High-Level Expression of a Thermally Stable Alginate Lyase Using Pichia pastoris, Characterization and Application in Producing Brown Alginate Oligosaccharide


Abstract
:1. Introduction
2. Results and Discussion
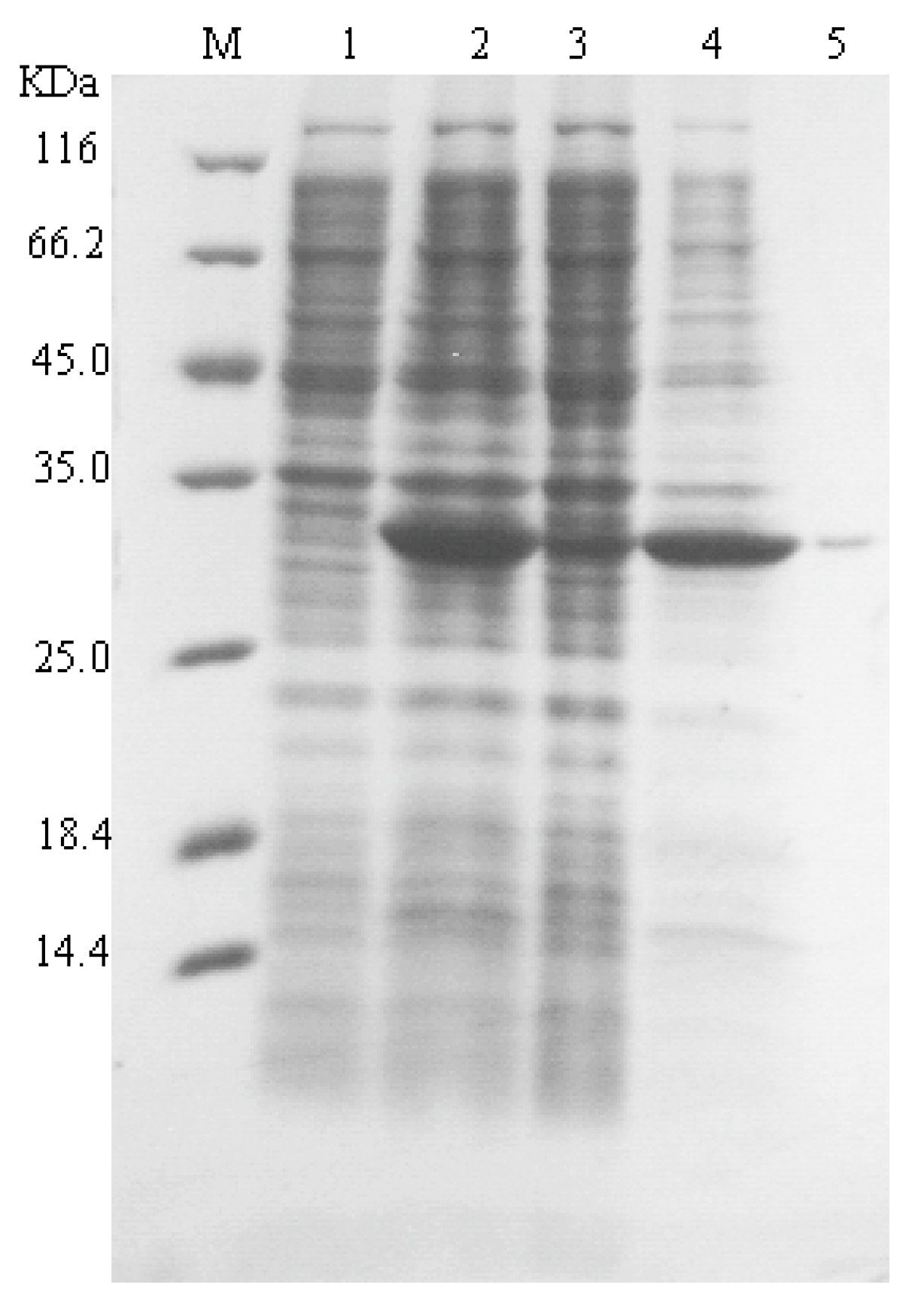
2.1. Recombinant Expression of SAGL in Escherichia coli
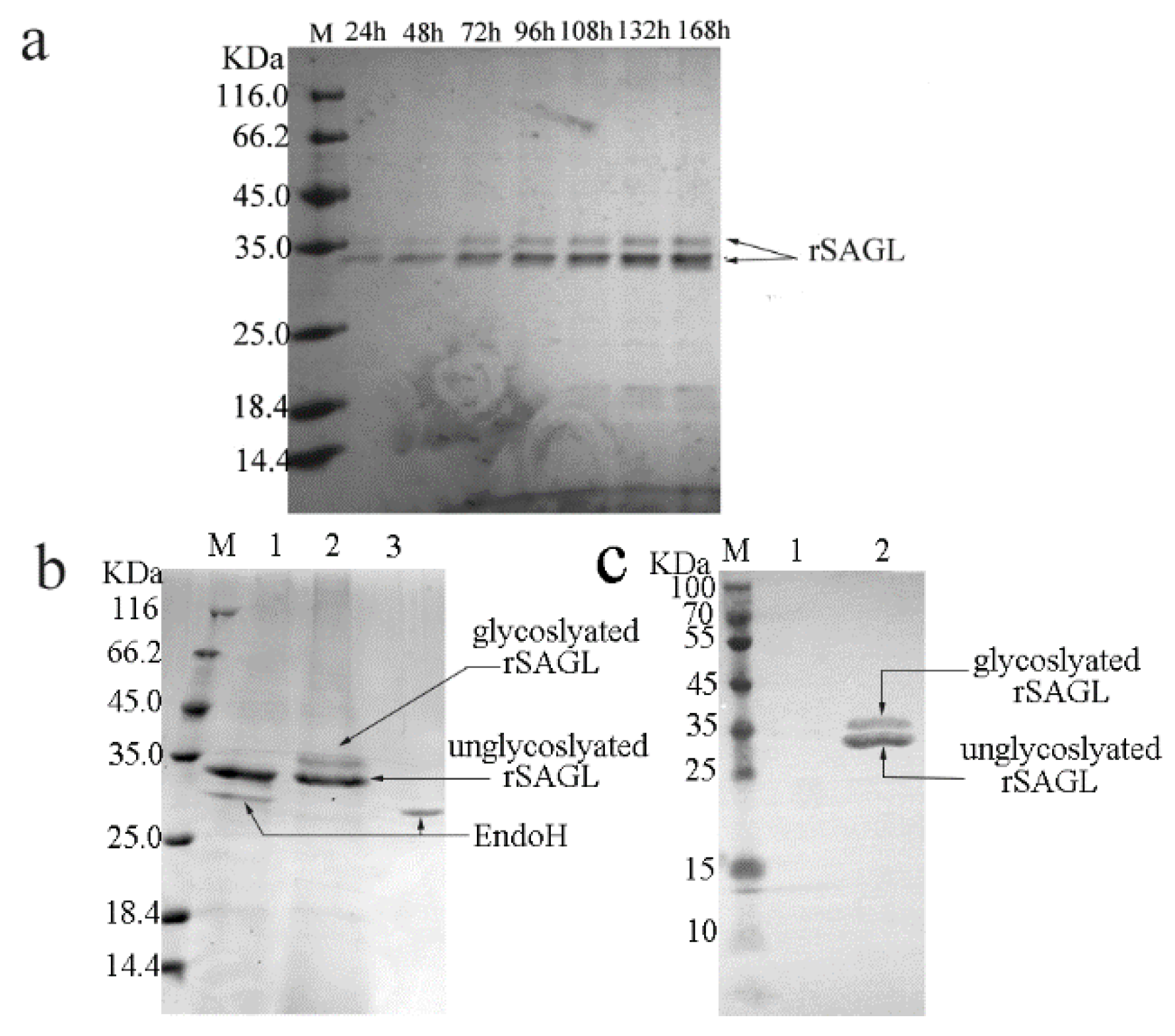
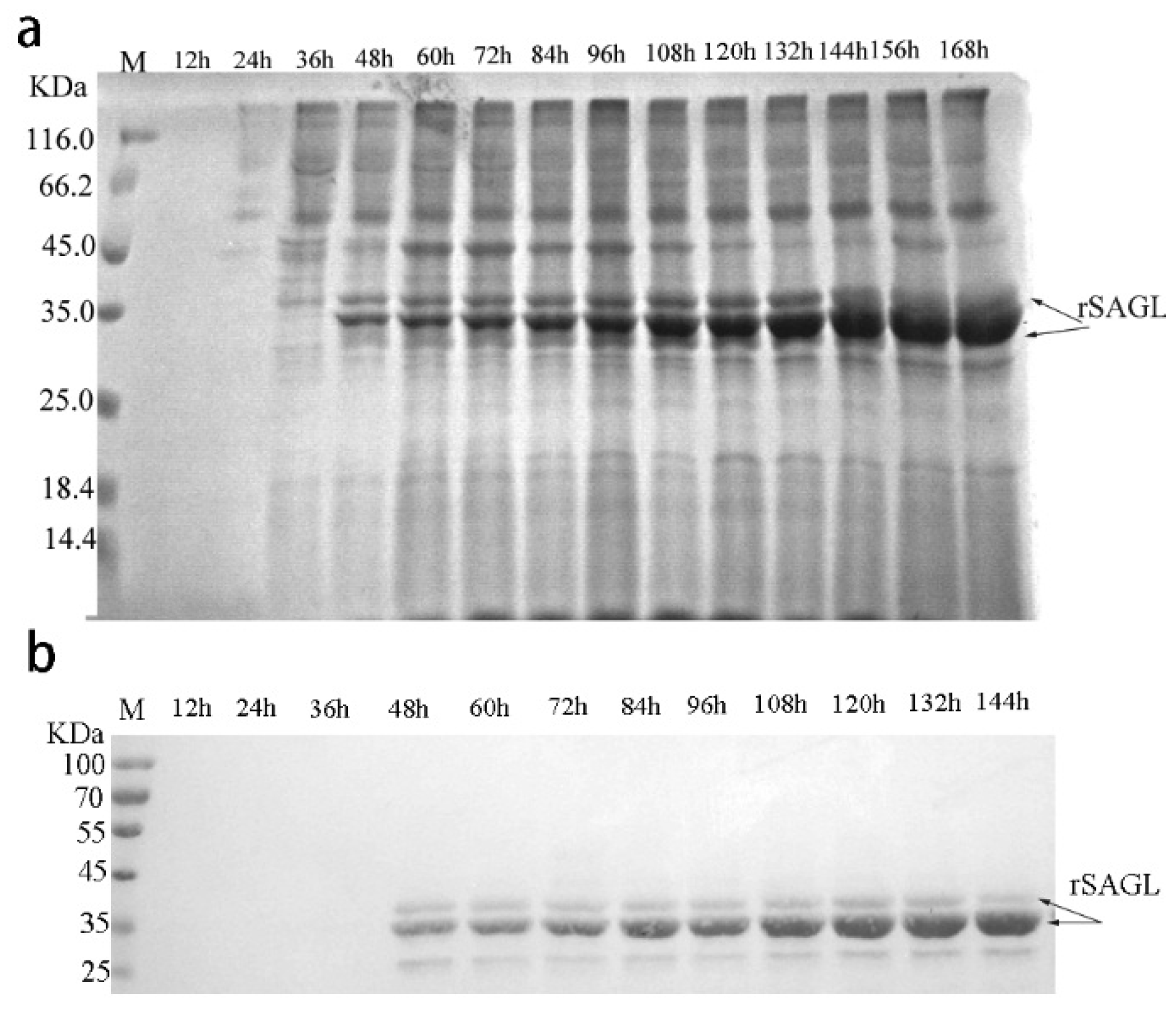
2.2. Recombinant Expression of SAGL Using P. pastoris in Flask
2.3. Characterization of Recombinant sagl from E. coli and P. pastoris Systems
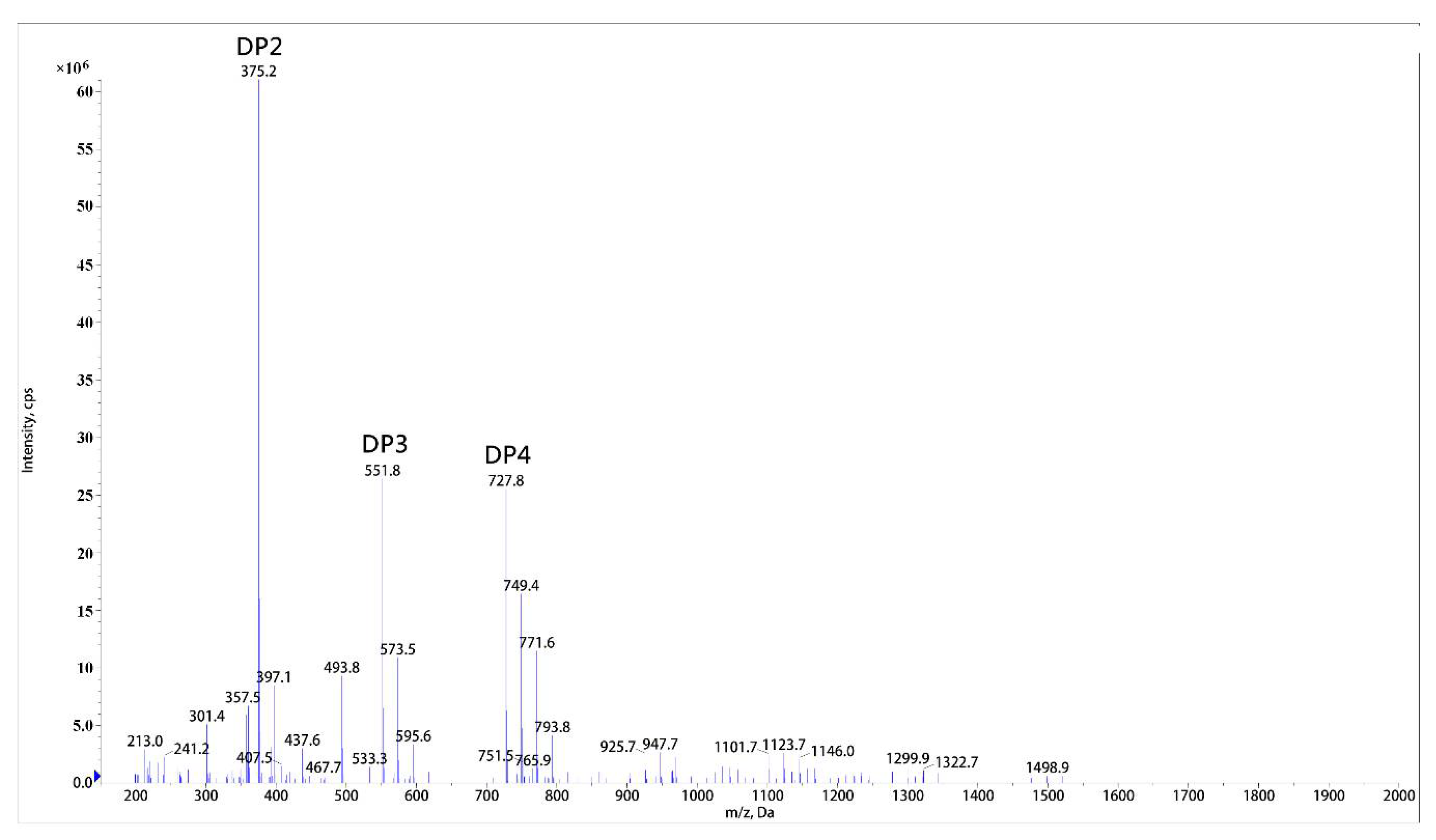
2.4. End Product Analysis
2.5. Characterization of Thermal Stability of rSAGL
2.6. Comparison of Catalytic Property
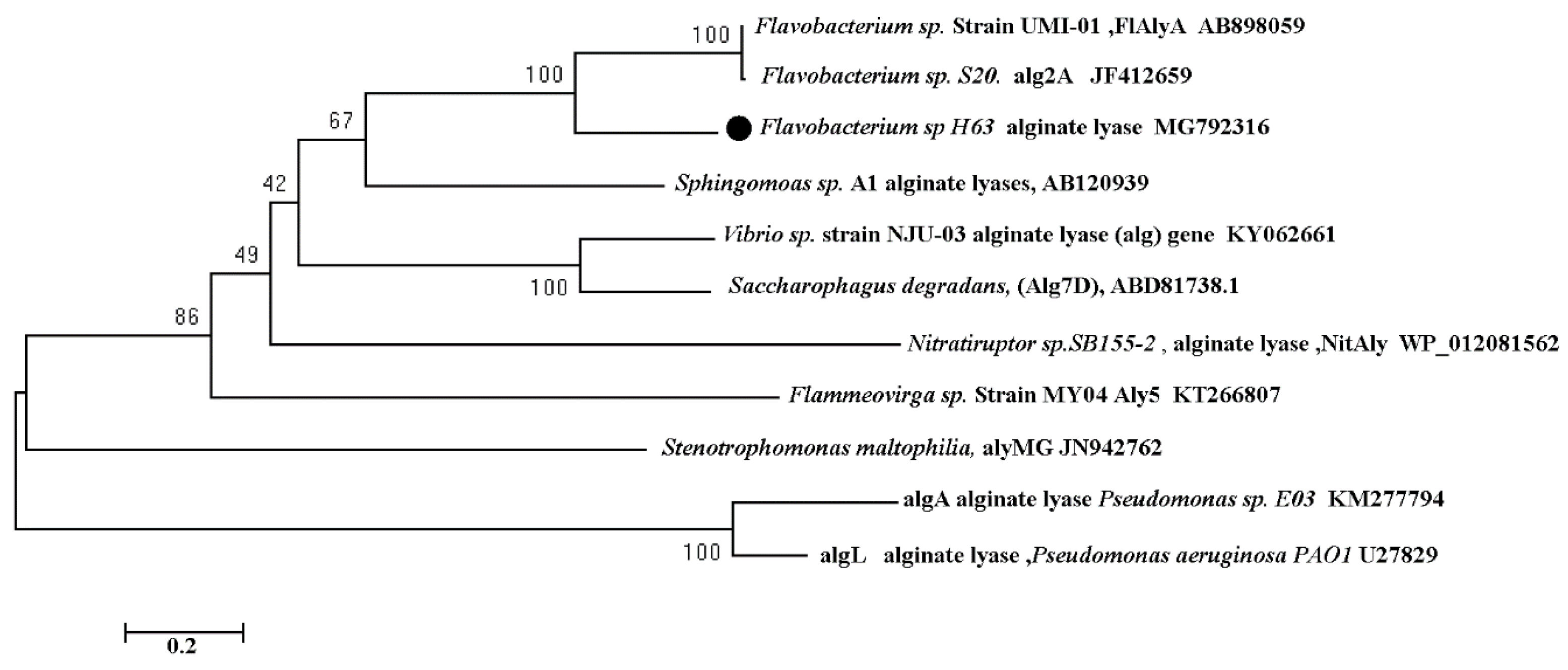
2.7. Phylogenetic Tree Analysis of Sagl Based on Amino Acid Sequence
2.8. High Density Fermentation
2.9. Production and Analysis of Brown Alginate Oligosaccharide
3. Materials and Methods
3.1. Expression of Alginate Lyase in E. coli System
3.2. Strain and Plasmid of Pichia System
3.3. Gene Cloning and Construction of Recombinant Yeast Expression Vector In Vitro
3.4. Transformation and Colony Screening of P. pastoris
3.5. Shaking Flask Fermentation Test
3.6. Recombinant Enzyme Purification, Western Blotting and Activity Assay
