Specific Molecular Signatures for Type II Crustins in Penaeid Shrimp Uncovered by the Identification of Crustin-Like Antimicrobial Peptides in Litopenaeus vannamei


Abstract
:1. Introduction
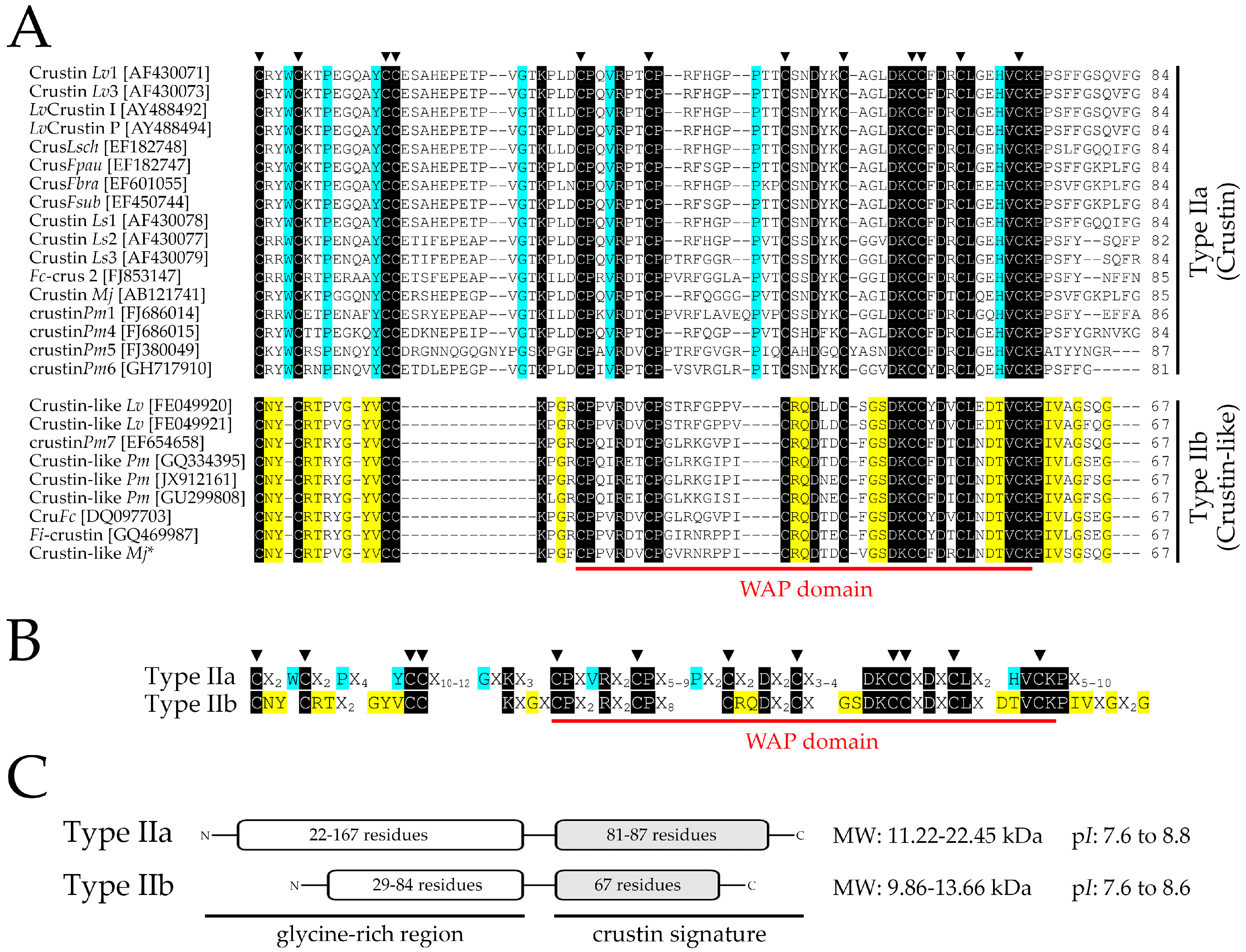
2. Results and Discussion
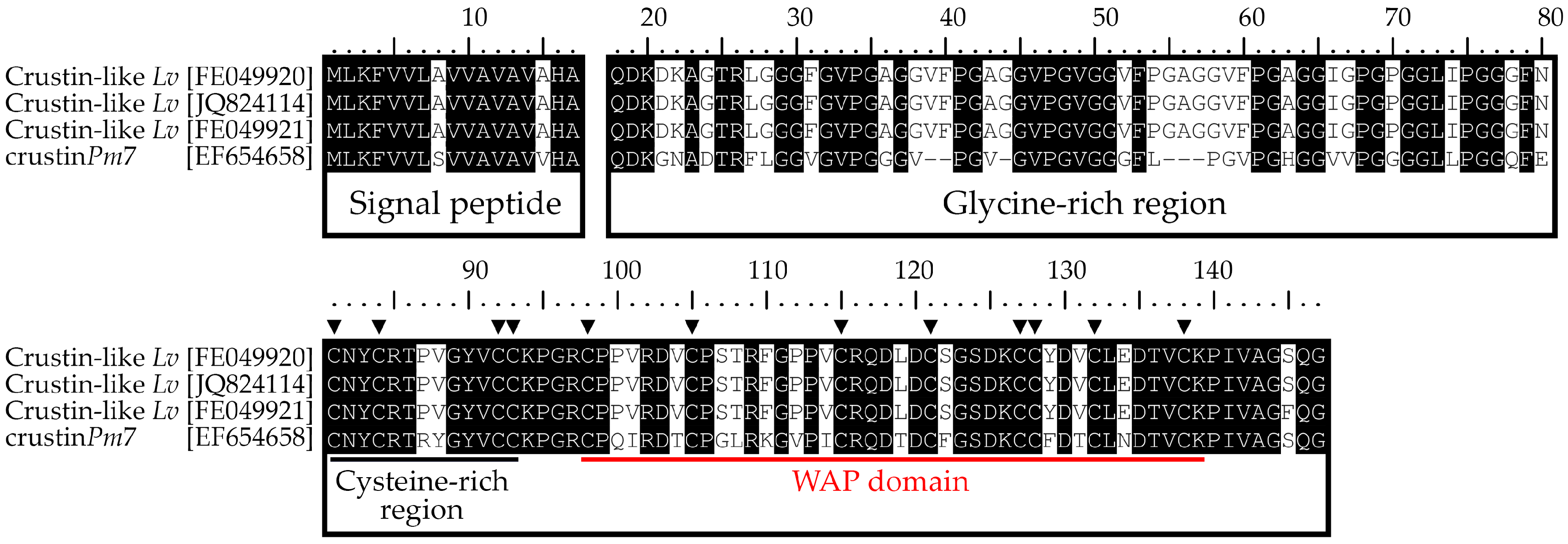
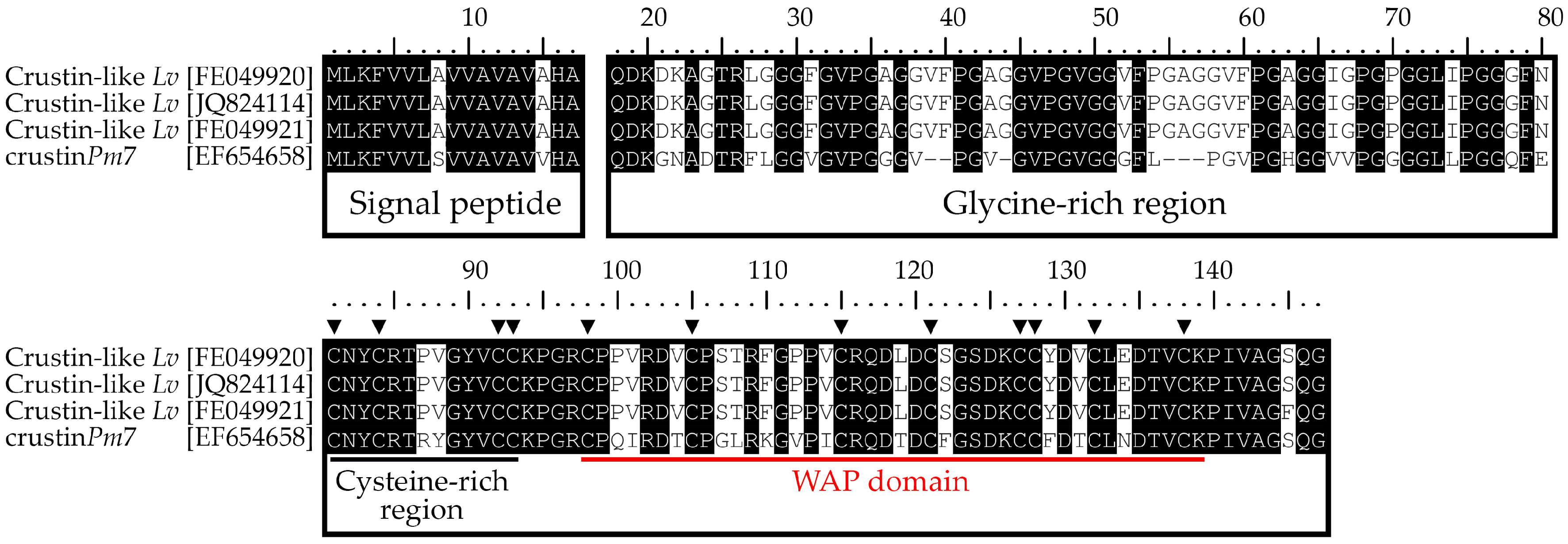
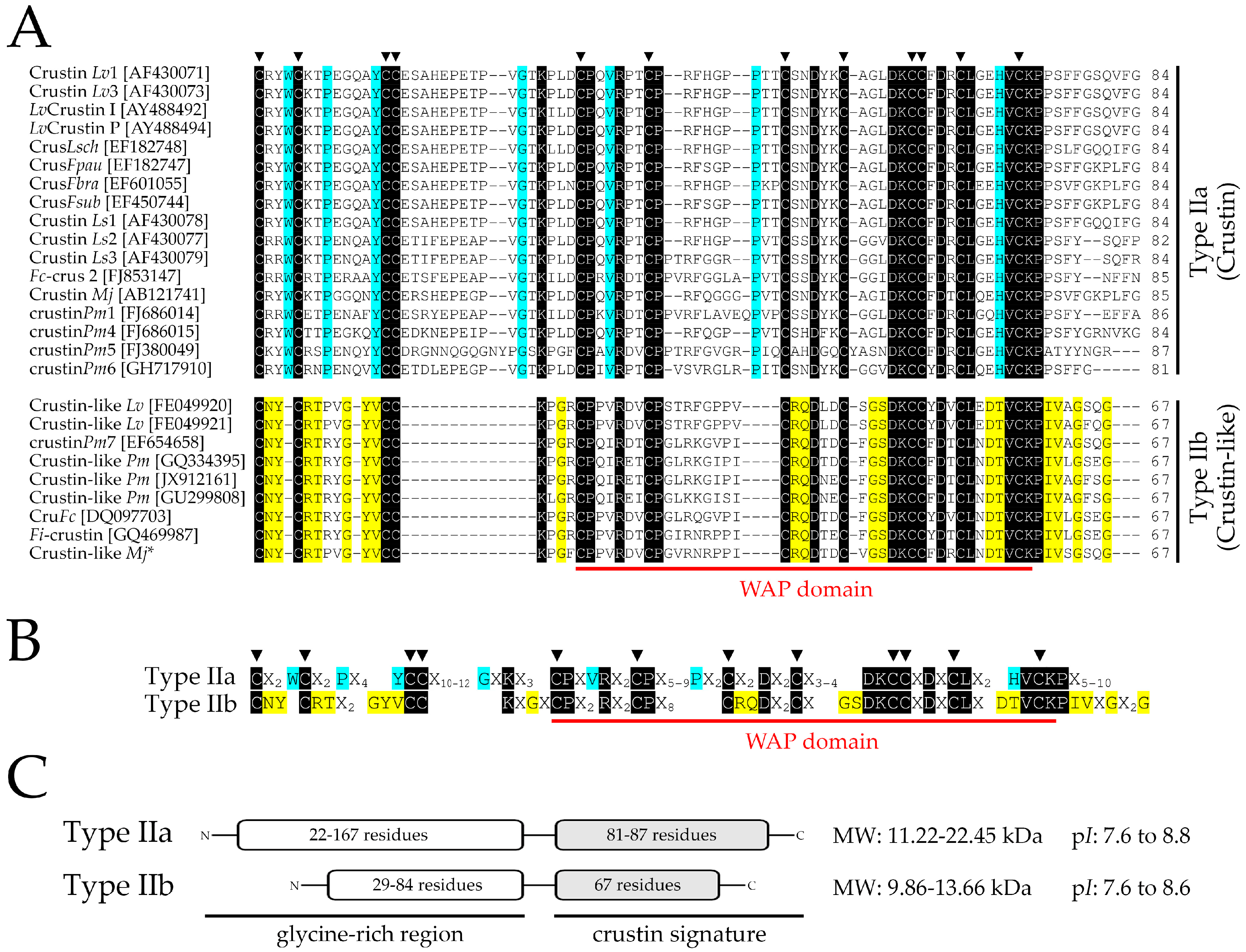
2.1. Identification and Molecular Characterization of Type IIb Crustins in L. vannamei
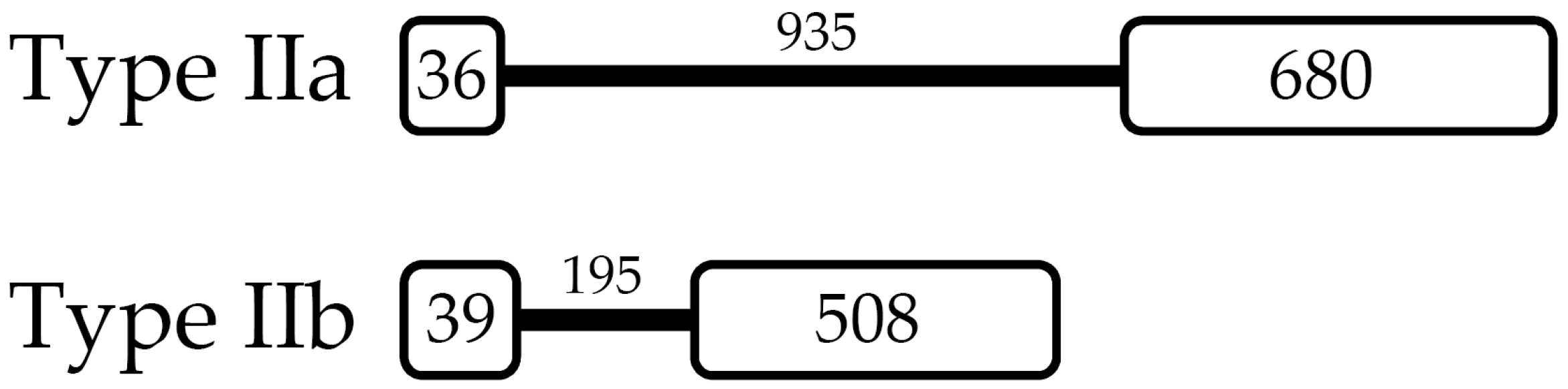
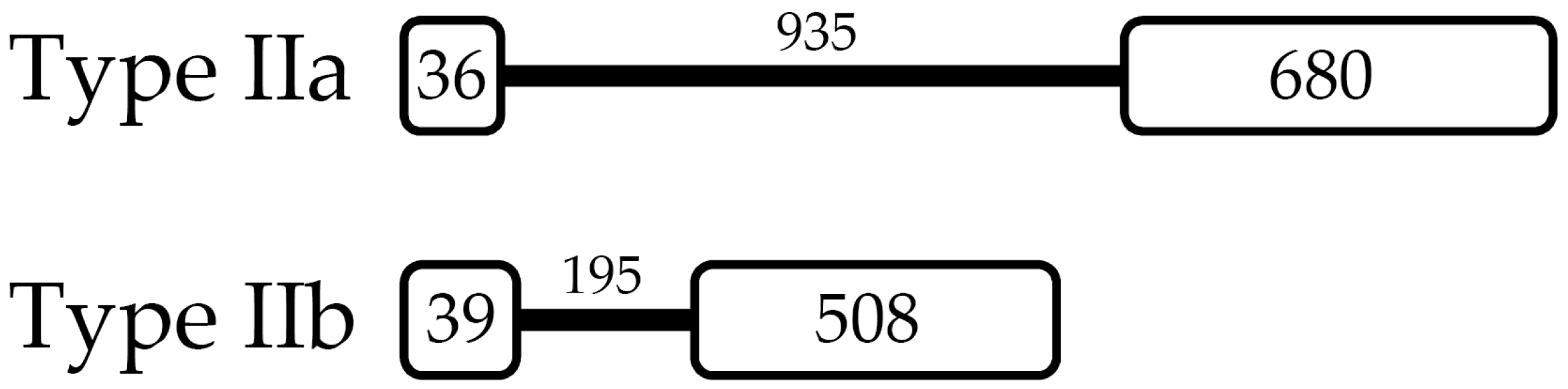
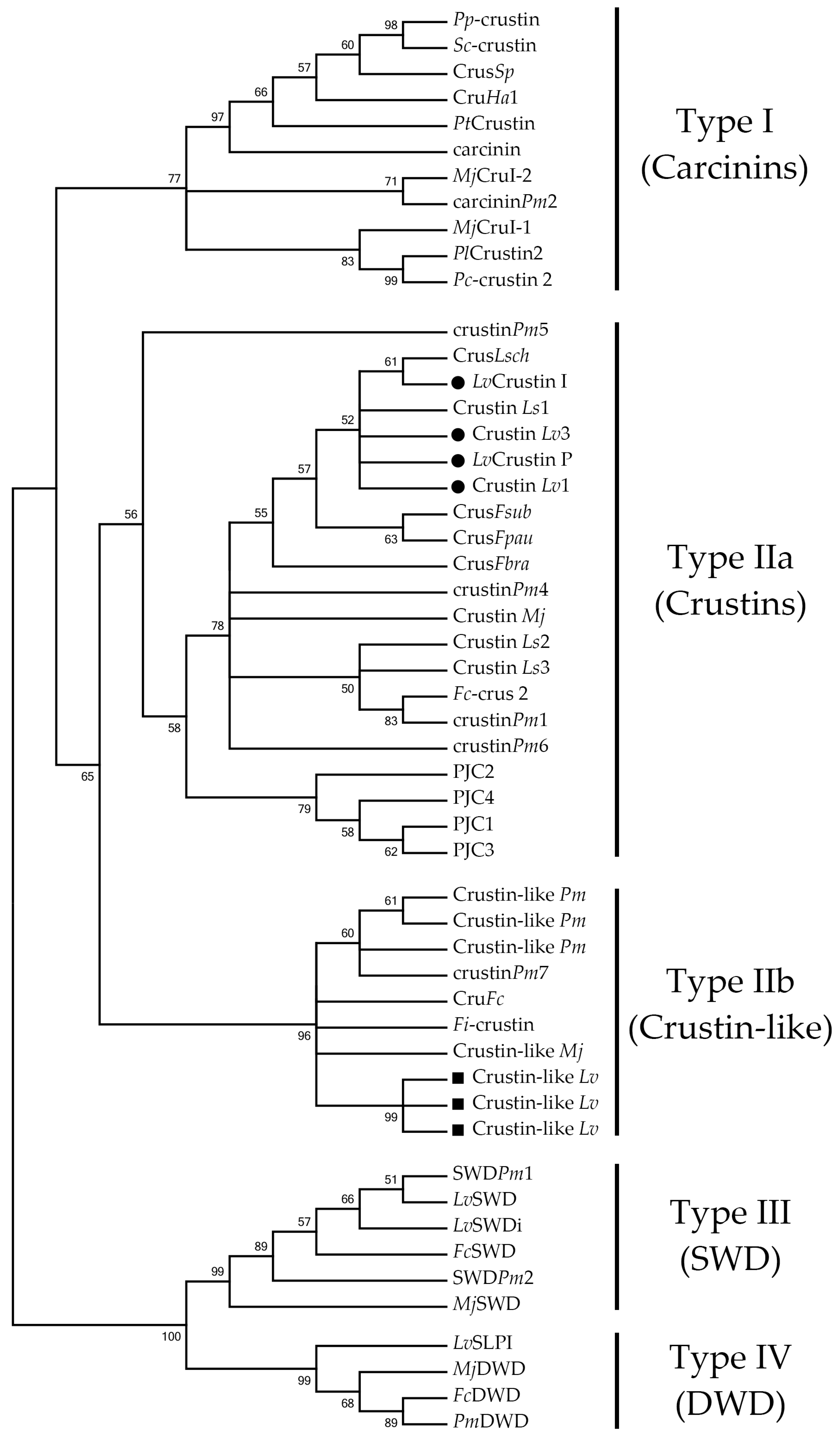
2.2. L. vannamei Type II Crustins Are Encoded by Distinct Genes
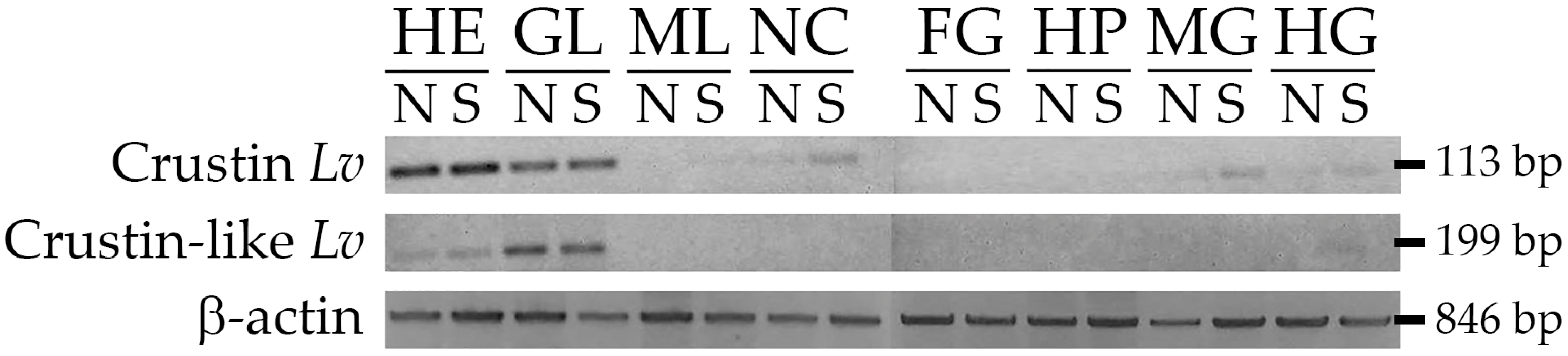
2.3. Gene Expression Distribution of Type II Crustins in Shrimp Tissues
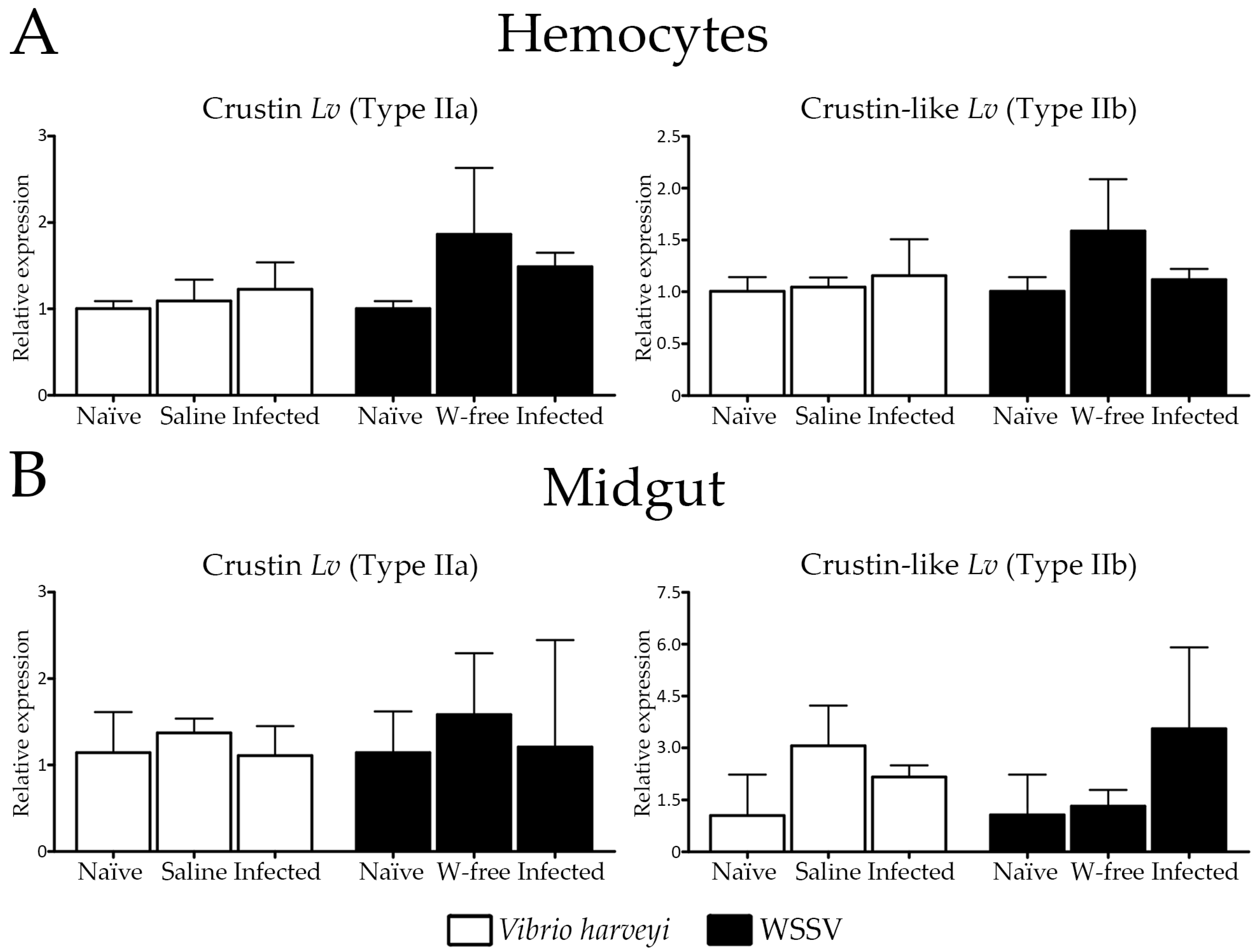
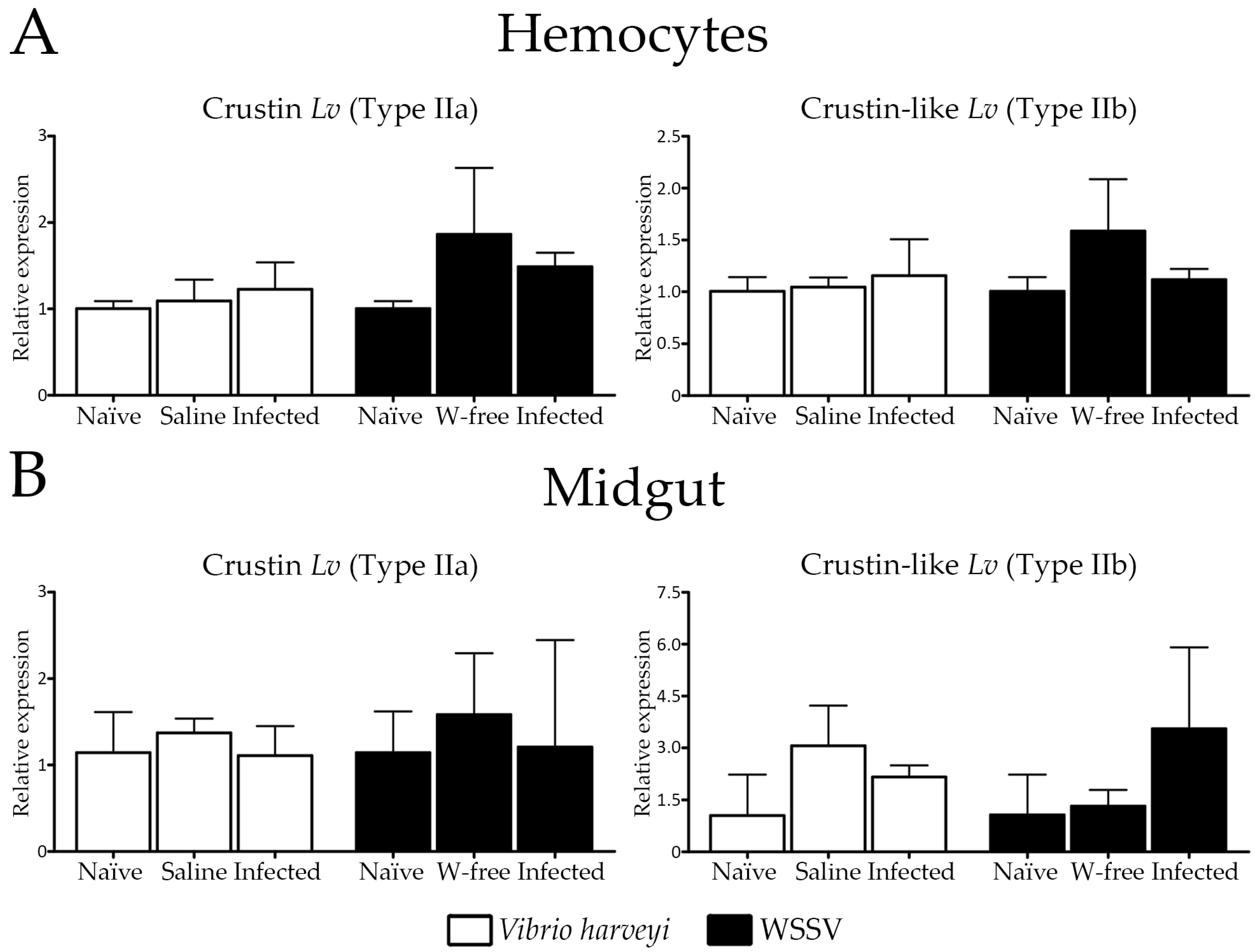
2.4. Expression Profile of Type II Crustins in Response to Bacterial and Viral Infections
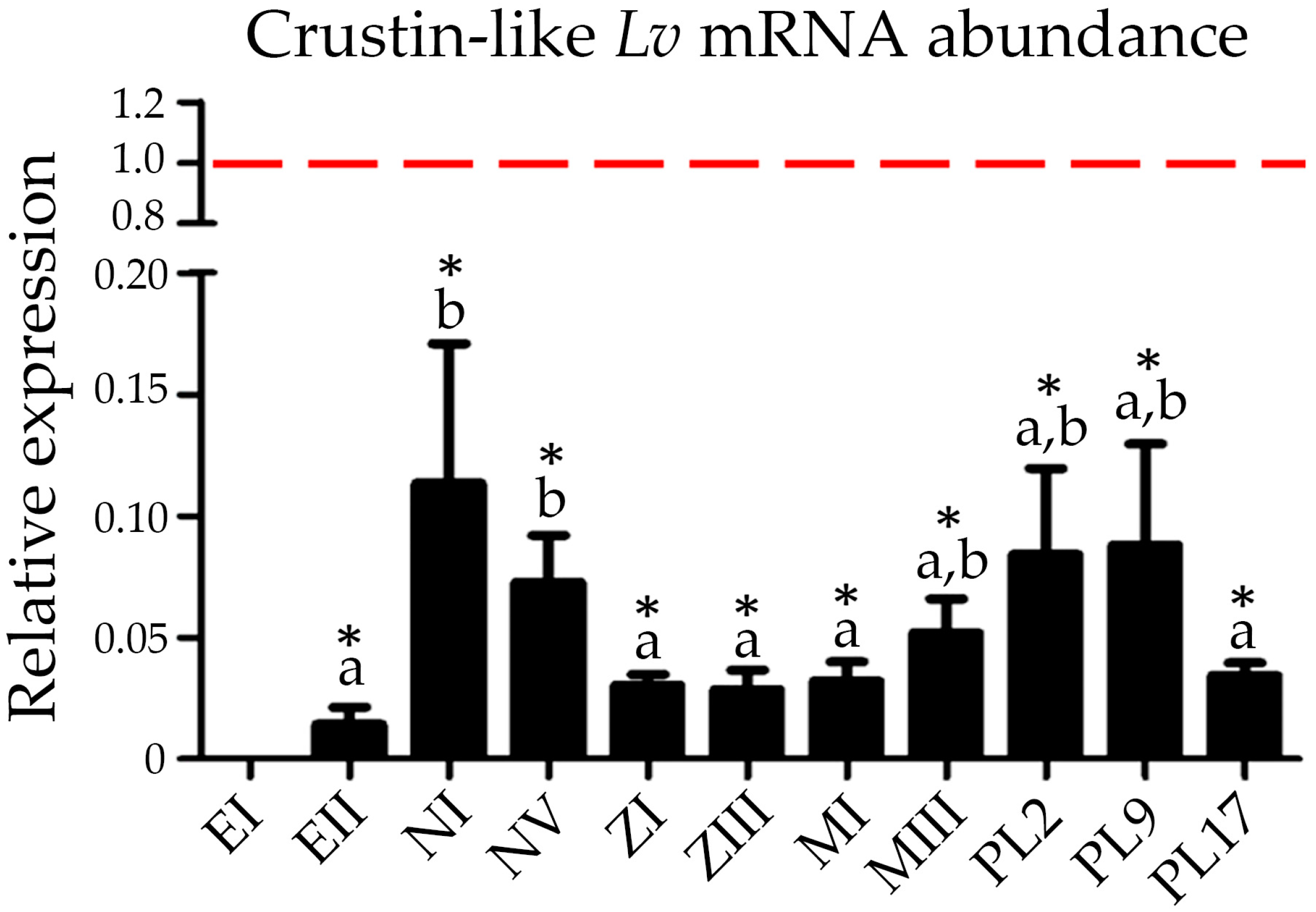
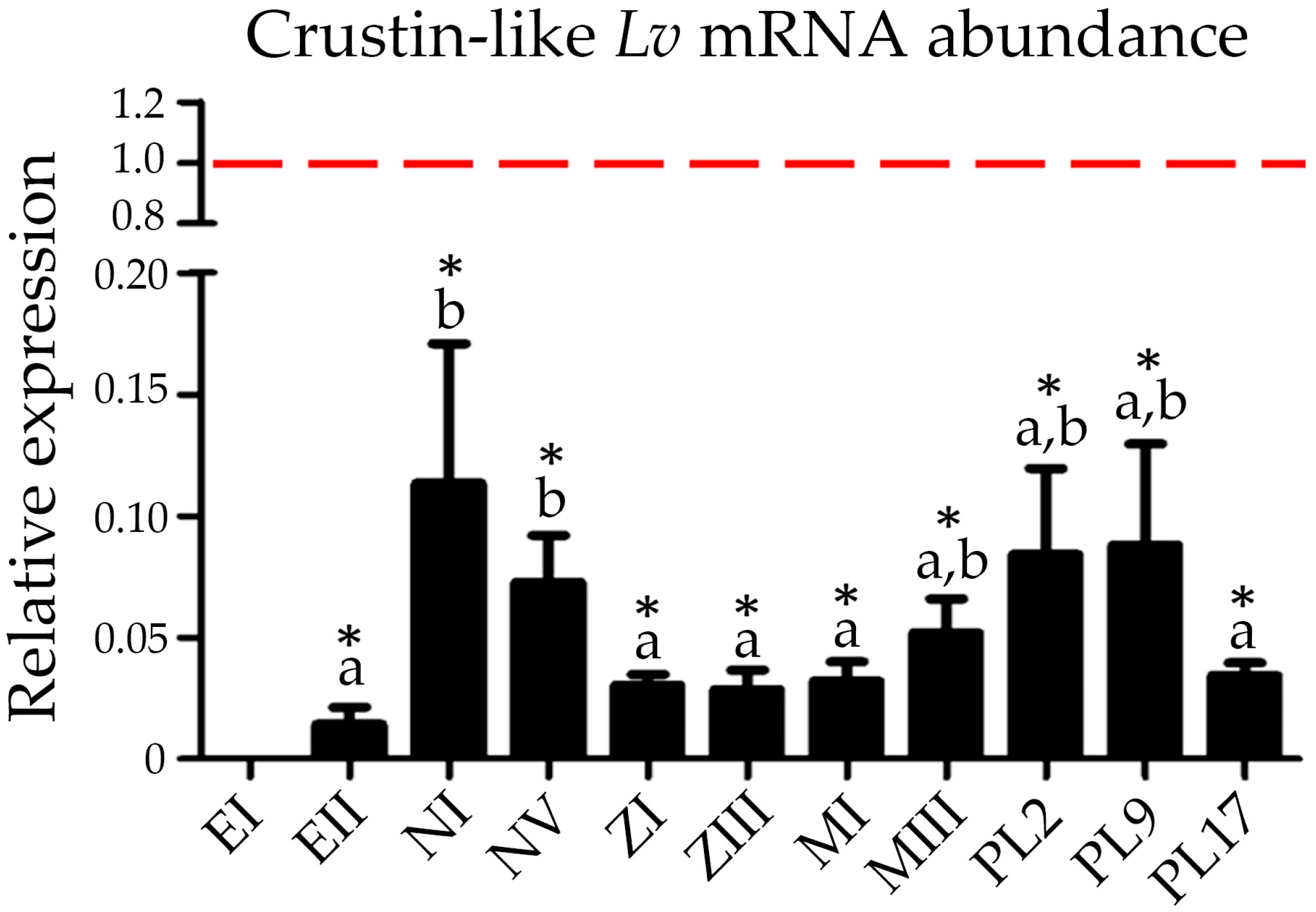
2.5. Expression of Type IIb Crustins during Shrimp Ontogenesis
2.6. Molecular Signatures for Type II Crustins in Penaeid Shrimp
3. Materials and Methods
3.1. Sequence Data Analysis and Phylogeny
3.2. Animals, Microbial Stimulation and Tissue Collection
3.3. Experimental Infections
3.4. Reverse Transcription-Polymerase Chain Reaction (RT-PCR) Analysis for Tissue Distribution of Gene Expression
3.5. Fluorescence-Based Reverse Transcription Real-Time Quantitative PCR (RT-qPCR)
3.6. Quantitative Gene Expression Analysis during Shrimp Development
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Smith, V.J.; Dyrynda, E.A. Antimicrobial proteins: From old proteins, new tricks. Mol. Immunol. 2015, 68, 383–398. [Google Scholar] [CrossRef] [PubMed]
