A Transcriptomic Survey of Ion Channel-Based Conotoxins in the Chinese Tubular Cone Snail (Conus betulinus)


1
BGI Education Center, University of Chinese Academy of Sciences, Shenzhen 518083, China
2
Shenzhen Key Lab of Marine Genomics, Guangdong Provincial Key Lab of Molecular Breeding in Marine Economic Animals, BGI Academy of Marine Sciences, BGI Marine, BGI, Shenzhen 518083, China
3
Hainan Provincial Key Laboratory of Research and Development of Tropical Medicinal Plants, Hainan Medical University, Haikou 571199, China
4
Laboratory of Aquatic Genomics, College of Life Sciences and Oceanography, Shenzhen University, Shenzhen 518060, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Mar. Drugs 2017, 15(7), 228; https://doi.org/10.3390/md15070228
Submission received: 31 May 2017
/
Revised: 10 July 2017
/
Accepted: 13 July 2017
/
Published: 18 July 2017
(This article belongs to the Special Issue Marine Drugs and Ion Currents)
Abstract
:Conotoxins in the venom of cone snails (Conus spp.) are a mixture of active peptides that work as blockers, agonists, antagonists, or inactivators of various ion channels. Recently we reported a high-throughput method to identify 215 conotoxin transcripts from the Chinese tubular cone snail, C. betulinus. Here, based on the previous datasets of four transcriptomes from three venom ducts and one venom bulb, we explored ion channel-based conotoxins and predicted their related ion channel receptors. Homologous analysis was also performed for the most abundant ion channel protein, voltage-gated potassium (Kv; with Kv1.1 as the representative), and the most studied ion channel receptor, nicotinic acetylcholine receptor (nAChR; with α2-nAChR as the representative), in different animals. Our transcriptomic survey demonstrated that ion channel-based conotoxins and related ion channel proteins/receptors transcribe differentially between the venom duct and the venom bulb. In addition, we observed that putative κ-conotoxins were the most common conotoxins with the highest transcription levels in the examined C. betulinus. Furthermore, Kv1.1 and α2-nAChR were conserved in their functional domains of deduced protein sequences, suggesting similar effects of conotoxins via the ion channels in various species, including human beings. In a word, our present work suggests a high-throughput way to develop conotoxins as potential drugs for treatment of ion channel-associated human diseases.
1. Introduction
Cone snail (Conus spp.) is the common name for a large genus of small to large-sized predatory marine gastropod molluscs that feed on a variety of prey, including worms, other molluscs, and even small fish [1,2,3]. Corresponding to these prey preferences, cone snails are classified into three groups, i.e., the vermivorous, molluscivorous, and piscivorous species [1]. Nowadays, after more than 55 million years of evolution, there are about 700 described cone snail species around the world [4,5]. All cone snails constitute a single, but the largest, genus of invertebrate marine animals, which is the only genus of the Conidae family within the Conoidea superfamily which belongs to the taxonomic class Neogastropoda [6,7]. Conus species inhabit throughout tropical, subtropical, and temperate oceans in the Indo-West Pacific region, such as South China sea, Philippines and Australia [8]. Despite the relatively young age of the genus Conus, the slow-moving cone snails have developed successful strategies to subdue prey and defence against foes [5,9], including a highly complex cocktail of potent venom components with a great specificity towards a wide range of physiological targets [10,11]. The sophisticated venom apparatus consists of venom bulb, venom duct, and the hollow harpoon-like radula [12,13].
The venom components are produced in the venom duct and pushed into a radula through systolic pressure of the venom bulb, and then discharged into prey through the radula [5,14]. The modified radulae can be fired at the targets in a harpoon-like action and, subsequently, the venom will be discharged into the victim’s circulatory system, interacting with a range of molecular targets in the nervous system and rapidly immobilizing prey or predators in a few seconds [5,10,11,14].
The major bioactive components of Conus venoms are a vast array of unique neurotoxic peptides, commonly referred to as conotoxins [13]. Each Conus species contains an average of over 100 unique conotoxin transcripts and 1000–2000 various conotoxin peptides [9,13,15,16,17,18]; therefore, up to 70,000 transcripts and 1,000,000 natural conotoxin peptides may exist in the world, which constitute a great potential resource for pharmacological research and development.
The small sizes of conotoxins, generally 10–30 amino acids in length [19], are produced as precursors comprising a highly-conserved N-terminal signal region followed by a less conserved propeptide region and a hypervariable C-terminal mature region [20,21]. Historically, the modified mature peptides were divided into disulfide-rich and disulfide-poor conotoxin groups based on the number of disulfide bond [22], although this distinction is now considered redundant [23]. The disulfide-poor conopeptides, having none or only one disulfide bond, are further subdivided into contulakines, conantokines, conorfamids, conolysines, conopressins, contryphans, conophans, conomarphines and conomaps [24,25]. With multiple disulfide bonds, the disulfide-rich conotoxins are classified based on three criteria, including the signal region for defining superfamilies (capitalized letters with Arabic numerals), the cysteine pattern (number and connectivity) for defining cysteine frameworks (Roman numerals), and the receptor targets and types of interaction for defining pharmacology families (using lower-case Greek letters) [25,26]. Conotoxins are currently classified into a total of 27 gene superfamilies [13,23,27,28] and 13 temporary gene superfamilies for those identified in the early divergent species [28,29,30]. Conotoxins with a similar cysteine network usually carry a similar signal sequence. So far, these distinct cysteine frameworks have been described in conotoxins and they are often considered to be associated with particular pharmacological families [21,28,29,31,32].
According to the wide range of distinct ion channels, receptors, and transporters in the nervous systems and the interaction types (agonist, antagonist, or delayed inactivation), 12 pharmacological families of conotoxins (α-, γ-, δ-, ε-, ι-, κ-, μ-, ρ-, σ-, τ-, and χ-families) have been defined. Please see more details about definitions of each family in Table 1 [25,28]. Due to their exquisite specificity for receptor subtypes, conotoxins are valuable tools in neurological studies. Several conotoxins are being developed as drugs or drug leads [33,34,35,36,37,38]. The most well-known calcium channel blocker, synthetic ω-conotoxin MVIIA, was approved by the American FDA in 2004 to treat severe and chronic pain [39]. More and more conotoxins are undergoing to be developed for curing many kinds of human diseases, including chronic pain, epilepsy, drug addiction, Parkinson’s disease, and various neurological diseases [40,41].
In our previous report [13], a total of 215 distinct conotoxin transcripts were identified from the vermivorous C. betulinus, using both next-generation sequencing (RNA-seq) and traditional Sanger sequencing technologies. In the current study, based on the reported four transcriptomes from the venom duct and the venom bulb, we are interested in exploring ion channel active conotoxins since they play important roles inhibiting or activating ion channel receptors, we also investigated related ion channel proteins and receptors in the transcription datasets. Homologous analysis was performed for the most abundant ion channel protein, voltage-gated potassium (Kv), and the most studied ion channel receptor, nicotinic acetylcholine receptor (nAChR), in different animal species. In turn, conservation of these examples between snails and humans could suggest potential drug development of related conotoxins.
2. Results
2.1. Summary of the Previously-Reported Transcriptome Sequencing and Achieved Data
Previously, we identified a total of 215 conotoxin transcripts from the Chinese tubular cone snail using five transcriptomes combined with one expressed sequence tag (EST) sequencing [13]. Here, we chose four transcriptome datasets (using the same Illumina sequencing strategy) to explore ion channel based conotoxins and related ion channel receptors/proteins. The four RNA-seq libraries were constructed from three venom ducts (Big, Middle, Small) and one venom bulb (Bulb), in which 4.67, 8.39, 4.58, and 9.77 Gb of paired-end raw reads were generated for each sample respectively. After filtering out low-quality reads, 4.42, 7.95, 4.37, and 9.77 Gb of corresponding clean data were obtained for the followed de novo assembly. All transcriptome assemblies were clustered into a single file for redundancy elimination. The final merged assembly contains a total of 300,069 unigenes with an N50 length of 554 bp and a mean length of 428 bp. Please see more details of the transcriptome assemblies in Table 2.
2.2. Annotation of Unigenes
To annotate the assembled unigenes, diverse public protein databases were applied in this study, including Nr, Nt, Swiss-Prot [53], Kyoto Encyclopedia of Genes and Genomes (KEGG) [54], Clusters of Orthologous Groups (COG) [55] and Gene Ontology (GO) [56]. Among the achieved 300,069 unigenes, 51,412 (17.13%), 43,885 (14.62%) and 37,392 (12.46%) have matches in the Nr, Nt, and Swiss-Prot databases, respectively; 31,682 (10.56%), 13,981 (4.66%), and 19,276 (6.42%) have hits in the KEGG, COG, and GO databases, respectively. In total, there are 76,317 (25.43%) unigenes functionally annotated on basis of the searched databases (Table S1).