3.7. Substrate Specificity Determination
3.8. Characterization of Optimal Reaction Temperature and pH
3.9. Characterization of Thermal Stability
3.10. Characterization of Metal Ions Effects
3.11. ESI-MS and TLC Analysis of End Oligosaccharide Product
3.12. High Cell-Density Fermentation of Recombinant Alginate Lyase in P. pastoris
3.13. Enzymatic Production of Alginate Oligosaccharide
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Zhu, Y.; Wu, L.; Chen, Y.; Ni, H.; Xiao, A.; Cai, H. Characterization of an extracellular biofunctional alginate lyase from marine Microbulbifer sp. ALW1 and antioxidant activity of enzymatic hydrolysates. Microbiol. Res. 2016, 182, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Inoue, A.; Takadono, K.; Nishiyama, R.; Tajima, K.; Kobayashi, T.; Ojima, T. Characterization of an Alginate Lyase, FlAlyA, from Flavobacterium sp. Strain UMI-01 and Its Expression in Escherichia coli. Mar. Drugs 2014, 12, 4693–4712. [Google Scholar] [CrossRef] [PubMed]
- Kang, L.X.; Chen, X.M.; Fu, L.; Ma, L.X. Recombinant expression of chitosanase from Bacillus subtilis HD145 in Pichia pastoris. Carbohyd. Res. 2012, 352, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Yue, M.M.; Gong, W.W.; Qiao, Y.; Ding, H. A Method for Efficient Expression of Pseudomonas aeruginosa Alginate Lyase in Pichia pastoris. Prep. Biochem. Biotechnol. 