- Ranganathan, S.; Simpson, K.J.; Shaw, D.C.; Nicholas, K.R. The whey acidic protein family: A new signature motif and three-dimensional structure by comparative modeling. J. Mol. Graph. Model. 1999, 17, 106–113. [Google Scholar] [CrossRef]
- Relf, J.M.; Chisholm, J.R.; Kemp, G.D.; Smith, V.J. Purification and characterization of a cysteine-rich 11.5-kDa antibacterial protein from the granular haemocytes of the shore crab, Carcinus maenas. Eur. J. Biochem. 1999, 264, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, T.C.; Cuthbertson, B.J.; Shepard, E.F.; Chapman, R.W.; Gross, P.S.; Warr, G.W. Crustins, homologues of an 11.5-kDa antibacterial peptide, from two species of penaeid shrimp, Litopenaeus vannamei and Litopenaeus setiferus. Mar. Biotechnol. 2002, 4, 278–293. [Google Scholar] [CrossRef] [PubMed]
- Smith, V.J.; Fernandes, J.M.O.; Kemp, G.D.; Hauton, C. Crustins: Enigmatic WAP domain-containing antibacterial proteins from crustaceans. Dev. Comp. Immunol. 2008, 32, 758–772. [Google Scholar] [CrossRef] [PubMed]
- Rosa, R.; Barracco, M. Antimicrobial peptides in crustaceans. Invertebr. Surviv. J. 2010, 7, 262–284. [Google Scholar]
- Tassanakajon, A.; Rimphanitchayakit, V.; Visetnan, S.; Amparyup, P.; Somboonwiwat, K.; Charoensapsri, W.; Tang, S. Shrimp humoral responses against pathogens: Antimicrobial peptides and melanization. Dev. Comp. Immunol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Destoumieux-Garzón, D.; Rosa, R.D.; Schmitt, P.; Barreto, C.; Vidal-Dupiol, J.; Mitta, G.; Gueguen, Y.; Bachère, E. Antimicrobial peptides in marine invertebrate health and disease. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016. [Google Scholar] [CrossRef] [PubMed]
