Purification and Characterization of a Novel Alginate Lyase from the Marine Bacterium Bacillus sp. Alg07

1
National Engineering Laboratory for Industrial Enzymes, Tianjin Institute of Industrial Biotechnology, Chinese Academy of Sciences, Tianjin 300308, China
2
University of Chinese Academy of Sciences, Beijing 100049, China
*
Author to whom correspondence should be addressed.
Mar. Drugs 2018, 16(3), 86; https://doi.org/10.3390/md16030086
Submission received: 9 February 2018
/
Revised: 28 February 2018
/
Accepted: 5 March 2018
/
Published: 9 March 2018
(This article belongs to the Special Issue Marine Enzymes: Sources, Biochemistry and Bioprocesses for Marine Biotechnology)
Abstract
:Alginate oligosaccharides with different bioactivities can be prepared through the specific degradation of alginate by alginate lyases. Therefore, alginate lyases that can be used to degrade alginate under mild conditions have recently attracted public attention. Although various types of alginate lyases have been discovered and characterized, few can be used in industrial production. In this study, AlgA, a novel alginate lyase with high specific activity, was purified from the marine bacterium Bacillus sp. Alg07. AlgA had a molecular weight of approximately 60 kDa, an optimal temperature of 40 °C, and an optimal pH of 7.5. The activity of AlgA was dependent on sodium chloride and could be considerably enhanced by Mg2+ or Ca2+. Under optimal conditions, the activity of AlgA reached up to 8306.7 U/mg, which is the highest activity recorded for alginate lyases. Moreover, the enzyme was stable over a broad pH range (5.0–10.0), and its activity negligibly changed after 24 h of incubation at 40 °C. AlgA exhibited high activity and affinity toward poly-β-d-mannuronate (polyM). These characteristics suggested that AlgA is an endolytic polyM-specific alginate lyase (EC 4.2.2.3). The products of alginate and polyM degradation by AlgA were purified and identified through fast protein liquid chromatography and electrospray ionization mass spectrometry, which revealed that AlgA mainly produced disaccharides, trisaccharides, and tetrasaccharide from alginate and disaccharides and trisaccharides from polyM. Therefore, the novel lysate AlgA has potential applications in the production of mannuronic oligosaccharides and poly-α-l-guluronate blocks from alginate.
1. Introduction
Alginate is a linear copolymer that is composed of homopolymeric blocks of (1–4)-linked α-l-guluronic acid (G) and its C5 epimer β-d-mannuronic acid (M), which forms three types of blocks: poly-α-l-guluronate (polyG), poly-β-d-mannuronate (polyM), and random heteropolymeric sequences (polyMG) [1]. Alginate is the most abundant carbohydrate in brown algae, and it accounts for up to 10–45% of the dry weight of brown algae [2]. Some bacteria that belong to the genera Azotobacter [3] and Pseudomonas [4] produce alginate as an extracellular polysaccharide. In contrast to algal alginate, bacterial alginate is acetylated. Commercial alginate manufactured from brown algae has been used as a thickening agent or gelling agent in the food and pharmaceutical industries [5]. Alginate can be degraded into alginate oligosaccharides (AOS) through a chemical process or by alginate lyase. Given that AOS can stimulate the growth of endothelial cells [6] and the production of multiple cytokines [7], they may be applied as growth-promoting agents in some plants [8] and bifidobacteria [9]. Furthermore, AOS demonstrate excellent antioxidant activity [10] and havepotential uses in protection against pathogens [11].
In alginate degradation, alginate lyases cleave the (1–4)-linked glucosidic bond of alginate via a β-elimination mechanism and generate unsaturated oligosaccharides with 4-deoxy-alpha-l-erythrohex-4-enopyranuronosyl uronate as the nonreducing terminal residue [12]. Numerous alginate lyases have been isolated from various organisms, such as marine algae [13], marine mollusks [14], marine and terrestrial bacteria [15,16], marine fungi [17], and viruses [18]. Alginate lyases can be categorized into polyM-specific, polyG-specific, and polyMG-specific lyases on the basis of their substrate preferences [19] or into endo- or exo-alginate lyases on the basis of their cleavage mode [19]. In the carbohydrate-active enzyme database, alginate lyases belong to the polysaccharide lyase family [20]. The structures of some alginate lyases have been elucidated.
Alginate lyases are widely used in many fields. For example, alginate lyases have been employed to explain the fine structures of alginate [21] and to prepare red and brown algal protoplasts [22]. These enzymes may be utilized in the treatment of cystic fibrosis [23] and have been used as catalysts for AOS production [24]. The application of alginate lyases in alginate degradation under mild conditions has recently attracted public attention given the high efficiency and specifi of these enzymes. Nevertheless, present studies on alginate lyases remain in development, and the low catalytic efficiency and poor thermostability of alginate lyases limit their utility in AOS production. Therefore, high-efficiency and thermostable alginate lyases should be identified for use in AOS production.
In our work, we isolated and identified Bacillus sp. Alg07, a novel marine bacterium. AlgA, the alginate lyase secreted by this strain, showed extremely high activity. Hence, we purified and characterized AlgA to confirm that it has potential applications in AOS production.
2. Results and Discussion
2.1. Screening and Identification of Strain Alg07
Twenty-one strains with alginate lyase activity were isolated using alginate as the sole carbon source. Alg07 secreted the alginate lyase with the highest activity in the fermentation culture.
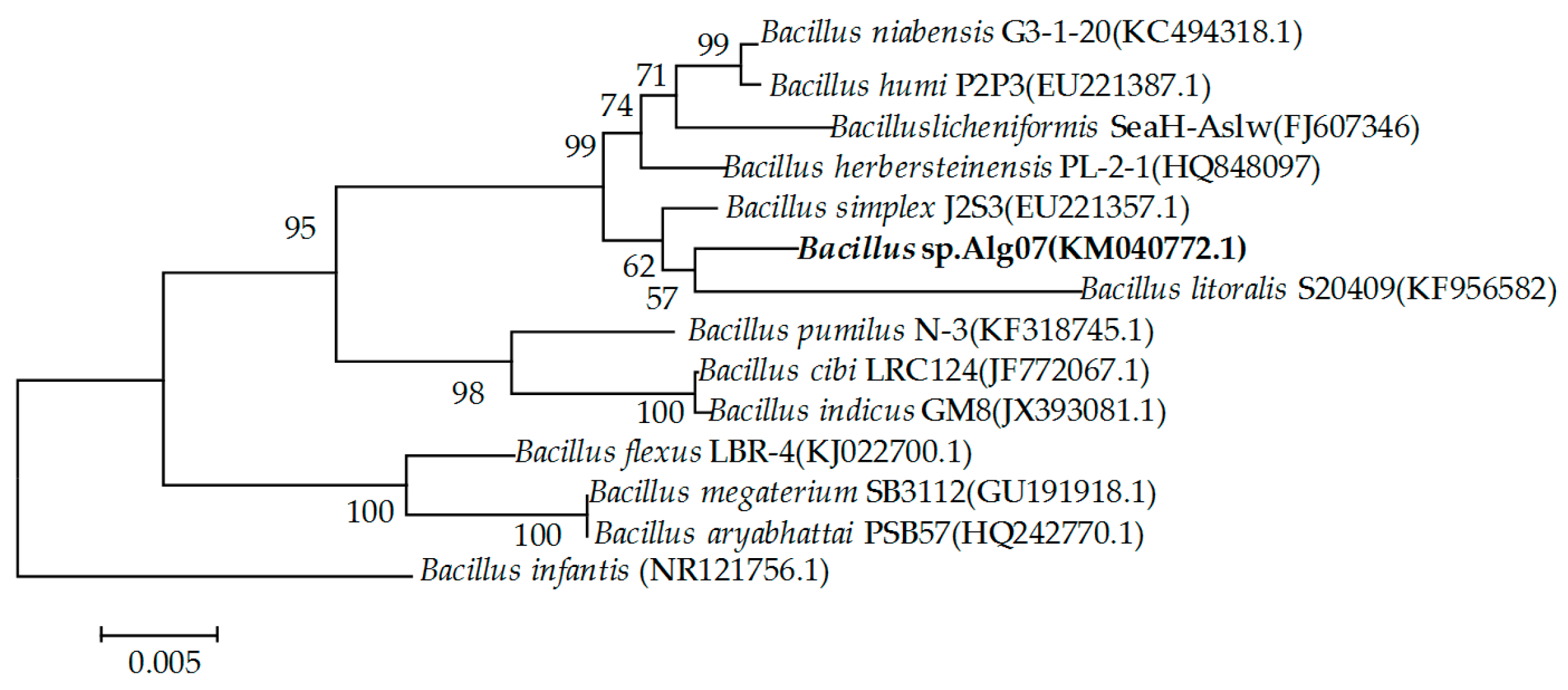
The 16S rRNA gene of Alg07 was cloned, sequenced, and submitted to GeneBank (accession number KM040772) for strain identification. The alignment of 16S rRNA gene sequences from different Bacillus species showed that strain Alg07 is closely related to Bacillus litoralis S20409 (97%) and Bacillus simplex J2S3 (97%). However, the low similarity shared by the 16s rRNA gene sequence of Alg07 with that of other known Bacillus species indicated that Alg07 may be a novel Bacillus species. In accordance with the neighbor-joining phylogenetic tree, the strain was assigned to the genus Bacillus and designated as Bacillus sp. Alg07 (Figure 1).
2.2. Purification of Alginate Lyase from Bacillus sp. Alg07
Bacillus sp. Alg07 was cultured in optimized liquid medium for 24 h until its alginate lyase reached the highest activity. The supernatant containing 510 U/mL of alginate lyase was subjected to further purification through the two simple steps of tangential flow filtration concentration and anion exchange chromatography with Source 15Q (Figure S1). After purification, the alginate lyase was purified 8.34-fold with a yield of 62.4% (Table 1). The final specific activity of the purified alginate lyase was 8306.7 U/mg. The purified alginate lyase from Bacillus sp. Alg07 was designated as AlgA.
The activities of AlgA and those of other well-studied strains are shown in Table 2, which shows that AlgA has the highest activity among all reported alginate lyases. The simple purification, high recovery, and high specific enzyme activity of AlgA indicated that it may be produced industrially on a large scale.
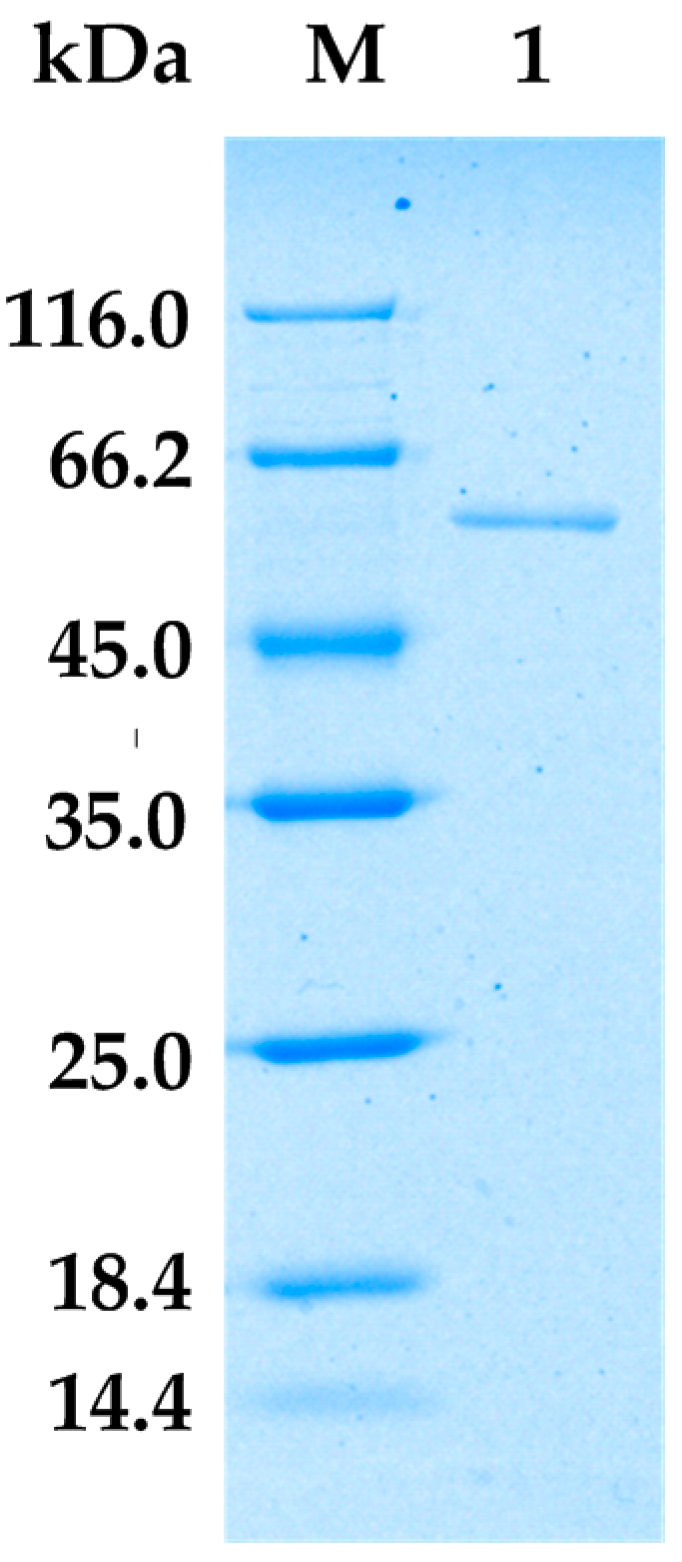
Figure 2 shows that the purified AlgA exhibited a clear and unique band on sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). This result suggested that the two-step purification process is successful. The molecular weight of AlgA was approximately 60 kDa, which is similar to molecular weights of alginate lyases from Vibrio sp.YKW-34 [30] and Saccharophagus degradans [26]. AlgA belongs to the class of large alginate lyases on the basis of its molecular weight [31].
