Frondanol, a Nutraceutical Extract from Cucumaria frondosa, Attenuates Colonic Inflammation in a DSS-Induced Colitis Model in Mice

Abstract
:1. Introduction
2. Results
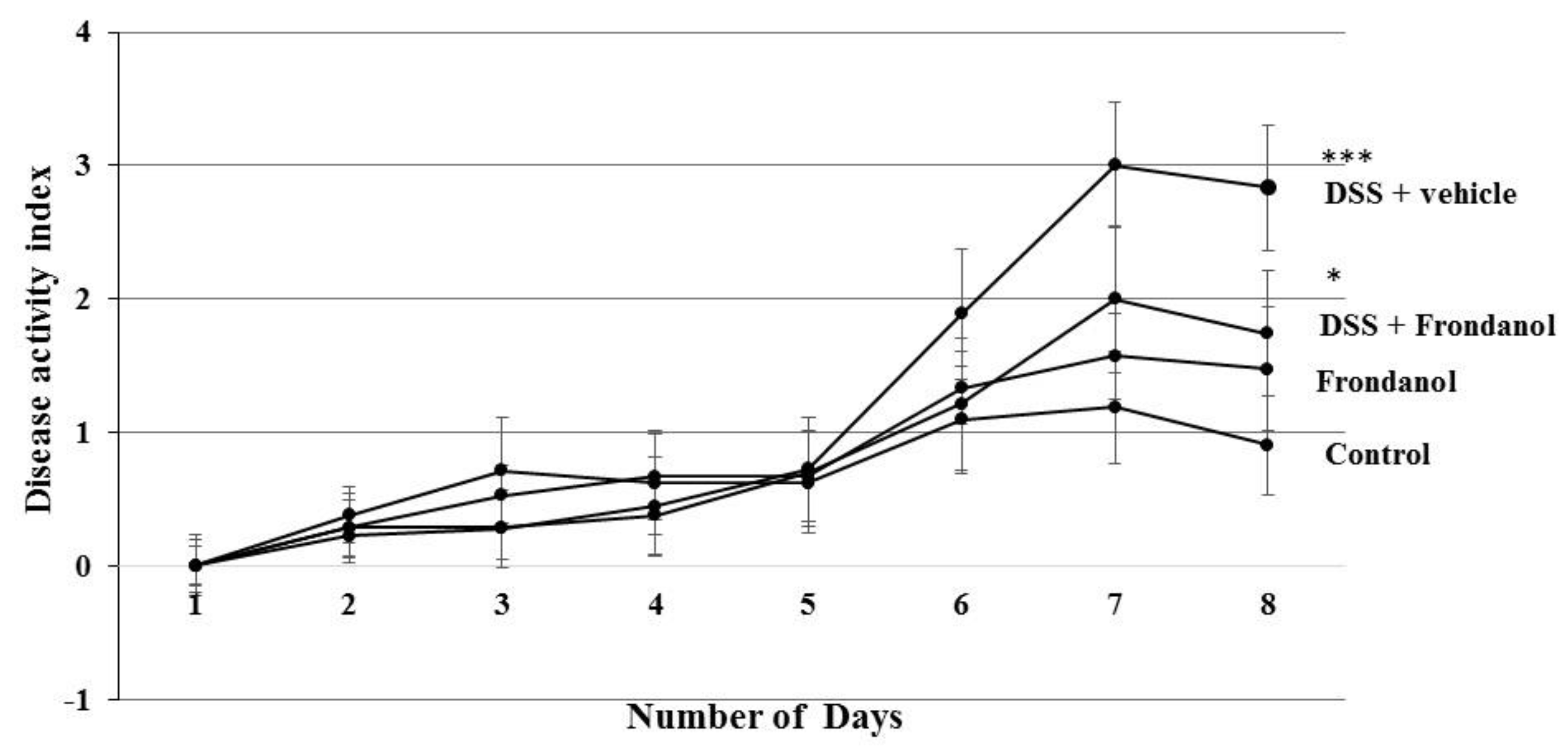
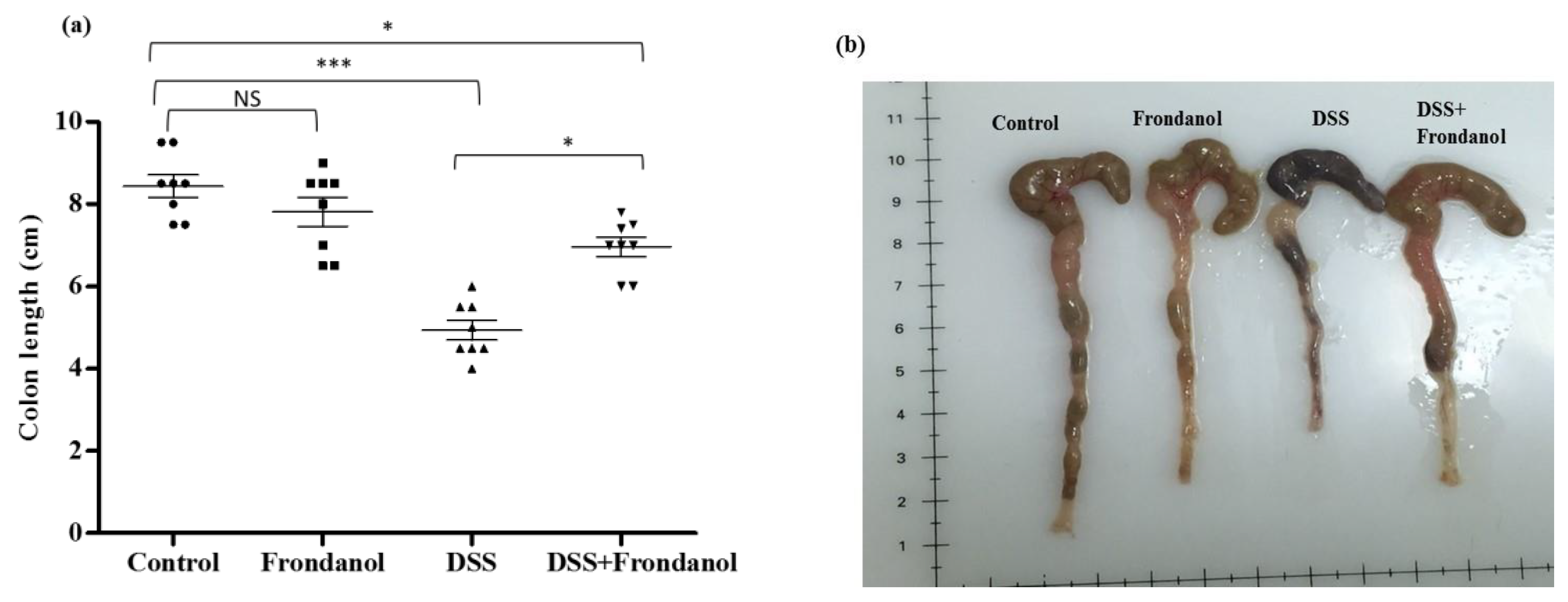
2.1. Effect of Frondanol on Disease Activity Index (DAI) and Colon Length
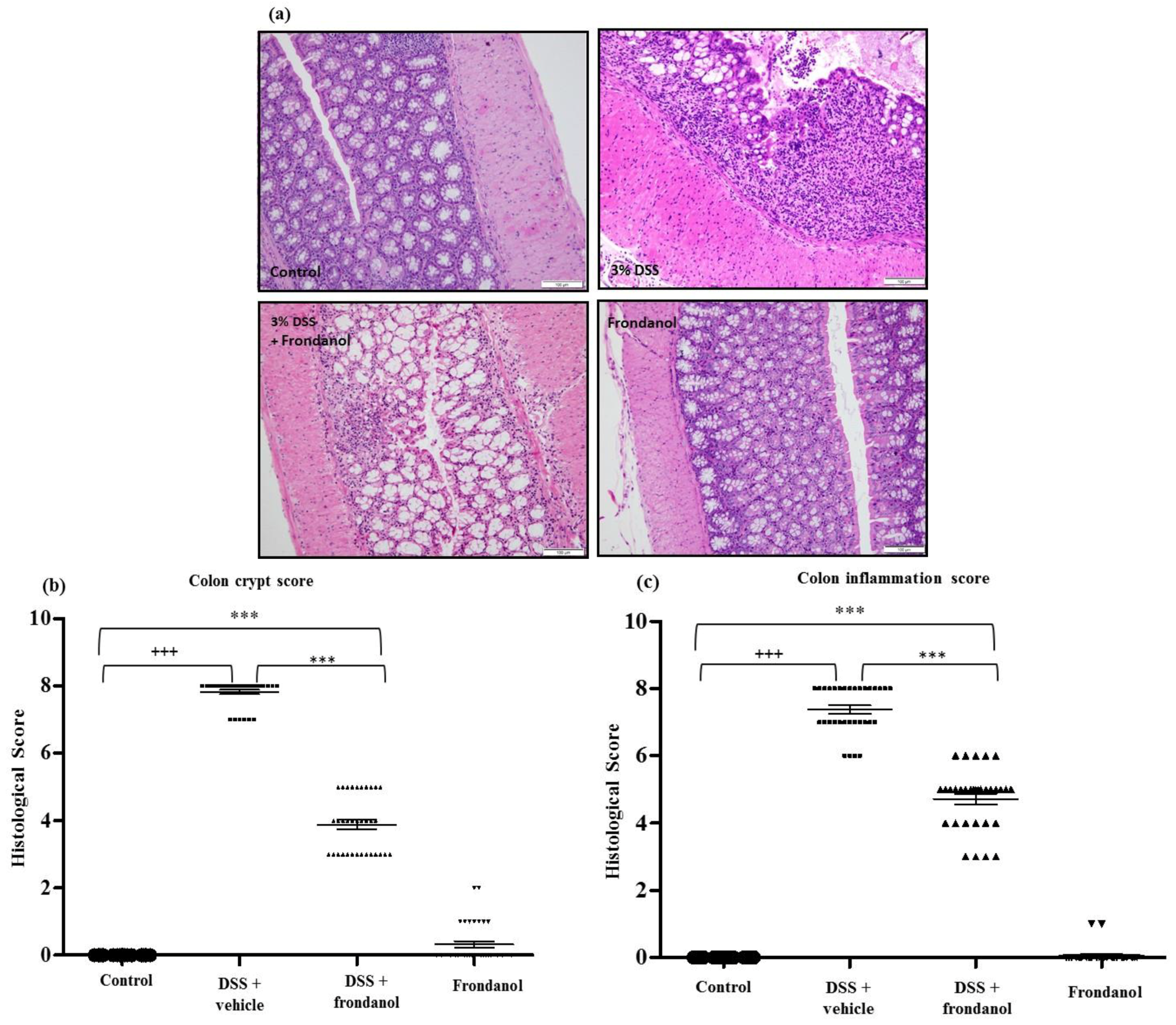
2.2. Effect of Frondanol on Microscopic Architecture
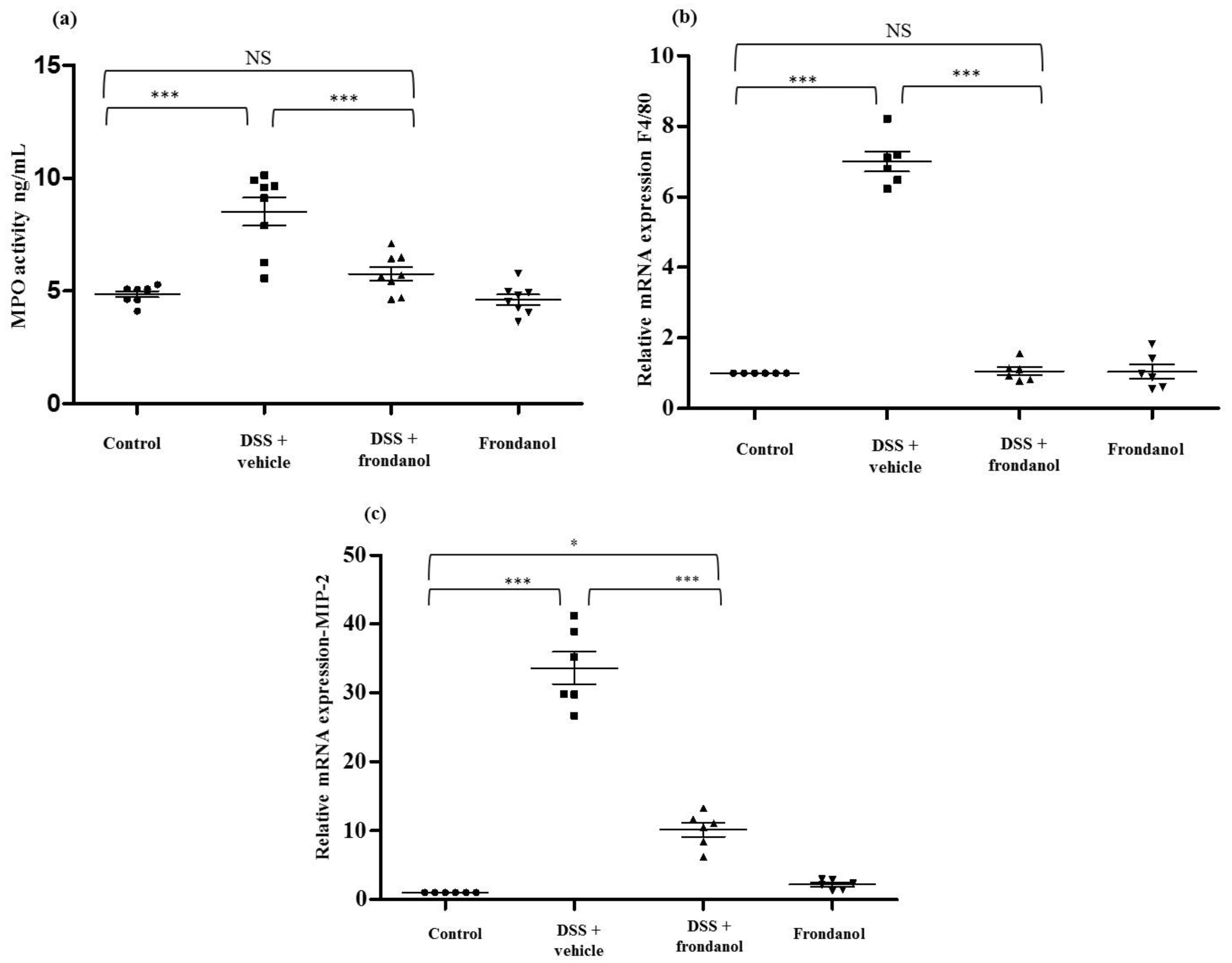
2.3. Effect of Frondanol on the Expression of Myeloperoxidase (MPO) Protein, F4/80 Macrophage Marker mRNA, and Macrophage Inflammatory Protein-2 (MIP-2) mRNA Concentrations
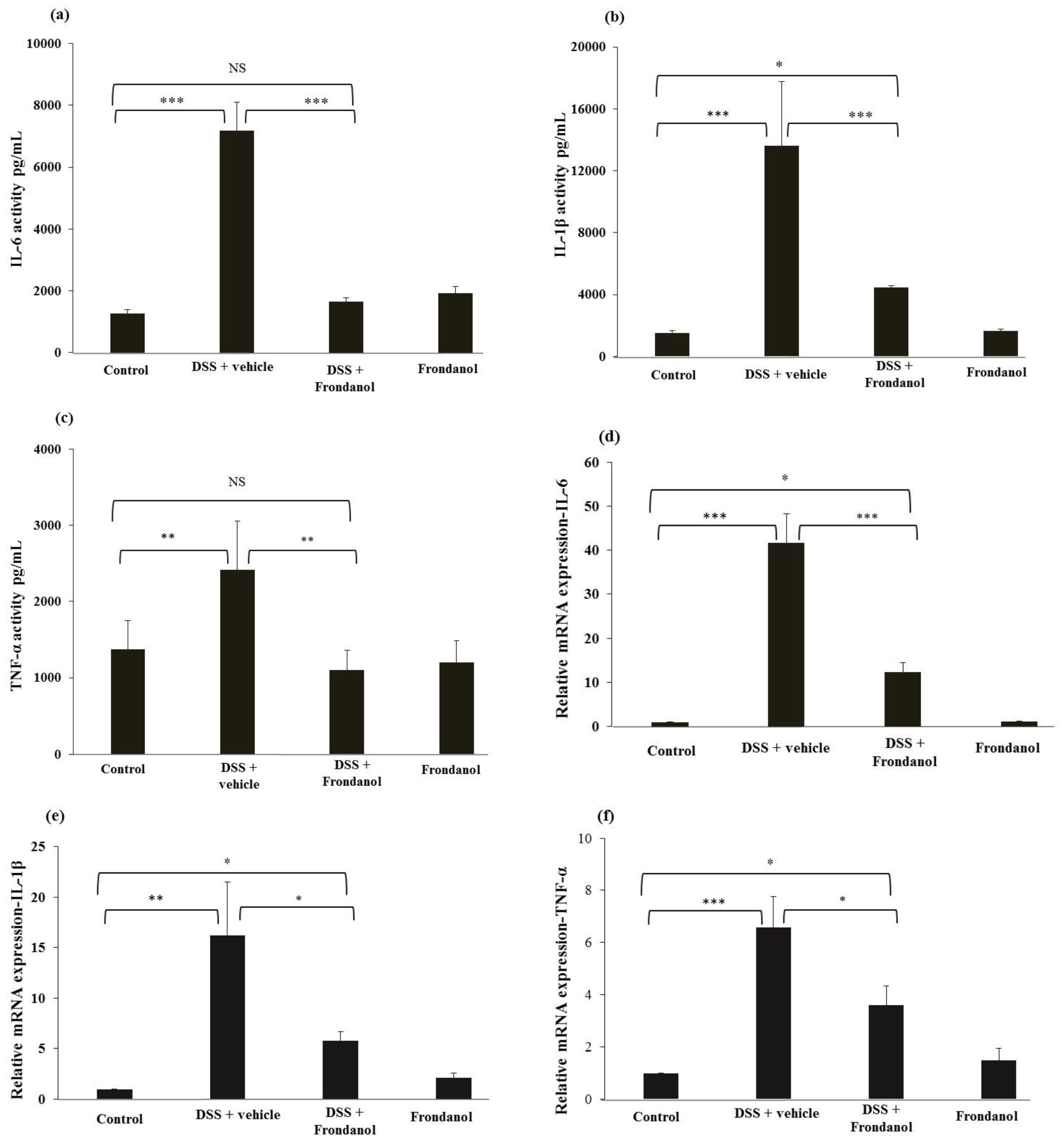
2.4. Effect of Frondanol on Pro-Inflammatory Cytokine Protein and mRNA Concentrations
2.5. Effect of Frondanol on Leukotriene B4 (LTB4)
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Experimental Animals
4.3. Experimental Design, Induction of Colitis and Tissue Collection
4.4. Macroscopic Assessment or Disease Activity Index (DAI)
4.5. ELISA
4.6. Colonic Cytokine mRNA Content Determined by Real-Time PCR
4.7. Histological Scoring
4.8. Statistics
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lee, S.H.; Kwon, J.E.; Cho, M.L. Immunological pathogenesis of inflammatory bowel disease. Intest. Res. 2018, 16, 26–42. [Google Scholar] [CrossRef] [PubMed]
- Ananthakrishnan, A.N. Epidemiology and risk factors for IBD. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Duijvestein, M.; Battat, R.; Vande Casteele, N.; D’Haens, G.R.; Sandborn, W.J.; Khanna, R.; Jairath, V.; Feagan, B.G. Novel therapies and treatment strategies for patients with inflammatory bowel disease. Curr. Treat. Options Gastroenterol. 2018, 16, 129–146. [Google Scholar] [CrossRef] [PubMed]