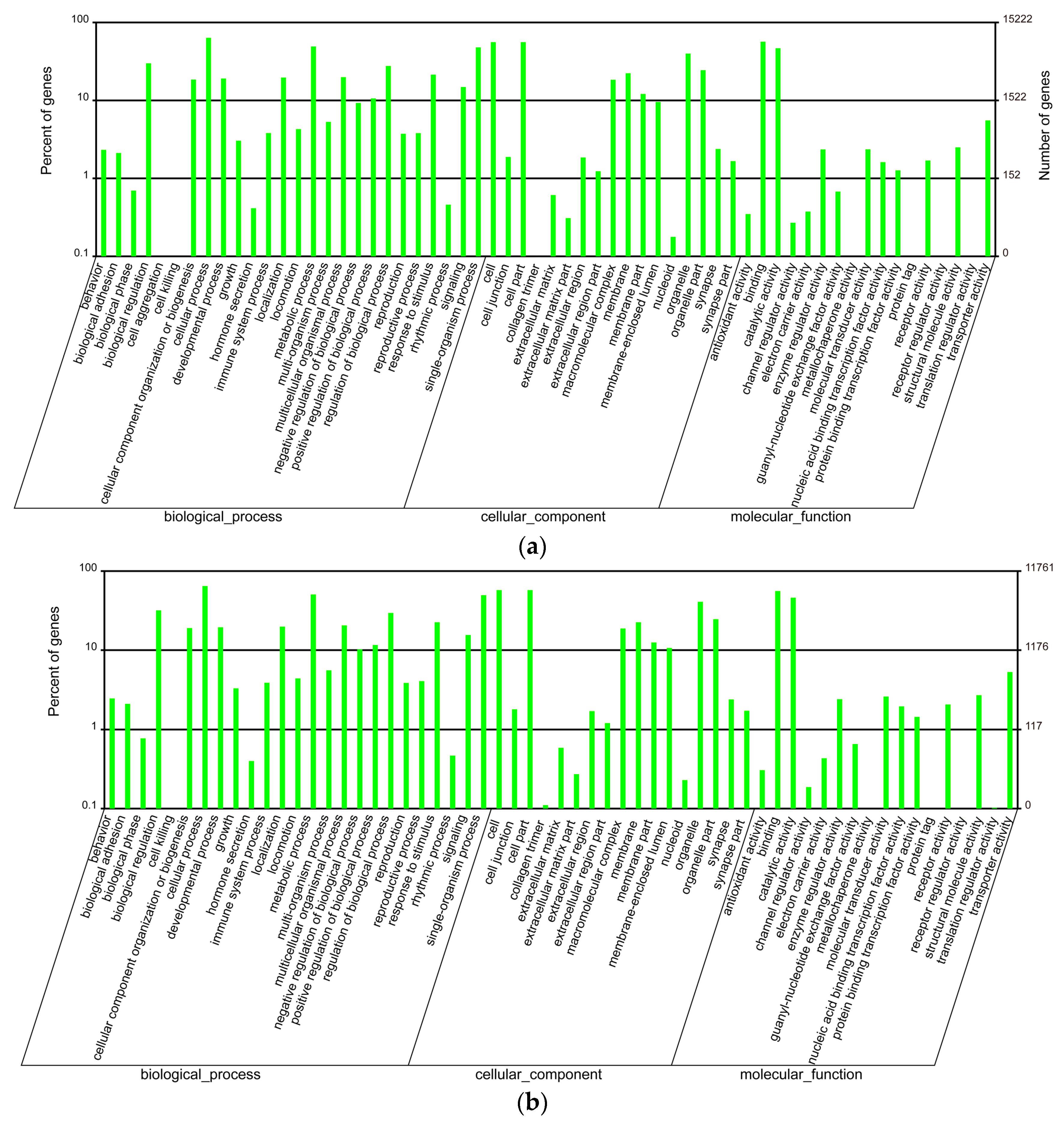
GO enrichment analysis was performed to classify gene functions of these unigenes. Unigenes in each tissue were selected from the merged assembly based on the corresponding clustering results and gene IDs, and then related annotations were applied for GO enrichment analysis. We found that unigenes in the venom duct were enriched into 60 GO classes, in which cellular process, binding, cell and cell part are the top four enriched classes, with 9679, 8621, 8512, and 8511 annotated unigenes, respectively (Figure 1a, Table S2). Interestingly, only one unigene (CL3594.Contig2_All) was classified into the cell aggregation group; while in the venom bulb, annotated unigenes were enriched into 59 GO classes with missing of the cell aggregation term. Likewise, cellular process, cell, cell part, and binding are also the top four enriched terms, although they are slightly different in the order, with 7613, 6764, 6763, and 6579 annotated unigenes in each term respectively (Figure 1b, Table S3). Furthermore, we carried out GO enrichment analysis to classify the achieved conotoxins. In all the reported 215 conotoxins from six datasets [13], 172 conotoxins were identified in the present work due to examination of only four transcriptome datasets (big, middle, small, and bulb). These genes were enriched into 36 GO terms, in which cellular process and metabolic process are the top two with 24 and 22 annotated conotoxins, respectively (Table S4).
2.3. Top Highly-Transcribed Channel and Ion Channel Genes in the Venom Duct and the Venom Bulb
Values of reads per kilobase transcriptome per million mapped reads (RPKM) were calculated to illustrate gene transcription levels in each sample (see more details in Table S1). The lack of specific ion channel classification in the COG, GO and KEGG databases urged us to search with the alternative keywords “ion channel” and “channel” against all the annotation results in Figure 1. Both of the keywords have their limitations since “ion channel” narrows the findings by eliminating genes with synonyms or much more sophisticated annotation (such as “ion transport” or “cation-selective channel”), whereas “channel” includes irrelevant genes such as “water channel protein”. Therefore, both keywords were used to obtain a brief and balanced result. Finally, 365 and 1066 hits by searching “ion channel” and “channel” (Tables S5 and S6) were obtained. The searching results were sorted by RPKM values from the highest to the lowest. Top 20 (ion channel) and 50 (channel) highly transcribed genes in each sample (Big, Middle, Small and Bulb) were picked out for comparison.
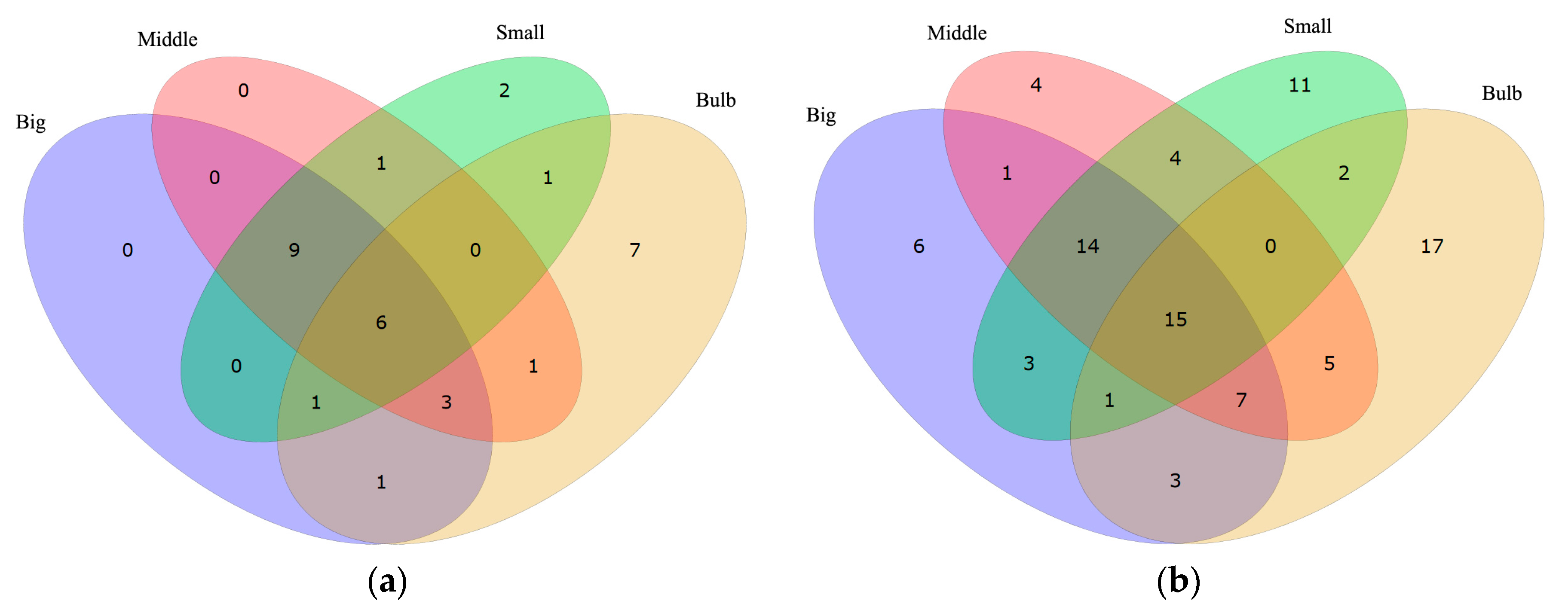
This survey showed that most of the top highly transcribed genes, whether tagged with “ion channel” or “channel”, are the same among Big, Middle and Small samples, while Bulb has other different genes with the highest RPKM values (Figure 2 and Table 3). These data are consistent with the different functions between the venom duct and the venom bulb [13]. In the top 20 “ion channel” genes, the number of individual-specific genes are 0, 0, 2 and 7 in Big, Middle, Small and Bulb datasets, respectively. Similar trend was observed in the top 50 “channel” genes. It seems that the “ion channel” and “channel” proteins transcribed differentially in various tissues. Based on the differential expression strategy of conotoxins in the venom duct and the venom bulb, we agree that cells in the venom duct (the dominant resource of endogenous conotoxins) may have evolved to form special ion channels for various roles of ion channel-based conotoxins.
2.4. Identified Conotoxins and Their Predicted Activities
Previously, we observed that most of the 215 identified conotoxins were synthesized in the venom duct while only few were identified in the venom bulb and, interestingly, conotoxins in the bulb were also transcribed at very low levels [13]. Here, we further examined those conotoxins identified in the four transcriptomes and conducted a detailed survey of their predicted activities (on the basis of distinct cysteine frameworks and gene superfamilies [29]). The top 20 highly-transcribed conotoxins in each specimen are summarized in Table 3. Interestingly, we predicted that in all the four transcriptome datasets, more than half (big: 10; middle: 11; small: 12; bulb: 12) of the top 20 conotoxins belong to the I2-superfamily and are labelled with κ activity (inhibiting voltage-gated potassium channels, VGPCs) [57,58]. The second superfamily in the top line is putative μ-conotoxins (big: 9; middle: 11; small: 11; bulb: 10), with the ability to block voltage-gated sodium channels (VGSCs) [59,60,61]. Only one or two ρ-, ε-, and τ-conotoxins were found in the top 20s of the big, middle, and small transcriptomes, while none of these families were identified in the top 20s of the bulb.
2.5. Ion Channel-Related Proteins and Homologous Analysis
Conotoxins with their diverse activities are proved to target a wide range of ion channels and receptors [62,63,64,65]. This feature has attracted great attentions from researchers to investigate their potential roles and functions for treatment of human diseases [66,67,68]. Here, we picked out the voltage-gated potassium (Kv) ion channels, which interact with the most abundant group (κ-conotoxins) in the top 20s of Table 3, and the widely studied nicotinic acetylcholine receptors (nAChRs) for further evolutionary analysis.