- Brockton, V.; Hammond, J.; Smith, V. Gene characterisation, isoforms and recombinant expression of carcinin, an antibacterial protein from the shore crab, Carcinus maenas. Mol. Immunol. 2007, 44, 943–949. [Google Scholar] [CrossRef] [PubMed]
- Donpudsa, S.; Visetnan, S.; Supungul, P.; Tang, S.; Tassanakajon, A.; Rimphanitchayakit, V. Type I and type II crustins from Penaeus monodon, genetic variation and antimicrobial activity of the most abundant crustinPm4. Dev. Comp. Immunol. 2014, 47, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.-S.; Jia, W.-M.; Zhao, X.-F.; Wang, J.-X. Four crustins involved in antibacterial responses in Marsupenaeus japonicus. Fish Shellfish Immunol. 2015, 43, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Amparyup, P.; Donpudsa, S.; Tassanakajon, A. Shrimp single WAP domain (SWD)-containing protein exhibits proteinase inhibitory and antimicrobial activities. Dev. Comp. Immunol. 2008, 32, 1497–1509. [Google Scholar] [CrossRef] [PubMed]
- Tassanakajon, A.; Somboonwiwat, K.; Amparyup, P. Sequence diversity and evolution of antimicrobial peptides in invertebrates. Dev. Comp. Immunol. 2015, 48, 324–341. [Google Scholar] [CrossRef] [PubMed]
- Shockey, J.E.; O’Leary, N.A.; de la Vega, E.; Browdy, C.L.; Baatz, J.E.; Gross, P.S. The role of crustins in Litopenaeus vannamei in response to infection with shrimp pathogens: An in vivo approach. Dev. Comp. Immunol. 2009, 33, 668–673. [Google Scholar] [CrossRef] [PubMed]
