The Roles of Spinochromes in Four Shallow Water Tropical Sea Urchins and Their Potential as Bioactive Pharmacological Agents

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
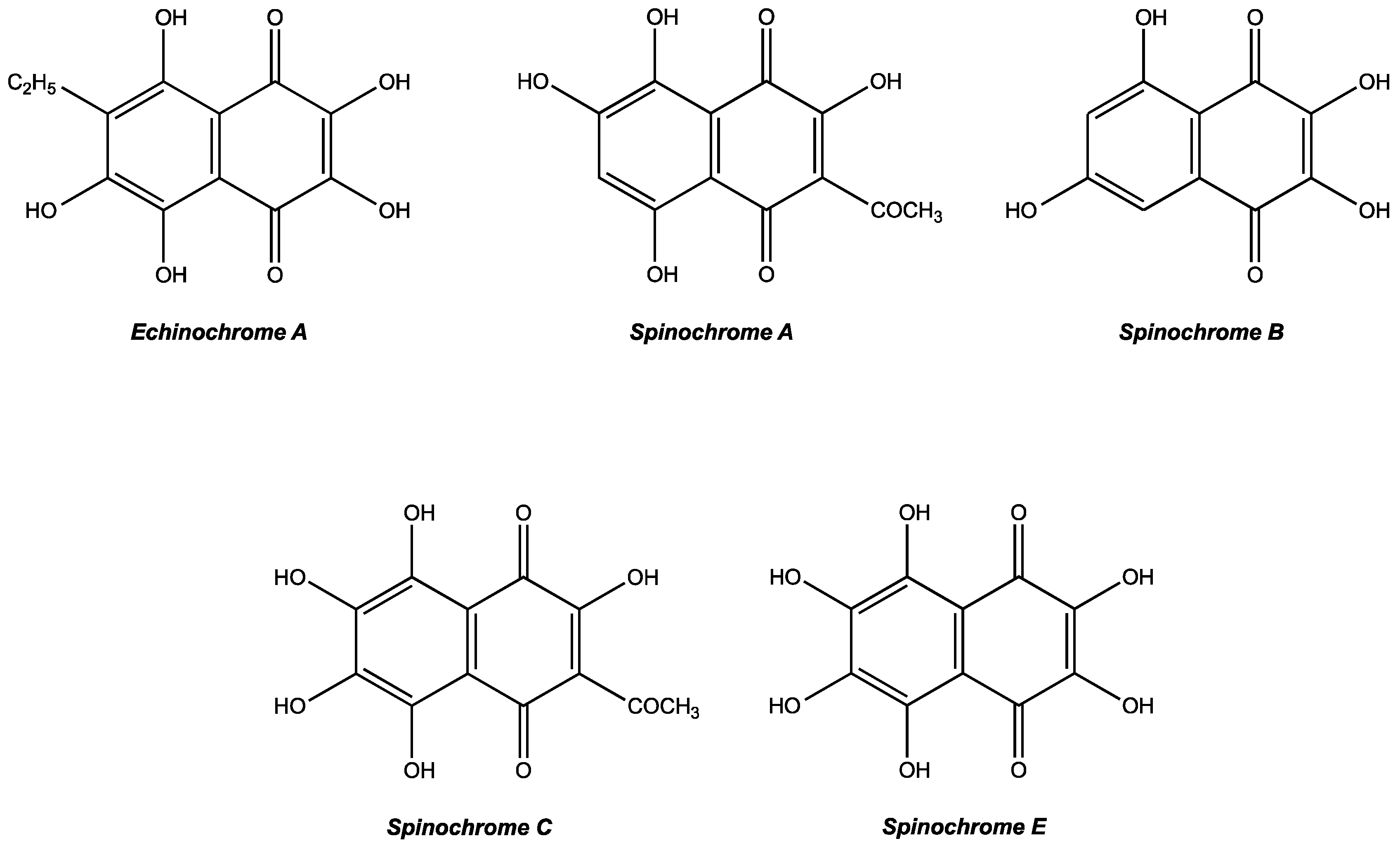
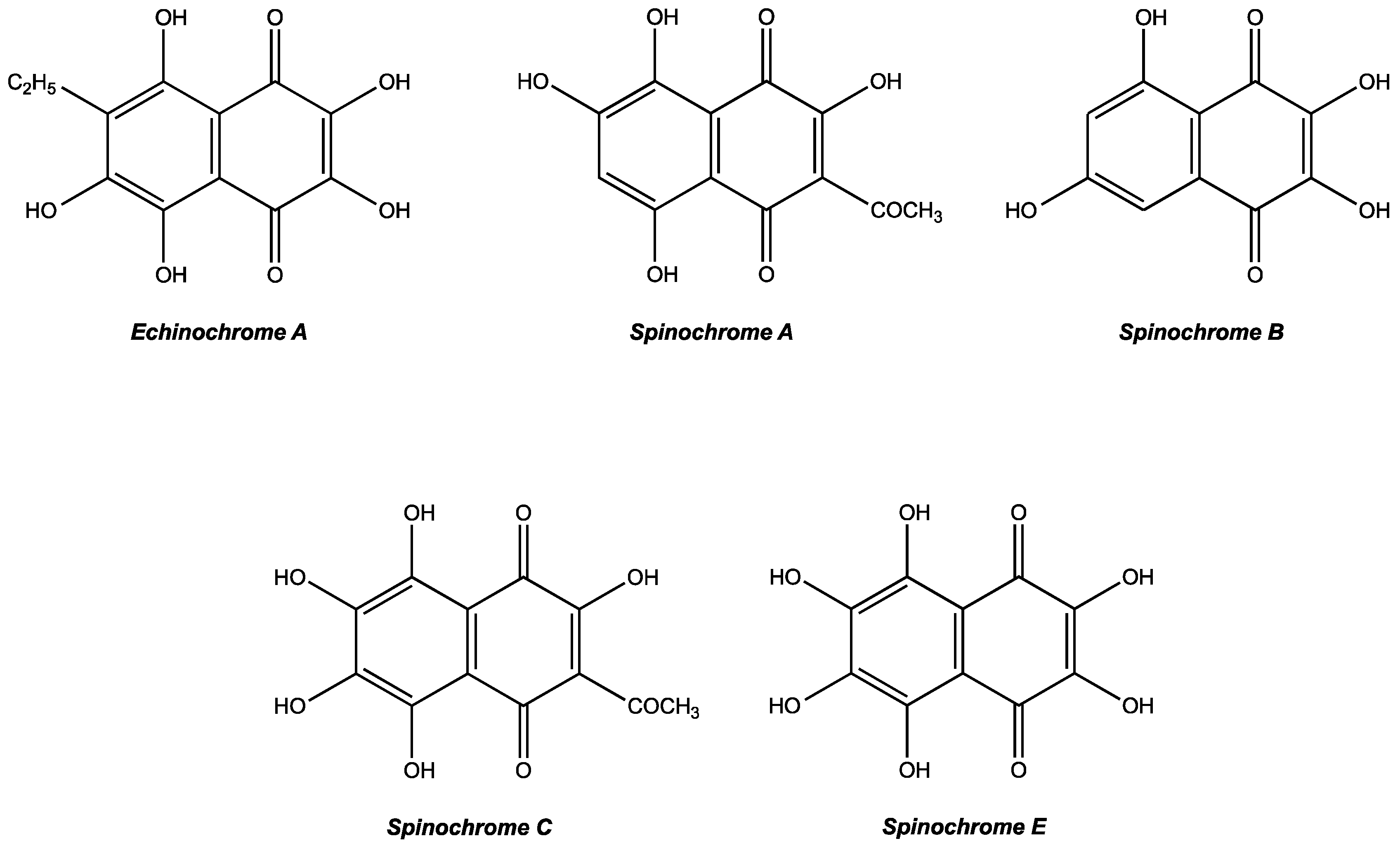
2.1. Extraction and Isolation of Spinochromes
2.2. Bioactive Effects of Spinochromes
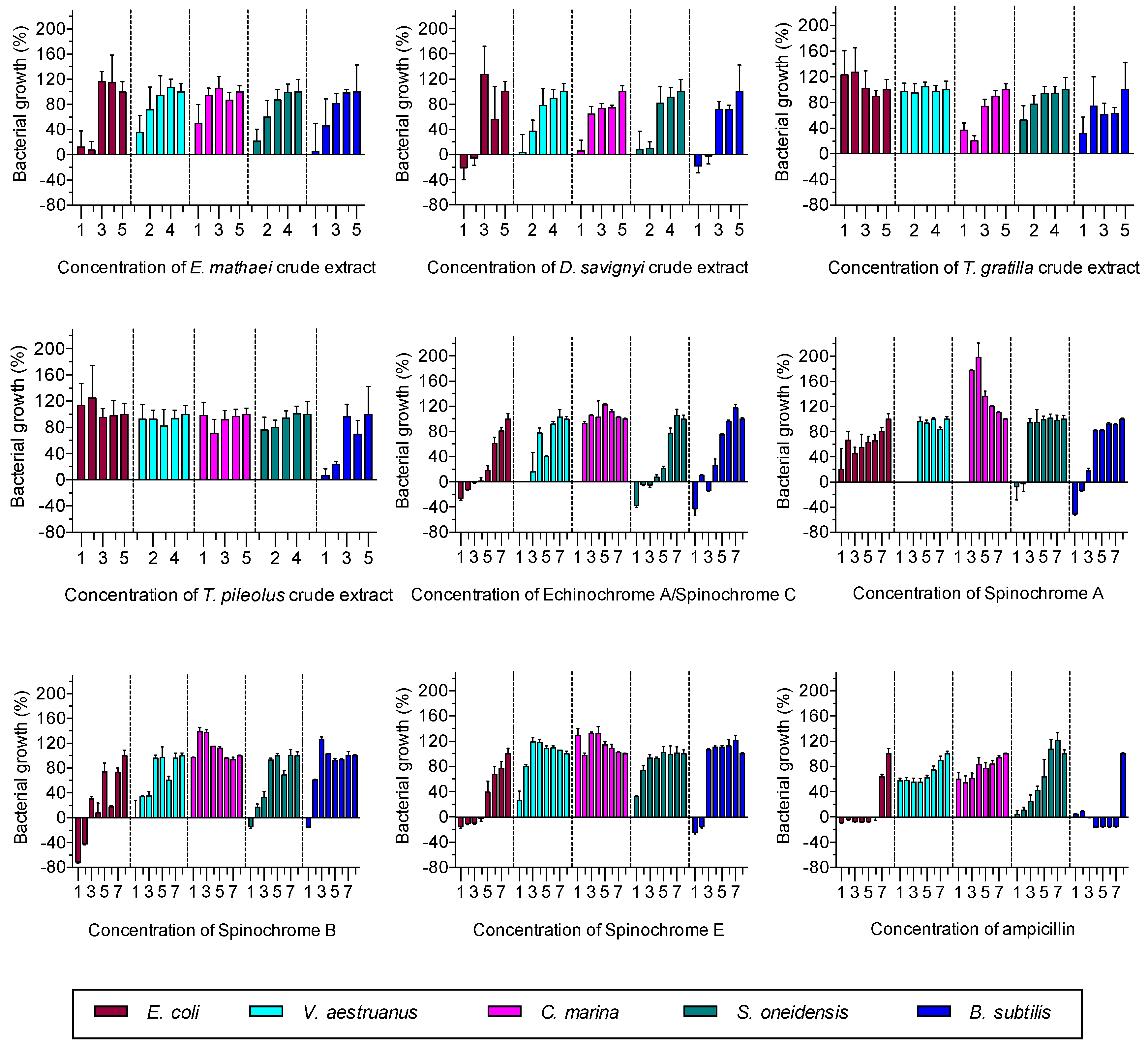
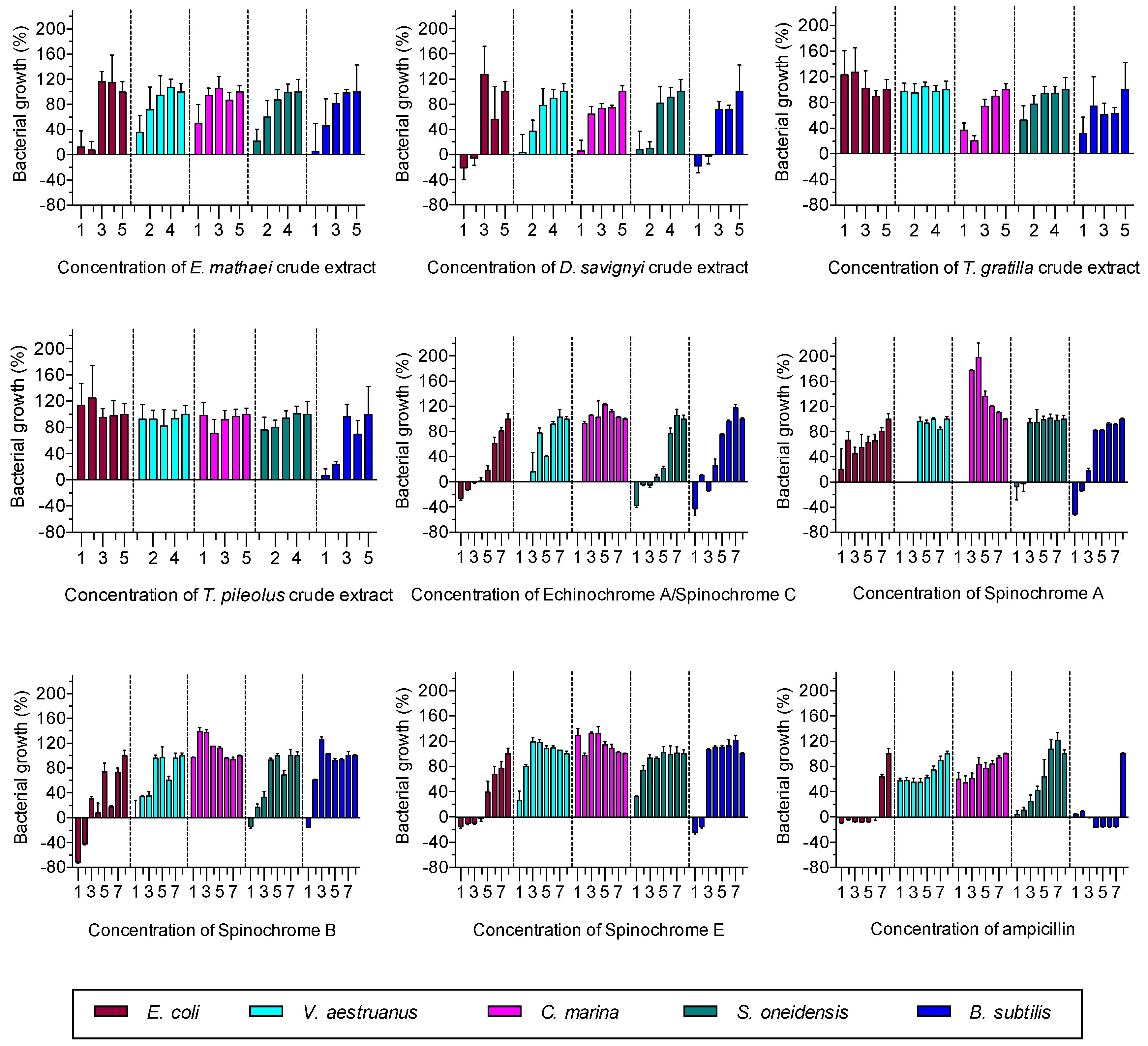
2.2.1. Antibacterial Activity Assay
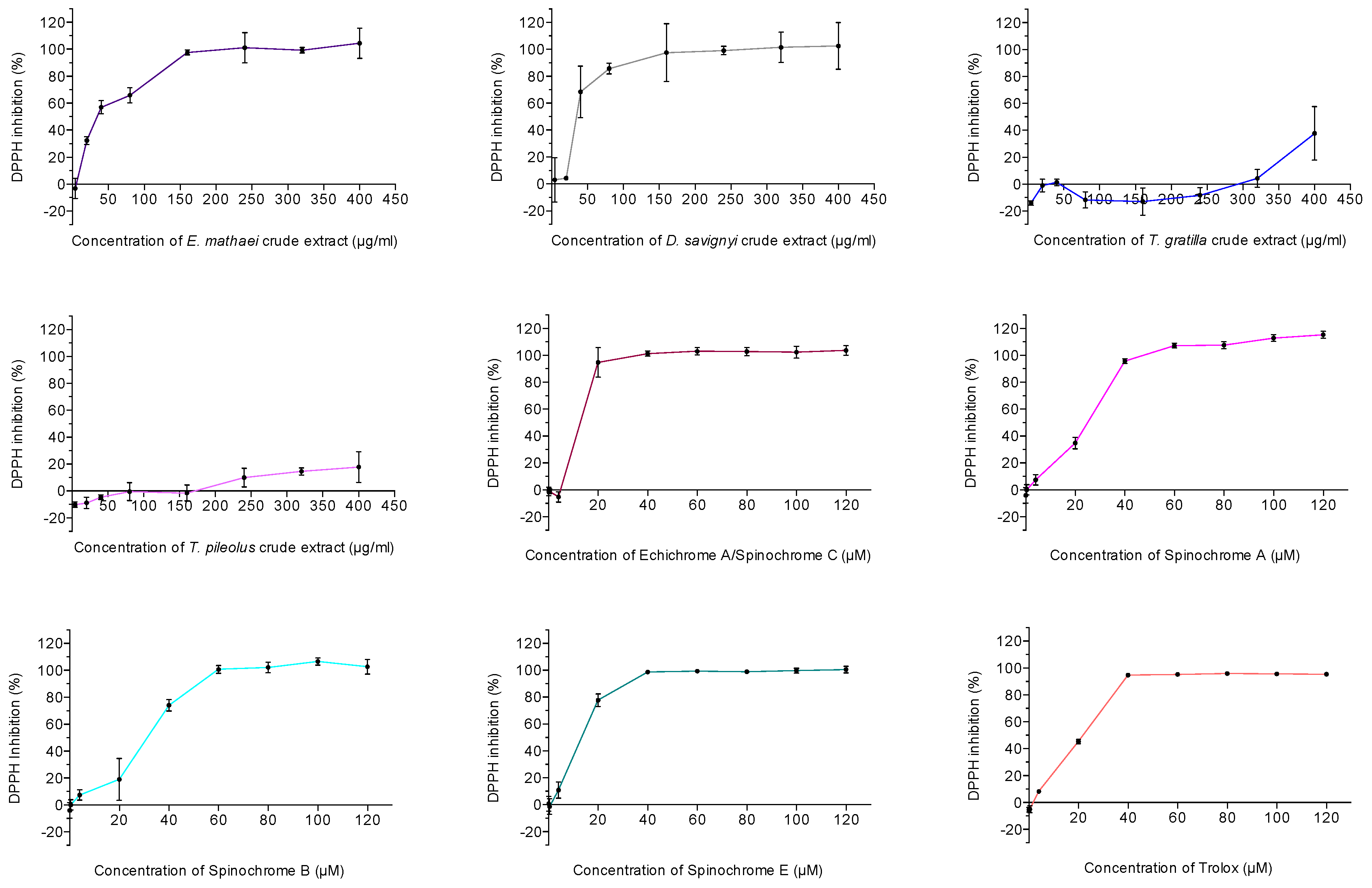
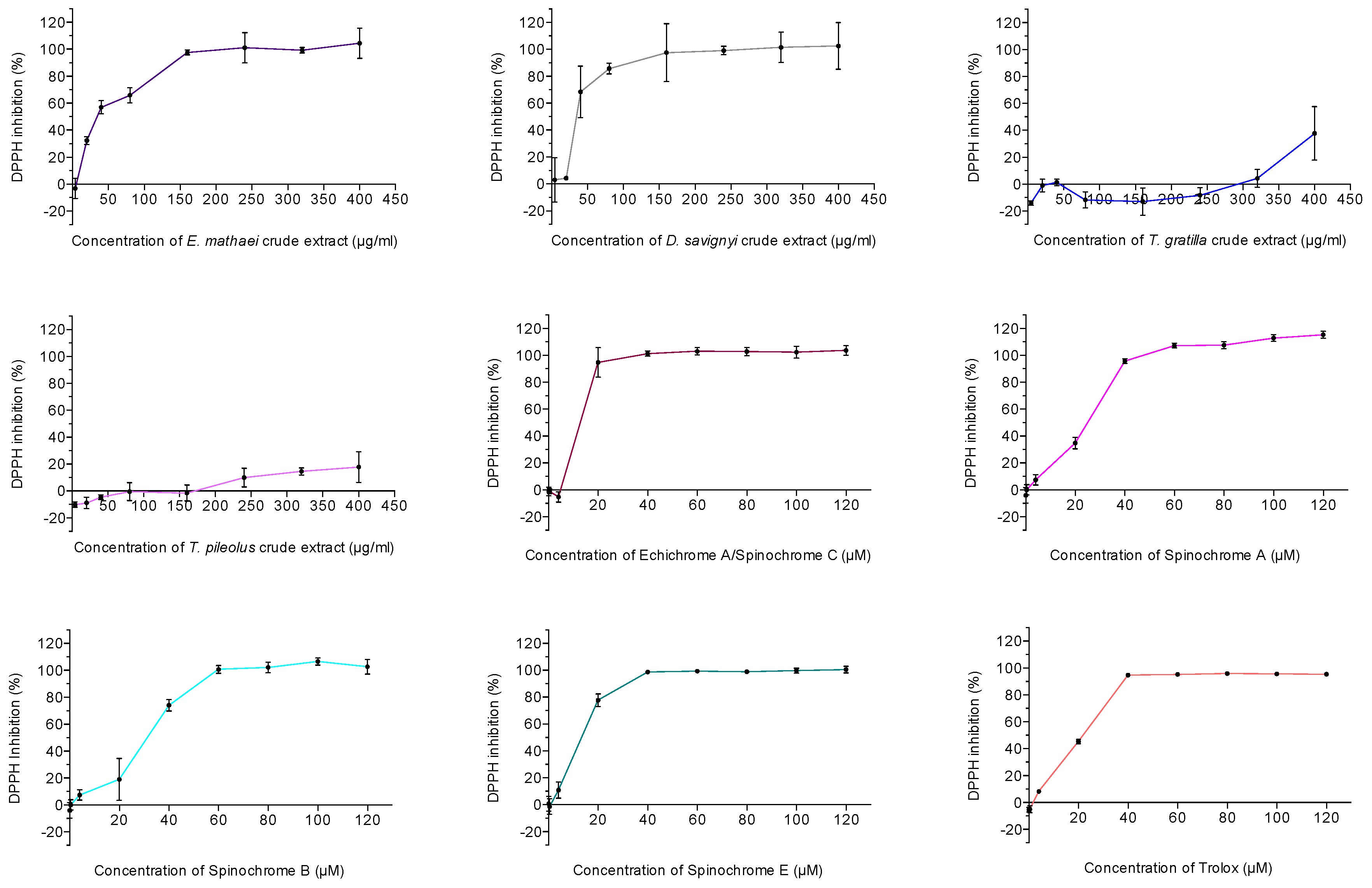
2.2.2. Antioxidant Activity Assay
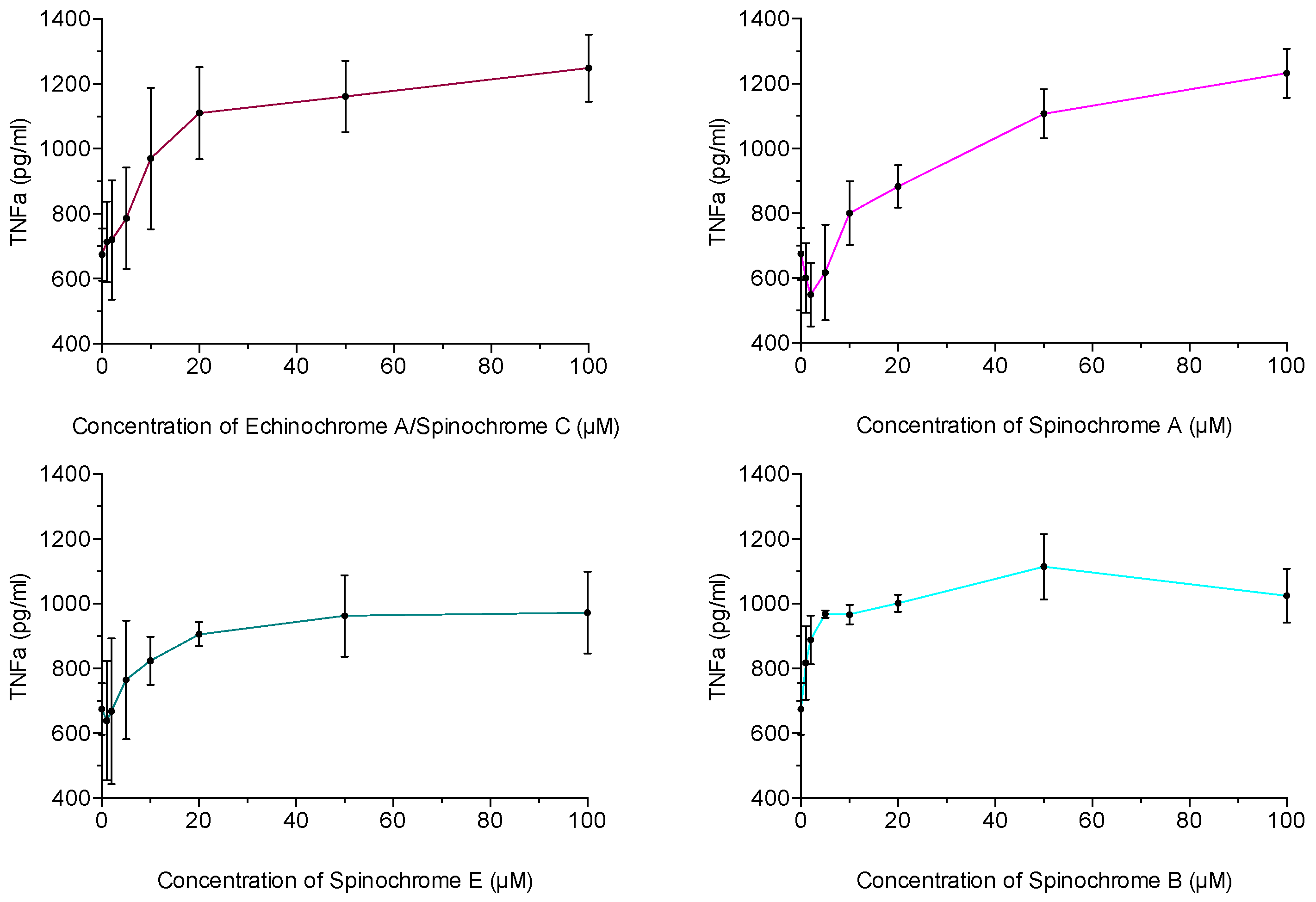
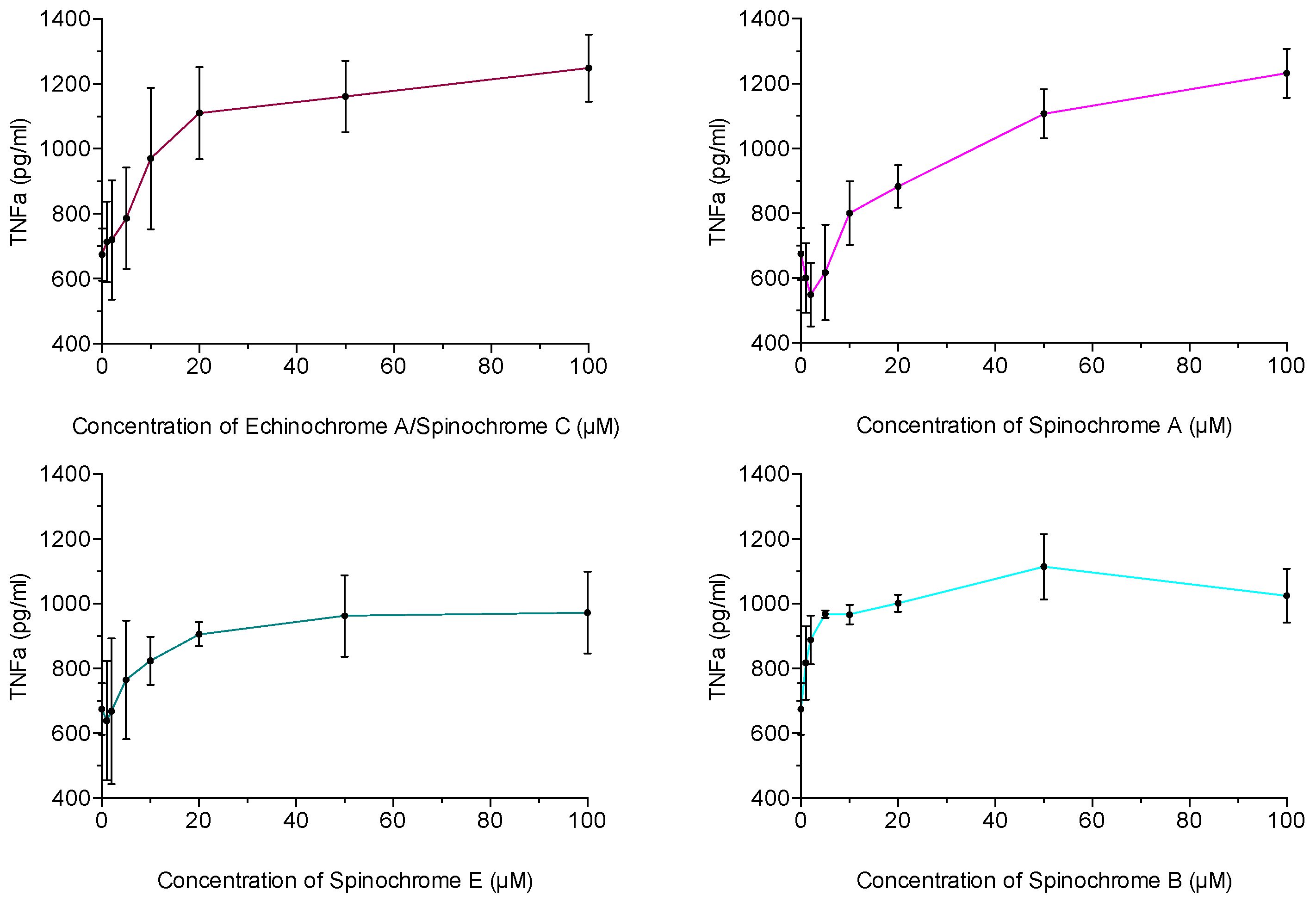
2.2.3. Inflammatory Activity Assay
2.2.4. Cytotoxic Activity Assay
3. Discussion
4. Materials and Methods
4.1. Extraction and Isolation of Spinochromes
4.1.1. Sampling
4.1.2. Preparation of Crude Extracts from Sea Urchins
4.1.3. Mass Spectrometry Analyses and Spinochrome Purification
4.2. Bioactive Effects of Spinochromes
4.2.1. Antibacterial Activity Assay
4.2.2. Antioxidant Activity Assay
4.2.3. Inflammatory Activity
4.2.4. Cytotoxic Activity
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Demain, A.L.; Fang, A. The natural functions of secondary metabolites. Adv. Biochem. Eng. Biotechnol. 2000, 69, 1–39. [Google Scholar] [CrossRef] [PubMed]
- Caulier, G.; Flammang, P.; Gerbaux, P.; Eeckhaut, I. When a repellent becomes an attractant: Harmful saponins are kairomones attracting the symbiotic Harlequin crab. Sci. Rep. 2013, 3, 2639. [Google Scholar] [CrossRef] [PubMed]
- Stonik, V.A.; Elyakov, G.B. Secondary Metabolites from Echinoderms as Chemotaxonomic Markers. In Bioorganic Marine Chemistry; Scheuer, P.P.J., Ed.; Springer: Berlin/Heidelberg, Germany, 1988; pp. 43–86. [Google Scholar]
- Kornprobst, J.-M. Substances Naturelles D’Origine Marine—Tome 2; LAVOISIER S.A.S.: Cachan, France, 2005. [Google Scholar]
- Brasseur, L.; Demeyer, M.; Decroo, C.; Caulier, G.; Flammang, P.; Gerbaux, P.; Eeckhaut, I. Identification and quantification of spinochromes in body compartments of Echinometra mathaei’s colored types. Mar. Biol. 2017, submitted. [Google Scholar]
- Service, M.; Wardlaw, A.C. Echinochrome-A as a bactericidal substance in the coelomic fluid of Echinus esculentus (L.). Comp. Biochem. Physiol. B Comp. Biochem. 1984, 79, 161–165. [Google Scholar] [CrossRef]
- Thomson, R.H. Distribution of naturally occurring quinones. Pharm. World Sci. 1991, 13, 70–73. [Google Scholar] [CrossRef]
- Mischenko, N.P.; Fedoreyev, S.A.; Pokhilo, N.D.; Anufriev, V.P.; Denisenko, V.A.; Glazunov, V.P. Echinamines A and B, first animated hydroxynaphthazarins from the sea urchin Scaphechinus mirabilis. J. Nat. Prod. 2005, 68, 1390–1393. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.-Y.; Qin, L.; Zhu, B.-W.; Wang, X.D.; Tan, H.; Yang, J.F.; Li, D.M.; Dong, X.P.; Wu, H.T.; Sun, L.M.; et al. Extraction and antioxidant property of polyhydroxylated naphthoquinone pigments from spines of purple sea urchin Strongylocentrotus nudus. Food Chem. 2011, 129, 1591–1597. [Google Scholar] [CrossRef]