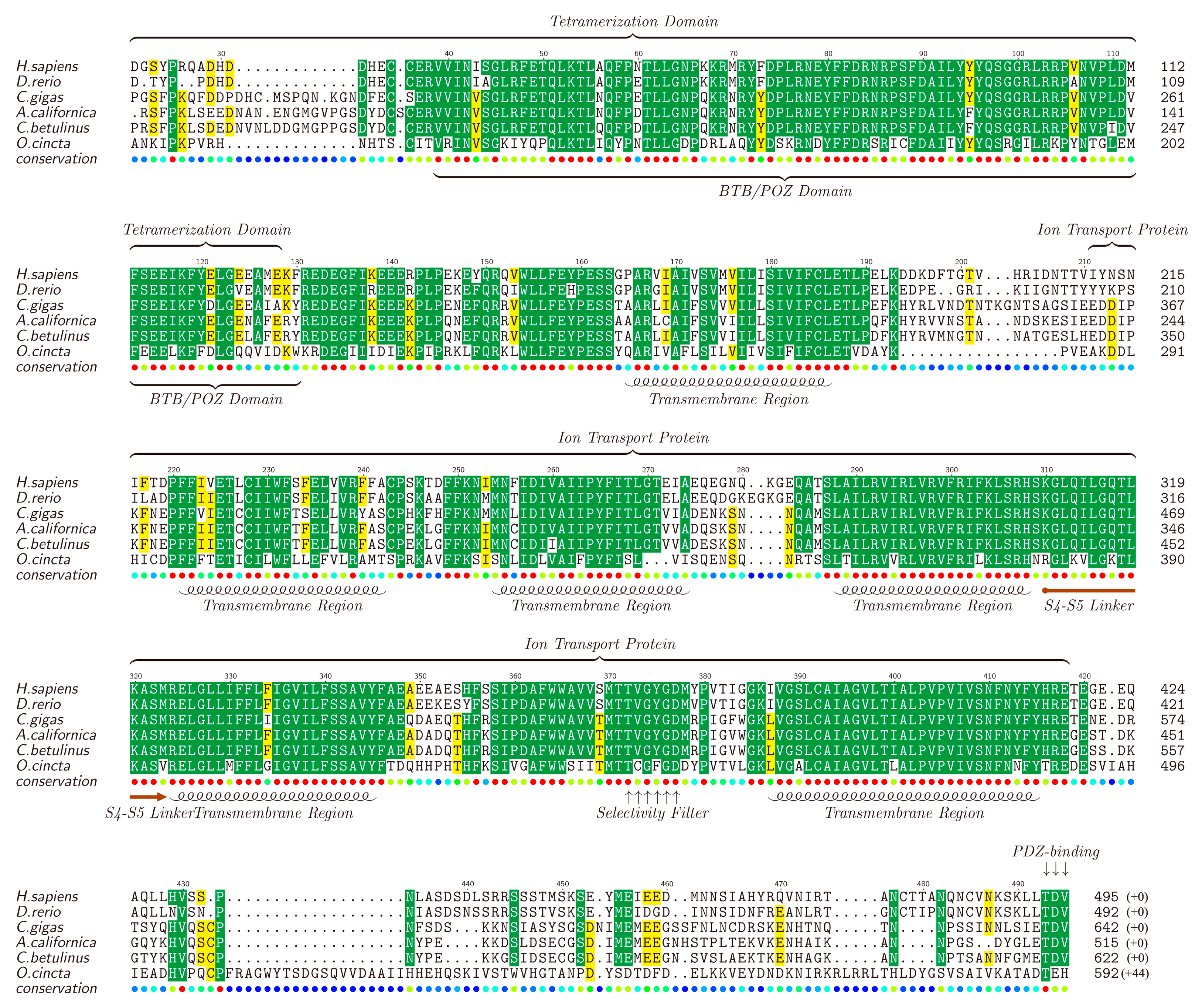
Kv channels, forming the most diverse group among all types of potassium ion channels, are divided into 12 families (Kv1 ~ Kv12) [69,70]. It is reported that Kv proteins play important regulatory roles and participate in various cellular processes including cell differentiation [71], cell proliferation [72], cell apoptosis [73], functioning of excitable cells [74], and so on. Different types of Kv channel members were identified in the four transcriptomes, including Kv1.1, Kv1.4, Kv2.1, Kv3.1 and Kv5.1, with full or partial sequences. The full-length Kv1.1 was picked out as a representative by chance, translated into the protein sequence, submitted to NCBI (MF179123) and aligned against Kv1.1 proteins of other animals (downloaded from NCBI). The lengths for different Kv1.1 proteins range from 492 to 642 amino acids (see Figure 3).
The results showed that, consisting of about 200 amino acids, the ion transport protein region of the Kv1.1, especially all the functional domains, including five transmembrane helices (TMs), S4S5 linker and selectivity filter, tend to be conserved in all the examined animals. Usually, these Kv proteins contain six TMs [75], and the remaining one TM (not included in the ion transport protein region) is also conserved. Furthermore, the tetramerization domain (T1) is somehow variable in the first 30–40 amino acids while the followed BTB (broad-complex, tramtrack and bric-à-brac) or POZ (Pox virus and zinc finger) domain (about 75 amino acids) is highly conserved. The non-conservative segment of the last 50–60 amino acids of Kv1.1 also contains a conserved PDZ-binding site. Existence of the highly-conservative functional domains in these Kv1.1 proteins among different animals suggests similar effects of the putative κ-conotoxins in various species.
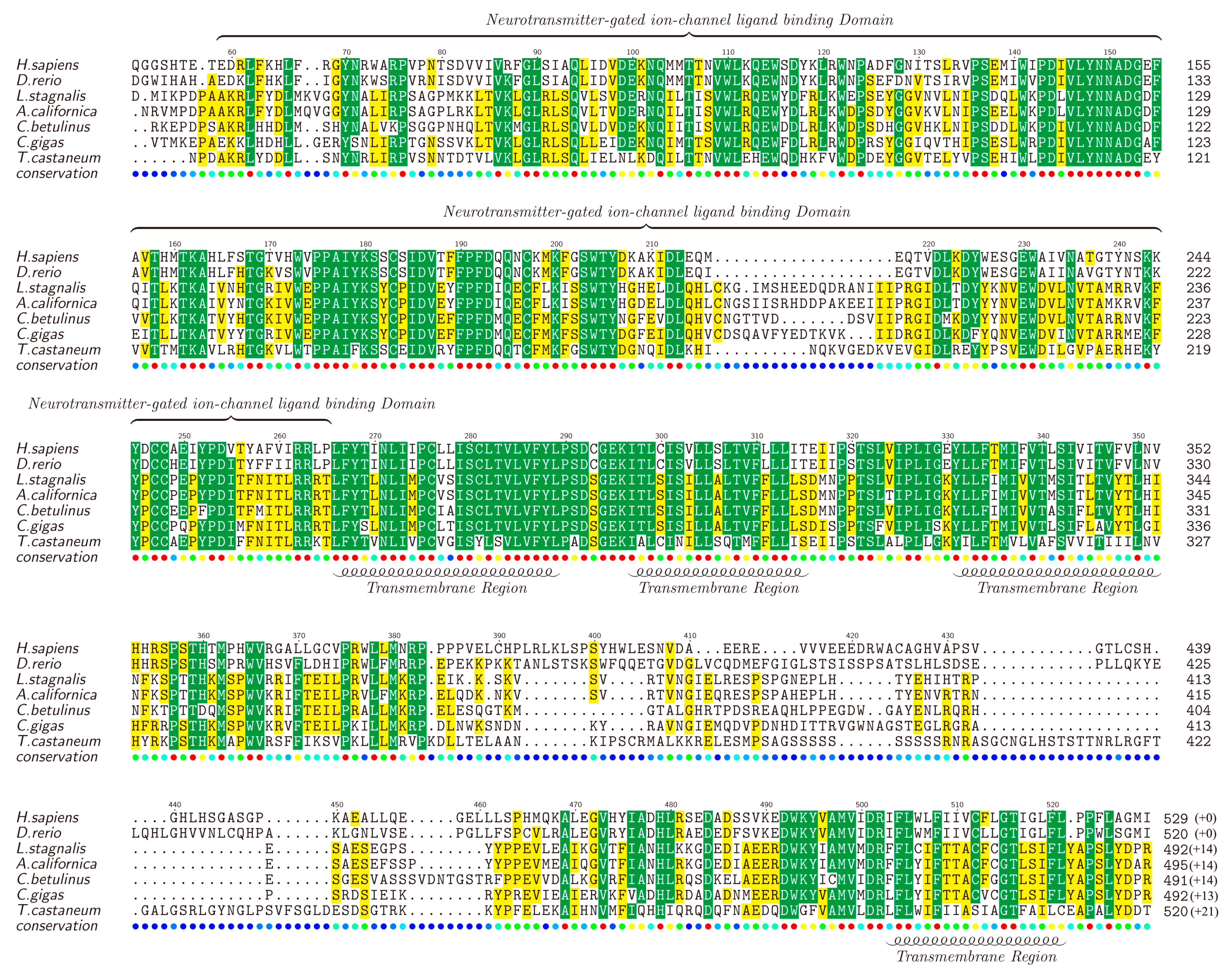
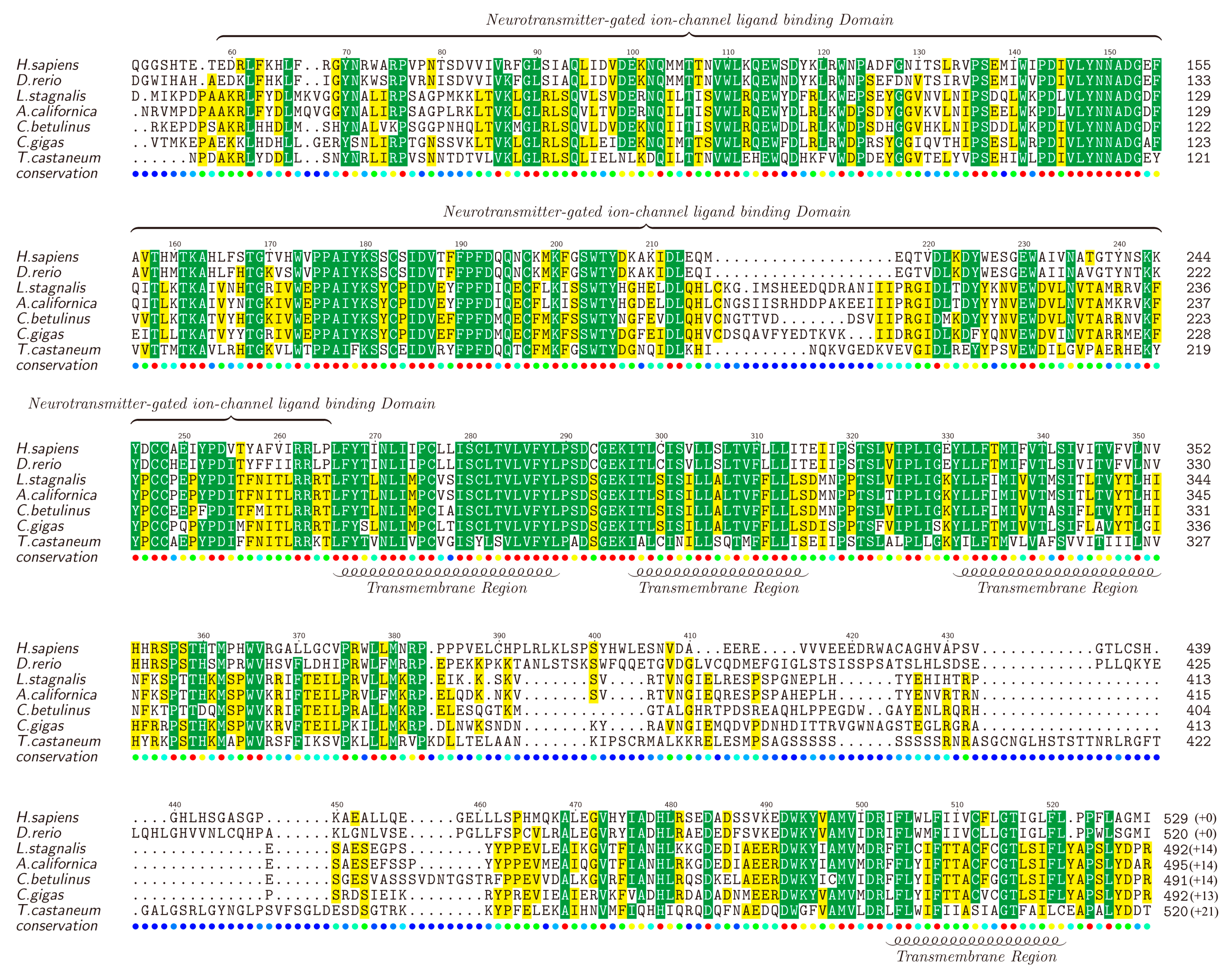
nAChRs are a family of receptors involved in modulation of neurotransmission in the central and peripheral nervous system [76,77,78], and have been reported to be associated with diseases such as schizophrenia, Alzheimer’s and Parkinson’s [79,80]. There are many nAChR subtypes, but generally they can be classified into two main groups, the homopentamers (e.g., α7) and the heteropentamers (e.g., α3β4) [81,82]. We also identified various nAChR genes in the examined transcriptome datasets, including α1, α2, α4, α5, α7, α9, α10 and β3. Likewise, the full-length α2-nAChR was selected as a representative by chance, translated into the protein sequence, submitted to NCBI (MF179124) and aligned against α2-nAChRs of other animals (downloaded from the NCBI). The lengths of different α2-nAChRs range from 505 to 541 amino acids (Figure 4).