2016, 46, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Gu, J.; Cheng, Y.; Liu, H.; Li, Y.; Li, F. Novel Alginate Lyase (Aly5) from a Polysaccharide-Degrading Marine Bacterium, Flammeovirga sp. Strain MY04: Effects of Module Truncation on Biochemical. Characteristics, Alginate Degradation Patterns, and Oligosaccharide-Yielding Properties. Appl. Environ. Microb. 2015, 82, 364. [Google Scholar] [CrossRef] [PubMed]
- Inoue, A.; Anraku, M.; Nakagawa, S.; Ojima, T. Discovery of a Novel Alginate Lyase from Nitratiruptor sp. SB155-2 Thriving at Deep-sea Hydrothermal Vents and Identification of the Residues Responsible for Its Heat Stability. J. Biol. Chem. 2016, 291, 15551–15563. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Zhu, Y.M.; Men, Y.; Zeng, Y.; Sun, Y.X. Purification and Characterization of a Novel Alginate Lyase from the Marine Bacterium Bacillus sp. Alg07. Mar. Drugs 2018, 16, 86. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.W.; Sun, Y.; Ni, F.; Ning, L.M.; Yao, Z. Characterization of a new endo-type alginate lyase from Vibrio sp. NJU-03. Int. J. Biol. Macromol. 2018, 108, 1140–1147. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.L.; Sheng, D.; Fei, X.; Fang, D.; Li, P.Y.; Zhang, X.Y.; Zhou, B.C.; Zhang, Y.Z.; Xie, B.B. Characterization of a New Cold-Adapted and Salt-Activated Polysaccharide Lyase Family 7 Alginate Lyase from Pseudoalteromonas sp. SM0524. Front. Microbiol. 2016, 7, 1120. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, L.; Hao, J.; Xing, M.; Sun, J.; Sun, M. Purification and Characterization of a New Alginate Lyase from Marine Bacterium Vibrio sp. SY08. Mar. Drugs 2017, 15, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Guo, E.W.; Yu, W.G.; Han, F. Purification and characterization of a new alginate lyase from a marine bacterium Vibrio sp. Biotechnol. Lett. 2013, 35, 703–708. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.W.; Huang, L.S.; Tan, H.D.; Qin, Y.Q.; Du, Y.G.; Yin, H. Characterization of a new endo-type polyM-specific alginate lyase from Pseudomonas sp. Biotechnol. Lett. 2015, 37, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Hye-Jin, Y.; Hashimoto, W.; Osamu Miyake, M.O.; Mikami, B.; Murata, K. Overexpression in Escherichia coli, purification, and characterization of Sphingomonas sp. A1 alginate lyases. Protein Expr. Purif. 2000, 19, 84–90. [Google Scholar] [CrossRef]