The N-terminal amino acid sequence of the purified AlgA was analyzed. The sequence was identified as Glutamic acid–Glutamic acid–Glutamic acid–Glutamic acid–Aspartic acid–Valine–Threonine–Tyrosine (Figure S2). The results from homology search using BLASTp and CLUSTAL X indicated that the N-terminal amino acid sequence of AlgA is absent from the sequences of the previously reported alginate lyases. Furthermore, a protein with four Glu residues at its N-terminal is unusual. Therefore, AlgA may be a novel alginate lyase.
2.3. Biochemical Characterization of AlgA
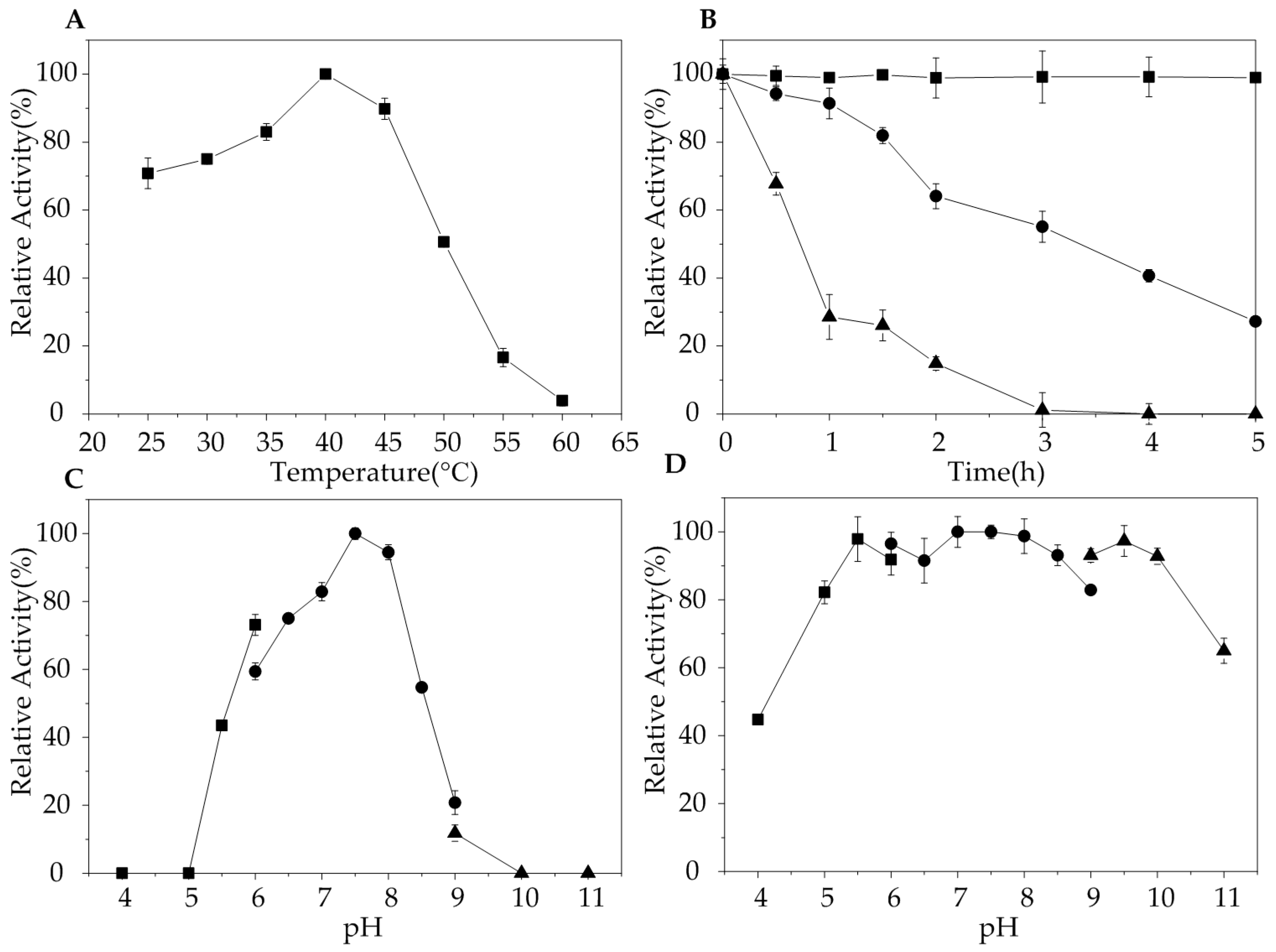
The optimal temperature of AlgA is 40 °C, which is similar to alginate lyases derived from Vibrio sp.YKW-34 [30], Flavobacterium sp. LXA [11], Pseudomonas aeruginosa PA1167 [32], and Stenotrophomnas maltophilia KJ-2 [33]. The activity of AlgA significantly decreased under temperatures exceeding 50 °C (Figure 3A). Thermostability analysis indicated that the activity of AlgA remained relatively unchanged after 5 h of incubation at 40 °C and did not decrease even after 24 h of incubation at 40 °C (data not shown). However, the enzyme was less stable under high temperatures, and the half-life of AlgA is approximately 3 h under 45 °C and 0.75 h under 50 °C (Figure 3B). This result indicated that at its optimal temperature, AlgA might be the most stable enzyme among all reported alginate lyases. Alginate lyases from Favobacterium sp. LXA [11], Vibrio sp. W13 [34], and Zobellia galactanivorans [35] are stable only under temperatures less than 40 °C. The activities of many alginate lyases considerably decrease after incubation at 40 °C. AlyL2 from Agarivorans sp. L11 has a half-life of 125 min at 40 °C [36], and OalS17 from Shewanella sp. Kz7 retains 88% of its activity after 1 h of incubation at 40 °C [25]. Furthermore, alginate lyases from Flammeovirga sp. MY04 [37] and Vibrio sp. SY08 [27] retain approximately 80 and 75%, respectively, of their activities after 2 h of incubation at 40 °C. Thermostable enzymes are more advantageous than thermolabile counterparts due totheir long half-lives and low production costs. Thus, AlgA has potential industrial applications given its excellent thermostability.
The optimal pH of AlgA was 7.5 in 20 mM Tris-HCl buffer. AlgA exhibited more than 90% of its maximal activity in pH 8.0 buffer. These results suggested that AlgA is basophilic (Figure 3C). Similarly, the optimal activities of most alginate lyases from marine bacteria are observed at pH 7.5–8.0 (Table 2). The results of the pH stability assay showed that AlgA presented the highest stability for an extended period over a pH range of 5.0–10.0 (Figure 3D). The good stability of AlgA over a wide pH range indicated its suitability for industrial application. Furthermore, the use of AlgA may decrease production costs given that pH adjustment will be unnecessary even when various alginates from different sources are employed as substrates.
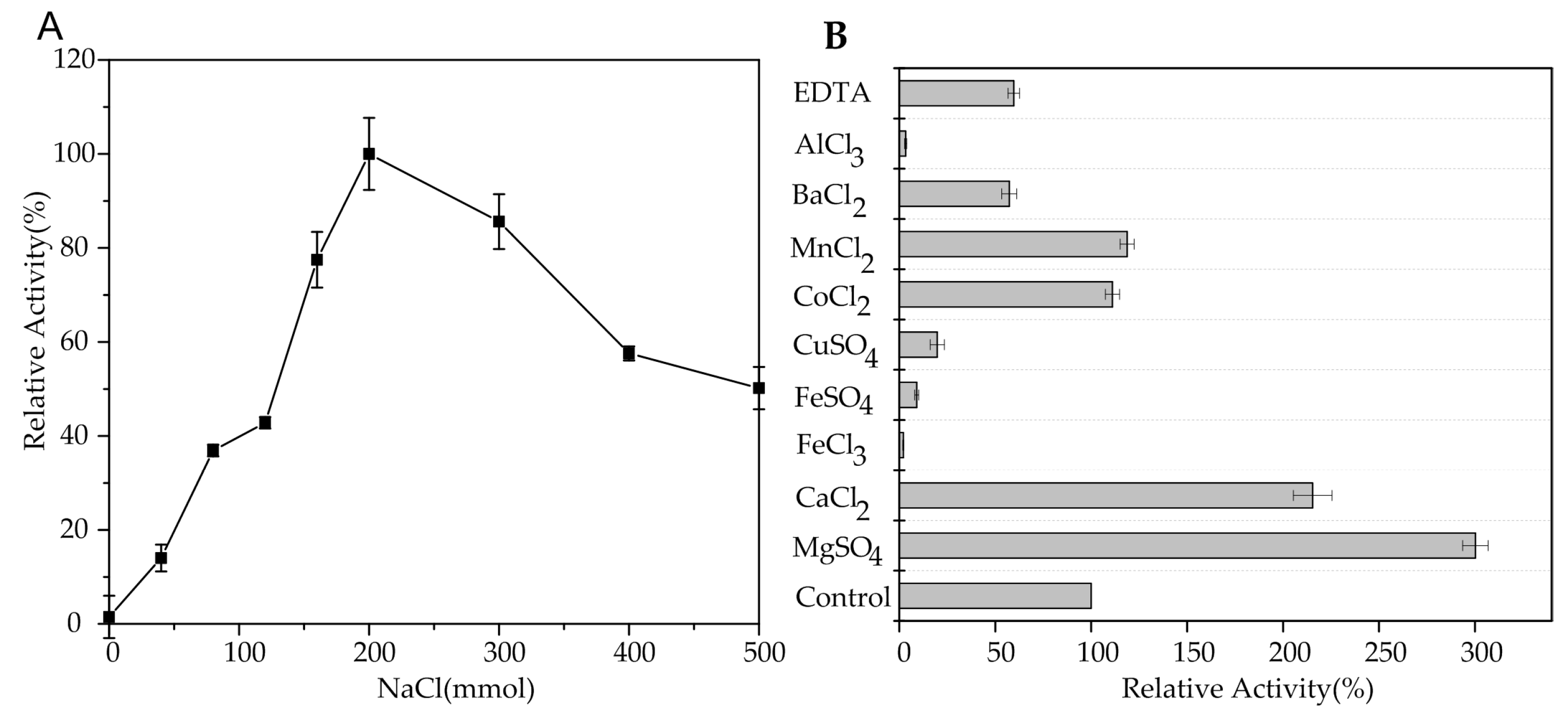
To determine the effect of NaCl on AlgA, the activity of AlgA in the presence of various NaCl concentrations was measured. As shown in Figure 4A, the optimal NaCl concentration for AlgA activity was 200 mM, and no activity was detected in the absence of NaCl. These results indicated that the activity of AlgA is dependent on NaCl. Thus, NaCl concentration is crucial for the activity of AlgA, which is a salt-activated alginate lyase. However, high NaCl concentrations decreased the activity of AlgA. Similarly, the activities of marine bacterial alginate lyases, such as AlyV5 from Vibrio sp. QY105 [28] and AlyYKW-34 from Vibrio sp. YKW-34 [30], are dependent on NaCl.