Similar to the Kv1.1, high similarities were observed in the TMs of α2-nAChRs. It is reported that α2-nAChRs contain four TMs and TM4 is the least conserved [82], which is also verified by us in this study. Furthermore, TM1, TM2, and TM3 are found to be located closely to each other, while there is a long highly-variable loop between TM3 and TM4. In addition, the neurotransmitter-gated ion channel ligand binding domain (LBD), with a length of about 210 amino acids, is conserved in most areas but not in the segment of 210–220 region. Different from the Kv1.1, α2-nAChR seems to be less conserved among the examined animals, but the functional domains still present a high homology, indicating that α-conotoxin activity may partially influence nervous systems of a wide range of diverse species through regulation of the conserved α2-nAChR.
3. Discussion
Combination of transcriptome sequencing and the subsequent bioinformatics analysis has become a powerful way to discover and predict novel conotoxins from cone snails [14,83,84,85]. Here, we illustrated the differences of ion channel based conotoxins and related ion channel proteins/receptors in C. betulinus, based on our previously reported transcriptomic datasets from both the venom ducts (big, middle, small) and the venom bulb (bulb) [13].
In this study, we regenerated a merged transcriptome assembly containing a total of 300,069 unigenes with an N50 length of 554 bp, and 25.43% of them were functionally annotated according to the public Nr, Nt, Swiss-Prot, KEGG, COG, and GO databases. The GO enrichment analysis of the venom bulb and the venom duct demonstrated similar results of the enriched classes and the distribution number of genes enriched in each term, with cellular process, cell, cell part, and binding to be the top four enriched groups. While all 172 conotoxins, reconfirmed in these transcriptomes, were enriched into 36 GO terms, with cellular process and metabolic process to be the top two enriched ones. These similarities and differences provide insights into these unique conotoxins among all the transcribed genes.
The “ion channel” and “channel” genes transcribed differentially in various samples. In both top 20s and top 50s, the bulb contains the most unique tagged genes, but the ducts share few channel proteins, suggesting differences of ion channels existing in the venom duct (the major tissue to encode conotoxin precursors [85,86]) and the venom bulb (to propel venoms out of the duct [87]). It is estimated that special channels in the venom duct may participate in the protective roles of conotoxins, while the venom bulb functions majorly as the aurilave to discharge conotoxins into the prey [5,14]. Therefore, the gene expression differences are consistent with the function differences.
Among the top 20 highly-transcribed conotoxins in the venom duct and the venom bulb, κ-conotoxins are predicted to be the most common in all samples, followed by putative μ-conotoxins. It seems that these putative κ-conotoxins may have the highest total concentration in the venom mixture of the studied cone snail. Therefore, we extracted the Kv1.1 out from the transcriptome datasets, as well as the most examined α2-nAChR, to act as representatives for further homologous analysis. We observed that all TMs in both Kv1.1 and α2-nAChR are conserved. T1 domain in Kv1.1 is somehow variable in the first 30–40 amino acids, but highly-conserved in the followed BTB/POZ domain, which mediates homomeric/heteromeric dimerization [88]. α2-nAChR contains a conserved neurotransmitter-gated ion channel LBD, which plays important role in the binding of ligands [89], but the TM3–TM4 loop is highly variable. Some studies have shown that this loop forms the largest part of the intracellular domain (ICD) and has special activities [90,91]. The conservation of the neurotransmitter-gated ion channel LBD domain in all species makes the binding of α-conotoxin to α2-nAChRs (from different species) possible, while the variable ICD makes various reactions in cells possible. These interesting data suggest possible ion channel-based toxic pathways for the majority of conotoxins (from cone snails) to be dangerous to human beings (for conservation of ion channels), which in turn proposes a potential hope for development of novel conotoxin-based marine drugs for treatment of ion channel associated human diseases.
4. Materials and Methods
4.1. Assembly, Annotation, and RPKM Calculation
The generated transcriptome raw data from our previous report [13] (NCBI SRS1009725, SRS1009726, SRS1009728, and SRS1009729) were filtered with SOAPnuke [92] to obtain high-quality clean reads. Reads with adaptors, more than 10% of non-sequenced bases and more than 50% of low-quality bases (base quality score ≤ 10) were discarded. The remaining clean data were then separately de novo assembled into contigs and unigenes with an optimized k-mer length of 25 using Trinity software (v2.1.1) [93]. TGICL (v2.0) [94] was used to eliminate the redundancy in each assembly. The final assembly was submitted for functional annotation based on diverse protein databases including Nr, Nt, Swiss-Prot, KEGG, and COG using BLASTX with the threshold of E-value ≤ 10−5. Blast2GO software (v2.8) [95] was used to obtain GO annotation with the Nr BLAST results, and WEGO software [96] was used to achieve the GO classification thereafter. RPKM values of the unigenes were calculated using the Cuffdiff package (FDR < 0.05) of Cufflink software (v2.1.1) [97].
4.2. Prediction and Identification of Ion Channel-Related Proteins/Receptors
We applied two methods to predict and identify interested ion channel related proteins or receptors from the four transcriptome datasets. The first method is to dig the potential proteins out according to the annotation results. Functional annotation results of diverse databases were merged together according to the gene IDs. Candidate ion channel-related genes were picked out from the merged data with the commonly used names (both abbreviation and full names). Subsequently, the sequences of those selected genes were submitted to NCBI using the tBLASTp tool for further verification. The correctly annotated genes were translated into peptide sequences.
Another method is based on homology search with those reported ion channel related proteins or receptors. We downloaded the interested sequences from the NCBI Protein database. Using these sequences as the queries and our assembled transcriptomes as the local database, we applied BLASTX with the threshold of E-value ≤ 10−5 to run the queries against the database. Likewise, the best hits of unigenes were submitted to NCBI using tBLASTp tool for further verification and subsequently translated into peptide sequences.
4.3. Alignment and Homology of Ion Channel Related Proteins/Receptors
The obtained full-length ion channel related proteins/receptors, whether predicted by us or downloaded from the NCBI (Table 4), were aligned and sorted by MAFFT software (v7.037) [98] based on their identity. Protein alignments were shaded in different colours by the TEXshade package [99]. Spatial structures and functional domains of each protein were presented according to the identification of that protein in NCBI Protein database combined with the PDB [100].
5. Conclusions
This is the first report of ion channel-based conotoxins and related ion channel proteins/receptors in the venomous C. betulinus. Similar GO enrichment results were proved in the venom duct and the venom bulb, but different channel and ion channel proteins were identified between the two different tissues. The most common conotoxins with highest transcription levels in the transcriptome datasets are predicted to be κ-conotoxins, followed by putative μ-conotoxins. The ion channel-related protein/receptor, Kv1.1 and α2-nAChR, are conserved in their functional domains, suggesting similar ion channel-based effects of conotoxins in various animal species including human beings, thus making conotoxins a valuable resource for the discovery and development of novel marine drugs to treat ion channel associated human diseases.
Supplementary Materials
The following documents are available online at www.mdpi.com/1660-3397/15/7/228/s1. Table S1. Annotation of all the unique genes and their RPKM values; Table S2. GO enrichment of genes from the venom duct; Table S3. GO enrichment of genes from the venom bulb; Table S4. GO enrichment of conotoxins identified from the transcriptomes; Table S5. Annotated genes tagged with “ion channel” and their RPKM values; Table S6. Annotated genes tagged with “channel” and their RPKM values.
Acknowledgments
This work was supported by International Cooperation Project of Shenzhen Science and Technology (no. GJHZ20160229173052805), Special Project on the Regional Development of Shenzhen Dapeng New District (no. KY20150207), Shenzhen Science and Technology Program (no. GJHS20160331150703934), and Zhenjiang Leading Talent Program for Innovation and Entrepreneurship.