- Zezos, P.; Nguyen, G.C. Use of complementary and alternative medicine in inflammatory bowel disease around the world. Gastroenterol. Clin. N. Am. 2017, 46, 679–688. [Google Scholar] [CrossRef] [PubMed]
- Malve, H. Exploring the ocean for new drug developments: Marine pharmacology. J. Pharm. Bioallied Sci. 2016, 8, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Bordbar, S.; Anwar, F.; Saari, N. High-value components and bioactives from sea cucumbers for functional foods—A review. Mar. Drugs 2011, 9, 1761–1805. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, Y.; Zhang, W.; Franco, C. Discovery of Novel Saponins from the Viscera of the Sea Cucumber Holothuria lessoni. Mar. Drugs 2014, 12, 2633–2667. [Google Scholar] [CrossRef] [PubMed]
- Collin, P.D. Sea Cucumber Carotenoid Lipid Fraction Products and Methods of Use. U.S. Patent 6,399,105B1, 4 June 2002. [Google Scholar]
- Hamabata, T.; Nakamura, T.; Masuko, S.; Maeda, S.; Murata, T. Production of lipid mediators across different disease stages of dextran sulfate sodium-induced colitis in mice. J. Lipid Res. 2018, 59, 586–595. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.P.; Patil, C.S.; Kulkarni, S.K. Effect of 5-lipoxygenase inhibition on events associated with inflammatory bowel disease in rats. Indian J. Exp. Biol. 2004, 42, 667–673. [Google Scholar] [PubMed]
- Song, E.M.; Jung, S.A.; Lee, J.S.; Kim, S.E.; Shim, K.N.; Jung, H.K.; Yoo, K.; Park, H.Y. Benzoxazole derivative B-98 ameliorates dextran sulfate sodium-induced acute murine colitis and the change of T cell profiles in acute murine colitis model. Korean J. Gastroenterol. 