- Hipolito, S.G.; Shitara, A.; Kondo, H.; Hirono, I. Role of Marsupenaeus japonicus crustin-like peptide against Vibrio penaeicida and white spot syndrome virus infection. Dev. Comp. Immunol. 2014, 46, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Wang, Z.; Zhu, F. The crustin-like peptide plays opposite role in shrimp immune response to Vibrio alginolyticus and white spot syndrome virus (WSSV) infection. Fish Shellfish Immunol. 2017, 66, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, F.; Wang, Z.; Xiang, J. Cloning and recombinant expression of a crustin-like gene from Chinese shrimp, Fenneropenaeus chinensis. J. Biotechnol. 2007, 127, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Amparyup, P.; Kondo, H.; Hirono, I.; Aoki, T.; Tassanakajon, A. Molecular cloning, genomic organization and recombinant expression of a crustin-like antimicrobial peptide from black tiger shrimp Penaeus monodon. Mol. Immunol. 2008, 45, 1085–1093. [Google Scholar] [CrossRef] [PubMed]
- Supungul, P.; Klinbunga, S.; Pichyangkura, R.; Jitrapakdee, S.; Hirono, I.; Aoki, T.; Tassanakajon, A. Identification of immune-related genes in hemocytes of black tiger shrimp (Penaeus monodon). Mar. Biotechnol. 2002, 4, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Rosa, R.D.; Bandeira, P.T.; Barracco, M.A. Molecular cloning of crustins from the hemocytes of Brazilian penaeid shrimps. FEMS Microbiol. Lett. 2007. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Du, X.-J.; Xu, W.-T.; Zhang, H.-W.; Zhao, X.-F.; Wang, J.-X. Molecular cloning and characterization of three crustins from the Chinese white shrimp, Fenneropenaeus chinensis. Fish Shellfish Immunol. 2010, 28, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Antony, S.P.; Bright Singh, I.S.; Philip, R. Molecular characterization of a crustin-like, putative antimicrobial peptide, Fi-crustin, from the Indian white shrimp, Fenneropenaeus indicus. Fish Shellfish Immunol. 2010, 28, 216–220. [Google Scholar] [CrossRef] [PubMed]