Author Contributions
Q.S. conceived and designed the experiments and revised the manuscript. Y.H. and C.P. analysed the data and wrote the paper. Y.Y. and B.G. contributed to the figures and tables.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kohn, A.J. The ecology of Conus in Hawaii. Ecol. Monogr. 1959, 29, 47–90. [Google Scholar] [CrossRef]
- Puillandre, N.; Bouchet, P.; Duda, T.F.; Kauferstein, S.; Kohn, A.J.; Olivera, B.M.; Watkins, M.; Meyer, C. Molecular phylogeny and evolution of the cone snails (Gastropoda, Conoidea). Mol. Phylogenet. Evol. 2014, 78, 290–303. [Google Scholar] [CrossRef] [PubMed]
- Duda, T.F., Jr.; Kohn, A.J.; Palumbi, S.R. Origins of diverse feeding ecologies within Conus, a genus of venomous marine gastropods. Biol. J. Linn. Soc. 2001, 73, 391–409. [Google Scholar] [CrossRef]
- Duda, T.F., Jr.; Kohn, A.J. Species-level phylogeography and evolutionary history of the hyperdiverse marine gastropod genus Conus. Mol. Phylogenet. Evol. 2005, 34, 257–272. [Google Scholar] [CrossRef] [PubMed]
- Olivera, B.M. Conus venom peptides, receptor and ion channel targets and drug design: 50 million years of neuropharmacology. Mol. Biol. Cell 1997, 8, 2101–2109. [Google Scholar] [CrossRef] [PubMed]
- Bouchet, P.; Kantor, Y.I.; Sysoev, A.; Puillandre, N. A new operational classification of the conoidea (Gastropoda). J. Molluscan Stud. 2011, 77, 273–308. [Google Scholar] [CrossRef]
- Modica, M.V.; Holford, M. The Neogastropoda: Evolutionary innovations of predatory marine snails with remarkable pharmacological potential. In Evolutionary Biology—Concepts, Molecular and Morphological Evolution; Pontarotti, P., Ed.; Springer: Heidelberg, Germany, 2010; pp. 249–270. [Google Scholar]
- Prashanth, J.R.; Dutertre, S.; Jin, A.H.; Lavergne, V.; Hamilton, B.; Cardoso, F.C.; Griffin, J.; Venter, D.J.; Alewood, P.F.; Lewis, R.J. The role of defensive ecological interactions in the evolution of conotoxins. Mol. Ecol. 2016, 25, 598–615. [Google Scholar] [CrossRef] [PubMed]
- Dutertre, S.; Jin, A.H.; Vetter, I.; Hamilton, B.; Sunagar, K.; Lavergne, V.; Dutertre, V.; Fry, B.G.; Antunes, A.; Venter, D.J.; et al. Evolution of separate predation- and defence-evoked venoms in carnivorous cone snails. Nat. Commun. 2014, 5, 3521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Violette, A.; Biass, D.; Dutertre, S.; Koua, D.; Piquemal, D.; Pierrat, F.; Stocklin, R.; Favreau, P. Large-scale discovery of conopeptides and conoproteins in the injectable venom of a fish-hunting cone snail using a combined proteomic and transcriptomic approach. J. Proteom. 2012, 75, 5215–5225. [Google Scholar] [CrossRef] [PubMed]
- Safavi-Hemami, H.; Siero, W.A.; Gorasia, D.G.; Young, N.D.; Macmillan, D.; Williamson, N.A.; Purcell, A.W. Specialisation of the venom gland proteome in predatory cone snails reveals functional diversification of the conotoxin biosynthetic pathway. J. Proteome Res. 2011, 10, 3904–3919. [Google Scholar] [CrossRef] [PubMed]
- Endean, R.; Duchemin, C. The venom apparatus of Conus magus. Toxicon 1967, 4, 275–284. [Google Scholar] [CrossRef]
- Peng, C.; Yao, G.; Gao, B.; Fan, C.; Bian, C.; Wang, J.; Cao, Y.; Wen, B.; Zhu, Y.; Ruan, Z.; et al. High-throughput identification of novel conotoxins from the chinese tubular cone snail (Conus betulinus) by multitranscriptome sequencing. Gigascience 2016, 5, 17. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, J.M.; Jones, R.M. Cone venom—From accidental stings to deliberate injection. Toxicon 2001, 39, 1447–1451. [Google Scholar] [CrossRef]
- Dutertre, S.; Jin, A.H.; Kaas, Q.; Jones, A.; Alewood, P.F.; Lewis, R.J. Deep venomics reveals the mechanism for expanded peptide diversity in cone snail venom. Mol. Cell Proteom. 2013, 12, 312–329. [Google Scholar] [CrossRef] [PubMed]
- Barghi, N.; Concepcion, G.P.; Olivera, B.M.; Lluisma, A.O. Comparison of the Venom Peptides and Their Expression in Closely Related Conus Species: Insights into Adaptive Post-speciation Evolution of Conus Exogenomes. Genome Biol. Evol. 2015, 7, 1797–1814. [Google Scholar] [CrossRef] [PubMed]
- Himaya, S.W.A.; Jin, A.H.; Dutertre, S.; Giacomotto, J.; Mohialdeen, H.; Vetter, I.; Alewood, P.F.; Lewis, R.J. Comparative Venomics Reveals the Complex Prey Capture Strategy of the Piscivorous Cone Snail Conus catus. J. Proteome Res. 2015, 14, 4372–4381. [Google Scholar] [CrossRef] [PubMed]
- Lavergne, V.; Harliwong, I.; Jones, A.; Miller, D.; Taft, R.J.; Alewood, P.F. Optimized deep-targeted proteotranscriptomic profiling reveals unexplored Conus toxin diversity and novel cysteine frameworks. Proc. Natl. Acad. Sci. USA 2015, 112, E3782–E3791. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zheng, Y.; Tang, H. Identifying the types of ion channel-targeted conotoxins by incorporating new properties of residues into pseudo amino acid composition. BioMed Res. Int. 2016, 2016, 3981478. [Google Scholar] [CrossRef] [PubMed]
- Woodward, S.R.; Cruz, L.J.; Olivera, B.M.; Hillyard, D.R. Constant and hypervariable regions in conotoxin propeptides. EMBO J. 1990, 9, 1015–1020. [Google Scholar] [PubMed]
- Olivera, B.M.; Walker, C.; Cartier, G.E.; Hooper, D.; Santos, A.D.; Schoenfeld, R.; Shetty, R.; Watkins, M.; Bandyopadhyay, P.; Hillyard, D.R. Speciation of cone snails and interspecific hyperdivergence of their venom peptides. Potential evolutionary significance of introns. Ann. N. Y. Acad. Sci. 1999, 870, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Lebbe, E.K.; Tytgat, J. In the picture: Disulfide-poor conopeptides, a class of pharmacologically interesting compounds. J. Venom. Anim. Toxins Incl. Trop. Dis. 2016, 22, 30. [Google Scholar] [CrossRef] [PubMed]
- Puillandre, N.; Koua, D.; Favreau, P.; Olivera, B.M.; Stocklin, R. Molecular phylogeny, classification and evolution of conopeptides. J. Mol. Biol. 2012, 74, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Vetter, I.; Lewis, R.J. Therapeutic potential of cone snail venom peptides (conopeptides). Curr. Top. Med. Chem. 2012, 12, 1546–1552. [Google Scholar] [CrossRef] [PubMed]
- Akondi, K.B.; Muttenthaler, M.; Dutertre, S.; Kaas, Q.; Craik, D.J.; Lewis, R.J.; Alewood, P.F. Discovery, synthesis, and structure-activity relationships of conotoxins. Chem. Rev. 2014, 114, 5815–5847. [Google Scholar] [CrossRef] [PubMed]
- Espiritu, D.J.; Watkins, M.; Dia-Monje, V.; Cartier, G.E.; Cruz, L.J.; Olivera, B.M. Venomous cone snails: Molecular phylogeny and the generation of toxin diversity. Toxicon 2001, 39, 1899–1916. [Google Scholar] [CrossRef]
- Lu, A.; Yang, L.; Xu, S.; Wang, C. Various conotoxin diversifications revealed by a venomic study of Conus flavidus. Mol. Cell Proteom. 2014, 13, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Kaas, Q.; Westermann, J.C.; Craik, D.J. Conopeptide characterization and classifications: An analysis using ConoServer. Toxicon 2010, 55, 1491–1509. [Google Scholar] [CrossRef] [PubMed]
- Kaas, Q.; Yu, R.; Jin, A.H.; Dutertre, S.; Craik, D.J. ConoServer: Updated content, knowledge, and discovery tools in the conopeptide database. Nucleic Acids Res. 2012, 40, D325–D330. [Google Scholar] [CrossRef] [PubMed]
- Biggs, J.S.; Watkins, M.; Puillandre, N.; Ownby, J.P.; Lopez-Vera, E.; Christensen, S.; Moreno, K.J.; Bernaldez, J.; Licea-Navarro, A.; Corneli, P.S.; et al. Evolution of Conus peptide toxins: Analysis of Conus californicus Reeve, 1844. Mol. Phylogenet. Evol. 2010, 56, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kancherla, A.K.; Meesala, S.; Jorwal, P.; Palanisamy, R.; Sikdar, S.K.; Sarma, S.P. A Disulfide Stabilized β-Sandwich Defines the Structure of a New Cysteine Framework M-Superfamily Conotoxin. ACS Chem. Biol. 2015, 10, 1847–1860. [Google Scholar] [CrossRef] [PubMed]
- Bernáldez, J.; Román-González, S.A.; Martínez, O.; Jiménez, S.; Vivas, O.; Arenas, I.; Corzo, G.; Arreguín, R.; García, D.E.; Possani, L.D.; et al. A Conus regularis conotoxin with a novel eight-cysteine framework inhibits CaV2.2 channels and displays an anti-nociceptive activity. Mar. Drugs 2013, 11, 1188–1202. [Google Scholar] [CrossRef] [PubMed]
- Olivera, B.M.; Cruz, L.J. Conotoxins, in retrospect. Toxicon 2001, 39, 7–14. [Google Scholar] [CrossRef]
- Dutton, J.L.; Craik, D.J. alpha-Conotoxins: Nicotinic acetylcholine receptor antagonists as pharmacological tools and potential drug leads. Curr. Med. Chem. 2001, 8, 327–344. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J. Conotoxins: Molecular and therapeutic targets. Prog. Mol. Subcell. Biol. 2009, 46, 45–65. [Google Scholar] [PubMed]
- Vincler, M.; McIntosh, J.M. Targeting the alpha9alpha10 nicotinic acetylcholine receptor to treat severe pain. Expert Opin. Ther. Targets 2007, 11, 891–897. [Google Scholar] [CrossRef] [PubMed]
- Twede, V.D.; Miljanich, G.; Olivera, B.M.; Bulaj, G. Neuroprotective and cardioprotective conopeptides: An emerging class of drug leads. Curr. Opin. Drug Discov. Dev. 2009, 12, 231–239. [Google Scholar]
- Clark, R.J.; Jensen, J.; Nevin, S.T.; Callaghan, B.P.; Adams, D.J.; Craik, D.J. The engineering of an orally active conotoxin for the treatment of neuropathic pain. Angew. Chem. Int. Ed. 2010, 49, 6545–6548. [Google Scholar] [CrossRef] [PubMed]
- Miljanich, G.P. Ziconotide: Neuronal calcium channel blocker for treating severe chronic pain. Curr. Med. Chem. 2004, 11, 3029–3040. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J.; Dutertre, S.; Vetter, I.; Christie, M.J. Conus venom peptide pharmacology. Pharmacol. Rev. 2012, 64, 259–298. [Google Scholar] [CrossRef] [PubMed]
- Han, T.S.; Teichert, R.W.; Olivera, B.M.; Bulaj, G. Conus venoms—A rich source of peptide-based therapeutics. Curr. Pharm. Des. 2008, 14, 2462–2479. [Google Scholar] [CrossRef] [PubMed]
- Gray, W.R.; Luque, A.; Olivera, B.M.; Barrett, J.; Cruz, L.J. Peptide toxins from Conus geographus venom. J. Biol. Chem. 1981, 256, 4734–4740. [Google Scholar] [PubMed]
- Fainzilber, M.; Nakamura, T.; Lodder, J.C.; Zlotkin, E.; Kits, K.S.; Burlingame, A.L. gamma-Conotoxin-PnVIIA, a gamma-carboxyglutamate-containing peptide agonist of neuronal pacemaker cation currents. Biochemistry 1998, 37, 1470–1477. [Google Scholar] [CrossRef] [PubMed]
- Fainzilber, M.; Gordon, D.; Hasson, A.; Spira, M.E.; Zlotkin, E. Mollusc-specific toxins from the venom of Conus textile neovicarius. Eur. J. Biochem. 1991, 202, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Rigby, A.C.; Lucas-Meunier, E.; Kalume, D.E.; Czerwiec, E.; Hambe, B.; Dahlqvist, I.; Fossier, P.; Baux, G.; Roepstorff, P.; Baleja, J.D.; et al. A conotoxin from Conus textile with unusual posttranslational modifications reduces presynaptic Ca2+ influx. Proc. Natl. Acad. Sci. USA 1999, 96, 5758–5763. [Google Scholar] [CrossRef] [PubMed]
- Buczek, O.; Wei, D.; Babon, J.J.; Yang, X.; Fiedler, B.; Chen, P.; Yoshikami, D.; Olivera, B.M.; Bulaj, G.; Norton, R.S. Structure and sodium channel activity of an excitatory I1-superfamily conotoxin. Biochemistry 2007, 46, 9929–9940. [Google Scholar] [CrossRef] [PubMed]
- Terlau, H.; Shon, K.J.; Grilley, M.; Stocker, M.; Stühmer, W.; Olivera, B.M. Strategy for rapid immobilization of prey by a fish-hunting marine snail. Nature 1996, 381, 148–151. [Google Scholar] [CrossRef] [PubMed]