2013, 62, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Kimura, I.; Nagahama, S.; Kawasaki, M.; Kataoka, M.; Sato, M. Study on the experimental ulcerative colitis (UC) model induced by dextran sulfate sodium (DSS) in rats (3). Nihon Yakurigaku Zasshi 1996, 108, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Kitamura, T.; Suzuki, M.; Fujii, T.; Shoji, H.; Tanaka, K.; Igarashi, J. Effects of alpha-linolenic acid on colonic secretion in rats with experimental colitis. J. Gastroenterol. 2007, 42, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Low, D.; Nguyen, D.D.; Mizoguchi, E. Animal models of ulcerative colitis and their application in drug research. Drug Des. Dev. Ther. 2013, 7, 1341–1357. [Google Scholar]
- Eichele, D.D.; Kharbanda, K.K. Dextran sodium sulfate colitis murine model: An indispensable tool for advancing our understanding of inflammatory bowel diseases pathogenesis. World J. Gastroenterol. 2017, 23, 6016–6029. [Google Scholar] [CrossRef] [PubMed]
- Chassaing, B.; Aitken, J.D.; Malleshappa, M.; Vijay-Kumar, M. Dextran sulfate sodium (DSS)-induced colitis in mice. Curr. Protoc. Immunol. 2014, 104, 15–25. [Google Scholar]
- Mizoguchi, A. Animal models of inflammatory bowel disease. Prog. Mol. Biol. Transl. Sci. 2012, 105, 263–320. [Google Scholar] [PubMed]
- Roginsky, A.B.; Ding, X.Z.; Woodward, C.; Ujiki, M.B.; Singh, B.; Bell, R.H., Jr.; Collin, P.; Adrian, T.E. Anti-pancreatic cancer effects of a polar extract from the edible sea cucumber, Cucumaria frondosa. Pancreas 2010, 39, 646–652. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, R.; Collin, P.D. Method of Inhibiting Rejection of Transplanted Material. World Patent Application No. WO200309267281, 13 November 2003. [Google Scholar]