- Stekhova, S.I.; Shentsova, E.B.; Kol’tsova, E.B.; Kulesh, N.I. Antimicrobial activity of polyhydroxynaphthoquinones from sea urchins. Antibiot. Khimioter. 1988, 33, 831–833. [Google Scholar] [PubMed]
- Haug, T.; Kjuul, A.K.; Styrvold, O.B.; Sandsdalen, E.; Olsen, Ø.M.; Stensvåg, K. Antibacterial activity in Strongylocentrotus droebachiensis (Echinoidea), Cucumaria frondosa (Holothuroidea), and Asterias rubens (Asteroidea). J. Invertebr. Pathol. 2002, 81, 94–102. [Google Scholar] [CrossRef]
- Shankarlal, S.; Prabu, K.; Natarajan, E. Antimicrobial and Antioxidant Activity of Purple Sea Urchin Shell (Salmacis virgulata L. Agassiz and Desor 1846). Am. J. Sci. Res. 2011, 6, 178–181. [Google Scholar]
- Fusaro, R.M.; Runge, W.J.; Johnson, J.A. Protection against light sensitivity with dihydroxyacetone/naphthoquinone. Int. J. Dermatol. 1972, 11, 67–70. [Google Scholar] [CrossRef] [PubMed]
- Lebedev, A.V.; Levitskaya, E.L.; Tikhonova, E.V.; Ivanova, M.V. Antioxidant Properties, Autooxidation, and Mutagenic Activity of Echinochrome A Compared with Its Etherified Derivative. Biochemistry 2001, 66, 885–893. [Google Scholar] [CrossRef] [PubMed]
- Lebedev, A.V.; Ivanova, M.V.; Levitsky, D.O. Echinochrome, a naturally occurring iron chelator and free radical scavenger in artificial and natural membrane systems. Life Sci. 2005, 76, 863–875. [Google Scholar] [CrossRef] [PubMed]
- Kuwahara, R.; Hatate, H.; Yuki, T.; Murata, H.; Tanaka, R.; Hama, Y. Antioxidant property of polyhydroxylated naphthoquinone pigments from shells of purple sea urchin Anthocidaris crassispina. LWT Food. Sci. Technol. 2009, 42, 1296–1300. [Google Scholar] [CrossRef]
- Kuwahara, R.; Hatate, H.; Chikami, A.; Murata, H.; Kijidani, Y. Quantitative separation of antioxidant pigments in purple sea urchin shells using a reversed-phase high performance liquid chromatography. LWT Food. Sci. Technol. 2010, 43, 1185–1190. [Google Scholar] [CrossRef]
- Li, D.-M.; Zhou, D.-Y.; Zhu, B.-W.; Miao, L.; Qin, L.; Dong, X.P.; Wang, X.D.; Murata, Y. Extraction, structural characterization and antioxidant activity of polyhydroxylated 1,4-naphthoquinone pigments from spines of sea urchin Glyptocidaris crenularis and Strongylocentrotus intermedius. Eur. Food Res. Technol. 2013, 237, 331–339. [Google Scholar] [CrossRef]
- Lee, S.R.; Pronto, J.R.D.; Sarankhuu, B.-E.; Ko, K.S.; Rhee, B.D.; Kim, N.; Mishchenko, N.P.; Fedoreyev, S.A.; Stonik, V.A.; Han, J. Acetylcholinesterase inhibitory activity of pigment echinochrome A from sea urchin Scaphechinus mirabilis. Mar. Drugs 2014, 12, 3560–3573. [Google Scholar] [CrossRef] [PubMed]
- Powell, C.; Hughes, A.D.; Kelly, M.S.; Conner, S.; McDougall, G.J. Extraction and identification of antioxidant polyhydroxynaphthoquinone pigments from the sea urchin, Psammechinus miliaris. LWT Food Sci. Technol. 2014, 59, 455–460. [Google Scholar] [CrossRef]
