The Effect of Fucoidan from the Brown Alga Fucus evanescence on the Activity of α-N-Acetylgalactosaminidase of Human Colon Carcinoma Cells


Abstract
:1. Introduction
2. Results and Discussion
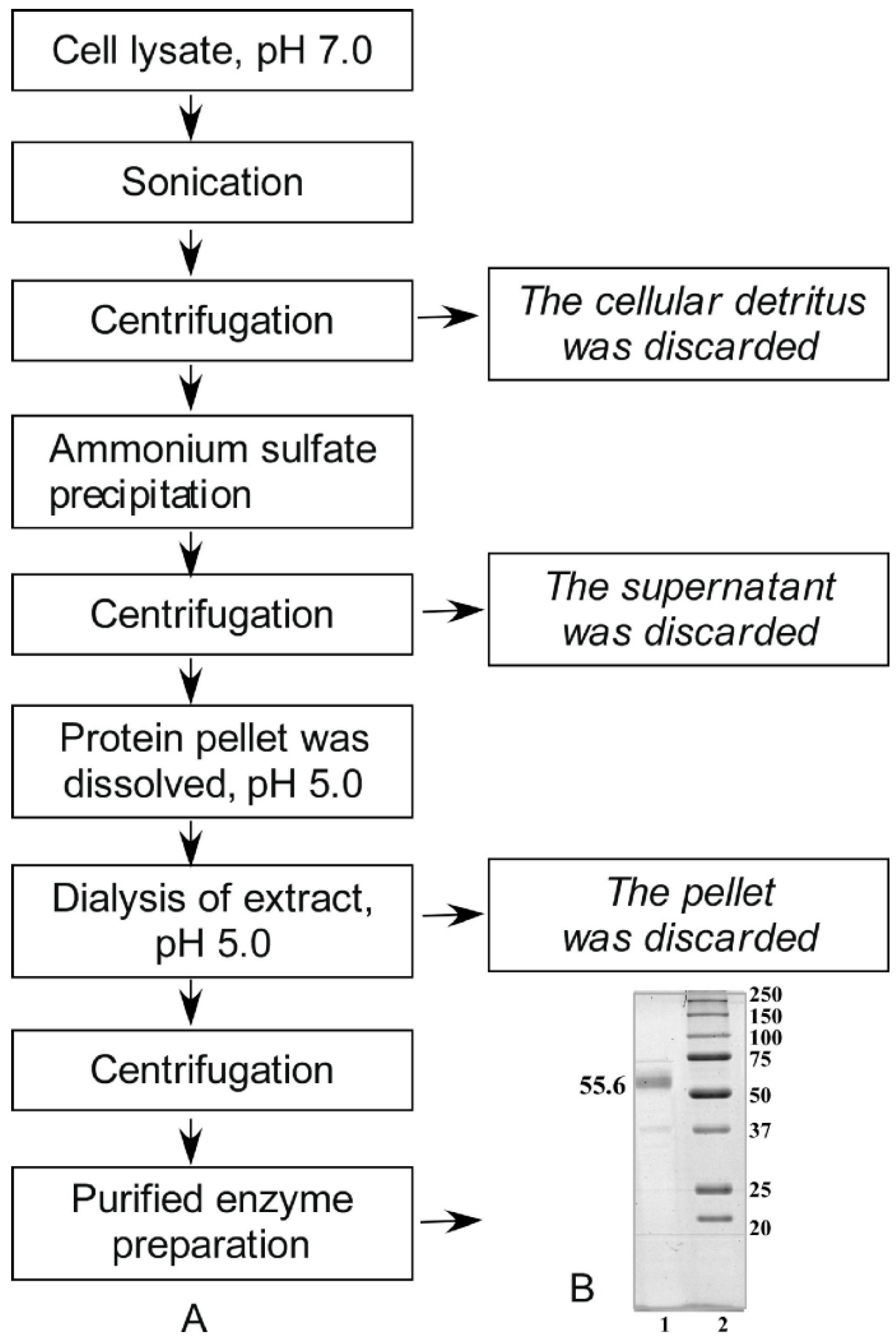
2.1. Isolation and Purification of Alpha-NaGalase from Cell Lysates
2.2. Biochemical and Enzyme Properties of Alpha-NaGalase from Cell Lysates
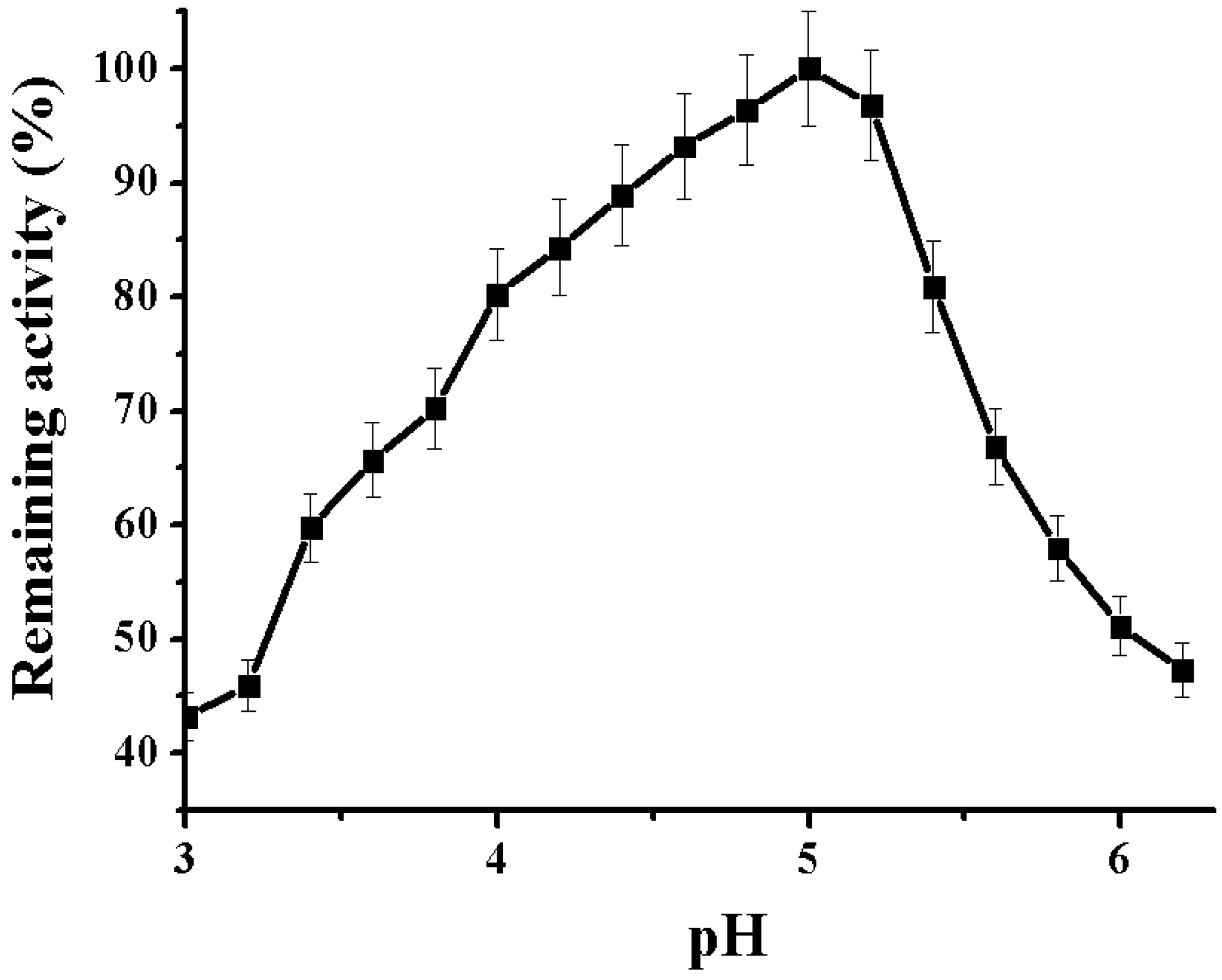
2.2.1. Determination of pH Optimum of the Alpha-NaGalase
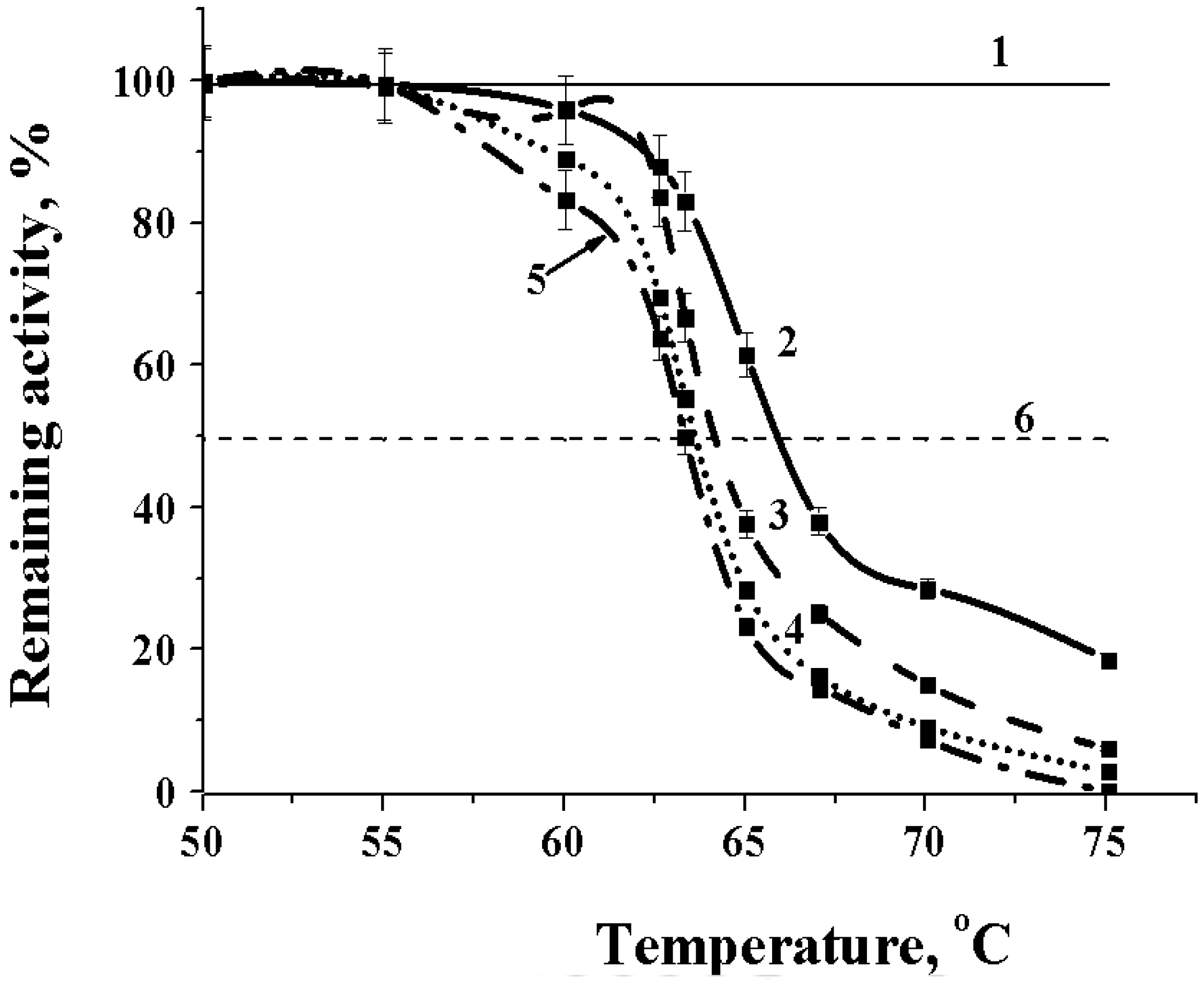
2.2.2. Effect of Temperature on the Alpha-NaGalase
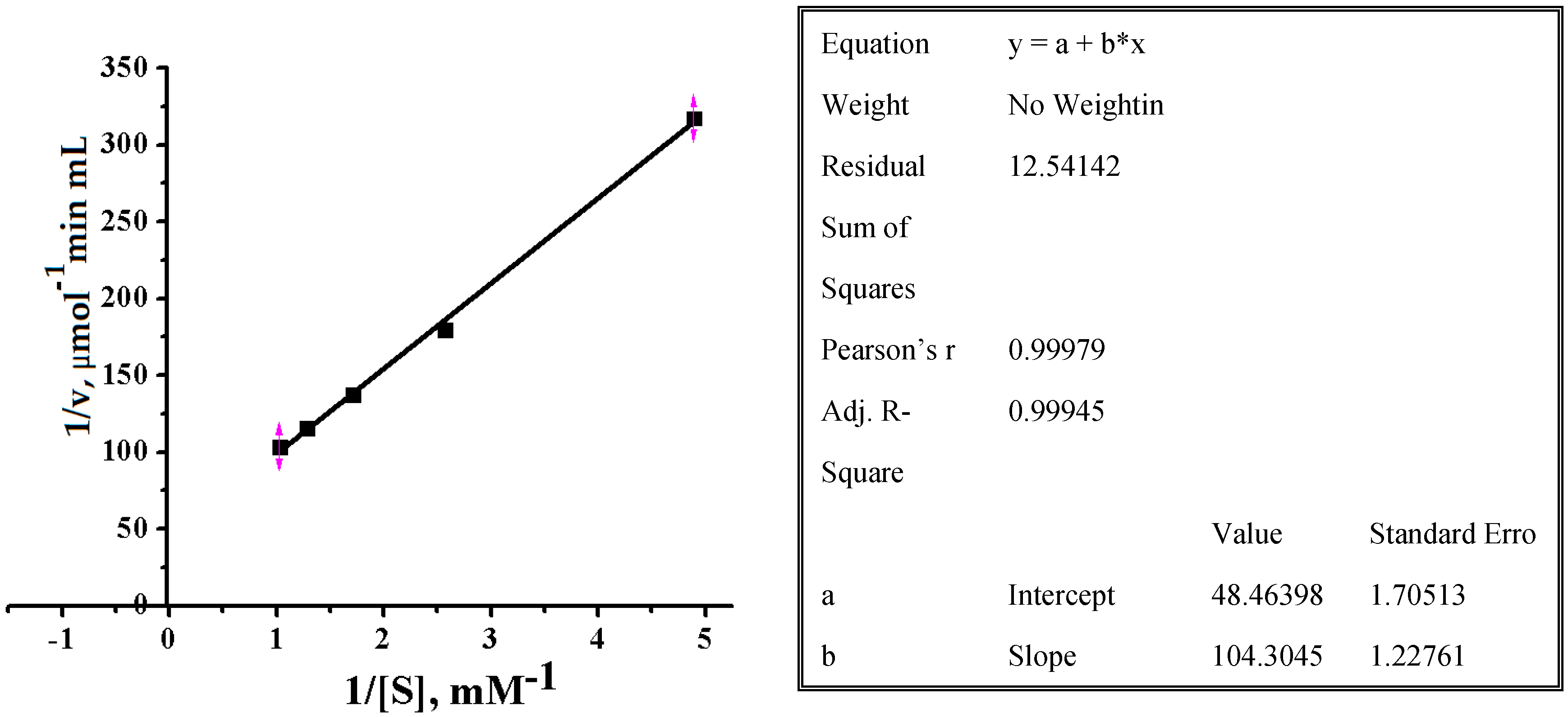
2.2.3. Effect of Substrate Concentrations on Reaction Rate of Alpha-NaGalase of DLD-1 Human Colon Carcinoma Cells
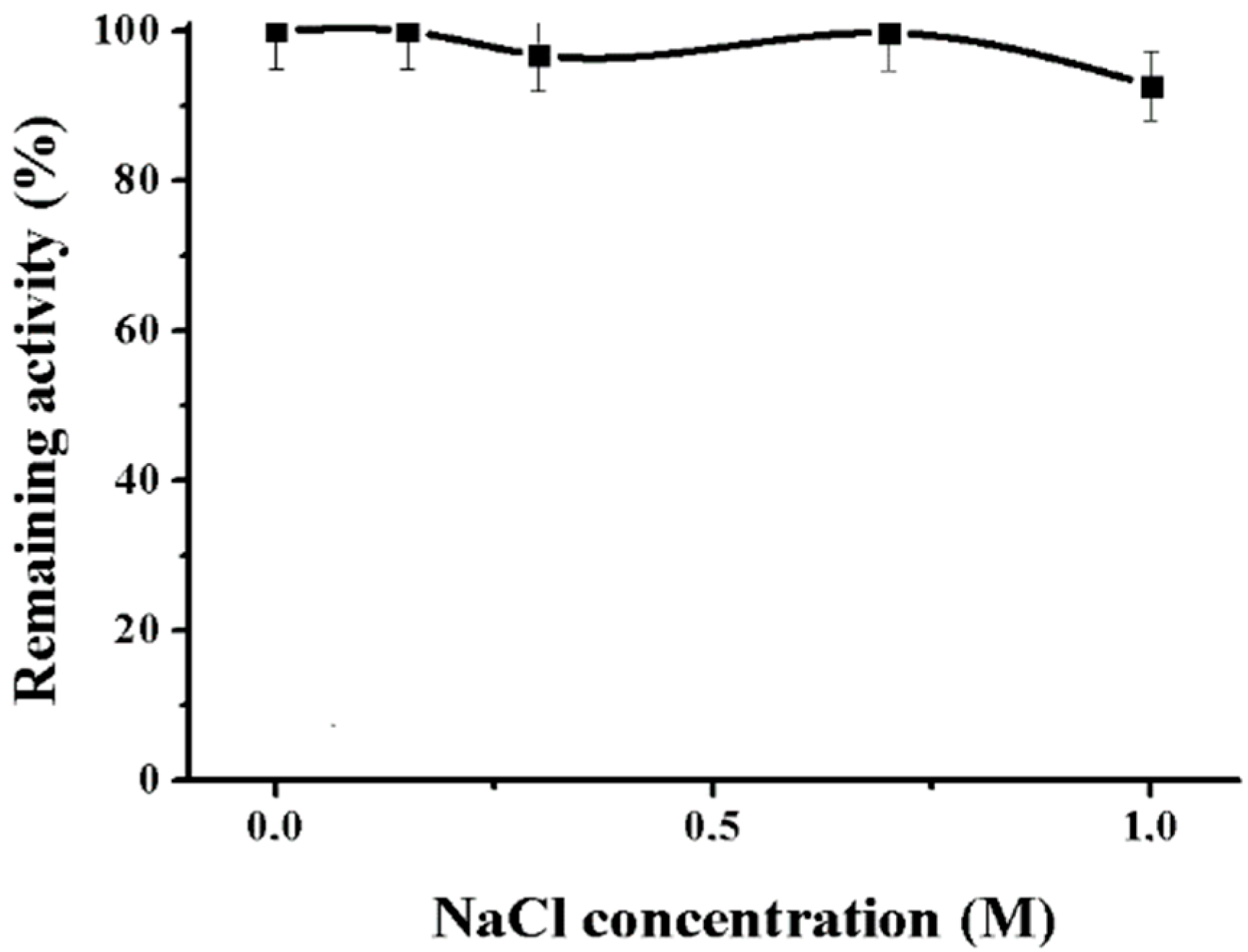
2.2.4. Effect of NaCl and NaN3 on the Alpha-NaGalase
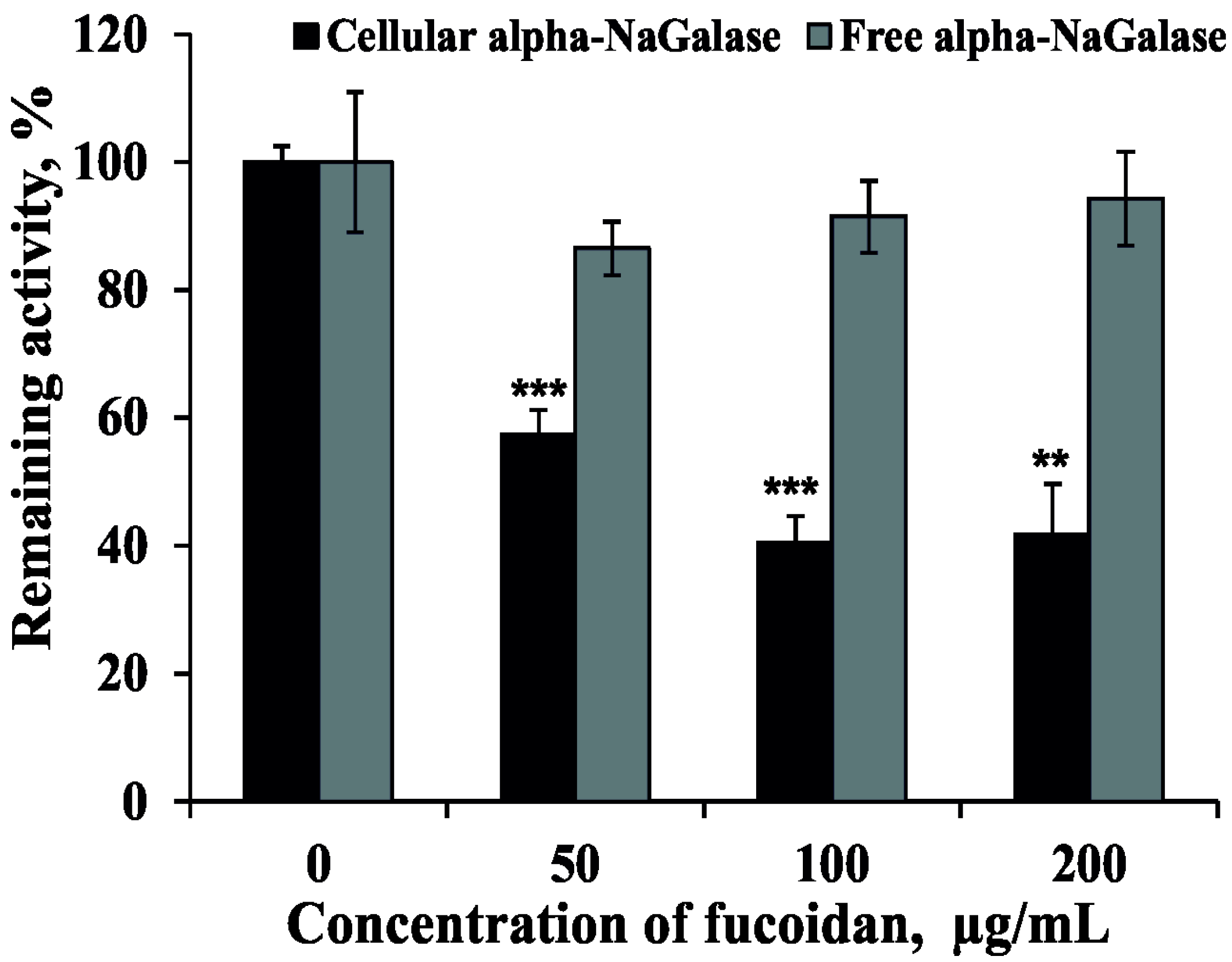
2.3. Effect of Fucoidan on the Activity of Alpha-NaGalase
3. Materials and Methods
3.1. Materials and Reagents
3.2. Brown Alga and Fucoidan Isolation
3.3. Cell Cultured and Treated with Fucoidan
3.3.1. Preparation of Cell Lysate
3.3.2. Fucoidan’s Treatment of Cells
3.4. Isolation and Purification of α-N-Acetylgalactosaminidase from Cell Lysates
3.5. Biochemical and Enzyme Properties of α-N-Acetylgalactosaminidase from Cell Lysates
3.5.1. The Molecular Weight
3.5.2. Enzyme Assay
3.5.3. pH Optimum of the Alpha-NaGalase
3.5.4. Effect of Temperature on the Alpha-NaGalase
3.5.5. Effect of NaCl and NaN3 on the Alpha-NaGalase
3.5.6. Determination of Km and Vmax Values
3.6. Effect of Fucoidan on Alpha-NaGalase
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Santos-Pereira, J.M.; Muñoz-Galván, S. Biomarkers of colorectal cancer: A genome-wide perspective. Cancer Transl. Med. 2016, 2, 182–188. [Google Scholar]
- Clausen, H.; Hakomori, S.-I. ABH and related histo-blood group antigens; immunochemical differences in carrier isotypes and their distribution. Vox Sang. 1989, 56, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.M.; Wu, J.H.; Chen, Y.Y.; Tsai, M.S.; Herp, A. Forssman pentasaccharide and polyvalent Galβ1→4GlcNAc as major ligands with affinity for Caragana arborescens agglutinin. FEBS Lett. 1999, 463, 223–230. [Google Scholar] [CrossRef]