- Collin, P.D.; Yang, P.; Newman, R. Methods and Compositions for Treating Lipoxygenase-Medaited Disease States. U.S. Patent 6541519B2, 1 April 2003. [Google Scholar]
- Cole, N.; Hume, E.B.; Jalbert, I.; Vijay, A.K.; Krishnan, R.; Willcox, M.D. Effects of topical administration of 12-methyl tetradecanoic acid (12-mta) on the development of corneal angiogenesis. Angiogenesis 2007, 10, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.-Z.; Collin, P.D.; Adrian, T.E. Myristoleic acid inhibits pancreatic cancer growth Via 5-lipoxygenase inhibition. Pancreas 2009, 38, 993. [Google Scholar]
- Yasui, Y.; Hosokawa, M.; Mikami, N.; Miyashita, K.; Tanaka, T. Dietary astaxanthin inhibits colitis and colitis-associated colon carcinogenesis in mice via modulation of the inflammatory cytokines. Chem. Biol. Interact. 2011, 193, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.J.; Shajib, M.S.; Manocha, M.M.; Khan, W.I. Investigating intestinal inflammation in DSS-induced model of IBD. J. Vis. Exp. 2012, 60, e3678. [Google Scholar] [CrossRef] [PubMed]
- Ohkusa, T.; Okayasu, I.; Tokoi, S.; Araki, A.; Ozaki, Y. Changes in bacterial phagocytosis of macrophages in experimental ulcerative colitis. Digestion 1995, 56, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Okayasu, I.; Hatakeyama, S.; Yamada, M.; Ohkusa, T.; Inagaki, Y.; Nakaya, R. A novel method in the induction of reliable experimental acute and chronic ulcerative colitis in mice. Gastroenterology 1990, 98, 694–702. [Google Scholar] [CrossRef]
- Ohkawara, T.; Mitsuyama, K.; Takeda, H.; Asaka, M.; Fujiyama, Y.; Nishihira, J. Lack of macrophage migration inhibitory factor suppresses innate immune response in murine dextran sulfate sodium-induced colitis. Scand. J. Gastroenterol. 2008, 43, 1497–1504. [Google Scholar] [CrossRef] [PubMed]