The effects of metal ions and EDTA on the activity of AlgA were determined in the presence of 200 mM NaCl. Mg2+ or Ca2+ significantly enhanced enzymatic activity by 300% or 215%, and Co2+ and Mn2+ slightly increased enzymatic activity (Figure 4B). By contrast, Hg2+, Fe3+, Fe2+, and Cu2+ completely inhibited lyase activity. Ba2+ and EDTA partially inhibited lyase activity. Ca2+ and Mg2+ increase the activity of many alginate lyases, such as aly-SJ02 from Pseudoalteromonas sp. SM0524 [38] and AlyV5 from Vibrio sp. QY105 [28]. However, Mg2+ and Ca2+ decrease the activity of Alg7D from S. degradans [26].
2.4. Substrate Specificity and Kinetic Parameters of AlgA
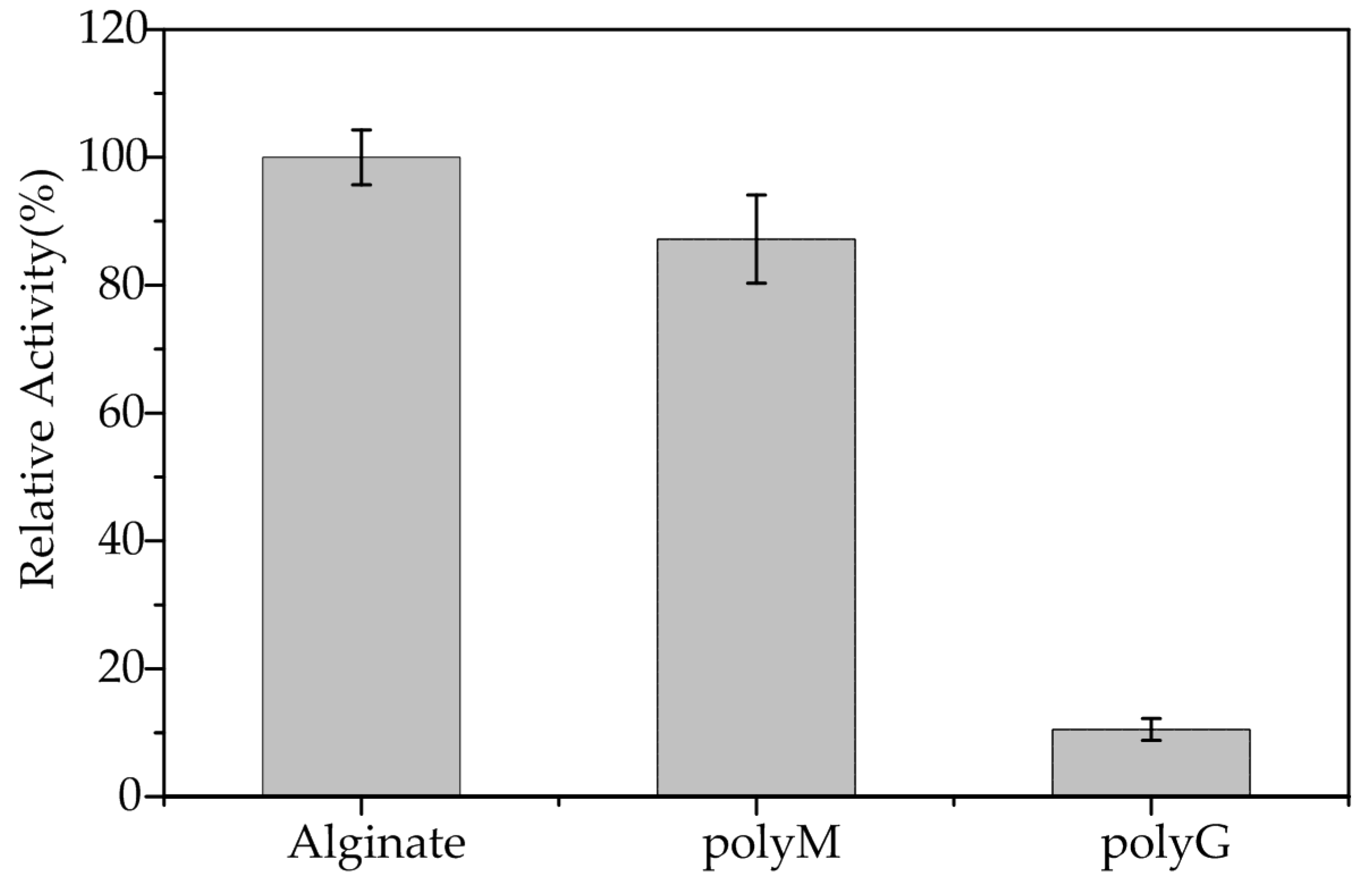
AlgA exhibited activity toward alginate but not toward pectin, hyaluronan, chitin, or agar. This behavior suggested that AlgA is indeed an alginate lyase. As shown in Figure 5, the relative activities of AlgA toward alginate, polyM, and polyG blocks are 100 ± 4.3, 87.2 ± 6.9, and 10.5 ± 1.7%, respectively. The slight activity toward polyG block might result from the presence of a few M residues in polyG substrates. These results indicated that the polyM block of substrates is the preferred substrate of AlgA. Thus, AlgA is a mannuronate lyase. Alginate lyases from Flavobacterium sp. UMI-01 [16], Vibrio sp. JAM-A9m [39], and Pseudomonas sp. QD03 [40] also belong to the mannuronate lyase class of enzymes.
The kinetic parameters of AlgA were determined through nonlinear regression analysis (Figure S3) and are shown in Table 3. AlgA has a lower Km value for polyM than for sodium alginate. This result suggested that AlgA has high affinity for polyM blocks and further confirmed that AlgA is a mannuronate lyase. However, the kcat values of AlgA for sodium alginate were higher than those for polyM. Therefore, AlgA has equivalent catalytic efficiency for sodium alginate and polyM.
2.5. Fast Protein Liquid Chromatography and Electrospray Ionization Mass Spectrometry Analysis of the Degradation Products of AlgA
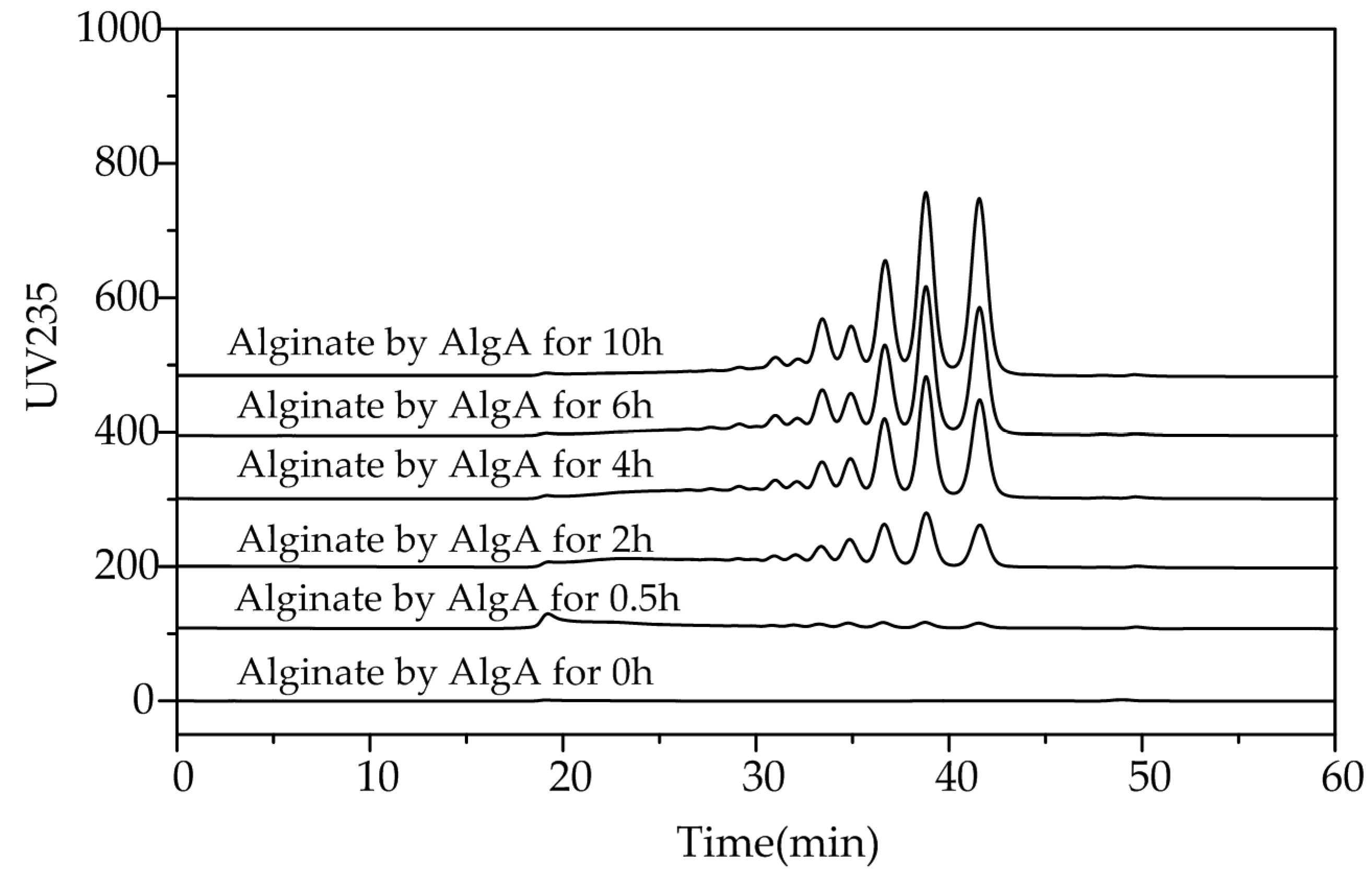
To investigate the action patterns of AlgA, degradation of alginate by this enzyme was performed. The degradation products at different time intervals were analyzed through gel chromatography. Fast protein liquid chromatography (FPLC) analysis indicated that AOS with different degrees of polymerization (DP) gradually accumulated (Figure 6). Therefore, AlgA is an endo-type alginate lyase.