- Kim, H.T.; Ko, H.J.; Kim, N.; Kim, D.; Lee, D.; Choi, I.G.; Woo, H.C.; Kim, M.D.; Kim, K.H. Characterization of a recombinant endo-type alginate lyase (Alg7D) from Saccharophagus degradans. Biotechnol. Lett. 2012, 34, 1087–1092. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zhou, J.; Li, X.; Peng, Q.; Lu, H.; Du, Y. Characterization of a new alginate lyase from newly isolated Flavobacterium sp. S20. J. Ind. Microbiol. Biotechnol. 2013, 40, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.I.; Choi, S.H.; Lee, E.Y.; Kim, H.S. Molecular cloning, purification, and characterization of a novel polyMG-specific alginate lyase responsible for alginate MG block degradation in Stenotrophomas maltophilia KJ-2. Appl. Microbiol. Biotechnol. 2012, 95, 1643–1653. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metal Ion | Ion Concentration | Relative Activity |
|---|---|---|
| Mn2+ | 1 mM | 0 |
| 10 mM | 0 | |
| Cu2+ | 1 mM | 0 |
| 10 mM | 0 | |
| Co2+ | 1 mM | 0 |
| 10 mM | 0 | |
| Ca2+ | 1 mM | 74 ± 0.2 |
| 10 mM | 0 | |
| Mg2+ | 1 mM | 106 ± 0.6 |
| 10 mM | 93 ± 1.3 | |
| 100 mM | 147 ± 0.9 | |
| Ag+ | 1 mM | 0 |
| 10 mM | 0 | |
| Zn2+ | 1 mM | 15 ± 0.4 |
| 10 mM | 0 | |
| Ni+ | 1 mM | 29 ± 0.7 |
| 10 mM | 0 | |
| Fe3+ | 1 mM | 5.3 ± 0.2 |
| 10 mM | 14 ± 1.1 | |
| Control enzyme | 0 | 100 ± 3.4 |