- Nakajima, H.; Kurosaka, A.; Fujisawa, A.; Kawasaki, T.; Matsuyana, M.; Nagayo, T.; Yamashina, I. Isolation and characterization of a glycoprotein from a human rectal adenocarcinoma. J. Biochem. 1983, 93, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.M. Carbohydrate structural units in glycosphingolipids as receptors for Gal and GalNAc reactive lectins. Neurochem. Res. 2002, 27, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Kenne, L.; Lindberg, B. Bacterial Polysaccharides. In The Polysaccharides; Aspinoll, G.O., Ed.; Academic Press: New York, NY, USA, 1983; Volume 2, pp. 287–363. [Google Scholar]
- Tomshich, S.V.; Isakov, V.V.; Komandrova, N.A.; Shevchenko, L.S. Structure of the O-specific polysaccharide of the marine bacterium Arenibacter palladensis KMM 3961T containing 2-acetamido-2-deoxy-l-galacturonic acid. Biochemistry 2012, 77, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Callahan, J.W.; Lassila, E.L.; Den Tandt, W.; Philippart, M. α-N-acetylgalactosaminidase: Isolation, properties and distribution of the human enzyme. Biochem. Med. 1973, 7, 424–431. [Google Scholar] [CrossRef]
- Weissman, B.; Hindrichsen, D.F. Mamalian α-N-acetylgalactosaminidase. Occurrence, partial purification and action on linkage in submaxillary mucins. Biochemistry 1969, 8, 2034–2043. [Google Scholar] [CrossRef]
- Hata, J.; Dhar, M.; Mitra, M.; Harmata, M.; Haibach, F.; Sun, P.; Smith, D. Purification and characterization of N-acetyl-alpha-d-galactosaminidase from Gallus domesticus. Biochem. Int. 1992, 28, 77–86. [Google Scholar] [PubMed]
- Tuppy, H.; Staudenbauer, W.L. The action on soluble blood group A substances of an alpha-N-acetylgalactosaminidase from Helix pomatia. Biochemistry 1966, 5, 1742–1747. [Google Scholar] [CrossRef] [PubMed]
- Uda, Y.; Li, S.C.; Li, Y.T. α-N-Acetylgalactosaminidase from the limpet Patella vulgate. J. Biol. Chem. 1977, 252, 5194–5200. [Google Scholar] [PubMed]