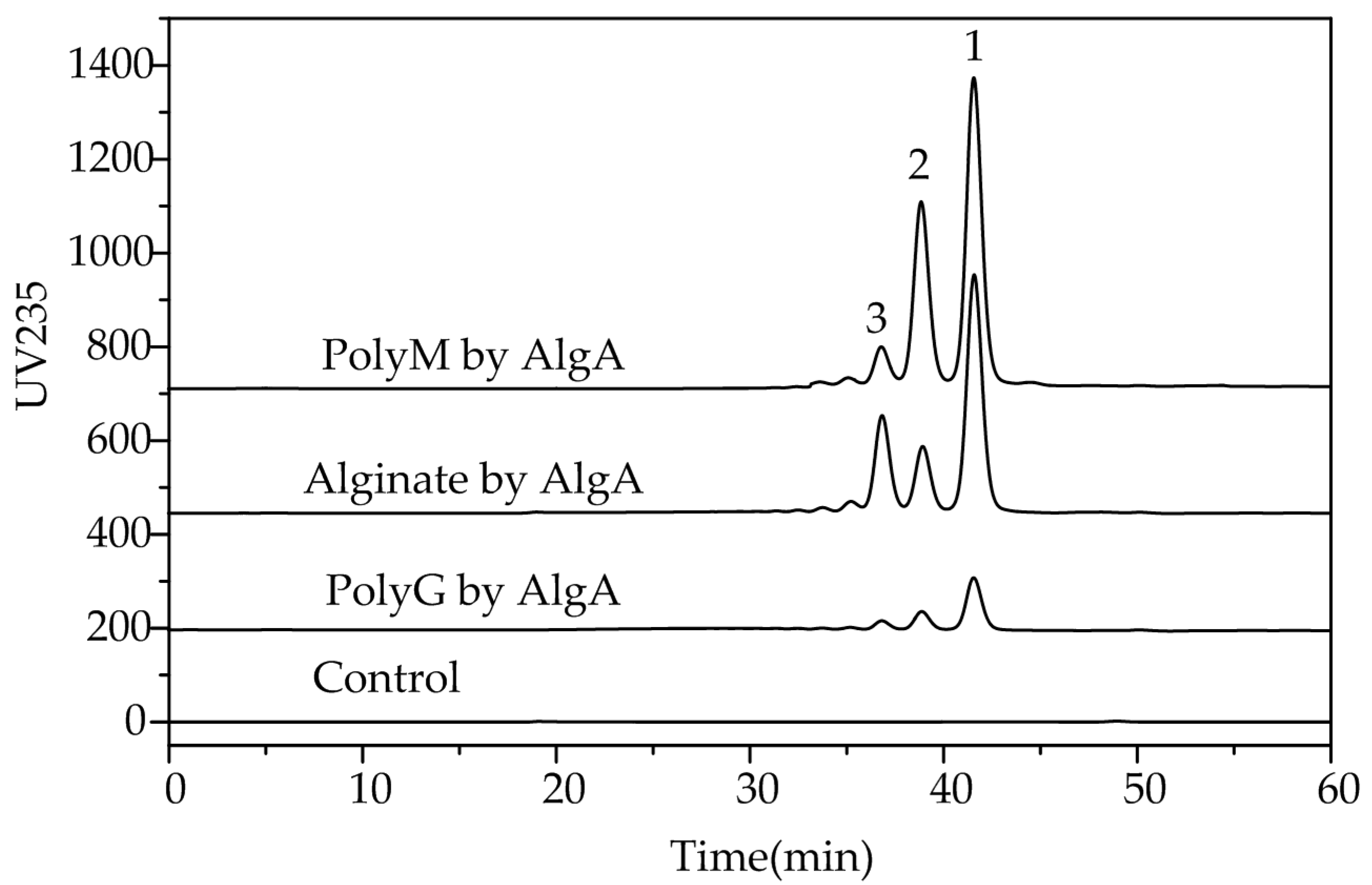
In addition, the alginate was completely digested with an excess of AlgA at 40 °C for 24 h. The products were separated through gel chromatography. The elution profiles (Figure 7) of the degradation products presented three major fractions (peaks 1, 2, and 3).
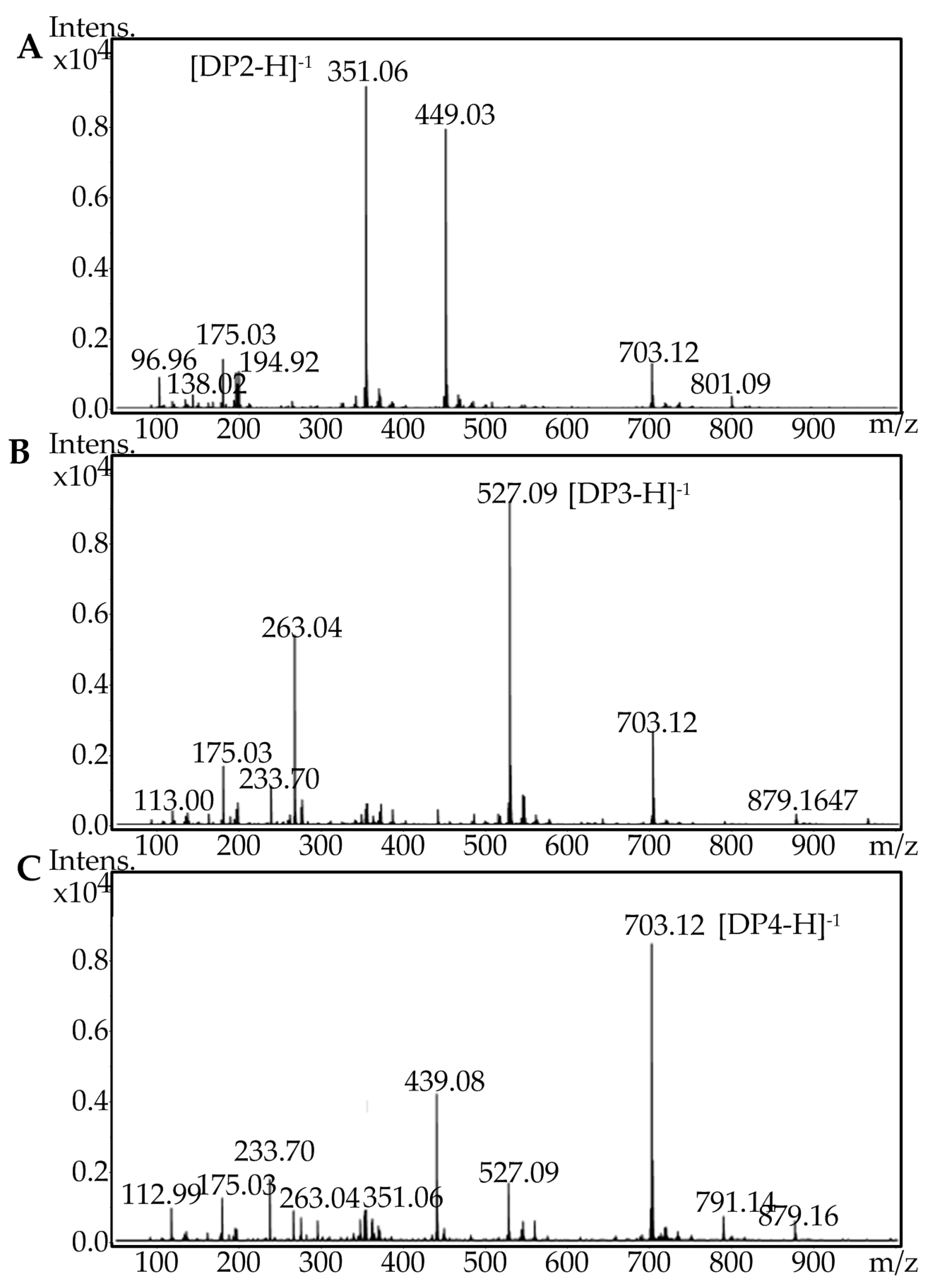
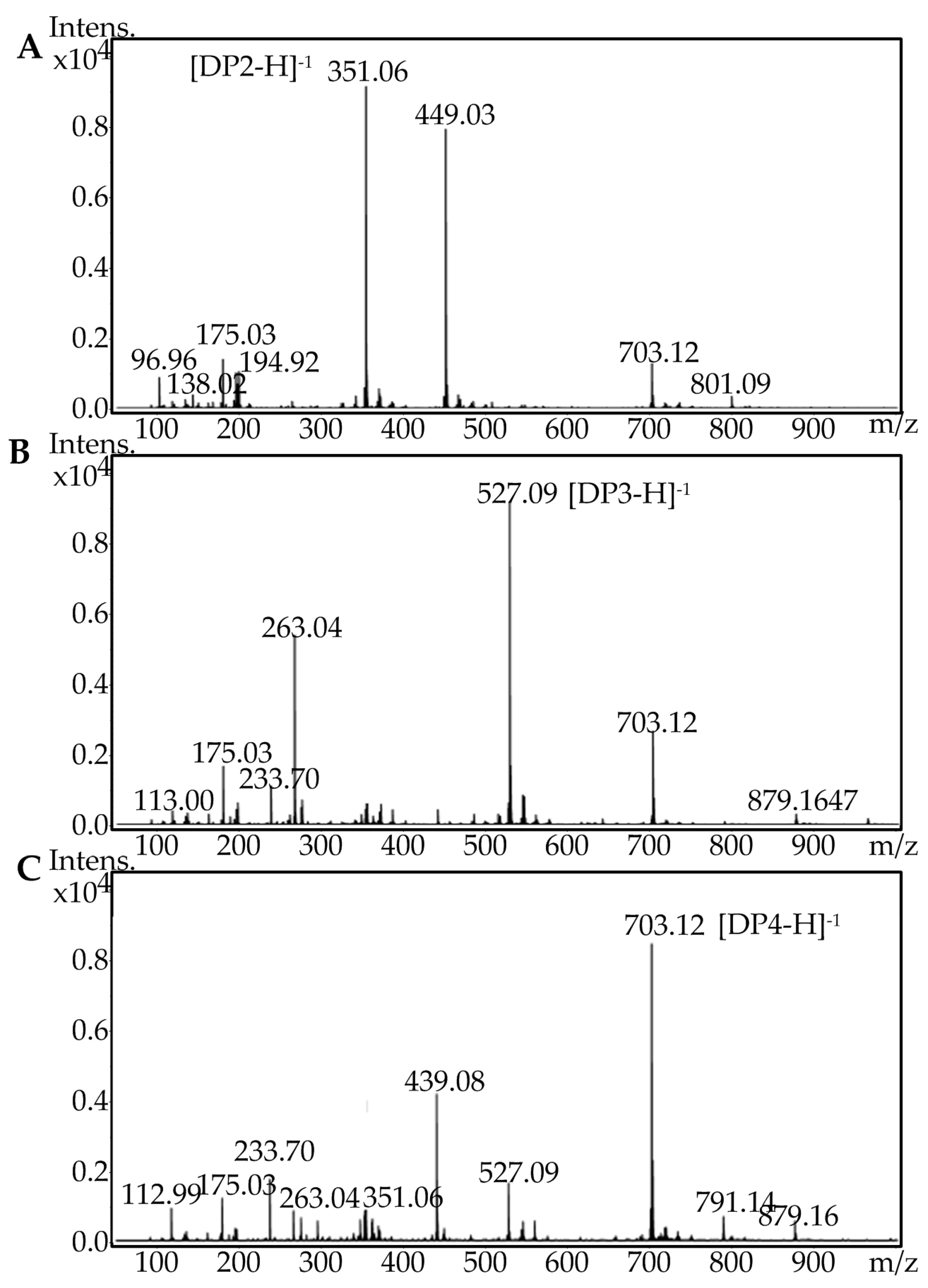
To identify the final oligosaccharide products of AlgA degradation and to determine their DP, three major fractions were subjected to electrospray ionization mass spectrometry (ESI-MS). The molecular masses of oligosaccharides in peaks 1, 2, and 3 were determined to be 351.06, 527.09, and 703.12, respectively (Figure 8). These results indicated that the main degradation products are di-, tri- and tetra-saccharides. The relative contents of di-, tri- and tetra-saccharides in alginate were 61.19%, 15.59%, and 23.22%, respectively, whereas those of di-, tri- and tetra-saccharides in the polyM substrate were 58.30%, 34.26%, and 7.43%, respectively. Meanwhile, AlgA has limited activity toward polyG. Therefore, AlgA may be used in the production of mannuronic oligosaccharides from polyM blocks and the preparation of polyG blocks from sodium alginate via the degradation of polyM blocks. The products of mannuronic oligosaccharides and polyG blocks possess special biological activity and have potential applications in many fields. For example, mannuronate oligosaccharides can promote the secretion of multiple cytokines [7], and polyG demonstrates higher macrophage-stimulation activity than polyM [41].
3. Materials and Methods
3.1. Materials
Sodium alginate derived from brown seaweed was purchased from Sigma (St. Louis, MO, USA). SOURCETM 15Q 4.6/100 PE and Superdex peptide 10/300 gel filtration columns were purchased from GE HealthCare Bio-Sciences (Uppsala, Sweden). DNA polymerase, protein molecular weight markers, and polyacrylamide were purchased from New England Biolabs (Ipswich, MA, USA). Other chemicals and reagents used in this study were of analytical grade.
3.2. Screening and Identification of Strain Alg07
Sea mud and rotten kelp samples were collected from a seaweed farm in Weihai, China. Five grams of the samples were added to 45 mL of modified marine broth 2216 medium containing 5 g/L (NH4)2SO4, 19.45 g/L NaCl, 12.6 g/L MgCl2·6H2O, 6.64 g/L MgSO4·7H2O, 0.55 g/L KCl, 0.16 g/L NaHCO3, 1 g/L ferric citrate, and 10 g/L sodium alginate. After 48 h of enrichment at 30 °C, the culture was serially diluted with deionized water and spread on modified marine broth 2216 agar containing 10 g/L sodium alginate. The plates were incubated at 30 °C for two days until colonies appeared. Single colonies were inoculated into marine broth 2216 medium containing 10 g/L sodium alginate and incubated for 48 h at 30 °C. Then, the activities of alginate lyase in the supernatants were determined. The alginate lyase from one of the isolates, strain Alg07, showed the highest activity among all lyases.