- Pisuttharachai, D.; Fagutao, F.F.; Yasuike, M.; Aono, H.; Yano, Y.; Murakami, K.; Kondo, H.; Aoki, T.; Hirono, I. Characterization of crustin antimicrobial proteins from Japanese spiny lobster Panulirus japonicus. Dev. Comp. Immunol. 2009, 33, 1049–1054. [Google Scholar] [CrossRef] [PubMed]
- Vatanavicharn, T.; Supungul, P.; Puanglarp, N.; Yingvilasprasert, W.; Tassanakajon, A. Genomic structure, expression pattern and functional characterization of crustinPm5, a unique isoform of crustin from Penaeus monodon. Comp. Biochem. Physiol. B. Biochem. Mol. Biol. 2009, 153, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Padhi, A. Adaptive evolution of crustin antimicrobial peptides in decapods. Genetica 2012, 140, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Supungul, P.; Tang, S.; Maneeruttanarungroj, C.; Rimphanitchayakit, V.; Hirono, I.; Aoki, T.; Tassanakajon, A. Cloning, expression and antimicrobial activity of crustinPm1, a major isoform of crustin, from the black tiger shrimp Penaeus monodon. Dev. Comp. Immunol. 2008, 32, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-C.; Chang, P.-S.; Chen, H.-Y. Tissue expressions of nine genes important to immune defence of the Pacific white shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2007, 23, 1161–1177. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-C.; Lu, C.-L.; Chen, S.; Liao, W.