- Carr, M.W.; Roth, S.J.; Luther, E.; Rose, S.S.; Springer, T.A. Monocyte chemoattractant protein 1 acts as a T-lymphocyte chemoattractant. Proc. Natl. Acad. Sci. USA 1994, 91, 3652–3656. [Google Scholar] [CrossRef] [PubMed]
- Ohtsuka, Y.; Sanderson, I.R. Dextran sulfate sodium-induced inflammation is enhanced by intestinal epithelial cell chemokine expression in mice. Pediatr. Res. 2003, 53, 143–147. [Google Scholar] [PubMed]
- Sklyarov, A.Y.; Panasyuk, N.B.; Fomenko, I.S. Role of nitric oxide-synthase and cyclooxygenase/lipooxygenase systems in development of experimental ulcerative colitis. J. Physiol. Pharmacol. 2011, 62, 65–73. [Google Scholar] [PubMed]
- Zijlstra, F.J.; Garrelds, I.M.; van Dijk, A.P.; Wilson, J.H. Experimental colitis in mice: Effects of olsalazine on eicosanoid production in colonic tissue. Agents Actions 1992, 36, C76–C78. [Google Scholar] [CrossRef]
- Bertran, X.; Mane, J.; Fernandez-Banares, F.; Castella, E.; Bartoli, R.; Ojanguren, I.; Esteve, M.; Gassull, M.A. Intracolonic administration of zileuton, a selective 5-lipoxygenase inhibitor, accelerates healing in a rat model of chronic colitis. Gut 1996, 38, 899–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauritsen, K.; Laursen, L.S.; Bukhave, K.; Rask-Madsen, J. Effects of topical 5-aminosalicylic acid and prednisolone on prostaglandin e2 and leukotriene b4 levels determined by equilibrium in vivo dialysis of rectum in relapsing ulcerative colitis. Gastroenterology 1986, 91, 837–844. [Google Scholar] [CrossRef]
- Melstrom, L.G.; Bentrem, D.J.; Salabat, M.R.; Kennedy, T.J.; Ding, X.Z.; Strouch, M.; Rao, S.M.; Witt, R.C.; Ternent, C.A.; Talamonti, M.S.; et al. Overexpression of 5-lipoxygenase in colon polyps and cancer and the effect of 5-lox inhibitors in vitro and in a murine model. Clin. Cancer Res. 2008, 14, 6525–6530. [Google Scholar] [CrossRef] [PubMed]