To identify the Alg07 strain, the 16S rRNA gene of the strain was amplified through PCR by using universal primers. The purified PCR fragment was sequenced and compared with reported 16s rRNA sequences in Genbank by using BLAST. A phylogenetic tree was constructed using CLUSTAL X and MEGA 6.0 [42] through neighbor-joining method [43].
3.3. Production and Purification of AlgA
Strain Alg07 was cultured for 24 h at 30 °C and 180 rpm in the optimized liquid medium, which contained 1g/L peptone, 3 g/L yeast extract, 9 g/L sodium alginate, 5 g/L NaCl, 1 g/L MgSO4·7H2O, 5 g/L KCl, and 4 g/L CaCl2 (pH 6.5). The supernatant was collected after 30 min of centrifugation at 10,000 rpm and 4 °C and then concentrated and desalted using a tangential flow filtration system (Vivaflow 50, Sartorius, Goettingen, Germany). The concentrated solution was subjected to AKTA FPLC (GE Healthcare Life Science, Marlborough, MA, USA) equipped with a SOURCETM (Barrie, ON, Canada) 15Q 4.6/100 PE column that had been equilibrated with 20 mM Tris–HCl buffer (pH 7.0). Adsorbed proteins were eluted with a linear gradient of 0–0.5 M NaCl in equilibrating buffer under a flow rate of 1 mL/min. Fractions possessing the highest specific activity among all fractions were pooled and dialyzed against 20 mM Tris-HCl buffer (pH 7.0) for further enzyme characterization. All purification procedures were performed at 4 °C. Protein concentration was determined with a protein quantitative analysis kit (Solarbio, Beijing, China) using bovine serum albumin as the calibration standard. The purity of the isolated alginate lyase was analyzed through 12.5% SDS-PAGE in accordance with the method of Laemmli (1970) [44].
3.4. Enzyme Activity Assay
To determine the activity of alginate lyase, 100 μL of appropriately diluted enzyme was added to 1900 μL of substrate solution containing 10 g/L sodium alginate, 20 mM Tris-HCl, and 200 mM NaCl (pH 7.5). The reaction was allowed to proceed for 20 min at 40 °C and terminated by the addition of 20 μL of 10 M NaOH. Absorbance at 235 nm was recorded. One unit (U) was defined as the amount of enzyme required to increase the absorbance at 235 nm by 0.1 per min. For kinetic parameter analysis, the 3,5-dinitrosalicylic acid method was performed to determine alginate lyase activity based on the release of reducing sugars from substrates [45]. One unit (U) was defined as the amount of enzyme required to release 1 μmol of reducing sugar per min. All enzyme reactions were performed in triplicate, and reaction parameters were expressed as mean ± standard deviation.
3.5. Characterization of AlgA
Enzyme reactions were carried out at different temperatures (25 °C–60 °C) to determine the effect of temperature on AlgA. To evaluate the thermal stability of AlgA, the enzyme was incubated for different intervals at 40 °C, 45 °C, and 50 °C. Then, the residual activities of the enzyme were tested. The activity of the enzyme stored at 4 °C was used to represent 100% enzyme activity. The effect of different pH values on AlgA was determined by calculating the residual activities of AlgA after 20 h of incubation at 4 °C in 20 mM sodium acetate (pH 3.0–6.0), Tris-HCl (pH 6.0–9.0), or glycine-NaOH (pH 9.0–11.0) buffers. The initial activities in different pH buffers represented 100% enzyme activity.
Enzyme reactions were performed in the presence of different concentrations of NaCl (0–500 mM) to evaluate the effect of NaCl on AlgA. To determine the effects of metal ions and EDTA on the activity of AlgA, the highest enzyme activity was considered as 100% enzyme activity. AlgA was subjected to an activity assay after 12 h of incubation at 4 °C in the presence of 2 mM of different metal ions and EDTA.
3.6. Analysis of the N-Terminal Amino Acid Sequence of AlgA
After SDS-PAGE, the protein band of AlgA was electro-transferred onto a polyvinylidene difluoride membrane (Imobulon; Millipore, Darmstadt, Germany). Amino acid sequences were determined with a PPSQ-31A protein sequencer (Shimadzu Corporation; Kyoto, Japan).
3.7. Substrate Specificity and Kinetic Parameters of AlgA
A standard enzymatic assay was performed to test the substrate preference of AlgA by using pectin, hyaluronan, chitin, agar, sodium alginate, polyM, and polyG as substrates. PolyM and polyG were prepared in accordance with the method of Haug et al. [46].
The kinetic parameters of AlgA toward alginate and polyM were determined by measuring the initial velocities of enzyme activity under various substrate concentrations and were calculated on the basis of the nonlinear regression fitting of the Michaelis–Menten equation using Prism 6 (GraphPad Software, Inc., La Jolla, CA, USA).
3.8. FPLC and ESI-MS Analysis of the Degradation Products of AlgA
To elucidate the mode of action of AlgA toward alginate, alginate degradation was performed at 40 °C with 10 g/L sodium alginate as a substrate. The reaction was initiated by the addition of 2 μg of purified AlgA in a 10 mL reaction volume. Reaction solutions were withdrawn at appropriate time intervals, and AlgA was inactivated by 5 min of boiling. The samples were analyzed through FPLC with a Superdex peptide 10/300 gel filtration column (GE Health, Marlborough, MA, USA) with 0.2 M ammonium bicarbonate as the mobile phase at a flow rate of 0.4 mL/min [34]. The reaction was monitored at 235 nm.
To determine the oligosaccharide compositions of the final digests, an excess of AlgA was used to completely degrade 10 g/L sodium alginate, polyM, or polyG. The reaction was carried out in a 10 mL reaction volume at 40 °C for 24 h and then terminated by boiling for 5 min. Oligosaccharides were separated through gel filtration as described above. Peak fractions containing unsaturated oligosaccharide products were collected and repeatedly freeze-dried to remove NH4HCO3 for ESI-MS analysis. The molecular weight of each oligosaccharide fraction was determined using the ESI-MS method on microTOF-Q II equipment (Bruker, Billerica, MA, USA) with the following conditions: capillary voltage of 4 kV, dry temperature of 180 °C, gas flow rate of 4.0 L/min, and scan range of 50–1000 m/z.
4. Conclusions
An alginate lyase–producing marine bacterium was isolated and identified as Bacillus sp. Alg07. AlgA, the alginate lyase derived from Bacillus sp. Alg07, was purified through the two simple steps of tangential flow filtration concentration and anion-exchange chromatography. The results of SDS-PAGE indicated that the molecular mass of AlgA is approximately 60 kDa. The optimal temperature and pH for the activity of AlgA is 40 °C and 7.5, respectively. The activity of AlgA is dependent on NaCl and is promoted by the addition of Mg2+ and Ca2+. Under optimal conditions, the specific activity of AlgA reaches up to 8306.7 U/mg, which is the highest activity recorded for all reported alginate lyases. Moreover, AlgA is stable over a broad pH range (5.0–10.0) and under its optimal temperature (40 °C). AlgA is an endolytic polyM-specific alginate lyase and mainly produces disaccharides, trisaccharides, and tetrasaccharides from alginate and disaccharides and trisaccharides from polyM. The highly efficient and thermostable AlgA can have potential applications in the production of mannuronic oligosaccharides and polyG blocks from alginate.
Supplementary Materials
The following are available online at https://www.mdpi.com/1660-3397/16/3/86/s1, Figure S1: Purification of alginate lyase by anion exchange chromatograph (Source 15Q); Figure S2. The N-terminal amino acid sequence of the purified AlgA; Figure S3. Effects of substrate concentration on the activity of AlgA.
Acknowledgments
This work was supported by the National key R&D Program of China (2017YFD200900), the Science Technology Planning Project of Tianjin (16YFXTNC00160), and Youth Innovation Promotion Association, Chinese Academy of Sciences (to Yueming Zhu).
Author Contributions
Peng Chen and Yueming Zhu conceived and designed the experiments; Peng Chen performed the experiments; Peng Chen and Yan Zeng analyzed the data; Yan Men contributed reagents/materials/analysis tools; Peng Chen wrote the paper. Yuanxia Sun edited the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gacesa, P. Enzymic degradation of alginates. Int. J. Biochem. 1992, 24, 545–552. [Google Scholar] [CrossRef]
- Mabeau, S.; Kloareg, B. Isolation and Analysis of the Cell Walls of Brown Algae: Fucus spiralis, F. ceranoides, F. vesiculosus, F. serratus, Bifurcaria bifurcata and Laminaria digitata. J. Exp. Bot. 1987, 38, 1573–1580. [Google Scholar] [CrossRef]
- Gorin, P.A.J.; Spencer, J.F.T. Exocellular alginic acid from Azotobacter vinelandii. Can. J. Chem. 1966, 44, 993–998. [Google Scholar] [CrossRef]
- Evans, L.R.; Linker, A. Production and Characterization of the Slime Polysaccharide of Pseudomonas aeruginosa. J. Bacteriol. 1973, 116, 915–924. [Google Scholar] [PubMed]
- Scott, C.D. Immobilized cells: A review of recent literature. Enzym. Microb. Technol. 1987, 9, 66–72. [Google Scholar] [CrossRef]
