–1682. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.S.; Park, S.Y.; Hwang, E.S.; Lee, D.G.; Song, H.G.; Mavlonov, G.T.; Yi, T.H. The inductive effect of ginsenoside F2 on hair growth by altering the Wnt signal pathway in telogen mouse skin. Eur. J. Pharmacol. 2014, 730, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Park, P.J.; Moon, B.S.; Lee, S.H.; Kim, S.N.; Kim, A.R.; Kim, H.J.; Park, W.S.; Choi, K.Y.; Cho, E.G.; Lee, T.R. Hair growth-promoting effect of aconiti ciliare tuber extract mediated by the activation of Wnt/beta-catenin signaling. Life Sci. 2012, 91, 935–943. [Google Scholar] [CrossRef] [PubMed]
- Hino, S.; Tanji, C.; Nakayama, K.I.; Kikuchi, A. Phosphorylation of β-catenin by cyclic amp-dependent protein kinase stabilizes β-catenin through inhibition of its ubiquitination. Mol. Cell. Biol. 2005, 25, 9063–9072. [Google Scholar] [CrossRef] [PubMed]
- Monick, M.M.; Carter, A.B.; Robeff, P.K.; Flaherty, D.M.; Peterson, M.W.; Hunninghake, G.W. Lipopolysaccharide activates Akt in human alveolar macrophages resulting in nuclear accumulation and transcriptional activity of β-catenin. J. Immunol. 2001, 166, 4713–4720. [Google Scholar] [CrossRef] [PubMed]
- Kwon, O.; Han, J.; Yoo, H.; Chung, J.; Cho, K.; Eun, H.; Kim, K. Human hair growth enhancement in vitro by green tea epigallocatechin-3-gallate (EGCG). Phytomedicine 2007, 14, 551–555. [Google Scholar] [CrossRef] [PubMed]
- Lavoie, J.N.; L’Allemain, G.; Brunet, A.; Müller, R.; Pouysségur, J. Cyclin D1 expression is regulated positively by the p42/p44MAPK and negatively by the p38/HOGMAPK pathway. J. Biol. Chem. 1996, 271, 20608–20616. [Google Scholar] [CrossRef] [PubMed]
- Picard, F.; Schulz, T.; Hartmann, R.W. 5-phenyl substituted 1-methyl-2-pyridones and 4′-substituted biphenyl-4-carboxylic acids. Synthesis and evaluation as inhibitors of steroid-5α-reductase type 1 and 2. Biorg. Med. Chem. 2002, 10, 437–448. [Google Scholar] [CrossRef]
- Filsell, W.; Little, J.C.; Stones, A.J.; Granger, S.P.; Bayley, S.A. Transfection of rat dermal papilla cells with a gene encoding a temperature-sensitive polyomavirus large T antigen generates cell lines retaining a differentiated phenotype. J. Cell Sci. 1994, 107, 1761–1772. [Google Scholar] [PubMed]
- Carmichael, J.; DeGraff, W.G.; Gazdar, A.F.; Minna, J.D.; Mitchell, J.B. Evaluation of a tetrazolium-based semiautomated colorimetric assay: Assessment of chemosensitivity testing. Cancer Res. 1987, 47, 936–942. [Google Scholar] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Image, J. Image Processing and Analysis in Java. Available online: https://imagej.nih.gov/ij/ (accessed on 10 February 2017).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibodies | Supplier | Species | dilution |
|---|---|---|---|
| phospho-(ser552)-β-catenin | Cell Signaling | Rabbit | 1:1000 |
| phospho-(ser675)-β-catenin | Cell Signaling | Rabbit | 1:1000 |
| β-catenin | Santa Cruz | Rabbit | 1:2000 |
| phospho(ser9)-GSK-3β | Cell Signaling | Rabbit | 1:1000 |
| GSK-3β | Cell Signaling | Rabbit | 1:1000 |
| phospho-Erk1/2 | Cell Signaling | Rabbit | 1:1000 |
| Cyclin D1 | BD Biosciences | Mouse | 1:1000 |
| phospho(ser780)-pRB | Cell Signaling | Rabbit | 1:1000 |
| Cyclin E | Santa Cruz | Rabbit | 1:1000 |
| phospho-CDK2 | Cell Signaling | Rabbit | 1:1000 |
| CDK2 | Santa Cruz | Rabbit | 1:1000 |
| β-actin | Sigma-Aldrich | Mouse | 1:5000 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, J.-I.; Kim, M.-K.; Lee, J.-H.; Jeon, Y.-J.; Hwang, E.-K.; Koh, Y.-S.; Hyun, J.-W.; Kwon, S.-Y.; Yoo, E.-S.; Kang, H.-K. Undariopsis peterseniana Promotes Hair Growth by the Activation of Wnt/β-Catenin and ERK Pathways. Mar. Drugs 2017, 15, 130. https://doi.org/10.3390/md15050130
Kang J-I, Kim M-K, Lee J-H, Jeon Y-J, Hwang E-K, Koh Y-S, Hyun J-W, Kwon S-Y, Yoo E-S, Kang H-K. Undariopsis peterseniana Promotes Hair Growth by the Activation of Wnt/β-Catenin and ERK Pathways. Marine Drugs. 2017; 15(5):130. https://doi.org/10.3390/md15050130
Chicago/Turabian StyleKang, Jung-Il, Min-Kyoung Kim, Ji-Hyeok Lee, You-Jin Jeon, Eun-Kyoung Hwang, Young-Sang Koh, Jin-Won Hyun, Soon-Young Kwon, Eun-Sook Yoo, and Hee-Kyoung Kang. 2017. "Undariopsis peterseniana Promotes Hair Growth by the Activation of Wnt/β-Catenin and ERK Pathways" Marine Drugs 15, no. 5: 130. https://doi.org/10.3390/md15050130