- Pozharitskaya, O.N.; Shikov, A.N.; Makarova, M.N.; Ivanova, S.A.; Kosman, V.M.; Makarov, V.G.; Bazgier, V.; Berka, K.; Otyepka, M.; Ulrichová, J. Antiallergic effects of pigments isolated from green sea urchin (Strongylocentrotus droebachiensis) shells. Planta Med. 2013, 79, 1698–1704. [Google Scholar] [CrossRef] [PubMed]
- Lennikov, A.; Kitaichi, N.; Noda, K.; Mizuuchi, K.; Ando, R.; Dong, Z.; Fukuhara, J.; Kinoshita, S.; Namba, K.; Ohno, S.; et al. Amelioration of endotoxin-induced uveitis treated with the sea urchin pigment echinochrome in rats. Mol. Vis. 2014, 20, 171–177. [Google Scholar] [PubMed]
- Sciani, J.M.; Zychar, B.C.; Gonçalves, L.R.D.C.; Nogueira, T.D.O.; Giorgi, R.; Pimenta, D.C. Pro-inflammatory effects of the aqueous extract of Echinometra lucunter sea urchin spines. Exp. Biol. Med. 2011, 236, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Suárez-Peñaranda, J.M.; Vieites, B.; Del Río, E.; Ortiz-Rey, J.A.; Antón, I. Histopathologic and immunohistochemical features of sea urchin granulomas. J. Cutan. Pathol. 2013, 40, 550–556. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, H. The Extracellular Release of Echinochrome. J. Gen. Physiol. 1946, 29, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y. Distribution and redistribution of pigment granules in the development of sea urchin embryos. Wilhelm Roux’s Arch. Dev. Biol. 1981, 190, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Floyd, R.A. Antioxidants, oxidative stress, and degenerative neurological disorders. Soc. Exp. Biol. Med. 1999, 222, 236–245. [Google Scholar] [CrossRef]
- Pham-huy, L.A.; He, H.; Pham-huy, C. Free Radicals, Antioxidants in Disease and Health. Int. J. Biomed. Sci. 2008, 4, 89–96. [Google Scholar] [PubMed]
- Uttara, B.; Singh, A.V.; Zamboni, P.; Mahajan, R.T. Oxidative Stress and Neurodegenerative Diseases: A Review of Upstream and Downstream Antioxidant Therapeutic Options. Curr. Neuropharmacol. 2009, 7, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Kovaleva, M.A.; Ivanova, S.A.; Makarova, M.N.; Pozharitskaia, O.N.; Shikov, A.N.; Makarov, V.G. Effect of a complex preparation of sea urchin shells on blood glucose level and oxidative stress parameters in type II diabetes model. Eksp. Klin. Farmakol. 2013, 76, 27–30. [Google Scholar] [PubMed]
- Egorov, E.A.; Alekhina, V.A.; Volobueva, T.M.; Fedoreev, S.A.; Mishchenko, N.P.; Kol’tsova, E.A. Histochrome, a new antioxidant, in the treatment of ocular diseases. Vestnik Oftalmol. 1999, 115, 34–35. [Google Scholar]
- Soleimani, S.; Moein, S.; Yousefzadi, M.; Bioki, N.A. Determination of In Vitro Antioxidant Properties, Anti-inflammatory Effects and A-Amylase Inhibition of Purple Sea Urchin Extract of Echinometra Mathaei from the Persian Gulf. Jundishapur J. Nat. Pharm. Prod. 2016, in press. [Google Scholar] [CrossRef]