- Kawada, A.; Hiura, N.; Tajima, S.; Takahara, H. Alginate oligosaccharides stimulate VEGF-mediated growth and migration of human endothelial cells. Arch. Dermatol. Res. 1999, 291, 542–547. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Kurachi, M.; Yamaguchi, K.; Oda, T. Stimulation of multiple cytokine production in mice by alginate oligosaccharides following intraperitoneal administration. Carbohydr. Res. 2007, 342, 1133–1137. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, K.I.; Matsubara, Y. Purification of Alginate Oligosaccharides with Root Growth-promoting Activity toward Lettuce. Biosci. Biotechnol. Biochem. 2000, 64, 1067–1070. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Han, F.; Hu, B.; Li, J.; Yu, W. In vivo prebiotic properties of alginate oligosaccharides prepared through enzymatic hydrolysis of alginate. Nutr. Res. 2006, 26, 597–603. [Google Scholar] [CrossRef]
- Falkeborg, M.; Cheong, L.Z.; Gianfico, C.; Sztukiel, K.M.; Kristensen, K.; Glasius, M.; Xu, X.; Guo, Z. Alginate oligosaccharides: enzymatic preparation and antioxidant property evaluation. Food Chem. 2014, 164, 185–194. [Google Scholar] [CrossRef] [PubMed]
- An, Q.D.; Zhang, G.L.; Wu, H.T.; Zhang, Z.C.; Zheng, G.S.; Luan, L.; Murata, Y.; Li, X. Alginate-deriving oligosaccharide production by alginase from newly isolated Flavobacterium sp. LXA and its potential application in protection against pathogens. J. Appl. Microbiol. 2009, 106, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Lee, C.G.; Lee, E.Y. Alginate lyase: Structure, property, and application. Biotechnol. Bioprocess Eng. 2011, 16, 843–851. [Google Scholar] [CrossRef]
- Inoue, A.; Mashino, C.; Uji, T.; Saga, N.; Mikami, K.; Ojima, T. Characterization of an Eukaryotic PL-7 Alginate Lyase in the Marine Red AlgaPyropia yezoensis. Curr. Biotechnol. 2015, 4, 240–258. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Suzuki, K.; Inoue, A.; Ojima, T. A novel oligoalginate lyase from abalone, Haliotis discus hannai, that releases disaccharide from alginate polymer in an exolytic manner. Carbohydr. Res. 2006, 341, 1809–1819. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Chen, M.; Yin, H.; Du, Y.; Ning, L. Enzymatic Hydrolysis of Alginate to Produce Oligosaccharides by a New Purified Endo-Type Alginate Lyase. Mar. Drugs. 2016, 14, 108. [Google Scholar] [CrossRef] [PubMed]
- Inoue, A.; Takadono, K.; Nishiyama, R.; Tajima, K.; Kobayashi, T.; Ojima, T. Characterization of an Alginate Lyase, FlAlyA, from Flavobacterium sp. Strain UMI-01 and Its Expression in Escherichia coli. Mar. Drugs 2014, 12, 4693–4712. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Gupta, V.; Kumari, P.; Kumar, M.; Reddy, C.R.K.; Prasad, K.; Jha, B. Purification and partial characterization of an extracellular alginate lyase from Aspergillus oryzae isolated from brown seaweed. J. Appl. Phycol. 2011, 23, 755–762. [Google Scholar] [CrossRef]
- Suda, K.; Tanji, Y.; Hori, K.; Unno, H. Evidence for a novel Chlorella virus-encoded alginate lyase. FEMS Microbiol. Lett. 1999, 180, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Wong, T.Y.; Preston, L.A.; Schiller, N.L. ALGINATE LYASE: Review of major sources and enzyme characteristics, structure-function analysis, biological roles, and applications. Annu. Rev. Microbiol. 2000, 54, 289–340. [Google Scholar] [CrossRef] [PubMed]
- Cantarel, B.L.; Coutinho, P.M.; Rancurel, C.; Bernard, T.; Lombard, V.; Henrissat, B. The Carbohydrate-Active EnZymes database (CAZy): an expert resource for Glycogenomics. Nucleic Acids Res. 2009, 37, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Aarstad, O.A.; Tøndervik, A.; Sletta, H.; Skjåkbræk, G. Alginate Sequencing: An Analysis of Block Distribution in Alginates Using Specific Alginate Degrading Enzymes. Biomacromolecules 2012, 13, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Inoue, A.; Mashino, C.; Kodama, T.; Ojima, T. Protoplast preparation from Laminaria japonica with recombinant alginate lyase and cellulase. Mar. Biotechnol. 2011, 13, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Islan, G.A.; Martinez, Y.N.; Illanes, A.; Castro, G.R. Development of novel alginate lyase cross-linked aggregates for the oral treatment of cystic fibrosis. RSC Adv. 2014, 4, 11758–11765. [Google Scholar] [CrossRef]
- Li, L.; Jiang, X.; Guan, H.; Wang, P. Preparation, purification and characterization of alginate oligosaccharides degraded by alginate lyase from Pseudomonas sp. HZJ 216. Carbohydr. Res. 2011, 346, 794–800. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, S.; Yu, W.; Gong, Q. Cloning, overexpression and characterization of a new oligoalginate lyase from a marine bacterium, Shewanella sp. Biotechnol. Lett. 2015, 37, 665–671. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.T.; Ko, H.J.; Kim, N.; Kim, D.; Lee, D.; Choi, I.G.; Woo, H.C.; Kim, M.D.; Kim, K.H. Characterization of a recombinant endo-type alginate lyase (Alg7D) from Saccharophagus degradans. Biotechnol. Lett. 2012, 34, 1087–1092. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, L.; Hao, J.; Xing, M.; Sun, J.; Sun, M. Purification and Characterization of a New Alginate Lyase from Marine Bacterium Vibrio sp. SY08. Mar. Drugs 2017, 15, 1. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Guo, E.W.; Yu, W.G.; Han, F. Purification and characterization of a new alginate lyase from a marine bacterium Vibrio sp. Biotechnol. Lett. 2013, 35, 703–708. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Uchimura, K.; Miyazaki, M.; Nogi, Y.; Horikoshi, K. A new high-alkaline alginate lyase from a deep-sea bacterium Agarivorans sp. Extremophiles 2009, 13, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Xiao, T.F.; Hong, L.; Sang, M.K. Purification and characterization of a Na+/K+ dependent alginate lyase from turban shell gut Vibrio sp. YKW-34. Enzym. Microb. Technol. 2007, 41, 828–834. [Google Scholar]
- Zhu, B.; Yin, H. Alginate lyase: Review of major sources and classification, properties, structure-function analysis and applications. Bioengineered 2015, 6, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, M.; Moriwaki, S.; Miyake, O.; Hashimoto, W.; Murata, K.; Mikami, B. Structure and function of a hypothetical Pseudomonas aeruginosa protein PA1167 classified into family PL-7. J. Biol. Chem. 2004, 279, 31863–31872. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.I.; Choi, S.H.; Lee, E.Y.; Kim, H.S. Molecular cloning, purification, and characterization of a novel polyMG-specific alginate lyase responsible for alginate MG block degradation in Stenotrophomas maltophilia KJ-2. Appl. Microbiol. Biotechnol. 2012, 95, 1643–1653. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Tan, H.; Qin, Y.; Xu, Q.; Du, Y.; Yin, H. Characterization of a new endo-type alginate lyase from Vibrio sp. W13. Int. J. Biol. Macromol. 2015, 75, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Thomas, F.; Lundqvist, L.C.E.; Jam, M.; Jeudy, A.; Barbeyron, T.; Sandström, C.; Michel, G.; Czjzek, M. Comparative Characterization of Two Marine Alginate Lyases from Zobellia galactanivorans Reveals Distinct Modes of Action and Exquisite Adaptation to Their Natural Substrate. J. Biol. Chem. 2013, 288, 23021–23037. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yang, X.; Bao, M.; Wu, Y.; Yu, W.; Han, F. Family 13 carbohydrate-binding module of alginate lyase from Agarivorans sp. L11 enhances its catalytic efficiency and thermostability, and alters its substrate preference and product distribution. FEMS Microbiol. Lett. 2015, 362. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Gu, J.; Cheng, Y.; Liu, H.; Li, Y.; Li, F. Novel Alginate Lyase (Aly5) from a Polysaccharide-Degrading Marine Bacterium, Flammeovirga sp. Strain MY04: Effects of Module Truncation on Biochemical Characteristics, Alginate Degradation Patterns, and Oligosaccharide-Yielding Properties. Appl. Environ. Microbiol. 2015, 82, 364–374. [Google Scholar] [CrossRef] [PubMed]
- Li, J.W.; Dong, S.; Song, J.; Li, C.B.; Chen, X.L.; Xie, B.B.; Zhang, Y.Z. Purification and Characterization of a Bifunctional Alginate Lyase from Pseudoalteromonas sp. SM0524. Mar. Drugs 2011, 9, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Uchimura, K.; Miyazaki, M.; Nogi, Y.; Kobayashi, T.; Horikoshi, K. Cloning and sequencing of alginate lyase genes from deep-sea strains of Vibrio and Agarivorans and characterization of a new Vibrio enzyme. Mar. Biotechnol. 2010, 12, 526–533. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Han, F.; Yang, Z.; Lu, X.Z.; Yu, W.G. A Novel Alginate Lyase with High Activity on Acetylated Alginate of Pseudomonas aeruginosa FRD1 from Pseudomonas sp. QD03. World J. Microbiol. Biotechnol. 2006, 22, 81–88. [Google Scholar] [CrossRef]
- Ueno, M.; Hiroki, T.; Takeshita, S.; Jiang, Z.; Kim, D.; Yamaguchi, K.; Oda, T. Comparative study on antioxidative and macrophage-stimulating activities of polyguluronic acid (PG) and polymannuronic acid (PM) prepared from alginate. Carbohydr. Res. 2012, 352, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Biochem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Haug, A.; Larsen, B. A study on the constitution of alginic acid by partial acid hydrolysis. Acta Chem. Scand. 1966, 20, 183–190. [Google Scholar] [CrossRef]
Figure 1.
Neighbor-joining phylogenetic tree generated on the basis of the 16S rRNA gene sequences of strain Alg07 and other known Bacillus species.
Figure 1.
Neighbor-joining phylogenetic tree generated on the basis of the 16S rRNA gene sequences of strain Alg07 and other known Bacillus species.
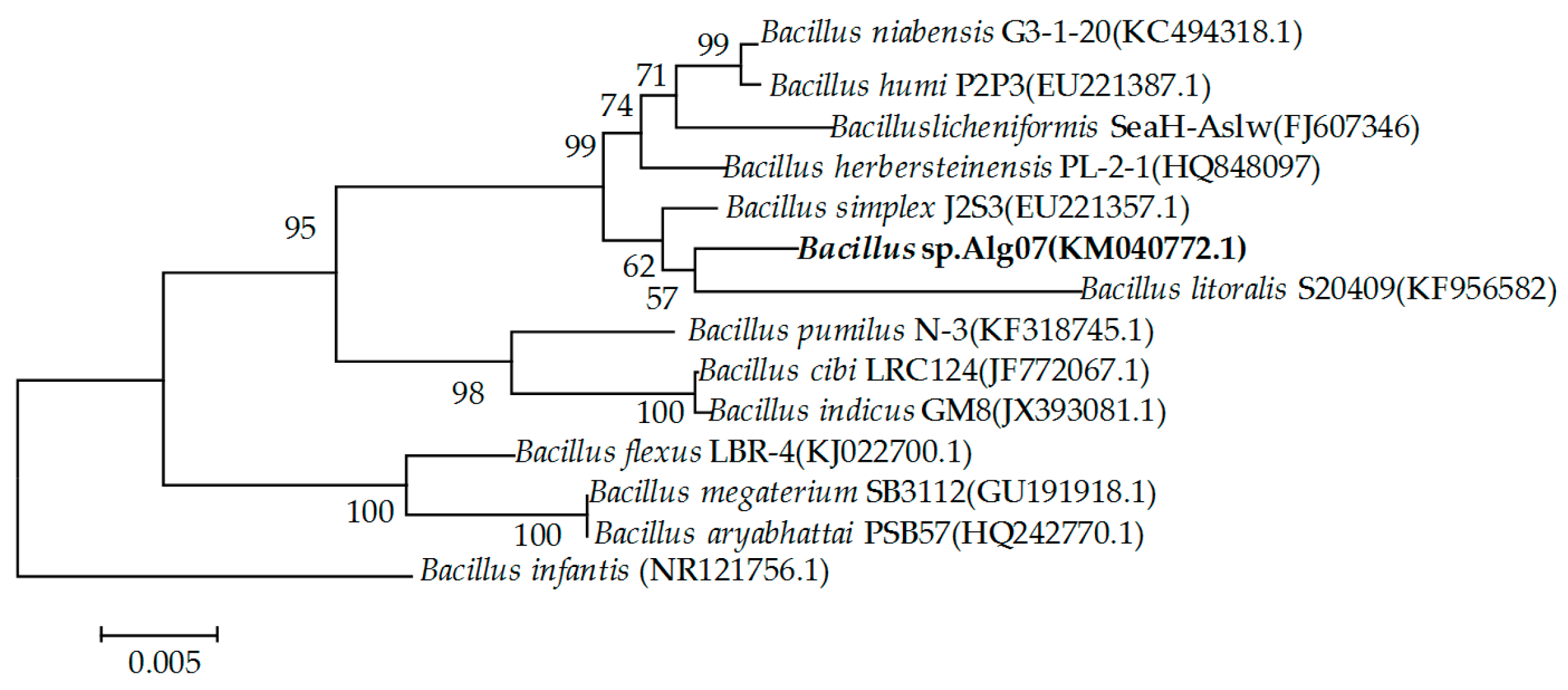
Figure 2.
Sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) result for AlgA. Lane M, protein ladder; Lane 1, purified AlgA.
Figure 2.
Sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) result for AlgA. Lane M, protein ladder; Lane 1, purified AlgA.