- Schauer, H.; Cottschalk, A. Studies on glycoproteins: XVII. Purification of O-seryl-N-acetylgalactosaminidine glycohydrolase. Its complete separation from N-acetyl-β-d-hexosaminidase and proteases. Biochim. Biophis. Acta 1968, 156, 304–310. [Google Scholar] [CrossRef]
- Ashida, H.; Tamaki, H.; Fujimoto, T.; Yamamoto, K.; Kumagai, H. Molecular cloning of cDNA encoding α-N-acetylgalactosaminidase from Acremonium sp., and its expression in yeast. Arch. Biochem. Biophys. 2000, 384, 305–310. [Google Scholar] [CrossRef] [PubMed]
- McDonald, M.J.; Bahl, O.P. [93a] α-N-acetylgalactosaminidase from Aspergillus niger. Methods Enzymol. 1972, 28, 734–738. [Google Scholar]
- Levy, G.N.; Aminoff, D. Purification and properties of α-N-acetylgalactosaminidase from Clostridium perfringens. J. Biol. Chem. 1980, 255, 11737–11742. [Google Scholar] [PubMed]
- Hoskins, L.C.; Boulding, E.T.; Larson, G. Purification and characterization of blood group A-degrading isoform of α-N-acetylgalactosaminidase from Ruminococcus torques strain IX-70. J. Biol. Chem. 1997, 272, 7932–7939. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.P.; Sulzenbacher, G.; Yuan, H.; Bennett, E.P.; Pietz, G.; Saunders, K.; Spence, J.; Nudelman, E.; Levery, S.B.; White, T.; et al. Bacterial glycosidases for the production of universal red blood cells. Nat. Biotechnol. 2007, 25, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Sadik, G.; Rashid, M.H.; Nashida, T.I.; Ishii, K.; Satoh, Y.; Shiraishi, T.; Uda, Y. Chemical and Immunological Characterization of the two α-N-acetylgalactosaminidases from Squid (Todarodes pacificus) Liver. Biol. Pharm. Bull. 2009, 32, 1469–1472. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.H.; Matsuzawa, T.; Satoh, Y.; Shiraishi, T.; Ando, M.; Sadik, G.; Uda, Y. Purification and characterization of α-N-Acetylgalactosaminidases I and II from Starfish, Asterina amurensis. Biosci. Biotech. Biochem. 2010, 74, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Asakawa, M.; Enomoto, N. Purification and characterization of α-N-acetylgalactosaminidase from Skipjack liver. J. Biochem. 1987, 101, 855–862. [Google Scholar] [CrossRef] [PubMed]