| Ions Concentration (mM) | Relative Activity (%) | |
|---|---|---|
| NaCl | KCl | |
| 0 | 100 | 100 |
| 10 | 201 ± 2.4 | 237 ± 5.6 |
| 20 | 292 ± 4.1 | 274 ± 3.5 |
| 50 | 168 ± 5.4 | 270 ± 1.7 |
| 80 | 135 ± 8.3 | 344.4 ± 2.9 |
| 100 | 246 ± 1.2 | 325 ± 2.7 |
| 150 | 120 ± 5.0 | 300 ± 6.2 |
| 200 | 131.5 ± 6.1 | 283.7 ± 7.5 |
| 300 | 140 ± 3.3 | 215 ± 1.8 |
| 400 | 215 ± 4.6 | 147 ± 0.4 |
| 500 | 197 ± 5.3 | 177 ± 8.0 |
| Origin | Source | Substrate Preference | Optimum Catalytic Temperature and Thermal Stability | Specific Activity | Main End Product | Yield | Reference |
|---|---|---|---|---|---|---|---|
| Microbulbifer sp. ALW1 | E. coli | Poly G and alginate | 45 °C and retained 68% activity at 45 °C for 1 h | 1.49 U/mg | DP 2,3 | N.D. | [1] |
| Flavobacterium sp. UMI-01 | E. coli | Poly M | 55 °C and inactivated at 50 °C for 30 min | 23478 EU/mg | DP 2,3,4 | 168 EU/mL | [2] |
| Pseudomonas aeruginosa PAOI | P. pastoris | Poly (MG) | 40 °C and inactivated at 50 °C for 30 min | 2440 U/mg. | DP 2,3 | 21 U/mL | [4] |
| Flammeovirga sp. MY04: | E. coli | Poly G | 40 °C and retained 80% of activity at 40 °C for 2 h | 620 U/mg | DP 2,3,4 | 496 U/mL (800 mg/L) | [5] |
| Nitratiruptor sp. SB155-2 | E. coli | Poly M | 70 °C and retain 20% of activity by incubation at 50 °C for 16 h | 1620 U/mg. | DP 3,4,5,6 | 1.944 U/mL (1.2 mg/L) | [6] |