- Shikov, A.N.; Ossipov, V.I.; Martiskainen, O.; Pozharitskaya, O.N.; Ivanova, S.A.; Makarov, V.G. The offline combination of thin-layer chromatography and high-performance liquid chromatography with diode array detection and micrOTOF-Q mass spectrometry for the separation and identification of spinochromes from sea urchin (Strongylocentrotus droebachi. J. Chromatogr. A 2011, 1218, 9111–9114. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Shavandi, A.; Carne, A.; Bekhit, A.A.; Ng, T.B.; Cheung, R.C.F.; Bekhit, A.E.D.A. Marine shells: Potential opportunities for extraction of functional and health-promoting materials. Crit. Rev. Environ. Sci. Technol. 2016, 46, 1047–1116. [Google Scholar] [CrossRef]
- Khamala, C.P.M. Ecology of Echinometra mathaei (Echinoidea: Echinodermata) at Diani Beach, Kenya. Mar. Biol. 1971, 11, 167–172. [Google Scholar] [CrossRef]
- McClanahan, T.R.; Muthiga, N.A. Ecology of Echinometra. In Edible Sea Urchins: Biology and Ecology; Lawrence, J.M., Ed.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 297–317. [Google Scholar]
- Muthiga, N.A.; McClanahan, T.R. Ecology of Diadema. In Edible Sea Urchins: Biology and Ecology; Lawrence, J.M., Ed.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 205–225. [Google Scholar]
- Vaïtilingon, D. The Biology and Ecology of the Echinoid Tripneustes Gratilla (Linneaus, 1758) off Toliara (Madagascar): Feeding, Reproduction, Larval Dévelopment, Population Dynamiics and Parasitism; Université Libre de Bruxelles: Bruxelles, Belgium, 2004. [Google Scholar]
- Ziegenhorn, M.A. Best Dressed Test: A Study of the Covering Behavior of the Collector Urchin Tripneustes gratilla. PLoS ONE 2016, 11, e0153581. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Soong, K. “Uncovering” behavior at spawning of the trumpet sea urchin Toxopneustes pileolus. Zool. Stud. 2010, 49, 9. [Google Scholar]
- Claereboudt, E.J.S.; Claereboudt, M.R. Quantifying sea urchins covering behavior by image analysis. Cah. Biol. Mar. 2013, 54, 587–592. [Google Scholar]
- Thormann, K.M.; Saville, R.M.; Shukla, S.; Pelletier, D.A.; Spormann, A.M. Initial Phases of Biofilm Formation in Initial Phases of Biofilm Formation in Shewanella oneidensis MR-1. J. Bacteriol. 2004, 186, 8096–8104. [Google Scholar] [CrossRef] [PubMed]
- Azandegbe, A. Etude de la Structure des Communautés Bactériennes du Sédiment et de L’Ecologie de Vibrio Aestuarianus Pathogène de L’Huître Creuse Crassostrea Gigas dans deux Sites Ostréicoles; Université de Brest: Brest, France, 2010. [Google Scholar]
- Mieszkin, S.; Martin-Tanchereau, P.; Callow, M.E.; Callow, J.A. Effect of bacterial biofilms formed on fouling-release coatings from natural seawater and Cobetia marina, on the adhesion of two marine algae. Biofouling 2012, 28, 953–968. [Google Scholar] [CrossRef] [PubMed]
- Blois, M. Antioxidant Determinations by the Use of a Stable Free Radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Fernandes, C.A.; Fievez, L.; Neyrinck, A.M.; Delzenne, N.M.; Bureau, F.; Vanbever, R. Biochemical and Biophysical Research Communications Sirtuin inhibition attenuates the production of inflammatory cytokines in lipopolysaccharide-stimulated macrophages. Biochem. Biophys. Res. Commun. 2012, 420, 857–861. [Google Scholar] [CrossRef] [PubMed]