- Cruz, L.J.; Gray, W.R.; Olivera, B.M.; Zeikus, R.D.; Kerr, L.; Yoshikami, D.; Moczydlowski, E. Conus geographus toxins that discriminate between neuronal and muscle sodium channels. J. Biol. Chem. 1985, 260, 9280–9288. [Google Scholar] [PubMed]
- Sharpe, I.A.; Gehrmann, J.; Loughnan, M.L.; Thomas, L.; Adams, D.A.; Atkins, A.; Palant, E.; Craik, D.J.; Adams, D.J.; Alewood, P.F.; et al. Two new classes of conopeptides inhibit the alpha1-adrenoceptor and noradrenaline transporter. Nat. Neurosci. 2001, 4, 902–907. [Google Scholar] [CrossRef] [PubMed]
- England, L.J.; Imperial, J.; Jacobsen, R.; Craig, A.G.; Gulyas, J.; Akhtar, M.; Rivier, J.; Julius, D.; Olivera, B.M. Inactivation of a serotonin-gated ion channel by a polypeptide toxin from marine snails. Science 1998, 281, 575–578. [Google Scholar] [CrossRef] [PubMed]
- Petrel, C.; Hocking, H.G.; Reynaud, M.; Upert, G.; Favreau, P.; Biass, D.; Paolini-Bertrand, M.; Peigneur, S.; Tytgat, J.; Gilles, N.; et al. Identification, structural and pharmacological characterization of τ-CnVA, a conopeptide that selectively interacts with somatostatin sst3 receptor. Biochem. Pharmacol. 2013, 85, 1663–1671. [Google Scholar] [CrossRef] [PubMed]
- Kerr, L.M.; Yoshikami, D. A venom peptide with a novel presynaptic blocking action. Nature 1984, 308, 282–284. [Google Scholar] [CrossRef] [PubMed]
- Bairoch, A.; Apweiler, R. The SWISS-PROT protein sequence database and its supplement TrEMBL in 2000. Nucleic Acids Res. 2000, 28, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Tatusov, R.L.; Galperin, M.Y.; Natale, D.A.; Koonin, E.V. The COG database: A tool for genome-scale analysis of protein functions and evolution. Nucleic Acids Res. 2000, 28, 33–36. [Google Scholar] [CrossRef] [PubMed]
- GO-EBI. The Gene Ontology (GO) database and informatics resource. Nucleic Acids Res. 2004, 32, D258–D261. [Google Scholar]
- Moran, O. Molecular simulation of the interaction of kappa-conotoxin-PVIIA with the Shaker potassium channel pore. Eur. Biophys. J. 2001, 30, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Terlau, H.; Olivera, B.M. Conus venoms: A rich source of novel ion channel-targeted peptides. Physiol. Rev. 2004, 84, 41–68. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.M.; Alewood, P.F.; Craik, D.J. Three-dimensional solution structure of mu-conotoxin GIIIB, a specific blocker of skeletal muscle sodium channels. Biochemistry 1996, 35, 8824–8835. [Google Scholar] [CrossRef] [PubMed]
- Halai, R.; Craik, D.J. Conotoxins: Natural product drug leads. Nat. Prod. Rep. 2009, 26, 526–536. [Google Scholar] [CrossRef] [PubMed]
- Green, B.R.; Bulaj, G.; Norton, R.S. Structure and function of μ-conotoxins, peptide-based sodium channel blockers with analgesic activity. Future Med. Chem. 2014, 6, 1677–1698. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J. Conotoxins as selective inhibitors of neuronal ion channels, receptors and transporters. IUBMB Life 2004, 56, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Favreau, P.; Le Gall, F.; Benoit, E.; Molgó, J. A review on conotoxins targeting ion channels and acetylcholine receptors of the vertebrate neuromuscular junction. Acta Physiol. Pharmacol. Ther. Latinoam. 1999, 49, 257–267. [Google Scholar] [PubMed]
- Deuis, J.R.; Dekan, Z.; Inserra, M.C.; Lee, T.H.; Aguilar, M.I.; Craik, D.J.; Lewis, R.J.; Alewood, P.F.; Mobli, M.; Schroeder, C.I.; et al. Development of a μO-Conotoxin Analogue with Improved Lipid Membrane Interactions and Potency for the Analgesic Sodium Channel NaV1.8. J. Biol. Chem. 2016, 291, 11829–11842. [Google Scholar] [CrossRef] [PubMed]
- Green, B.R.; Gajewiak, J.; Chhabra, S.; Skalicky, J.J.; Zhang, M.M.; Rivier, J.E.; Bulaj, G.; Olivera, B.M.; Yoshikami, D.; Norton, R.S. Structural Basis for the Inhibition of Voltage-gated Sodium Channels by Conotoxin μO§-GVIIJ. J. Biol. Chem. 2016, 291, 7205–7220. [Google Scholar] [CrossRef] [PubMed]
- Bellacchio, E. Mechanism of neurotoxicity of prion and Alzheimer’s disease-related proteins: Molecular insights from bioinformatically identified ω-conotoxin-like pharmacophores. Crit. Rev. Eukaryot. Gene Expr. 2013, 23, 355–373. [Google Scholar] [CrossRef] [PubMed]
- Reimers, C.; Lee, C.H.; Kalbacher, H.; Tian, Y.; Hung, C.H.; Schmidt, A.; Prokop, L.; Kauferstein, S.; Mebs, D.; Chen, C.C.; et al. Identification of a cono-RFamide from the venom of Conus textile that targets ASIC3 and enhances muscle pain. Proc. Natl. Acad. Sci. USA 2017, 114, E3507–E3515. [Google Scholar] [CrossRef] [PubMed]
- Jayamanne, A.; Jeong, H.J.; Schroeder, C.I.; Lewis, R.J.; Christie, M.J.; Vaughan, C.W. Spinal actions of ω-conotoxins, CVID, MVIIA and related peptides in a rat neuropathic pain model. Br. J. Pharmocol. 2013, 170, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Gutman, G.A.; Chandy, K.G.; Grissmer, S.; Lazdunski, M.; McKinnon, D.; Pardo, L.A.; Robertson, G.A.; Rudy, B.; Sanguinetti, M.C.; Stühmer, W.; et al. International Union of Pharmacology. LIII. Nomenclature and molecular relationships of voltage-gated potassium channels. Pharmacol. Rev. 2005, 57, 473–508. [Google Scholar] [CrossRef] [PubMed]
- Grizel, A.V.; Glukhov, G.S.; Sokolova, O.S. Mechanisms of Activation of Voltage-Gated Potassium Channels. Acta Nat. 2014, 6, 10–26. [Google Scholar]
- Li, G.R.; Deng, X.L. Functional ion channels in stem cells. World J. Stem Cells 2011, 3, 19–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pardo, L.A. Voltage-Gated Potassium Channels in Cell Proliferation. Physiology 2004, 19, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, S.; Archer, S.L.; Allalunis-Turner, J.; Haromy, A.; Beaulieu, C.; Thompson, R.; Lee, C.T.; Lopaschuk, G.D.; Puttagunta, L.; Bonnet, S.; et al. A mitochondria-K+ channel axis is suppressed in cancer and its normalization promotes apoptosis and inhibits cancer growth. Cancer Cell 2007, 11, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Hoshi, T.; Zagotta, W.N.; Aldrich, R.W. Biophysical and molecular mechanisms of Shaker potassium channel inactivation. Science 1990, 250, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Kuang, Q.; Purhonen, P.; Hebert, H. Structure of potassium channels. Cell Mol. Life Sci. 2015, 72, 3677–3693. [Google Scholar] [CrossRef] [PubMed]
- Paterson, D.; Nordberg, A. Neuronal nicotinic receptors in the human brain. Prog. Neurobiol. 2000, 61, 75–111. [Google Scholar] [CrossRef]
- Dani, J.A.; Bertrand, D. Nicotinic acetylcholine receptors and nicotinic cholinergic mechanisms of the central nervous system. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 699–729. [Google Scholar] [CrossRef] [PubMed]
- Changeux, J.P. The nicotinic acetylcholine receptor: The founding father of the pentameric ligand-gated ion channel superfamily. J. Biol. Chem. 2012, 287, 40207–40215. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, G.K.; Williams, M. Neuronal nicotinic acetylcholine receptors as novel drug targets. J. Pharmacol. Exp. Ther. 2000, 292, 461–467. [Google Scholar] [PubMed]
- Kawasaki, Y.; Freire, E. Finding a better path to drug selectivity. Drug Discov. Today 2011, 16, 985–990. [Google Scholar] [CrossRef] [PubMed]
- Alkondon, M.; Albuquerque, E.X. The nicotinic acetylcholine receptor subtypes and their function in the hippocampus and cerebral cortex. Prog. Brain Res. 2004, 145, 109–120. [Google Scholar] [PubMed]
- Albuquerque, E.X.; Pereira, E.F.R.; Alkondon, M.; Rogers, S.W. Mammalian nicotinic acetylcholine receptors: From structure to function. Physiol. Rev. 2009, 89, 73–120. [Google Scholar] [CrossRef] [PubMed]
- Barghi, N.; Concepcion, G.P.; Olivera, B.M.; Lluisma, A.O. High conopeptide diversity in Conus tribblei revealed through analysis of venom duct transcriptome using two high-throughput sequencing platforms. Mar. Biotechnol. 2015, 17, 81–98. [Google Scholar] [CrossRef] [PubMed]
- Lavergne, V.; Dutertre, S.; Jin, A.H.; Lewis, R.J.; Taft, R.J.; Alewood, P.F. Systematic interrogation of the Conus marmoreus venom duct transcriptome with ConoSorter reveals 158 novel conotoxins and 13 new gene superfamilies. BMC Genom. 2013, 14, 708. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Bandyopadhyay, P.K.; Olivera, B.M.; Yandell, M. Elucidation of the molecular envenomation strategy of the cone snail Conus geographus through transcriptome sequencing of its venom duct. BMC Genom. 2012, 13, 284. [Google Scholar] [CrossRef] [PubMed]
- Marshall, J.; Kelley, W.P.; Rubakhin, S.S.; Bingham, J.P.; Sweedler, J.V.; Gilly, W.F. Anatomical correlates of venom production in Conus californicus. Biol. Bull. 2002, 203, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Safavi-Hemami, H.; Young, N.D.; Williamson, N.A.; Purcell, A.W. Proteomic interrogation of venom delivery in marine cone snails: novel insights into the role of the venom bulb. J. Proteome Res. 2010, 9, 5610–5619. [Google Scholar] [CrossRef] [PubMed]
- Bardwell, V.J.; Treisman, R. The POZ domain: A conserved protein-protein interaction motif. Genes Dev. 1994, 8, 1664–1677. [Google Scholar] [CrossRef] [PubMed]
- Brejc, K.; van Dijk, W.J.; Klaassen, R.V.; Schuurmans, M.; van Der Oost, J.; Smit, A.B.; Sixma, T.K. Crystal structure of an ACh-binding protein reveals the ligand-binding domain of nicotinic receptors. Nature 2001, 411, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Kukhtina, V.; Kottwitz, D.; Strauss, H.; Heise, B.; Chebotareva, N.; Tsetlin, V.; Hucho, F. Intracellular domain of nicotinic acetylcholine receptor: the importance of being unfolded. J. Neurochem. 2006, 97, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Kracun, S.; Harkness, P.C.; Gibb, A.J.; Millar, N.S. Influence of the M3-M4 intracellular domain upon nicotinic acetylcholine receptor assembly, targeting and function. Br. J. Pharmacol. 2008, 153, 1474–1484. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Li, Y.; Kristiansen, K.; Wang, J. SOAP: Short oligonucleotide alignment program. Bioinformatcis 2008, 24, 713–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Trinity: Reconstructing a full-length transcriptome without a genome from RNA-Seq data. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Pertea, G.; Huang, X.; Liang, F.; Antonescu, V.; Sultana, R.; Karamycheva, S.; Lee, Y.; White, J.; Cheung, F.; Parvizi, B.; et al. TIGR Gene Indices clustering tools (TGICL): A software system for fast clustering of large EST datasets. Bioinformatics 2003, 19, 651–652. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Gotz, S.; Garcia-Gomez, J.M.; Terol, J.; Talon, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Fang, L.; Zheng, H.; Zhang, Y.; Chen, J.; Zhang, Z.; Wang, J.; Li, S.; Li, R.; Bolund, L.; et al. WEGO: A web tool for plotting GO annotations. Nucleic Acids Res. 2006, 34, W293–W297. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and abundance estimation from RNA-Seq reveals thousands of new transcripts and switching among isoforms. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Beitz, E. TEXshade: Shading and labeling multiple sequence alignments using LATEX2 epsilon. Bioinformatics 2000, 16, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Parasuraman, S. Protein data bank. J. Pharmacol. Pharmocother. 2012, 3, 351–352. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
GO enrichment of the annotated unigenes in the venom duct (a) and the venom bulb (b).