-L.; Chen, S. Immune gene expression for diverse haemocytes derived from pacific white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol. 2015, 44, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Davies, L.C.; Taylor, P.R. Tissue-resident macrophages: Then and now. Immunology 2015, 144, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Suleiman, S.; Smith, V.J.; Dyrynda, E.A. Unusual tissue distribution of carcinin, an antibacterial crustin, in the crab, Carcinus maenas, reveals its multi-functionality. Dev. Comp. Immunol. 2017, 76, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, R.G.; Kang, S.; Simões, M.L.; Angleró-Rodríguez, Y.I.; Dimopoulos, G. Mosquito gut antiparasitic and antiviral immunity. Dev. Comp. Immunol. 2016, 64, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, P.; Guertler, C.; Bachère, E.; de Souza, C.R.B.; Rosa, R.D.; Perazzolo, L.M. Molecular signatures at imminent death: Hemocyte gene expression profiling of shrimp succumbing to viral and fungal infections. Dev. Comp. Immunol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Soonthornchai, W.; Rungrassamee, W.; Karoonuthaisiri, N.; Jarayabhand, P.; Klinbunga, S.; Söderhäll, K.; Jiravanichpaisal, P. Expression of immune-related genes in the digestive organ of shrimp, Penaeus monodon, after an oral infection by Vibrio harveyi. Dev. Comp. Immunol. 2010, 34, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Destoumieux, D.; Muñoz, M.; Cosseau, C.; Rodriguez, J.; Bulet, P.; Comps, M.; Bachère, E. Penaeidins, antimicrobial peptides with chitin-binding activity, are produced and stored in shrimp granulocytes and released after microbial challenge. J. Cell Sci. 2000, 113, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Quispe, R.L.; Justino, E.B.; Vieira, F.N.; Jaramillo, M.L.; Rosa, R.D.; Perazzolo, L.M. Transcriptional profiling of immune-related genes in Pacific white shrimp (Litopenaeus vannamei) during ontogenesis. Fish Shellfish Immunol. 