- Hennig, R.; Grippo, P.; Ding, X.Z.; Rao, S.M.; Buchler, M.W.; Friess, H.; Talamonti, M.S.; Bell, R.H.; Adrian, T.E. 5-lipoxygenase, a marker for early pancreatic intraepithelial neoplastic lesions. Cancer Res. 2005, 65, 6011–6016. [Google Scholar] [CrossRef] [PubMed]
- Hennig, R.; Ding, X.Z.; Tong, W.G.; Witt, R.C.; Jovanovic, B.D.; Adrian, T.E. Effect of LY293111 in combination with gemcitabine in colonic cancer. Cancer Lett. 2004, 210, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Mao, F.; Xu, M.; Zuo, X.; Yu, J.; Xu, W.; Moussalli, M.J.; Elias, E.; Li, H.S.; Watowich, S.S.; Shureiqi, I. 15-lipoxygenase-1 suppression of colitis-associated colon cancer through inhibition of the IL-6/stat3 signaling pathway. FASEB J. 2015, 29, 2359–2370. [Google Scholar] [CrossRef] [PubMed]
- Mao, F.; Wang, M.; Wang, J.; Xu, W.R. The role of 15-LOX-1 in colitis and colitis-associated colorectal cancer. Inflamm. Res. 2015, 64, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Cooper, H.S.; Murthy, S.N.; Shah, R.S.; Sedergran, D.J. Clinicopathologic study of dextran sulfate sodium experimental murine colitis. Lab. Investig. 1993, 69, 238–249. [Google Scholar] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Pervin, M.; Hasnat, M.A.; Lim, J.H.; Lee, Y.M.; Kim, E.O.; Um, B.H.; Lim, B.O. Preventive and therapeutic effects of blueberry (vaccinium corymbosum) extract against dss-induced ulcerative colitis by regulation of antioxidant and inflammatory mediators. J. Nutr. Biochem. 