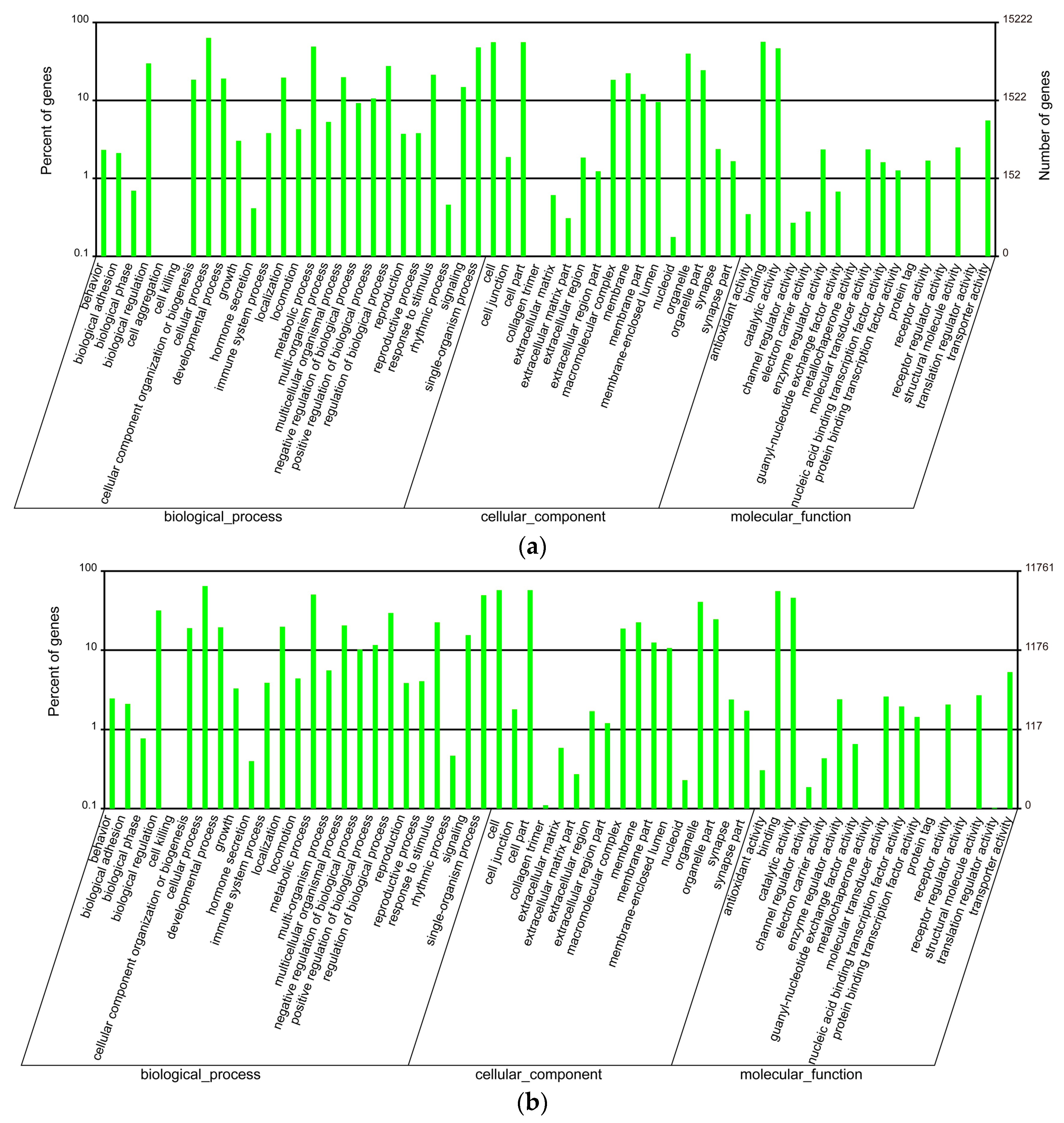
Figure 2.
Comparison of highly-transcribed genes tagged with “ion channel” (a) and “channel” (b) in the venom duct and the venom bulb.
Figure 2.
Comparison of highly-transcribed genes tagged with “ion channel” (a) and “channel” (b) in the venom duct and the venom bulb.
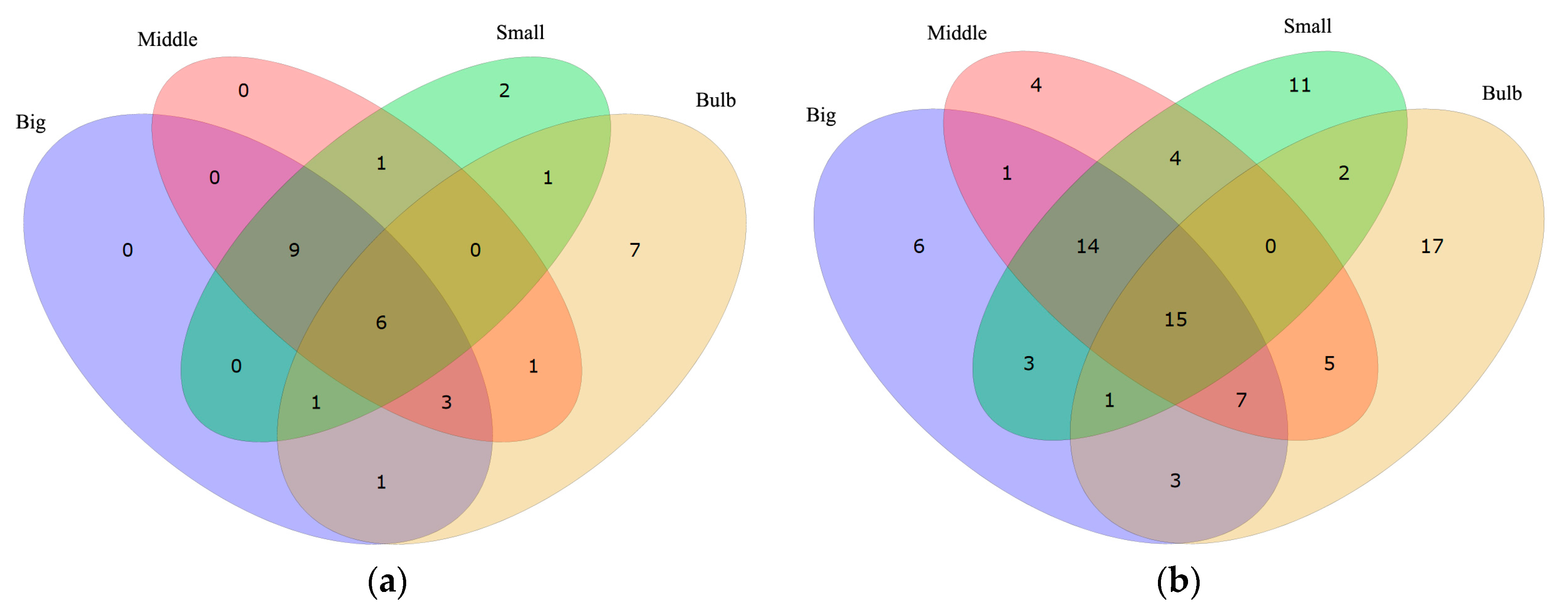
Figure 3.
Alignment of Kv1.1 proteins obtained from different species. Yellow blocks show the areas with sequence identity >50% and green blocks indicate the identity >80%.
Figure 3.
Alignment of Kv1.1 proteins obtained from different species. Yellow blocks show the areas with sequence identity >50% and green blocks indicate the identity >80%.