2016, 58, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Hauton, C.; Brockton, V.; Smith, V.J. Cloning of a crustin-like, single whey-acidic-domain, antibacterial peptide from the haemocytes of the European lobster, Homarus gammarus, and its response to infection with bacteria. Mol. Immunol. 2006, 43, 1490–1496. [Google Scholar] [CrossRef] [PubMed]
- Christie, A.E.; Rus, S.; Goiney, C.C.; Smith, C.M.; Towle, D.W.; Dickinson, P.S. Identification and characterization of a cDNA encoding a crustin-like, putative antibacterial protein from the American lobster Homarus americanus. Mol. Immunol. 2007, 44, 3333–3337. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wang, J. The antimicrobial peptides of the immune response of shrimp. Invertebr. Surviv. J. 2008, 5, 162–179. [Google Scholar]
- Du, Z.-Q.; Yuan, J.-J.; Ren, D.-M. A novel single WAP domain-containing protein isoform with antibacterial relevance in Litopenaeus vannamei. Fish Shellfish Immmunol. 2015, 44, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Abdelrahman, H.; ElHady, M.; Alcivar-Warren, A.; Allen, S.; Al-Tobasei, R.; Bao, L.; Beck, B.; Blackburn, H.; Bosworth, B.; Buchanan, J.; et al. Aquaculture genomics, genetics and breeding in the United States: Current status, challenges, and priorities for future research. BMC Genom. 2017, 18, 191. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Amplicon |
|---|---|---|---|
| Primers for Tissue Distribution Analysis (RT-PCR) | |||
| β-actin | TAATCCACATCTGCTGGAAGGTGG | TCACCAACTGGGATGACATGG | 846 bp |