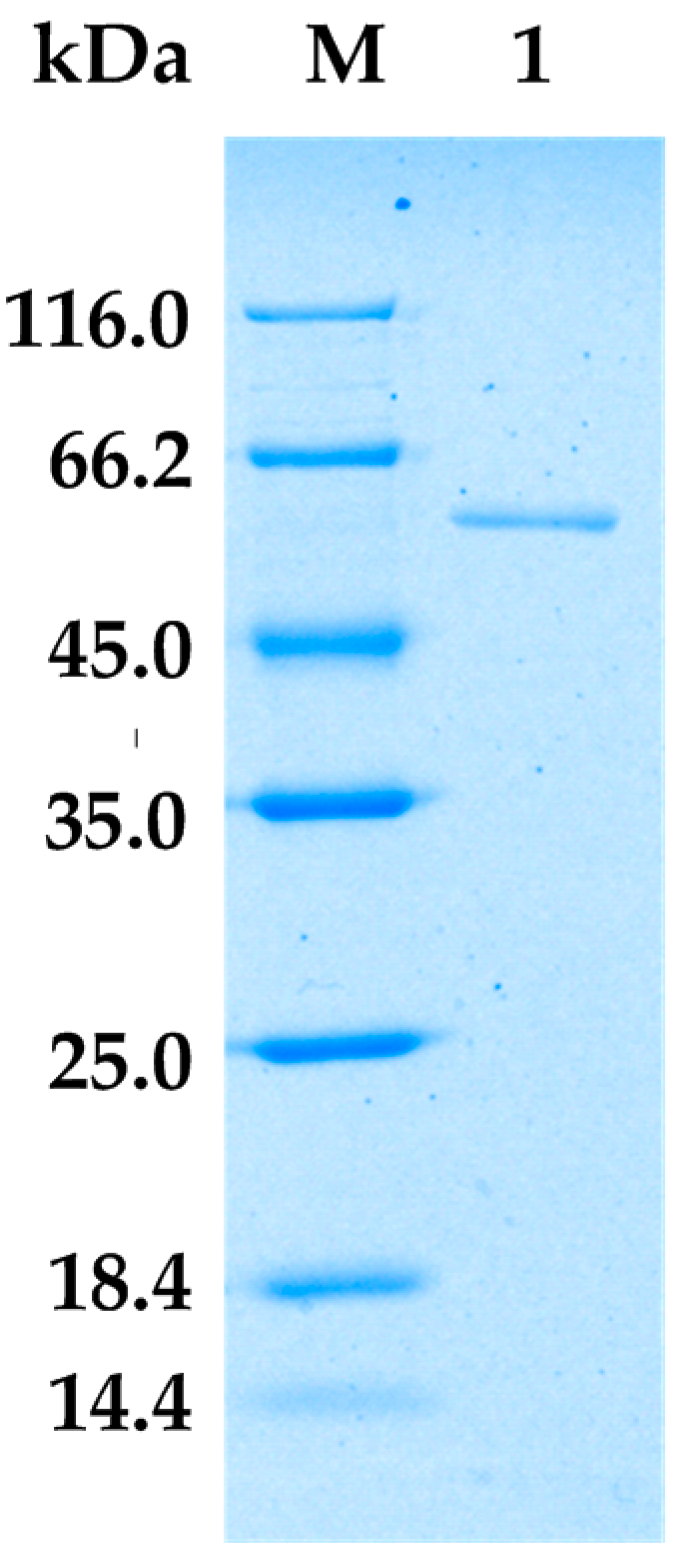
Figure 3.
Effects of temperature and pH on the relative activity of AlgA. (A) Optimal temperature of AlgA. (B) Thermostability of AlgA at 40 °C (filled square), 45 °C (filled circle), and 50 °C (filled triangle). (C) Optimal pH for the relative activity of AlgA was determined in 20 mM CH3COOH-CH3COONa buffer (filled square), 20 mM Tris-HCl buffer (filled circle), or 20 mM Glycine-NaOH buffer (filled triangle). (D) pH stability of AlgA in 20 mM CH3COOH-CH3COONa buffer (filled square), 20 mM Tris-HCl (filled circle), and 20 mM Glycine-NaOH (filled triangle).
Figure 3.
Effects of temperature and pH on the relative activity of AlgA. (A) Optimal temperature of AlgA. (B) Thermostability of AlgA at 40 °C (filled square), 45 °C (filled circle), and 50 °C (filled triangle). (C) Optimal pH for the relative activity of AlgA was determined in 20 mM CH3COOH-CH3COONa buffer (filled square), 20 mM Tris-HCl buffer (filled circle), or 20 mM Glycine-NaOH buffer (filled triangle). (D) pH stability of AlgA in 20 mM CH3COOH-CH3COONa buffer (filled square), 20 mM Tris-HCl (filled circle), and 20 mM Glycine-NaOH (filled triangle).
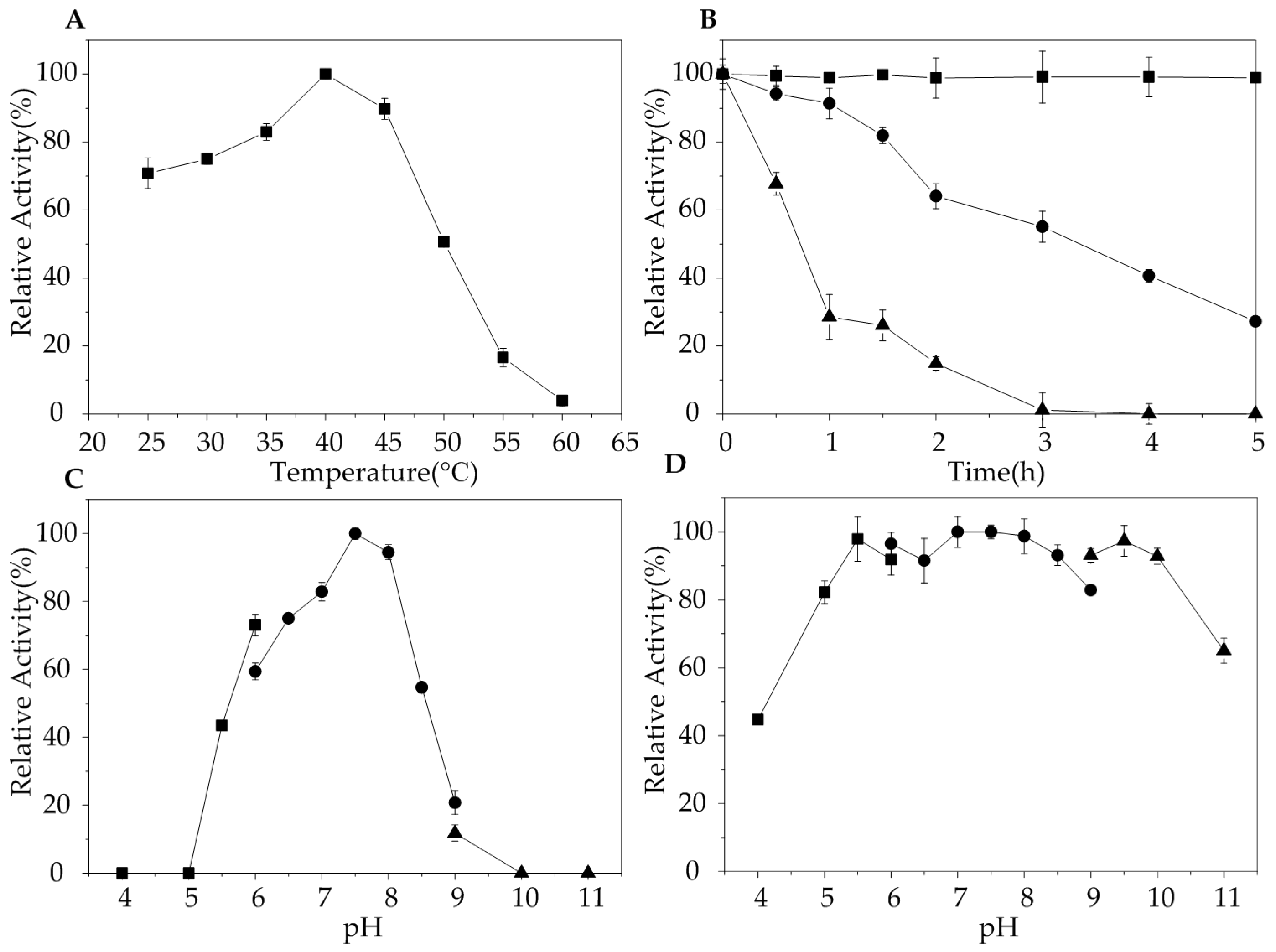
Figure 4.
Effect of NaCl (A) and metal ions (B) on the activity of AlgA.
Figure 5.
Relative activities of AlgA toward alginate, polyM, and polyG.
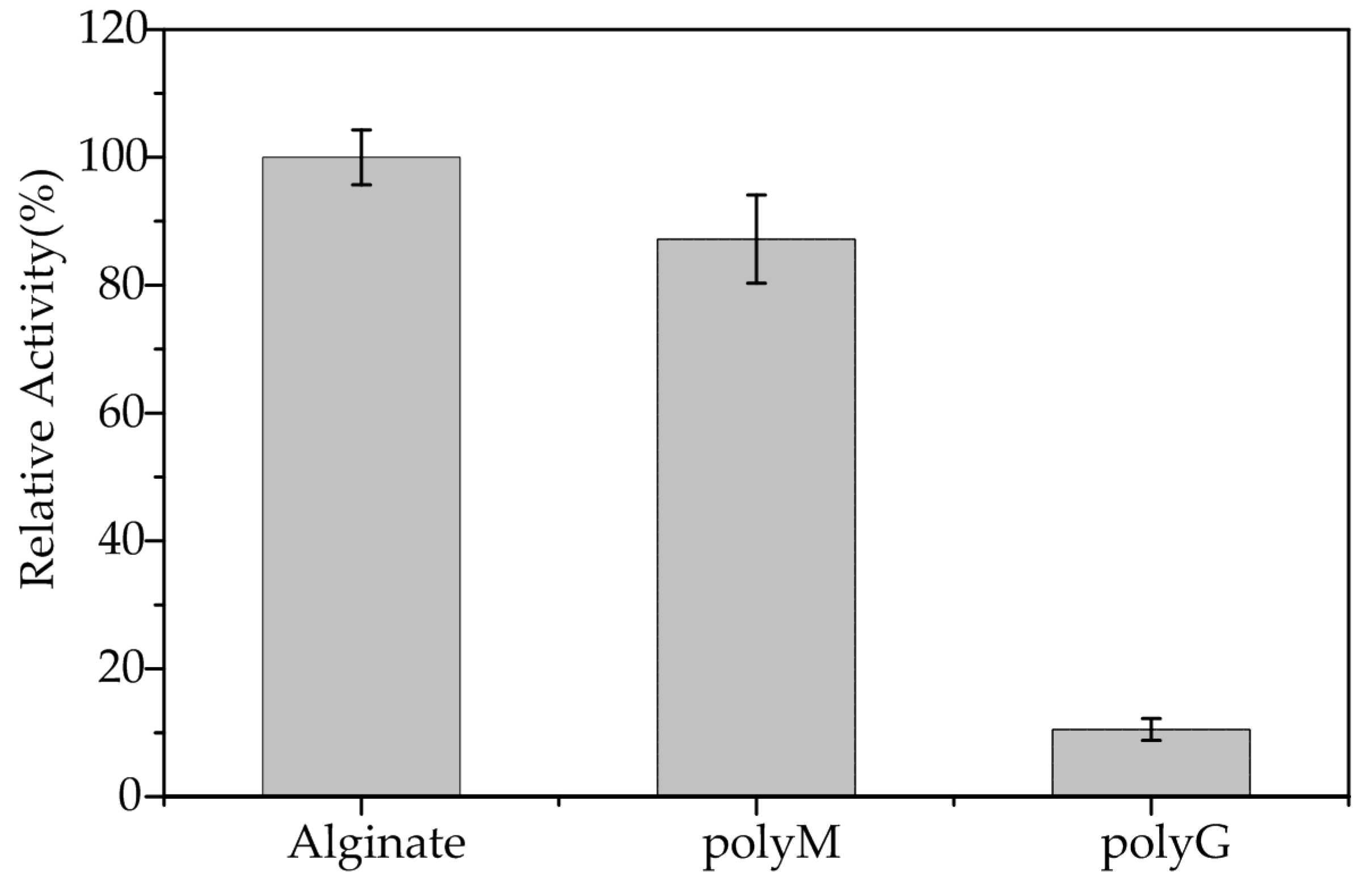
Figure 6.
Patterns of the polysaccharide degradation products of AlgA. Enzymatic degradation products collected at 0.5, 2, 4, 6, and 10 h were subjected to gel filtration with a Superdex peptide 10/300 GL column. The absorbances of the products were monitored at 235 nm.
Figure 6.
Patterns of the polysaccharide degradation products of AlgA. Enzymatic degradation products collected at 0.5, 2, 4, 6, and 10 h were subjected to gel filtration with a Superdex peptide 10/300 GL column. The absorbances of the products were monitored at 235 nm.