- Strober, W. Trypan Blue Exclusion Test of Cell Viability. Curr. Protoc. Immunol. 2001, 111, A3-B. [Google Scholar] [CrossRef]
- Bellamakondi, P.K.; Godavarthi, A.; Ibrahim, M.; Kulkarni, S.; Naik, R.M.; Maradam, S. In Vitro cytotoxicity of caralluma species by MTT and trypan blue dye exclusion. Asian J. Pharm. Clin. Res. 2014, 7, 7–9. [Google Scholar]
- Strober, W. Trypan Blue Exclusion Test of Cell Viability. Curr. Protoc. Immunol. 2015, 111, A3-B. [Google Scholar] [CrossRef] [PubMed]
- Anderson, H.A.; Mathieson, J.W.; Thomson, R.H. Distribution of spinochrome pigments in echinoids. Comp. Biochem. Physiol. 1969, 28, 333–345. [Google Scholar] [CrossRef]
- Thomson, R.H. Naturally Occuring Quinones, 2nd ed.; Academic Press Inc.: New York, NY, USA, 1971. [Google Scholar]
- Hatate, H.; Murata, H.; Hama, Y.; Tanaka, R.; Suzuki, N. Antioxidative activity of spinochromes extracted from shells of sea urchins. Fish Sci. 2002, 68, 1641–1642. [Google Scholar] [CrossRef]
- McClanahan, T.R.; Muthiga, N.A. Patterns of preedation on a sea urchin, Echinometra mathaei (de Blainville), on Kenyan coral reefs. J. Exp. Mar. Biol. Ecol. 1989, 126, 77–94. [Google Scholar] [CrossRef]
- Sala, E. Fish predators and scavengers of the sea urchin Paracentrotus lividus in protected areas of the north-west Mediterranean Sea. Mar. Biol. 1997, 129, 531–539. [Google Scholar] [CrossRef]
- Jangoux, M. Diseases of echinoderms. Helgol. Meeresunters. 1984, 37, 207–216. [Google Scholar] [CrossRef]
- Jangoux, M. Diseases of Echinodermata. III. Agents metazoans (Annelida to Pisces). Dis. Aquat. Organ. 1987, 3, 59–83. [Google Scholar] [CrossRef]
- Vaïtilingon, D.; Eeckhaut, I.; Fourgon, D.; Jangoux, M. Population dynamics, infestation and host selection of Vexilla vexillum, an ectoparasitic muricid of echinoids, in Madagascar. Dis. Aquat. Organ. 2004, 61, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Becker, P.T.; Egea, E.; Eeckhaut, I. Characterization of the bacterial communities associated with the bald sea urchin disease of the echinoid Paracentrotus lividus. J. Invertebr. Pathol. 2008, 98, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Becker, P.T.; Gillan, D.C.; Eeckhaut, I. Characterization of the bacterial community associated with body wall lesions of Tripneustes gratilla (Echinoidea) using culture-independent methods. J. Invertebr. Pathol. 2009, 100, 127–130. [Google Scholar] [CrossRef] [PubMed]
- Shikov, A.N.; Ossipov, V.I.; Karonen, M.; Pozharitskaya, O.N.; Krishtopina, A.S.; Makarov, V.G. Comparative stability of dimeric and monomeric pigments extracted from sea urchin Strongylocentrotus droebachiensis. Nat. Prod. Res. 2017, 6419, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Becker, P.; Gillan, D.; Eeckhaut, I. Microbiological study of the body wall lesions of the echinoid Tripneustes gratilla. Dis. Aquat. Organ. 2007, 77, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Tanigawa, T.; Tomita, K.; omihara, Y.; Araki, Y.; Tachikawa, E. Recent Studies on the Pathological Effects of Purified Sea Urchin Toxins. J. Toxicol. Toxin Rev. 2003, 22, 633–649. [Google Scholar] [CrossRef]