2016, 28, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Urushima, H.; Nishimura, J.; Mizushima, T.; Hayashi, N.; Maeda, K.; Ito, T. Perilla frutescens extract ameliorates dss-induced colitis by suppressing proinflammatory cytokines and inducing anti-inflammatory cytokines. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 308, G32–G41. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Weight Loss | Score | Stool Consistency | Score | Rectal Bleeding | Score |
|---|---|---|---|---|---|
| No loss | 0 | Normal | 0 | No Blood | 0 |
| 1–5% | 1 | Loose stool | 2 | Heme occult + ve and visual pellet bleeding | 2 |
| 5–10% | 2 | Diarrhea | 4 | Gross bleeding and blood around anus | 4 |
| 10–20% | 3 | ||||
| >20% | 4 |
| Name | Forward | Reverse |
|---|---|---|
| IL-6 | 5′-ACAACCACGGCCTTCCCTACTT-3′ | 5′-CACGATTTCCCAGAGAACATGTG-3′ |
| IL-1β | 5′-ACCTGCTGGTGTGTGACGTT-3′ | 5′-TCGTTGCTTGGTTCTCCTTG-3′ |
| TNF-α | 5′-CACGTCCGTAGCAAACCACCAA-3′ | 5′-GTTGGTTGTCTTTGAGATCCAT-3′ |
| F4/80 | 5′-TGTGTCGTGCTGTTCAGAACC-3′ | 5′-AGGAATCCCGCAATGATGG-3′ |
| MIP-2 | 5′-GGATGGCTTTCATGGAAGGAG-3′ | 5′-TTGCTAAGCAAGGCACTGTGC-3′ |
| 18S | 5′-AAATCAGTTATGGTTCCTTTGGTC-3′ | 5′-GCTCTAGAATTACCACAGTTATCCAA-3′ |
| Crypt Grade | Quantification of the Percentage of Crypt Change | Crypt Distortion—Graded Based on the Extent of Involvement | |||
|---|---|---|---|---|---|
| Grade 0: | Intact crypt | 1 | 1–25% | 0 | No crypt distortion |
| Grade 1: | Shortening and loss of basal 1/3 of crypts | 2 | 26–50% | 1 | 1–25% |
| Grade 2: | Loss of basal 2/3 of crypts | 3 | 51–75% | 2 | 26–50% |
| Grade 3: | Loss of entire crypt with intact surface epithelium | 4 | 76–100% | 3 | 51–75% |
| Grade 4: | Loss of both entire crypt and surface epithelium (erosion) | 4 | 76–100% | ||