- Bakunina, I.Y.; Nedashkovskaya, O.I.; Kim, S.B.; Zvyagintseva, T.N.; Mikhailov, V.V. Diversity of glycosidase activities in the bacteria of the phylum Bacteroidetes isolated from marine algae. Microbiology 2012, 81, 688–695. [Google Scholar] [CrossRef]
- Bakunina, I.Y.; Nedashkovskaya, O.I.; Kim, S.B.; Zvyagintseva, T.N.; Mikhailov, V.V. Distribution of α-N-acetylgalactosaminidases among marine bacteria of the phylum Bacteroidetes, epiphytes of marine algae of the Seas of Okhotsk and Japan. Microbiology 2012, 81, 375–380. [Google Scholar] [CrossRef]
- Bakunina, I.Y.; Nedashkovskaya, O.I.; Balabanova, L.A.; Zvyagintseva, T.N.; Rasskasov, V.A.; Mikhailov, V.V. Comparative analysis of glycoside hydrolases activities from phylogenetically diverse marine bacteria of the genus Arenibacter. Mar. Drugs 2013, 11, 1977–1998. [Google Scholar] [CrossRef] [PubMed]
- Van Diggelen, O.P.; Schindler, D.; Kleijer, W.J.; Huijmans, J.M.G.; Galjaard, H.; Linden, H.-U.; Peter-Katalinic, J.; Egge, H.; Cantz, M. Lysosomal alpha-NaGalase deficiency: A new inherited metabolic disease. Lancet 1987, 2, 804. [Google Scholar] [CrossRef]
- Yamamoto, N.; Urade, M. Pathogenic significance of α-N-acetylgalactosaminidase activity found in the hemagglutinin of influenza virus. Microbes Infect. 2005, 7, 674–681. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, N. Pathogenic significance of α-N-acetylgalactosaminidase activity found in the envelope glycoprotein gp160 of human immunodeficiency virus Type I. AIDS Res. Hum. Retroviruses 2006, 22, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Lombard, V.; Ramulu, H.G.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, N.; Naraparaju, V.R. Vitamin D3-binding protein as a precursor for macrophage activating factor in the inflammation-primed macrophage activation cascade in rats. Cell Immunol. 1996, 170, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, N.; Naraparaju, V.R.; Moore, M.; Brent, L.H. Deglycosylation of serum vitamin D3-binding protein by alpha-N-acetylgalactosaminidase detected in the plasma of patients with systemic lupus erythematosus. Clin. Immunol. Immunopathol. 1997, 82, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Reddi, A.L.; Sankaranarayanan, K.; Arulraj, H.S.; Devaraj, N.; Devaraj, H. Serum α-N-acetylgalactosaminidase is associated with diagnosis/prognosis of patients with squamous cell carcinoma of the uterine cervix. Cancer Lett. 2000, 158, 61–64. [Google Scholar] [CrossRef]