| Bacillus sp. Alg07 | native | Poly M | 40 °C and retained 50% activity at 50 °C for 0.75 h | 8306.7 U/mg | DP 2,3,4 | N.D. | [7] |
| Vibrio sp. NJU-03 | E. coli | Poly G | 30 °C and retained 40% activity at 40 °C for 30 min | 6468.99 U/mg | DP 2,3,4 | N.D. | [8] |
| Pseudoalteromonas sp. SM0524 | E. coli | Poly M | 30 °C and lost 80% activity at 40 °C for 15 min | 62.6 U/mg | DP 2,3,4 | N.D. | [9] |
| Vibrio sp. SY08 | native | Poly G and Poly M | 40 °C and retained 75% activity at 40 °C for 2 h | 1070.2 U/mg | DP 2 | 2.247 U/mL (2.1 mg/L) | [10] |
| Vibrio sp. QY105 | native | Poly G and alginate | 38 °C and retained 58% of activity at 50 °C for 20 min | 2152 U/mg | DP 2,3,4,5 | 15.8 U/mL | [11] |
| Pseudomonas sp. E03 | E. coli | Poly M | 30 °C and lost 50% activity at 50 °C for 30 min | 222 EU/mg | DP 2,3,4,5 | N.D. | [12] |
| Sphingomonas sp. A1-II | E. coli | Poly G | 70 °C and lost 50% of activity at 50 °C for 10 min | 109 U/mg | DP 3,4 | 3040 U/L | [13] |
| Saccharophagus degradans 2–40 | E. coli | Poly G and Poly M | 50 °C and lost 58% of activity at 50 °C for 30 min | 4.6 U/mg | DP 2,3,4,5 | 41.4 U/L | [14] |