- Akinbowale, O.L.; Peng, H.; Barton, M.D. Antimicrobial resistance in bacteria isolated from aquaculture sources in Australia. J. Appl. Microbiol. 2006, 100, 1103–1113. [Google Scholar] [CrossRef] [PubMed]
- Kathleen, M.M.; Samuel, L.; Felecia, C.; Reagan, E.L.; Kasing, A.; Lesley, M.; Toh, S.C. Antibiotic Resistance of Diverse Bacteria from Aquaculture in Borneo. Int. J. Microbiol. 2016, 2016, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lesser, M.P.; Farrell, J.H.; Walker, C.W. Oxidative stress, DNA damage and p53 expression in the larvae of atlantic cod (Gadus morhua) exposed to ultraviolet (290–400 nm) radiation. J. Exp. Biol. 2001, 204, 157–164. [Google Scholar] [PubMed]
- Lesser, M.P.; Barry, T.M. Survivorship, development, and DNA damage in echinoderm embryos and larvae exposed to ultraviolet radiation (290–400 nm). J. Exp. Mar. Biol. Ecol. 2003, 292, 75–91. [Google Scholar] [CrossRef]
- Lesser, M.P. Exposure to ultraviolet radiation causes apoptosis in developing sea urchin embryos. J. Exp. Biol. 2003, 206, 4097–4103. [Google Scholar] [CrossRef] [PubMed]
- Asada, K. Production and Scavenging of Reactive Oxygen Species in Chloroplasts and Their Functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Flora, S.J.S. Structural, Chemical and Biological Aspects of Antioxidants for Strategies Against Metal and Metalloid Exposure. Oxid. Med. Cell. Longev. 2009, 2, 191–206. [Google Scholar] [CrossRef] [PubMed]
- Slavova-Kazakova, A.K.; Angelova, S.E.; Veprintsev, T.L.; Denev, P.; Fabbri, D.; Dettori, M.A.; Kratchanova, M.; Naumov, V.V.; Trofimov, A.V.; Vasil’ev, R.F.; et al. Antioxidant potential of curcumin-related compounds studied by chemiluminescence kinetics, chain-breaking efficiencies, scavenging activity (ORAC) and DFT calculations. Beilstein J. Org. Chem. 2015, 11, 1398–1411. [Google Scholar] [CrossRef] [PubMed]
- Pozharitskaya, O.N.; Ivanova, S.A.; Shikov, A.N.; Makarov, V.G. Evaluation of Free Radical-Scavenging Activity of Sea Urchin Pigments Using HPTLC with Post-Chromatographic Derivatization. Chromatographia 2013, 76, 1353–1358. [Google Scholar] [CrossRef]
- Inoue, K.; Takano, H.; Hiyoshi, K.; Ichinose, T.; Sadakane, K.; Yanagisawa, R.; Tomura, S.; Kumaga, Y. Naphthoquinone enhances antigen-related airway inflammation in mice. Eur. Respir. J. 2006, 29, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Bodet, C.; Chandad, F.; Grenier, D. Hemoglobin and LPS Act in Synergy to Amplify the Inflammatory Response. J. Dent. Res. 2007, 86, 878–882. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.C.; Britten, R.J.; Davidson, E.H. Lipopolysaccharide activates the sea urchin immune system. Dev. Comp. Immunol. 1995, 19, 217–224. [Google Scholar] [CrossRef]