- Yamamoto, N.; Naraparaju, V.R.; Asbell, S.O. Deglycosylation of serum vitamin D3-binding protein leads to immunosuppression in cancer patients. Cancer Res. 1996, 56, 2827–2831. [Google Scholar] [PubMed]
- Yamamoto, N.; Naraparaju, V.R.; Urade, M. Prognostic utility of serum alpha-N-acetylgalactosaminidase and immunosuppression resulted from deglycosylation of serum Gc protein in oral cancer patients. Cancer Res. 1997, 57, 295–299. [Google Scholar] [PubMed]
- Mohamad, S.B.; Nagasawa, H.; Uto, Y.; Hori, H. Tumor cell alpha-N-acetylgalactosaminidase activity and its involvement in GcMAF-related macrophage activation. Comp. Biochem. Physiol. Part A 2002, 132, 1–8. [Google Scholar] [CrossRef]
- Matsuura, T.; Uematsu, T.; Yamaoka, M.; Furusawa, K. Effect of salivary gland adenocarcinoma cell-derived α-N-acetylgalactosaminidase on the bioactivity of macrophage activating factor. Int. J. Oncol. 2004, 24, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Ale, M.T.; Maruyama, H.; Tamauchi, H.; Mikkelsen, J.D.; Meyer, A.S. Fucoidan from Sargassum sp. and Fucus vesiculosus reduces cell viability of lung carcinoma and melanoma cells in vitro and activates natural killer cells in mice in vivo. Int. J. Biol. Macromol. 2011, 49, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Azuma, K.; Ishihara, T.; Nakamoto, H.; Amaha, T.; Osaki, T.; Tsuka, T.; Imagawa, T.; Minami, S.; Takashima, O.; Ifuku, S.; et al. Effects of oral administration of fucoidan extracted from Cladosiphon okamuranus on tumor growth and survival time in a tumor-bearing mouse model. Mar. Drugs 2012, 10, 2337–2348. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, J.; Wada-Funada, U.; Mano, H.; Matahira, Y.; Kawaguchi, M.; Wada, M. Proportion of murine cytotoxic T-cells is increased by high molecular-weight fucoidan extracted from okinawa mozuku (Cladosiphon okamuranus). J. Health Sci. 2005, 51, 394–397. [Google Scholar] [CrossRef]