| Flavobacterium S20 | E. coli | Poly G | 45 °C and lost 20% of activity at 45 °C for 60 min | 365.38 U/mg | DP 5,6,7 | 19.6 U/mL | [15] |
| Stenotrophomas maltophilia KJ-2 | E. coli | Poly (MG) | 40 °C and inactivitaed at higher than 40 °C for 30 min | 848.3 U/mg | DP 2,3,4 | N.D. | [16] |
| Flavobacterium sp. H63 | P. pastoris | Poly M and alginate | 45 °C and retained 49.0% activity at 50 °C for 72 h | 4044 U/mg | DP 2,3,4 | 915.5 U/mL (226.4 μg/mL) | This study |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Wang, S.; Zhang, Y.; Chen, L. High-Level Expression of a Thermally Stable Alginate Lyase Using Pichia pastoris, Characterization and Application in Producing Brown Alginate Oligosaccharide. Mar. Drugs 2018, 16, 158. https://doi.org/10.3390/md16050158
Li H, Wang S, Zhang Y, Chen L. High-Level Expression of a Thermally Stable Alginate Lyase Using Pichia pastoris, Characterization and Application in Producing Brown Alginate Oligosaccharide. Marine Drugs. 2018; 16(5):158. https://doi.org/10.3390/md16050158
Chicago/Turabian StyleLi, Haifeng, Shuling Wang, Yunyi Zhang, and Liehuan Chen. 2018. "High-Level Expression of a Thermally Stable Alginate Lyase Using Pichia pastoris, Characterization and Application in Producing Brown Alginate Oligosaccharide" Marine Drugs 16, no. 5: 158. https://doi.org/10.3390/md16050158