| Crustin Lv | CGAACCAGAGACACCTGTTG | CAGCACACTTGTAGTCGTTG | 113 bp |
| Crustin-like Lv | GCAGGATAAAGACAAGGC | GTAATTGCAGTTGAATCCGCC | 199 bp |
| Primers for Quantitative Analysis of Gene Expression (RT-qPCR) | |||
| LvEF1α | TGGCTGTGAACAAGATGGACA | TTGTAGCCCACCTTCTTGACG | 103 bp |
| LvRpS6 | AGCAGATACCCTTGGTGAAG | GATGCAACCACGGACTGAC | 193 bp |
| LvL40 | GAGAATGTGAAGGCCAAGATC | TCAGAGAGAGTGCGACCATC | 104 bp |
| LvRpS3A | GGCTTGCTATGGTGTGCTCC | TCATGCTCTTGGCTCGCTG | 101 bp |
| β-actin | CCACGAGACCACCTACAAC | AGCGAGGGCAGTGATTTC | 142 bp |
| Crustin Lv | CGAACCAGAGACACCTGTTG | CAGCACACTTGTAGTCGTTG | 113 bp |
| Crustin-like Lv | GCAGGATAAAGACAAGGC | GTAATTGCAGTTGAATCCGCC | 199 bp |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barreto, C.; Coelho, J.D.R.; Yuan, J.; Xiang, J.; Perazzolo, L.M.; Rosa, R.D. Specific Molecular Signatures for Type II Crustins in Penaeid Shrimp Uncovered by the Identification of Crustin-Like Antimicrobial Peptides in Litopenaeus vannamei. Mar. Drugs 2018, 16, 31. https://doi.org/10.3390/md16010031
Barreto C, Coelho JDR, Yuan J, Xiang J, Perazzolo LM, Rosa RD. Specific Molecular Signatures for Type II Crustins in Penaeid Shrimp Uncovered by the Identification of Crustin-Like Antimicrobial Peptides in Litopenaeus vannamei. Marine Drugs. 2018; 16(1):31. https://doi.org/10.3390/md16010031
Chicago/Turabian StyleBarreto, Cairé, Jaqueline Da Rosa Coelho, Jianbo Yuan, Jianhai Xiang, Luciane Maria Perazzolo, and Rafael Diego Rosa. 2018. "Specific Molecular Signatures for Type II Crustins in Penaeid Shrimp Uncovered by the Identification of Crustin-Like Antimicrobial Peptides in Litopenaeus vannamei" Marine Drugs 16, no. 1: 31. https://doi.org/10.3390/md16010031