- Hu, Y.; Cheng, S.C.; Chan, K.T.; Ke, Y.; Xue, B.; Sin, F.W.; Zeng, C.; Xie, Y. Fucoidin enhances dendritic cell-mediated T-cell cytotoxicity against ny-eso-1 expressing human cancer cells. Biochem. Biophys. Res. Commun. 2010, 392, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.O.; Zhang, W.; Du, J.Y.; Wong, K.W.; Oda, T.; Yu, Q. Fucoidan can function as an adjuvant in vivo to enhance dendritic cell maturation and function and promote antigen-specific T-cell immune responses. PLoS ONE 2014, 9, e99396. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-T.; Rioux, L.-E.; Turgeon, S.L. Alpha-amylase and alpha-glucosidase inhibition is differentially modulated by fucoidan obtained from Fucus vesiculosus and Ascophyllum nodosum. Phytochemistry 2014, 98, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-T.; Rioux, L.-E.; Turgeon, S.L. Molecular weight and sulfate content modulate the inhibition of α-amylase by fucoidan relevant for type 2 diabetes management. PharmaNutrition 2015, 3, 108–114. [Google Scholar] [CrossRef]
- Kumar, T.V.; Lakshmanasenthil, S.; Geetharamani, D.; Marudhupandic, T.; Sujaa, G.; Suganyaa, P. Fucoidan–A α-d-glucosidase inhibitor from Sargassum wightii with relevance to type 2 diabetes mellitus therapy. Int. J. Biol. Macromol. 2015, 72, 1044–1047. [Google Scholar] [CrossRef] [PubMed]
- Morozova, O.V.; Shumakovich, G.P.; Gorbacheva, M.A.; Shleev, S.V.; Yaropolov, A.I. Review: ‘‘Blue’’ laccases. Biochemistry 2007, 72, 1136–1150. [Google Scholar] [PubMed]
- Bakunina, I.Yu.; Kuhlmann, R.A.; Likhosherstov, L.M.; Martynova, M.D.; Nedashkovskaya, O.I.; Mikhailov, V.V.; Elyakova, L.A. α-N-Acetylgalactosaminidase from marine bacterium Arenibacter latericius KMM 426T removing blood type specificity of A erythrocytes. Biochemistry 2002, 67, 689–695. [Google Scholar] [PubMed]
- Bakunina, I.Yu.; Sova, V.V.; Nedashkovskaya, O.I.; Kuhlmann, R.A.; Likhosherstov, L.M.; Martynova, M.D.; Mikhailov, V.V.; Elyakova, L.A. α-Galactosidase of marine bacterium Pseudoalteromonas sp. KMM 701. Biochemistry 1998, 63, 1420–1427. [Google Scholar]