- Nair, S.V.; Del Valle, H.; Gross, P.S.; Terwilliger, D.P.; Smith, L.C. Macroarray analysis of coelomocyte gene expression in response to LPS in the sea urchin. Identification of unexpected immune diversity in an invertebrate. Physiol. Genom. 2005, 22, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Majeske, A.J.; Bayne, C.J.; Smith, L.C. Aggregation of Sea Urchin Phagocytes Is Augmented In Vitro by Lipopolysaccharide. PLoS ONE 2013, 8, e61419. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Aravena, M.; Perez-troncoso, C.; Urtubia, R.; Branco, P.; Da Silva, J.R.M.C.; Mercado, L.; De Lorgeril, J.; Bethke, J.; Paschke, K. Immune response of the Antarctic sea urchin Sterechinus neumayeri: Cellular, molecular and physiological approach. Rev. Biol. Trop. 2015, 63, 309–320. [Google Scholar] [CrossRef]
- Ioset, J.-R.; Brun, R.; Wenzler, T.; Kaiser, M.; Yardley, V. Drug Screening for Kinetoplastid Diseases: A Training Manual for Screening in Neglected Diseases; DNDi and Pan-Asian Screening Network: London, UK, 2009. [Google Scholar]
- Lambert, R.J.W.; Pearson, J. Susceptibility testing: Accurate and reproducible minimum inhibitory concentration (MIC) and non-inhibitory concentration (NIC) values. J. Appl. Microbiol. 2000, 88, 784–790. [Google Scholar] [CrossRef] [PubMed]
- Kedare, S.B.; Singh, R.P. Genesis and development of DPPH method of antioxidant assay. J. Food Sci. Technol. 2011, 48, 412–422. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PHNQ | Retention Time (min) | MW (U) | Predicted Formula A | PHNQ Concentration (MG of PHNQ/KG of Dried Tests/Spines) B | |||
|---|---|---|---|---|---|---|---|
| E. mathaei | D. savignyi | T. gratilla | T. pileolus | ||||
| Spinochrome B | 2.75 | 222 | C10H606 | 9.2 ± 6.9 | - | - | 4.5 ± 4.4 |
| Spinochrome D — Iso 1 | 2.65 | 238 | C10H6O7 | - | 1.6 ± 1.7 | - | 0.6 ± 0.8 |
| Spinochrome D — Iso 3 | 8.65 | 238 | C10H6O7 | - | - | 0.1 ± 0.1 | - |
| Spinochrome E | 1.73 | 254 | C10H6O8 | 4.9 ± 2.1 | - | 0.2 ± 0.2 | 0.4 ± 0.6 |
| Spinochrome A — Iso 2 | 7.95 | 264 | C12H8O7 | 13.0 ± 5.0 | - | - | 0.6 ± 0.7 |
| Echinochrome A | 6.69 | 266 | C12H10O7 | 10.3 ± 2.2 | 17.6 ± 8.8 | 0.1 ± 0.1 | 0.2 ± 0.2 |
| Spinochrome C | 6.73 | 280 | C12H8O8 | 2.4 ± 1.2 | - | - | 0.6 ± 0.9 |
| Antibacterial Activity | |||||||
|---|---|---|---|---|---|---|---|
| EC50 and MIC—Bacterial Growth | |||||||
| E. coli | B. subtilis | V. aestuarianus | C. marina | S. oneidensis | |||
| Tests/Spines Crude Extracts (μG/mL) | |||||||
| E. mathaei | 688.40 | 716.70 | 1518.00 | 1995.00 | 1118.00 | EC50 | |
| >2000 | >2000 | >2000 | >2000 | >2000 | MIC | ||
| D. savignyi | 358.70 | 252.40 | 534.80 | 681.00 | 391.00 | EC50 | |
| 406.80 | 1219.00 | >2000 | >2000 | >2000 | MIC | ||
| T. gratilla | >2000 | 1227.00 | >2000 | 543.80 | >2000 | EC50 | |
| >2000 | >2000 | >2000 | >2000 | >2000 | MIC | ||
| T. pileolus | >2000 | 512.60 | >2000 | >2000 | >2000 | EC50 | |
| >2000 | >2000 | >2000 | >2000 | >2000 | MIC | ||
| Isolated Spinochromes (μM; μG/mL) | |||||||
| Echinochrome A/Spinochrome C | 22.56 | 60.98 | - | >1000 | 27.88 | μM | EC50 |