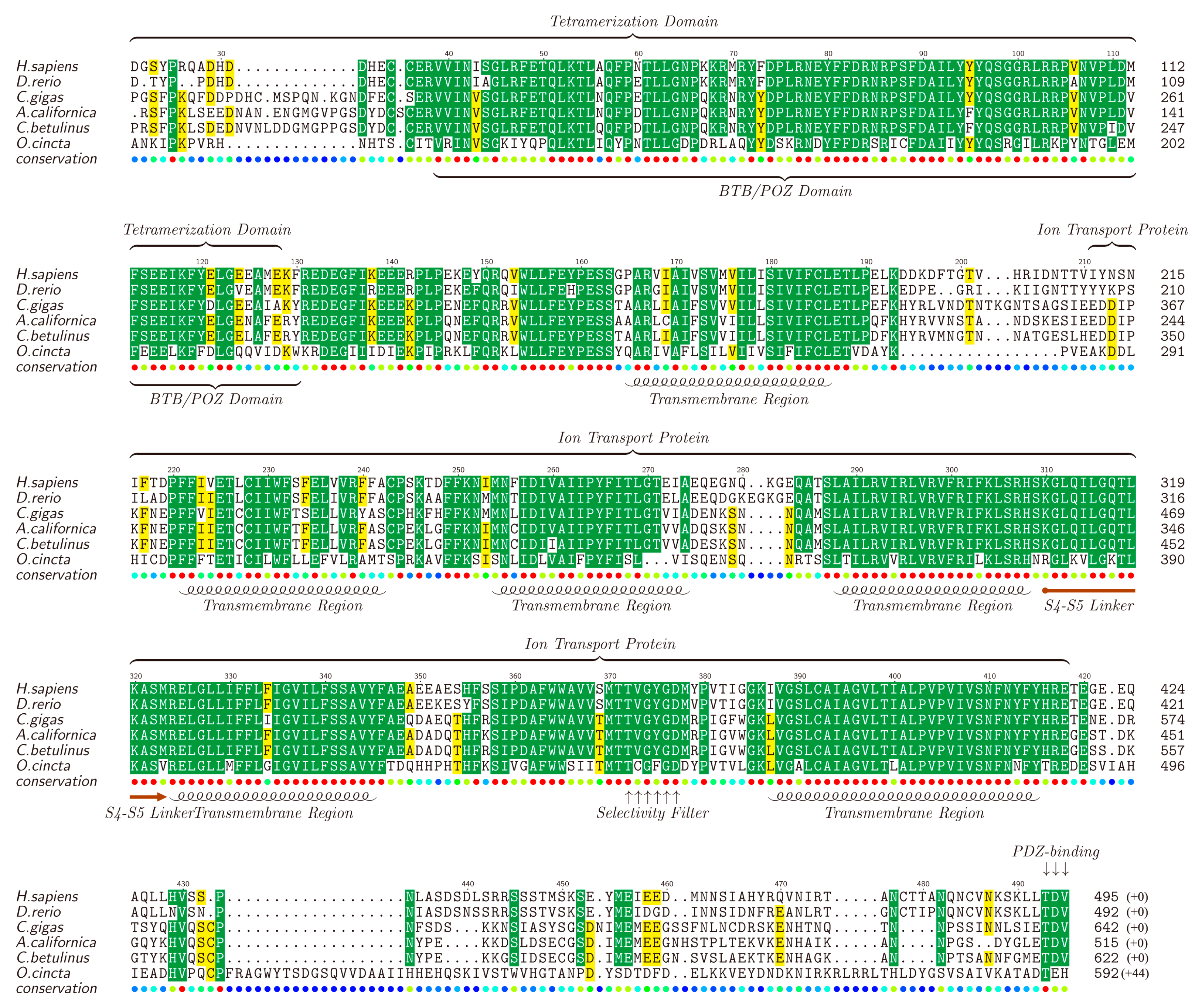
Figure 4.
Alignment of α2-nAChRs obtained from different species. Yellow blocks show the areas with sequence identity >50% and green blocks indicate the identity >80%.
Figure 4.
Alignment of α2-nAChRs obtained from different species. Yellow blocks show the areas with sequence identity >50% and green blocks indicate the identity >80%.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Pharmacological families and related definitions of classified conotoxins [28].
Table 1.
Pharmacological families and related definitions of classified conotoxins [28].
| Family | Definition | Reference |
|---|---|---|
| α (alpha) | Nicotinic acetylcholine receptors (nAChR) | [42] |
| γ (gamma) | Neuronal pacemaker cation currents (inward cation current) | [43] |
| δ (delta) | Voltage-gated Na channels (agonist, delay inactivation) | [44] |
| ε (epsilon) | Presynaptic Ca channels or G protein-coupled presynaptic receptors | [45] |
| ι (iota) | Voltage-gated Na channels (agonist, no delayed inactivation) | [46] |
| κ (kappa) | Voltage-gated K channels (blocker) | [47] |
| μ (mu) | Voltage-gated Na channels (antagonist, blocker) | [48] |
| ρ (rho) | Alpha1-adrenoceptors (GPCR) | [49] |
| σ (sigma) | Serotonin-gated ion channels (5-HT3R) | [50] |
| τ (tau) | Somatostatin receptor | [51] |
| χ (chi) | Neuronal noradrenaline transporter | [49] |
| ω (omega) | Voltage-gated Ca channels (blocker) | [52] |
Table 2.
Summary of the transcriptome assemblies.
| Sample | Venom Duct | Bulb | All | |||
|---|---|---|---|---|---|---|
| Unigene | Big | Middle | Small | |||
| Total Number | 94,026 | 52,387 | 114,057 | 124,004 | 300,069 | |
| Total Length | 37,880,261 | 23,128,493 | 44,918,779 | 67,451,577 | 128,471,163 | |
| Mean Length | 403 | 441 | 394 | 544 | 428 | |
| N50 | 413 | 464 | 398 | 681 | 554 | |
Table 3.
Top 20 highly transcribed conotoxins in the big, middle, small, and bulb datasets.
| Big | Middle | Small | Bulb | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Conotoxin | RPKM | Predicted Activity 1 | Conotoxin | RPKM | Predicted Activity | Conotoxin | RPKM | Predicted Activity | Conotoxin | RPKM | Predicted Activity |
| Bt035 | 84,466.31 | NMDARi 2 | Bt055 | 77,776.43 | κ | Bt035 | 58,816.45 | NMDARi | Bt070 | 179.21 | α, ι, κ, μ |
| Bt057 | 72,563.89 | κ | Bt018 | 71,975.85 | unknown | Bt018 | 32,503.63 | unknown | Bt035 | 143.27 | NMDARi |
| Bt018 | 57,233.21 | unknown | Bt082 | 57,532.96 | unknown | Bt075 | 25,533.62 | α, ι, κ, μ | Bt055 | 106.38 | κ |
| Bt005 | 21,270.26 | α, ρ | Bt035 | 53,262.22 | NMDARi | Bt055 | 20,861.06 | κ | Bt141 | 91.35 | δ, γ, κ, μ, ω |
| Bt082 | 20,710.97 | unknown | Bt213 | 43,598.05 | ε, μ, τ | Bt005 | 19,553.90 | α, ρ | Bt145 | 77.8 | δ, γ, κ, μ, ω |
| Bt055 | 19,799.15 | κ | Bt013 | 38,601.60 | NMDARi | Bt070 | 11,309.30 | α, ι, κ, μ | Bt043 | 56.26 | δ, γ, κ, μ, ω |
| Bt087 | 11,394.34 | α, ι, κ, μ | Bt076 | 27,361.60 | α, ι, κ, μ | Bt043 | 11,144.72 | δ, γ, κ, μ, ω | Bt017 | 51.57 | NMDARi |
| Bt043 | 10,995.23 | δ, γ, κ, μ, ω | Bt072 | 25,964.06 | α, ι, κ, μ | Bt213 | 10,602.62 | ε, μ, τ | Bt041 | 42.9 | unknown |
| Bt200 | 7552.24 | unknown | Bt071 | 22,016.06 | α, ι, κ, μ | Bt111 | 9823.73 | unknown | Bt076 | 34.75 | α, ι, κ, μ |
| Bt044 | 6067.80 | δ, γ, κ, μ, ω | Bt077 | 21,344.69 | α, ι, κ, μ | Bt057 | 9268.87 | κ | Bt005 | 34.05 | α |
| Bt186 | 5760.52 | δ, γ, κ, μ, ω | Bt125 | 18,909.36 | unknown | Bt013 | 8930.31 | NMDARi | Bt018 | 28.91 | unknown |
| Bt213 | 5745.23 | ε, μ, τ | Bt145 | 18,505.50 | δ, γ, κ, μ, ω | Bt210 | 6581.46 | ε, μ, τ | Bt186 | 27.08 | δ, γ, κ, μ, ω |
| Bt141 | 4616.86 | δ, γ, κ, μ, ω | Bt185 | 17,584.41 | δ, γ, κ, μ, ω | Bt081 | 5527.25 | α, ι, κ, μ | Bt054 | 16.35 | κ |
| Bt075 | 4512.67 | α, ι, κ, μ | Bt043 | 14,968.06 | δ, γ, κ, μ, ω | Bt082 | 3706.28 | unknown | Bt044 | 15.82 | δ, γ, κ, μ, ω |
| Bt081 | 4368.21 | α, ι, κ, μ | Bt192 | 12,120.02 | unknown | Bt086 | 3465.02 | α, ι, κ, μ | Bt075 | 15.36 | α, ι, κ, μ |
| Bt100 | 4154.73 | KCb 3 | Bt075 | 11,667.17 | α, ι, κ, μ | Bt058 | 3431.21 | κ | Bt077 | 13.29 | α, ι, κ, μ |
| Bt042 | 4079.54 | unknown | Bt141 | 11,364.97 | δ, γ, κ, μ, ω | Bt087 | 3429.93 | α, ι, κ, μ | Bt150 | 11.54 | δ, γ, κ, μ, ω |
| Bt138 | 3860.72 | δ, γ, κ, μ, ω | Bt136 | 10,248.83 | δ, γ, κ, μ, ω | Bt186 | 3353.26 | δ, γ, κ, μ, ω | Bt100 | 10.06 | KCb |
| Bt041 | 3662.06 | unknown | Bt005 | 9612.27 | α,ρ | Bt072 | 3110.04 | α, ι, κ, μ | Bt048 | 9.73 | ι |
| Bt172 | 3633.23 | γ | Bt040 | 8273.34 | unknown | Bt044 | 2827.06 | δ, γ, κ, μ, ω | Bt020 | 8.01 | NMDARi |
1 The Greek letters denote pharmacological families as conotoxins sharing the same receptor specificities; See more details about related definitions in the ConoServer website [29]; 2 NMDARi: NMDA receptor inhibitor; 3 KCb: potassium channels blocker.
Table 4.
Sequences used for the alignments of ion channel proteins.
| Gene Name | Genbank Accession No. | Species |
|---|---|---|
| Kv1.1 | NP_000208.2 | human (Homo sapiens) |
| XP_005163101.1 | zebrafish (Danio rerio) | |
| NP_001191634.1 | sea hare (Aplysia californica) | |
| MF179123 | cone snail (Conus betulinus) | |
| XP_011413619.1 | oyster (Crassostrea gigas) | |
| ODM96669.1 | springtail insect (Orchesella cincta) | |
| α2-nAchR | EAW63552.1 | human (Homo sapiens) |
| NP_001035417.1 | zebrafish (Danio rerio) | |
| NP_001267757.1 | sea hare (Aplysia californica) | |
| MF179124 | cone snail (Conus betulinus) | |
| XP_011450331.1 | oyster (Crassostrea gigas) | |
| ABA60382.1 | great pond snail (Lymnaea stagnalis) | |
| NP_001103423.1 | darkling beetle (Tribolium castaneum) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Huang, Y.; Peng, C.; Yi, Y.; Gao, B.; Shi, Q. A Transcriptomic Survey of Ion Channel-Based Conotoxins in the Chinese Tubular Cone Snail (Conus betulinus). Mar. Drugs 2017, 15, 228. https://doi.org/10.3390/md15070228
AMA Style
Huang Y, Peng C, Yi Y, Gao B, Shi Q. A Transcriptomic Survey of Ion Channel-Based Conotoxins in the Chinese Tubular Cone Snail (Conus betulinus). Marine Drugs. 2017; 15(7):228. https://doi.org/10.3390/md15070228
Chicago/Turabian StyleHuang, Yu, Chao Peng, Yunhai Yi, Bingmiao Gao, and Qiong Shi. 2017. "A Transcriptomic Survey of Ion Channel-Based Conotoxins in the Chinese Tubular Cone Snail (Conus betulinus)" Marine Drugs 15, no. 7: 228. https://doi.org/10.3390/md15070228
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.