- Comfort, D.A.; Bobrov, K.S.; Ivanen, D.R.; Shabalin, K.A.; Harris, J.M.; Kulminskaya, A.A.; Brumer, H.; Kelly, R.M. Biochemical analysis of Thermotoga maritima GH36 α-d-galactosidase (TmGalA) confirms the mechanistic commonality of clan GH-D glycoside hydrolases. Biochemistry. 2007, 46, 3319–3330. [Google Scholar] [CrossRef] [PubMed]
- Viladot, J.L.; de Ramon, E.; Durany, O.; Planas, A. Probing the mechanism of Bacillus 1,3–1,4-β-d-glucan 4-glucanohydrolases by chemical rescue of inactive mutants at catalytically essential residues. Biochemistry 1998, 37, 11332–11342. [Google Scholar] [CrossRef] [PubMed]
- Sirimontree, P.; Fukamizo, T.; Suginta, W. Azide anions inhibit GH-18 endochitinase and GH-20 exo-β-N-acetylglucosaminidase from the marine bacterium Vibrio harveyi. J. Biochem. 2016, 159, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Vishchuk, O.S.; Ermakova, S.P.; Zvyagintseva, T.N. The fucoidans from brown algae of Far-Eastern seas: Anti-tumor activity and structure–function relationship. Food Chem. 2013, 141, 1211–1217. [Google Scholar] [CrossRef] [PubMed]
- Anastyuk, S.D.; Shevchenko, N.M.; Ermakova, S.P.; Vishchuk, O.S.; Nazarenko, E.L.; Dmitrenok, P.S.; Zvyagintseva, T.N. Anticancer activity in vitro of a fucoidan from the brown alga Fucus evanescens and its low molecular fragments, structurally characterized by tandem mass-spectrometry. Carbohydr. Polym. 2012, 87, 186–194. [Google Scholar] [CrossRef]
- Schepetkin, A.B.; Quinn, M.T. Botanical polysaccharides: Macrophage immunomodulation and therapeutic potential. Int. Immunopharmacol. 2006, 6, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.L.; Wang, J.Z.; Noda, H.; Amano, H.; Ogawa, S. The effects of an algal polysaccharide from Gloiopeltis tenax on transplantable tumors and immune activities in mice. Planta Med. 1995, 61, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Mishima, T.; Murata, J.; Toyoshima, M.; Fujii, H.; Nakajima, M.; Hayashi, T.; Kato, T.; Saiki, I. Inhibition of tumor invasion and metastasis by calcium spirulan (Ca-SP), a novel sulfated polysaccharide derived from a blue-green alga, Spirulina platensis. Clin. Exp. Metastasis 1998, 16, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Okai, Y.; Higashi-Okai, K.; Ishizaka, K.S.; Yamashita, U. Enhancing effect of polysaccharides from an edible brown alga, Hijikia fusiforme (Hijiki), on release of tumor necrosis factor-α from macrophages of endotoxin-nonresponder C3H/HeJ Mice. Nutr. Cancer 1997, 27, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Liang, H.; Tang, Q.; Xue, Ch.; He, X.; Zhang, L.; Zhang, Z.; Liang, Z.; Bian, K.; Zhang, L.; Li, Z. The protective and immunomodulatory effects of fucoidan against 7,12-dimethyl benz[a]anthracene-induced experimental mammary carcinogenesis through the PD1/PDL1 signaling pathway in rats. Nutr. Cancer 2017, 69, 1234–1244. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, V.K. Cleavage of structural proteins during of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecular Mass, kDa | 55.6 | |
| Optimum pH range | 4.6–5.0 | |
| Temperature stability | Optimum temperature range | 50 °C |
| Start of inactivation | 62 °C | |
| 50% inactivation | 63–67 °C | |
| 100% inactivation | 75 °C | |
| Catalytic properties | Km, mM | 2.15 |
| Vmax, μmol min−1 mL−1 | 0.021 | |
| kcat, min−1 | 1.55 | |
| kcat/Km, min−1 mM−1 | 0.72 | |
| NaN3, % | Remaining Activity, % |
|---|---|
| 0 | 100 |
| 0.1 | 85 |
| 0.3 | 67 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakunina, I.; Chadova, O.; Malyarenko, O.; Ermakova, S. The Effect of Fucoidan from the Brown Alga Fucus evanescence on the Activity of α-N-Acetylgalactosaminidase of Human Colon Carcinoma Cells. Mar. Drugs 2018, 16, 155. https://doi.org/10.3390/md16050155
Bakunina I, Chadova O, Malyarenko O, Ermakova S. The Effect of Fucoidan from the Brown Alga Fucus evanescence on the Activity of α-N-Acetylgalactosaminidase of Human Colon Carcinoma Cells. Marine Drugs. 2018; 16(5):155. https://doi.org/10.3390/md16050155
Chicago/Turabian StyleBakunina, Irina, Oksana Chadova, Olesya Malyarenko, and Svetlana Ermakova. 2018. "The Effect of Fucoidan from the Brown Alga Fucus evanescence on the Activity of α-N-Acetylgalactosaminidase of Human Colon Carcinoma Cells" Marine Drugs 16, no. 5: 155. https://doi.org/10.3390/md16050155