| 6.08 | 16.43 | >269 | 7.51 | μG/mL | |||
| 54.19 | 149.40 | - | 628.50 | 48.26 | μM | MIC | |
| 14.60 | 40.26 | 169.38 | 13.00 | μG/mL | |||
| Spinochrome A | 199.40 | 139.20 | - | >1000 | 238.20 | μM | EC50 |
| 52.64 | 36.75 | >264 | 62.88 | μG/mL | |||
| >1000 | 228.40 | - | >1000 | 435.70 | μM | MIC | |
| >264 | 60.30 | >254 | 115.02 | μG/mL | |||
| Spinochrome B | 16.22 | 510.50 | 214.70 | >1000 | 172.40 | μM | EC50 |
| 3.60 | 113.331 | 47.66 | >222 | 38.27 | μG/mL | ||
| 468.30 | 583.50 | >1000 | >1000 | 238.60 | μM | MIC | |
| 103.96 | 129.54 | >222 | >222 | 52.97 | μG/mL | ||
| Spinochrome E | 28.53 | 295.20 | 746.70 | >1000 | 742.60 | μM | EC50 |
| 7.25 | 74.98 | 189.66 | >254 | 188.62 | μG/mL | ||
| 90.33 | 422.40 | >1000 | >1000 | >1000 | μM | MIC | |
| 22.94 | 107.29 | >254 | >254 | >254 | μG/mL | ||
| Ampicilin (μM; μG/mL) | 4.26 | 1.31 | >1000 | >1000 | 83.55 | μM | EC50 |
| 1.49 | 0.46 | >349 | >349 | 65.82 | μG/mL | ||
| 8.10 | 1.80 | >1000 | >1000 | 272.00 | μM | MIC | |
| 2.83 | 0.63 | >349 | >349 | 94.93 | μG/mL | ||
| Antioxidant Activity | ||
|---|---|---|
| EC50—DPPH Inhibition | ||
| Sea Urchin Crude Extracts | (μG/mL) | |
| E. mathaei | 35.63 | |
| D. savignyi | 34.46 | |
| T. gratilla | >400 | |
| T. pileolus | >400 | |
| Isolated Spinochromes | (μM) | (μG/mL) |
| Echinochrome A/Spinochrome C | 16.31 | 4.40 |
| Spinochrome A | 22.17 | 5.85 |
| Spinochrome B | 29.38 | 6.52 |
| Spinochrome E | 10.85 | 2.76 |
| Trolox (μM) | 20.88 | 5.22 |
| Cytotoxic Activity | |
|---|---|
| EC50—Cell Viability | |
| Isolated Spinochromes (μM) | |
| Echinochrome A/Spinochrome C | 380.9 |
| Spinochrome A | 341.7 |
| Spinochrome B | >500 |
| Spinochrome E | 258.6 |
| Time (Min) | Eluent A (%) | Eluent B (%) | Curve |
|---|---|---|---|
| 00 | 80 | 20 | Equilibration |
| 00 → 15 | 80 → 50 | 20 → 50 | Linear gradient |
| 15 → 16 | 50 → 80 | 50 → 80 | Linear gradient |
| 16 → 18 | 80 | 20 | Isocratic |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brasseur, L.; Hennebert, E.; Fievez, L.; Caulier, G.; Bureau, F.; Tafforeau, L.; Flammang, P.; Gerbaux, P.; Eeckhaut, I. The Roles of Spinochromes in Four Shallow Water Tropical Sea Urchins and Their Potential as Bioactive Pharmacological Agents. Mar. Drugs 2017, 15, 179. https://doi.org/10.3390/md15060179
Brasseur L, Hennebert E, Fievez L, Caulier G, Bureau F, Tafforeau L, Flammang P, Gerbaux P, Eeckhaut I. The Roles of Spinochromes in Four Shallow Water Tropical Sea Urchins and Their Potential as Bioactive Pharmacological Agents. Marine Drugs. 2017; 15(6):179. https://doi.org/10.3390/md15060179
Chicago/Turabian StyleBrasseur, Lola, Elise Hennebert, Laurence Fievez, Guillaume Caulier, Fabrice Bureau, Lionel Tafforeau, Patrick Flammang, Pascal Gerbaux, and Igor Eeckhaut. 2017. "The Roles of Spinochromes in Four Shallow Water Tropical Sea Urchins and Their Potential as Bioactive Pharmacological Agents" Marine Drugs 15, no. 6: 179. https://doi.org/10.3390/md15060179