| Inflammation Graded | Percentage of Inflammation Involvement of Mucosal Surface Area | Hyperplastic Epithelium—Graded Based on the Extent of Involvement | ||
|---|---|---|---|---|
| 0 | 0 | No inflammation | 0 | None |
| 1 | 1 | 1–25% | 1 | 1–25% |
| 2 | 2 | 26–50% | 2 | 26–50% |
| 3 | 3 | 51–75% | 3 | 51–75% |
| 4 | 4 | 76–100% | 4 | 76–100% |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Subramanya, S.B.; Chandran, S.; Almarzooqi, S.; Raj, V.; Al Zahmi, A.S.; Al Katheeri, R.A.; Al Zadjali, S.A.; Collin, P.D.; Adrian, T.E. Frondanol, a Nutraceutical Extract from Cucumaria frondosa, Attenuates Colonic Inflammation in a DSS-Induced Colitis Model in Mice. Mar. Drugs 2018, 16, 148. https://doi.org/10.3390/md16050148
Subramanya SB, Chandran S, Almarzooqi S, Raj V, Al Zahmi AS, Al Katheeri RA, Al Zadjali SA, Collin PD, Adrian TE. Frondanol, a Nutraceutical Extract from Cucumaria frondosa, Attenuates Colonic Inflammation in a DSS-Induced Colitis Model in Mice. Marine Drugs. 2018; 16(5):148. https://doi.org/10.3390/md16050148
Chicago/Turabian StyleSubramanya, Sandeep B., Sanjana Chandran, Saeeda Almarzooqi, Vishnu Raj, Aisha Salem Al Zahmi, Radeya Ahmed Al Katheeri, Samira Ali Al Zadjali, Peter D. Collin, and Thomas E. Adrian. 2018. "Frondanol, a Nutraceutical Extract from Cucumaria frondosa, Attenuates Colonic Inflammation in a DSS-Induced Colitis Model in Mice" Marine Drugs 16, no. 5: 148. https://doi.org/10.3390/md16050148
APA StyleSubramanya, S. B., Chandran, S., Almarzooqi, S., Raj, V., Al Zahmi, A. S., Al Katheeri, R. A., Al Zadjali, S. A., Collin, P. D., & Adrian, T. E. (2018). Frondanol, a Nutraceutical Extract from Cucumaria frondosa, Attenuates Colonic Inflammation in a DSS-Induced Colitis Model in Mice. Marine Drugs, 16(5), 148. https://doi.org/10.3390/md16050148