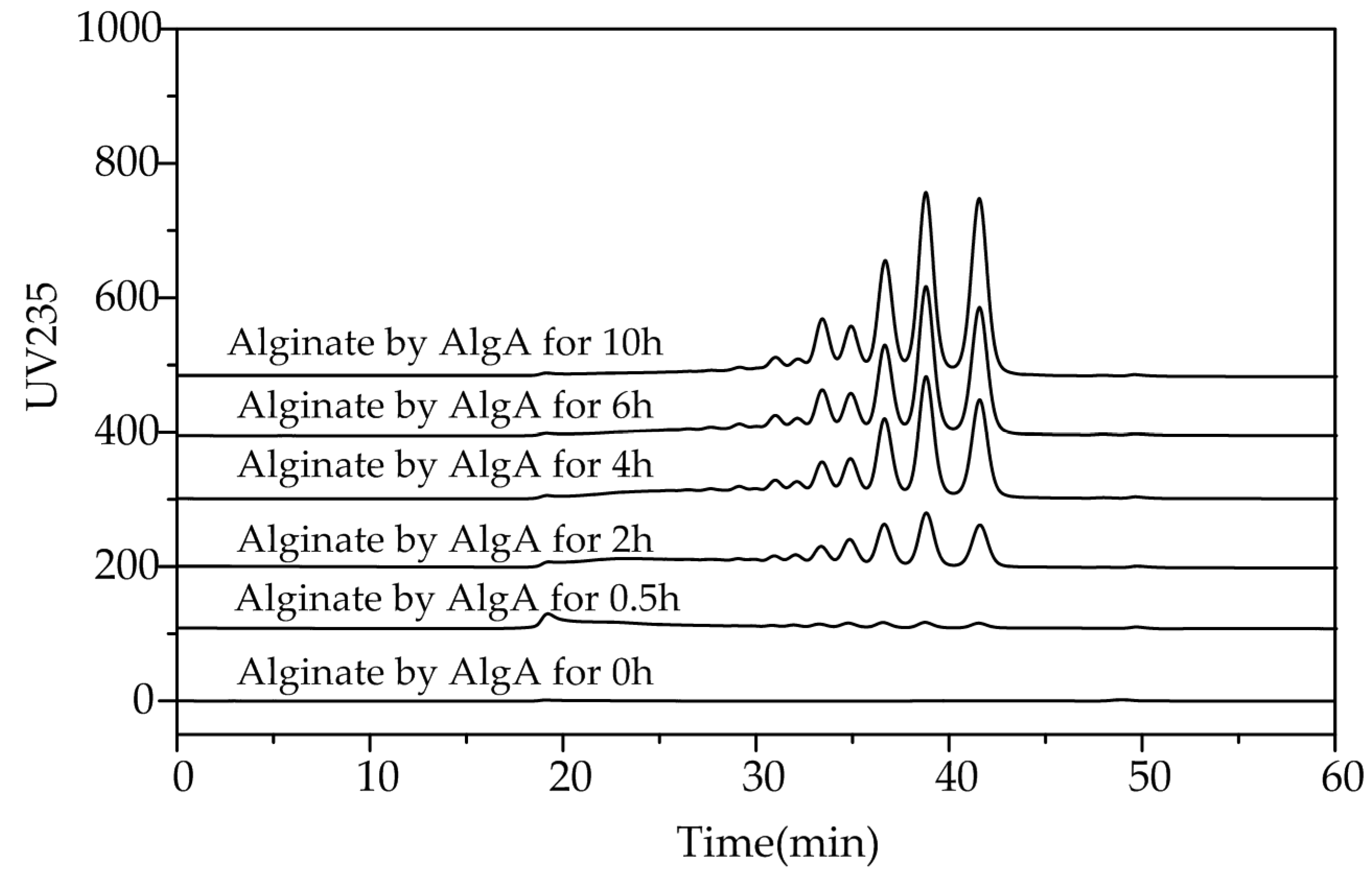
Figure 7.
Final products of alginate, polyM, and polyG after degradation by AlgA. Oligosaccharide products were gel-filtered through a Superdex peptide 10/300 GL column and monitored at a wavelength of 235 nm.
Figure 7.
Final products of alginate, polyM, and polyG after degradation by AlgA. Oligosaccharide products were gel-filtered through a Superdex peptide 10/300 GL column and monitored at a wavelength of 235 nm.
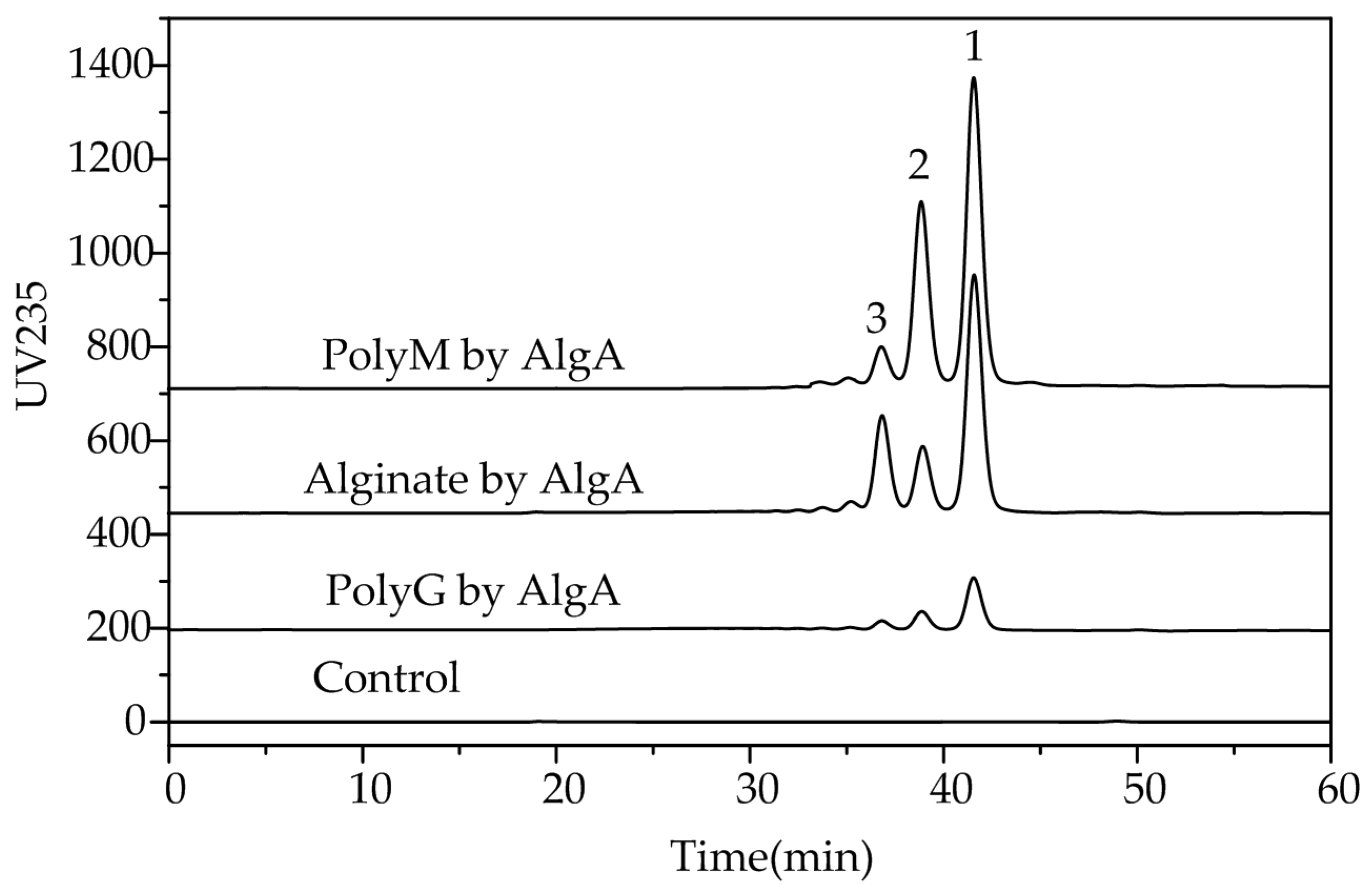
Figure 8.
Electrospray ionization mass spectrometry (ESI-MS) analysis of the final oligosaccharide products. (A) Fraction peak 1 separated through fast protein liquid chromatography (FPLC), (B) fraction peak 2 separated through FPLC, and (C) fraction peak 3 separated through FPLC.
Figure 8.
Electrospray ionization mass spectrometry (ESI-MS) analysis of the final oligosaccharide products. (A) Fraction peak 1 separated through fast protein liquid chromatography (FPLC), (B) fraction peak 2 separated through FPLC, and (C) fraction peak 3 separated through FPLC.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of the purification of AlgA.
| Step | Total Protein (mg) | Total Activity (U) | Specific Activity (U/mg) | Folds | Yield (%) |
|---|---|---|---|---|---|
| Culture broth | 23.82 | 23,724.72 | 996.01 | 1.00 | 100 |
| Vivaflow50 | 10.30 | 22,672.14 | 2201.18 | 2.21 | 95.4 |
| Source 15Q | 1.78 | 14,785.9 | 8306.71 | 8.34 | 62.4 |
Table 2.
Comparison of the properties of AlgA with those of alginate lyases from different microorganisms.
Table 2.
Comparison of the properties of AlgA with those of alginate lyases from different microorganisms.
| Enzyme | Source | Specific Activity (U/mg) | Molecular Mass (kDa) | Optimal Temperature (°C) | Optimal pH | Cation Activators | Substrate Specificity |
|---|---|---|---|---|---|---|---|
| AlgA | This study | 8306.71 | 60 | 40 | 7.5 | Na+, Mg2+, Ca2+, Mn2+ | PM |
| Oal17 | Shewanella sp.Kz7 [25] | 32 | 82 | 50 | 6.2 | Na+ | PM |
| Cel32 | Cellulophaga sp. NJ-1 [15] | 2417.8 | 32 | 50 | 8.0 | Ca2+, Mg2+, K+ | PM, PG |
| Alg7D | Saccharophagus degradans [26] | 4.6 | 63.2 | 50 | 7.0 | Na+ | PM, PG |
| AlySY08 | Vibrio sp. Aly08 [27] | 1070.2 | 33 | 40 | 7.6 | Na+, K+, Ca2+, Mg2+ | PM, PG |
| AlyV5 | Vibrio sp. QY105 [28] | 2152 | 37 | 38 | 7.0 | Na+, Mg2+, Ca2+, Mn2+ | PM, PG |
| Alm | Agarivorans sp. JAM-Alm [29] | 108.5 | 31 | 30 | 10.0 | Na+, K+ | PG, PMG |
| FlAlyA | Flavabacterium sp. UMI-01 [16] | 2347.8 | 30 | 55 | 7.7 | Na+, K+, Ca2+, Mg2+ | PM |
Table 3.
Kinetic parameters of the activity of AlgA toward sodium alginate and polyM blocks.
| Parameter | Sodium Alginate | PolyM |
|---|---|---|
| Vmax (U mg of protein−1) | 1052.0 ± 214.6 | 547.6 ± 22.4 |
| Km (mg mL−1) | 9.0 ± 3.3 | 3.6 ± 0.4 |
| kcat (s−1) | 911.7 ± 185.9 | 474.6 ± 19.4 |
| kcat/Km (mg−1 mL s−1) | 101.3 ± 20.7 | 132.0 ± 5.4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chen, P.; Zhu, Y.; Men, Y.; Zeng, Y.; Sun, Y. Purification and Characterization of a Novel Alginate Lyase from the Marine Bacterium Bacillus sp. Alg07. Mar. Drugs 2018, 16, 86. https://doi.org/10.3390/md16030086
AMA Style
Chen P, Zhu Y, Men Y, Zeng Y, Sun Y. Purification and Characterization of a Novel Alginate Lyase from the Marine Bacterium Bacillus sp. Alg07. Marine Drugs. 2018; 16(3):86. https://doi.org/10.3390/md16030086
Chicago/Turabian StyleChen, Peng, Yueming Zhu, Yan Men, Yan Zeng, and Yuanxia Sun. 2018. "Purification and Characterization of a Novel Alginate Lyase from the Marine Bacterium Bacillus sp. Alg07" Marine Drugs 16, no. 3: 86. https://doi.org/10.3390/md16030086
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.